NL9201716A - Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. - Google Patents
Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. Download PDFInfo
- Publication number
- NL9201716A NL9201716A NL9201716A NL9201716A NL9201716A NL 9201716 A NL9201716 A NL 9201716A NL 9201716 A NL9201716 A NL 9201716A NL 9201716 A NL9201716 A NL 9201716A NL 9201716 A NL9201716 A NL 9201716A
- Authority
- NL
- Netherlands
- Prior art keywords
- omp
- outer membrane
- saccharide
- polypeptide
- membrane vesicle
- Prior art date
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/195—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria
- C07K14/22—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from bacteria from Neisseriaceae (F)
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P31/00—Antiinfectives, i.e. antibiotics, antiseptics, chemotherapeutics
- A61P31/04—Antibacterial agents
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
Landscapes
- Chemical & Material Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Medicinal Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Molecular Biology (AREA)
- Genetics & Genomics (AREA)
- Biophysics (AREA)
- Biochemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- Veterinary Medicine (AREA)
- Oncology (AREA)
- Animal Behavior & Ethology (AREA)
- Pharmacology & Pharmacy (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Public Health (AREA)
- Communicable Diseases (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Medicinal Preparation (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Polysaccharides And Polysaccharide Derivatives (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
Description
Buitenmembraanvesikel dat voorzien is van een groep pnlvoeptiden welkp ten minste de inunuunwerking van aan membraan gebonden buitenmembraanp-i -witten (OMP's) hebben, werkwi.ize ter bereiding ervan alsmede een varn’n dat een dereeli.lk buitenmembraanvesikel omvat.
De onderhavige uitvinding heeft betrekking op een buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben. De uitvinding heeft eveneens betrekking op een vaccin dat een dergelijk buitenmembraanvesikel omvat en op een werkwijze ter bereiding van een dergelijk buitenmembraanvesikel.
Het is bekend dat vaccins van gezuiverde kapselpolysacchariden (CPS) beschermende immuniteit kunnen induceren. Deze immuniteit is afhankelijk van de leeftijd van de ingeënte persoon en is slechts van korte duur. De intrinsieke nadelen van kapselpolysacchariden als vaccins worden omzeild door de klassieke benadering kapselpolysacchariden of daarvan afgeleide oligosacchariden te koppelen met eiwitten (Goebel, W.F., en O.T. Avery 1929 J. Exp. Med. 50:533-550 en Cruse, J.M. en R.E. Lewis (Erd.) 1989 Contrib. Microbiol. Immunol. Basel, Krager, 10:1-196). Het koppelen van polysacchariden aan eiwitten heeft tot gevolg dat het karakter van dit type antigeen verandert van thymus onafhankelijk in thymus afhankelijk. Dergelijke poly- of oligosaccharide-eiwitconjugaten zijn in het algemeen bij jonge kinderen zeer immunogeen en kunnen geheugen induceren.
Hier volgen een aantal voorbeelden van bekende saccharide-peptide-conjugaten:
Uit de Nederlandse octrooiaanvrage 86.023.25 zijn conjugaten van kapselpolysacchariden (CPS) van H. influenza k met tetanustoxoïde (TT) bekend.
Er zijn conjugaten bereid van kapselpolysacchariden van meningococcus groep A en C met tetanustoxoïde, welke conjugaten bij muizen en konijnen zeer immunogeen blijken te zijn ([Beuvery, E.C., A. Kaaden. V. Kanhai, en A.B. Leussink. 1983· Physicochemical and immunological characterization of meningococcal group A and C polysaccharide-tetanus toxoid conjugates prepared by two methods. Vaccine 1:31-36], [Beuvery, E.C., F Miedema, R van Delft en K. Haverkamp. 1983* Preparation and immunochemical characterization of meningococcal group C polysaccharide-tetanus toxoid conjugates as a new generation of vaccines. Infect. Immun. 40:39-^5] en [Jennings, H.J. en H.C. Lugowski. 1981. Immunochemistry of groups A, B and C meningococcal polysaccharide-tetanus toxoid conjugates. J. Immunol. 127:1011-1018]).
Een bacteriegroep waarvan kapselpolysacchariden geen of weinig immuunreactie opwekken is groep B meningococcus, een bacteriegroep die in vele landen meer dan 50% van de meningococcus ziektegevallen veroorzaakt (Poolman et al., The Lancet, september 1986, blz. 555*558). Voor de groep B meningococcus is derhalve gezocht naar andere saccharide-peptide-conjugaten die bruikbaar kunnen zijn in een vaccin, aangezien kapselpolysacchariden van deze groep geen immuunreactie geven tegen gramnegatieve bacteriën bij proefdieren en humane vrijwilligers. Hiertoe zijn sacchari-de-peptide-conjugaten gemaakt met gemodificeerd kapselpolysaccharide en dragereiwitten ([Jennings, H.J., R. Roy, en A. Gamian. 1986. Induction of meningococcus group B polysaccharide-specific IgG antibodies in mice using an N-proprionylated B polysaccharide-tetanus toxoid conjugate vaccine. J. Immunol. 137:1708-1713] en [Jennings. H.J. 1989= The capsular polysaccharide of group B Neisseria meningitidis as a vehicle for vaccine development. In Cruse, J. M., and R. E. Lewis, Conjugate Vaccines. Con-trib. Microbiol. Immunol. Basel, Krager. 10:151*165]).
Omdat men vermoedt dat anti groep B antilichamen (in het bijzonder IgG) in vivo kruisreactie vertonen met gastheerantigenen en het gebruik van een saccharide-peptide-conjugaat dat gemodificeerd kapselpolysaccharide van groep B meningococcus omvat dientengevolge de oorzaak van auto-immuunziekte zou kunnen vormen zijn de meeste onderzoeken die betrekking hebben op een vaccin tegen groep B meningococcus gericht op het potentiële gebruik van subkapselbestanddelen zoals buitenmembraaneiwitten (OMP) en lipopolysacchariden (LPS).
Zo beschrijven Jennings et al. conjugaten van tetanustoxoïde (ΊΤ) met van meningococcus LPS afgeleide gedefosforyleerde oligosacchariden (OS*). De immunogeniciteit van deze OS*-TT-conjugaten werd in konijnen onderzocht (Infect. Immun. 43:^07~4l2). Hierbij werd fosforethanolamine door behandeling met waterstoffluoride van de meningococcus oligosacchariden (OS) verwijderd. De gedefosforyleerde oligosacchariden werden vervolgens aan tetanustoxoïde gekoppeld. Het aldus verkregen immunotype L3 OS-eiwitconjugaat was bij konijnen slechts enigszins immunogeen, hetgeen waarschijnlijk kan worden verklaard als zijnde een gevolg van verwijdering van de PEA-groepen.
In Infection and Immunity, maart 1991 blz 843"851. beschrijven Verheul et al. de bereiding van meningococcus OS-eiwit-conjugaten met teta- nustoxoïde, waarbij de fosforethanolaminegroepen van de oligosacchariden gehandhaafd zijn.
Oligosaccharide-peptideconjugaten, waarbij saccharidedeel en pepti-dedeel afkomstig zijn van verschillende organismen (meestal een tetanus-of difteriepeptide) hebben het nadeel dat overgevoeligheid of tolerantie voor genoemde peptidedelen (tetanus- of difteriedrager) kan ontstaan en tot verminderde respons op het B-cel activerende deel zal kunnen leiden. Derhalve is gezocht naar saccharide-peptide-conjugaten die voorzien zijn van homoloog dragerpeptide. Een dergelijk saccharide-peptide-conjugaat zal bij contact met het micro-organisme in kwestie behalve B-cel geheugen tevens T-cel geheugen reactiveren.
Er zijn saccharide-peptide-conjugaten bekend die een homologe dragerpeptide omvatten. Paton et al. beschrijven conjugatie van pneumolysi-ne-toxoïde aan type 19F kapselpolysaccharide van Streptococcus pneumoniae (Infection and Immunity, juli 1991* biz 2297-2304).
Tevens zijn er saccharide-peptide-conjugaten bekend die een homoloog dragerpetide omvatten die als immuniteit verlenend B-cel activerend deel een saccharidedeel omvatten dat van een lipopolysaccharide (LPS) van een gramnegatieve bacterie is afgeleid. Een dergelijk saccharide-peptide-conjugaat biedt als voordeel de mogelijkheid een vaccin te maken dat immuniteit verschaft tegen gramnegatieve bacteriën, waarvan de kapsel-polysacchariden geen of onvoldoende immuunreactie opwekken. Een saccharide-peptide-conjugaat dat als immuniteit verlenend B-cel activerend deel lipopolysaccharide omvat heeft echter ook nadelen. Lipopolysaccharide bevat toxische delen en een saccharide-peptide-conjugaat dat een lipopolysaccharide met toxische delen omvat zal eveneens toxisch zijn.
In Contrib. Microbiol. Immunol. Basel, Karger ,1989. vol 10, blz I66-I89 beschrijven Cryz, J.C. et al. vaccins tegen Pseudomonas aeruginosa die een saccharide-peptide-conjugaat omvatten dat gedetoxificeerde lipopolysaccharide van Pseudomonas aeruginosa immunotype 5 omvat, welke lipopolysaccharide gekoppeld is met diverse dragereiwitten. De drager-eiwitten kunnen zowel homoloog als niet homoloog zijn. De dragereiwitten die in dit artikel genoemd worden zijn tetanustoxoïde, toxine A en pili van Pseudomonas aeruginosa.
Het LPS van Pseudomonas aeruginosa wordt volgens dit artikel onschadelijk gemaakt door estergebonden vetzuren van Lipide A te verwijderen, zodat het serologisch actieve O-polysaccharidedeel kan worden opgenomen in een saccharide-peptide-conjugaat. Dergelijk gedetoxificeerd LPS (D-LPS) werd omgezet in een actieve ester door koppeling aan N-hydroxysucci- nimide en vervolgens werd de actieve ester gekoppeld aan een eiwit dat ter vergemakkelijking van de koppeling voorzien was van een spacer (1,4-diaminobutaan). Het D-LPS-TA-conjugaat was niet immunogeen maar het D-LPS-pili-conjugaat en het D-LPS-TT-conjugaat wel.
In een artikel van Boons et al (Bioorganic and Medicinal Chemistry Letters, Vol. 1, Nr 6, pp303"308, 1991) wordt beschreven dat men als onderdeel van vaccinontwikkeling met een brede werking tegen N. meningitidis een saccharide-peptideconjugaat (SPC) met twee essentiele immunologische domeinen (d.w.z. de B- en T-epitopen die voor antilichaam-specifi-citeit en T-helperactiviteit verantwoordelijk zijn) die covalent zijn verbonden door middel van een kunstmatige spacer heeft gemaakt. De B-epitoopfunctie wordt bij dit bekende conjugaat geleverd door een fragment van het IC-gebied van het LPS-immunotype-6 (IC=inner core). Voor het opwekken van homologe T-helperrespons (geheugen) is een T-cel-epitoop bevattend peptide gekozen. Het T-cel-epitoop bevattende peptidedeel bij het bekende homologe SPC is het 47=59 deel van een meningococcus OMP, dat door Wiertz E.J.H.J. et al (in Proceedings of the Eleventh American Peptide Symposium, red. River, J.E.; Marshall, G.R.; ESCOM, Leiden, 1990) is beschreven. Nadelen van dit bekende SPC zijn enerzijds de vereiste ingewikkelde en kostbare chemische synthese die vereist is, hetgeen produktie op grote schaal om economische redenen hindert en anderzijds het feit dat slechts een deel van de immuunwerking van OMP wordt benut omdat slechts -een klein fragment van OMP dat een T-cel activerend deel omvat wordt benut, zowel wat beteft B- en T-cel activerende delen.
Tot heden leek het niet mogelijk een saccharidedeel aan een volledig OMP te koppelen of aan een groot OMP-fragment met meer dan een epitoop zonder daarbij de struktuur of samenstelling te veranderen hetgeen de immuunwerking van het OMP zou verlagen.
Verder is bekend dat microorganismen die OMP's produceren deze in hun membranen in een bepaalde configuratie inbouwen, waarbij delen van het OMP in het membraan zijn verankerd en delen in de vorm van lussen uitsteken. De specifieke configuratie is vereist voor de immuunwerking van de in het OMP aanwezige T-cel- en B-cel- activerende delen. In (•♦•REFERENTIE VAN NOREN OF OCTROOIAANVRAGE???*·*) wordt beschreven dat men microorganismen kan behandelen op zodanige wijze dat zij gaan "bleb-ben", dat wil zeggen buitenmembraanvesikels (OMV's) gaan vormen. Dergelijke buitenmembraanvesikels zijn aan hun oppervlak voorzien van duizenden OMP's en bieden de mogelijkheid als krachtig vaccinbestanddeel te worden toegepast omdat er een grote hoeveelheid OMP's in de voor immuun- werking vereiste conformatie aanwezig is. Aan een vaccin dat een dergelijk vaccinbestanddeel omvat kleven echter een aantal van de nadelen van andere vaccins die enkel OMP omvatten als immuunwerking activerend bestanddeel. Bestaande OMV-vaccins blijken bij de mens intermediaire bescherming te kunnen bieden d.w.z. 50-80# bescherming.
De onderhavige uitvinding beoogt de problemen die bestaan voor de diverse bekende vaccinbestanddelen op te lossen. Hiertoe heeft men onderzocht hoe men OMP kan koppelen aan een tweede bestanddeel met immuunwer-kingen zonder de immuunwerking van het OMP te beschadigen. De onderhavige uitvinding heeft derhalve betrekking op een buitenmembraanvesikel dat voorzien is van polypeptiden die ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP’s) hebben, welk buitenmembraanvesikel wordt gekenmerkt doordat de polypeptiden in het membraan verankerde OMP's of OMP-fragmenten zijn met een mutatie in een van de aan het oppervlak gelegen lussen van het OMP waar het polypeptide van is afgeleid. De uitvinding heeft bij voorkeur betrekking op een dergelijk buitenmembraanvesikel, waarbij de polypeptiden in het membraan verankerde OMP's of OMP-fragmenten zijn met een mutatie in een van de aan het oppervlak gelegen lussen van het klasse-l-OMP waar het polypeptide van is afgeleid. In het bijzonder heeft de uitvinding betrekking op een dergelijk buitenmembraanvesikel waarbij het polypeptide ten minste een mutatie in een van de lussen 2, 3. 5. 6, 7 of 8 omvat.
Gevonden is dat het aanbrengen van mutaties in deze lussen de immuunwerking van aan een membraan gebonden OMP niet verlaagt. Hierdoor wordt de mogelijkheid geboden mutaties op deze lokaties aan te brengen die specifieke koppeling van het OMP aan een ander gewenst bestanddeel, bij voorkeur een bestanddeel met aanvullende immuunwerking op de plaats van de mutatie mogelijk maakt. Een buitenmembraanvesikel volgens de uitvinding kan worden beschouwd als een voor koppeling geactiveerd buitenmembraanvesikel .
De mutatie bevindt zich bij voorkeur in een van de lussen 6 of 7· Deze lussen bevinden zich in de tertiaire struktuur van het OMP op de meest gunstige posities ten opzichte van lussen 1 en 4 die een aantal belangrijke epitopen van het OMP omvatten.
De mutatie kan bestaan uit ten minste de aanwezigheid van een additioneel aminozuur met een reactieve zijketen in een van de lussen 2, 3. 5, 6, 7 of 8. Het kan hier gaan om een insertie, deletie of vervanging in een van de lussen. De voorkeur gaat uit naar een vervanging omdat de natuurlijke situatie dan het meest wordt nagebootst. De vorm van de mutatie is niet kritiek, de lokatie wel.
Omdat in natuurlijk OMP geen cysteine aanwezig is en de sulfhydryl-groep een goede reactieve groep is wordt dit aminozuur bij voorkeur aangebracht. Ander aminozuren zoals lysine zijn mogelijk maar door aanwezigheid van dergelijke aminozuren in het natieve OMP wordt voor de koppeling een aantal reactieplaatsen geboden en zullen de resulterende SPV's ge-screened moeten worden om te bepalen welke op de gewenste lokatie een conjugatie hebben ondergaan. Het aanbrengen van beschermende groepen zal in de meeste gevallen leiden tot ingewikkelde chemische reacties met een groot risico van denaturatie van het polypeptide.
Zoals aangegeven kan het polypeptide volledig OMP omvatten maar ook een OMP-fragment. Het OMP-fragment dient voldoende lengte en een zodanige struktuur te bezitten dat het kan worden verankerd in het membraan van een microorganisme en ten minste immuun activerende werking heeft, bij voorkeur evenveel of meer dan het overeenkomstige natieve OMP waar het van afgeleid is.
Een toepassing van een OMV volgens de uitvinding is gelegen in pro-duktie van een vaccinbestanddeel dat tevens een geconjugeerd saccharide-deel omvat. Het produkt van koppeling van een saccharidedeel aan een polypeptidedeel van het voor koppeling geactiveerde buitenmembraanve'si-kel, een buitenmembraanvesikel (OMV) dat een saccharide-peptide-conjugaat (SPC) omvat, een zogenaamd saccharide-peptide-vesikel (SPV) zal een bredere immuunwerking hebben dan de bestaande buitenmembraanvesikels die OMP omvatten. Het SPV zal ook een grotere en bredere immuunwerking hebben dan de bekende SPC's. Een verder voordeel is dat men nu ook het natuurlijke mechanisme, waarbij een microorganisme OMP's in het membraan verankert kan benutten om een vaccinbestanddeel met bredere en sterkere immuunwerking op een economische en eenvoudige wijze te produceren. De onderhavige uitvinding heeft betrekking op een SPV zoals zojuist is beschreven.
De voorkeur gaat uit naar het koppelen van een peptidedeel van een geactiveerd buitenmembraanvesikel aan een saccharidedeel, welk saccharidedeel ten minste een van een lipopolysaccharide (LPS) van een gramne-gatieve bacterie afgeleid immuniteit verlenend B-cel activerend deel met ten minste een epitoop omvat en een buitenmembraanvesikel dat eeen fragment van een uit een gramnegatieve bacterie geïsoleerd B-cel activerend deel met ten minste een epitoop omvat. De uitvinding omvat ook een SPV voorzien van polypeptiden met ten minste de immuunwerking van OMP, welke polypeptiden deel uitmaken van saccharide-peptide-conjugaten, waarbij het saccharidedeel en het peptidedeel van een dergelijk conjugaat gekoppeld zijn op de lokatie van de mutatie in een van de lussen 2, 3. 5» 6, 7 of 8 van het polypeptide.
Een SPV dat als B-cel activerend deel van het saccharidedeel de LPS "core region" of een daarvan afgeleid fragment omvat zal vanwege het ontbreken van toxische delen veiliger zijn als bestanddeel van een vaccin dan een SPV met een saccharide-peptide-conjugaat dat natieve LPS omvat. Een dergelijk "core" bevattend saccharide-peptide-conjugaat heeft bovendien het voordeel dat het een groter aantal verschillende LPS "core" B-cel activerende delen kan bevatten dan een SPC met natieve LPS dat toxisch wordt, wanneer teveel van het toxische bestanddeel lipide A wordt ingebouwd.
Lipopolysaccharide van meningococcus is toxisch en omvat drie delen. In figuur 1 wordt een LPS van meningococcus weergegeven. Het lipide A deel is toxisch en de lacto-N-neotetraose-eenheid, kan eventueel tot inductie van auto-antilichamen leiden. Het oligosaccharidedeel van meningococcus LPS, de zogenaamde "core region" is niet toxisch. De zogenaamde "inner core region" is dat deel van de "core region" van het oligosaccharidedeel van meningococcus LPS waarvan de lacto-N-neotetraose-eenheid ontbreekt. Een saccharide-peptide-conjugaat, dat een B-cel activerend deel omvat, dat de "core" oligosaccharide van meningococcus of een daarvan afgeleid fragment omvat, is een geschikt voorbeeld van een SPC die deel uit kan maken van een SPV volgens de uitvinding.
Weliswaar is in het reeds geciteerde artikel van Boons et al een saccharide-peptide-conjugaat, dat in het van meningococcus afgeleide saccharidedeel geen toxische lipide A en evenmin de volledige lacto-N-ne-otetraose-eenheid omvat beschreven, maar bij dit bekende vaccinbestand-deel is het saccharidedeel gekoppeld aan een van OMP afgeleid fragment dat slechts een T-cel-epitoop omvat en vormt het SPC geen onderdeel van een buitenmembraanvesikel.
Men kan voor een SPV volgens de uitvinding toe te passen natieve LPS isoleren en vervolgens lipide A en een deel van de tetraose-eenheid verwijderen. Lipopolysaccharide (LPS) van meningococcus bijvoorbeeld kan worden geïsoleerd via de extractie werkwijze met heet water en fenol van Westphal (Westphal, 0. en Jann, J.K., 1965, Methods Carbohydr. Chem. 5· 83“91) · In Let kort wordt LPS gehydrolyseerd in 1/Ü-azijnzuur totdat floc-culatie optreedt. Lipide A wordt verwijderd door te centrifugeren en de oligosacchariden worden gezuiverd over een Biogel kolom. Vervolgens kan de hoofd-oligosaccharide worden gekoppeld aan het peptidedeel.
Men kan op een dergelijke wijze verkregen "core" oligosaccharide of een werkzame afgeleide daarvan opnemen in een saccharide-peptide-vesikel volgens de uitvinding.
Omdat het verkrijgen van een van natieve LPS afgeleid B-cel activerend deel een moeizaam proces is en het volledig zuiveren ven de "core" oligosaccharide eveneens problematisch is, kan het B-cel activerende deel van het saccharidedeel van een saccharide-peptide-vesikel volgens de uitvinding worden gesynthetiseerd. Een saccharide-peptide-vesikel volgens de uitvinding kan derhalve een synthetisch B-cel activerend deel in het saccharidedeel omvatten.
Het is ook mogelijk een niet toxisch van een LPS afgeleid B-cel activerend deel van het SPV langs biochemische weg te verkrijgen. Deze route is mogelijk indien men via moleculair biologische methoden een produktiestam maakt die een veranderd lipide A en/of lacto-N-neotetraose kan produceren. In ieder geval zal de terminale galactose van het lacto-N-tetraose verwijderd moeten zijn.
De saccharide-peptide-vesikelen volgens de uitvinding kunnen geconjugeerd zijn via een spacer. Dit heeft als voordeel dat geen intramolecu-laire reacties optreden tussen de reactieve groepen van het saccharidedeel en het peptidedeel. Dergelijke reacties kunnen namelijk leiden tot een gewijzigde tertiaire structuur van het saccharidedeel en/of peptidedeel, met een verslechterde immuunwerking als gevolg.
Een aantal LPS immunotype specifieke epitopen hebben hun werking te danken aan de aanwezigheid van ten minste een fosforethanolamine (PEA) groep. Derhalve verdient een saccharide-peptide-vesikel volgens de uitvinding de voorkeur waarbij PEA-groepen van het B-cel activerende deel van het saccharidedeel vrije aminogroepen omvatten. Een voorbeeld van een dergelijk saccharide-peptide-vesikel volgens de uitvinding, dat een im-muunspecifiek B-cel activerend deel in het saccharidedeel omvat, is een saccharide-peptide-vesikel waarvan het saccharidedeel LPS van meningococcus met immunotype L3 omvat. Een dergelijk saccharide-peptide-vesikel, dat een immuunspecifieke epitoop van immunotype L3 omvat, zal bij voorkeur een B-cel activerend deel omvatten waarvan de PEA-groepen vrije aminogroepen hebben.
Saccharide-peptide-vesikelen volgens de uitvinding omvatten met voordeel een saccharidedeel dat een B-cel activerend deel omvat dat kruisreactie kan geven met meer dan een immunotype. Een vaccin dat een dergelijk saccharide-peptide-vesikel volgens de uitvinding omvat zal bescherming geven tegen meer dan één immunotype.
Momenteel zijn twaalf verschillende typen lipopolysacchariden bekend voor meningococcus stammen (hetgeen overeenkomt met 12 immunotypen). De verschillen in de meningococcus LPS immunotypen zijn gelokaliseerd in het oligosaccharide deel van het LPS ("core region"). Sinds kort zijn de volledige primaire structuren van de oligosacchariden opgehelderd voor immunotypen Llt L2, L3, L5 en L5; deze worden weergegeven in figuur 2 (Difabio J.L. et al, 1990, Structure of the LI and L6 core oligosaccharide of Neisseria meningitidis. Can. J. Chem. 86:1029-1034} Jennings, H.J. et al, 1987, Structure and Immunochemistry of meningococcal lipopolysaccharides, Anthonie van Leeuwenhoek 53:519-522; Michon, F. et al, 1990, Structure of the L5 lipopolysaccharide core oligosaccharide of Neisseria meningitidis, J. Biol. Chem. 256:7243-7247; Verheul, A.F.M. et Infection and Immunity (1991), 51: p 3566-3573). In figuur 3 zijn de structuren van de grootste Llt L2, L3, moleculen weergegeven .
Een saccharide-peptide-vesikel volgens de uitvinding, dat een of meer oligosacchariden met de structuren die in figuur 2 worden weergegeven omvat, is bruikbaar als bestanddeel van een vaccin dat tegen ten minste een immunotype van meningococcus is gericht. Een saccharide-peptide-vesikel volgens de uitvinding, dat ten minste in het saccharidedeel het B-cel activerende deel van ten minste een van de oligosacchariden uit figuur 2 omvat kan derhalve met voordeel als bestanddeel van een vaccin, dat tegen ten minste een immunotype van meningococcus is gericht, worden gebruikt.
De oligosacchariden van de immunotypen verschillen in monosacchari-de-samenstelling, hoeveelheid en locatie van fosforethanolamine (PEA) groepen en de mate van acetylering van de a(l~2) gebonden GlcNAc-eenheid of ander eenheden. Voor de meeste immunotypen is de basis-structuur van de oligosaccharide "core" gelijk (figuur 1 en figuur 2).
Omdat de "core" oligosacchariden van meningococcus LPS voor diverse immunotypen overeenkomsten vertonen, omvat de "core" meer dan een immunotype specifieke epitoop. Een SPV volgens de uitvinding dat een dergelijke "core" als B-cel activerend deel van het saccharidedeel omvat, zal de voorkeur verdienen. Een dergelijk SPV volgens de uitvinding kan voor een aantal verschillende immunotypen tegelijk B-cel activerend werken. Een saccharide-peptide-vesikel volgens de uitvinding, dat voor een aantal immunotypen tegelijk B-cel activerend kan werken kan ook een afgeleid fragment omvatten, dat meer dan een immunotype specifieke epitoop omvat.
Het is ook mogelijk slechts een relevant gedeelte van de "core" oligosaccharide als fragment op te nemen in een SPV volgens de uitvinding, dat specifiek immuniteit voor een specifiek immunotype verschaft. Het is ook mogelijk in het saccharidedeel van een SPV volgens de uitvinding meer dan één B-cel activerend deel op te nemen, zodat ëen dergelijk SPV diverse specifieke immuniteit verlenende B-cel activerende delen omvat en dus immuniteit voor diverse specifieke immunotypen kan verschaffen.
Men kan met voordeel B-cel activerende delen van het saccharidedeel met specifieke immuniteit verlenende werking tegen verschillende immunotypen of B-cel activerende delen van het saccharidedeel met kruisreactie-ve immuniteit verlenende werking in het saccharidedeel van een sacchari-de-peptide-vesikel volgens de uitvinding opnemen. Het is mogelijk geworden goed gedefinieerde oligosacchariden met toenemende complexiteit en molecuulgewicht te synthetiseren, zodat oligosacchariden die een of meer B-cel activerende structuren omvatten gesynthetiseerd kunnen worden.
Van de meningococcus is bekend dat immunotypen L3 en L2 respectievelijk ongeveer 70# en 30# van de groep B meningococcen meningitis veroorzaken. Derhalve verdienen saccharide-peptide-vesikelen volgens de uitvinding de voorkeur die ten minste B-cel activerende delen van L3 en/of L2 immunotypen in het saccharidedeel van het saccharide-peptide-vesikel omvatten.
Een saccharide-peptide-vesikel volgens de uitvinding dat ten minste een B-cel activerend deel van het saccharidedeel omvat dat een afgeleide is van
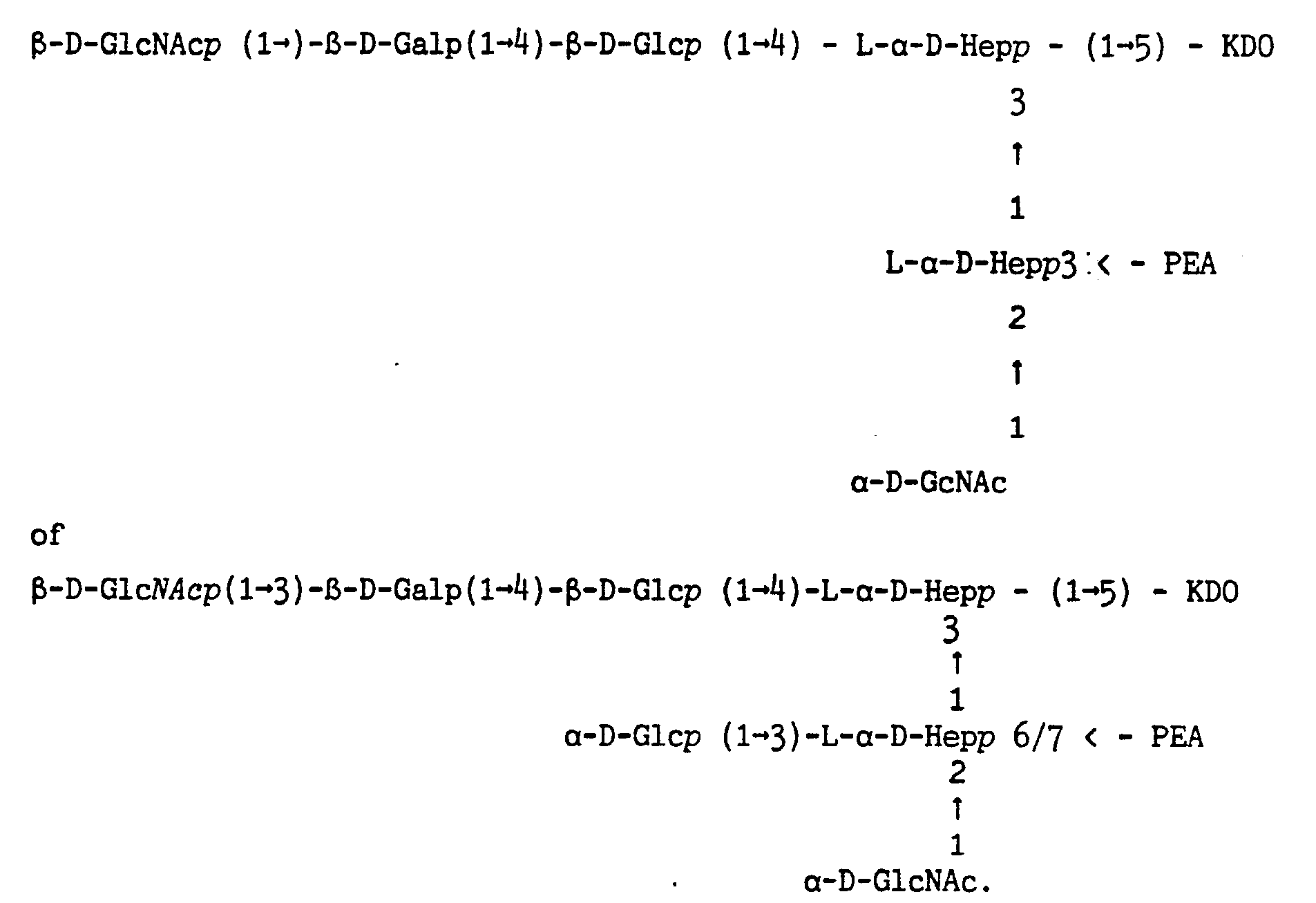
Een saccharide-peptide-vesikel dat de voorkeur verdient is een SPV dat een B-cel activerend deel in het saccharidedeel omvat dat ten minste kruisreactie vertoont met twee immunotypen van gram-negatieve bacteriën. Een voorbeeld van een dergelijk saccharide-peptide-vesikel vertoont ten minste kruisreactie met meningococcus immunotypen L2 en L3.
Een saccharide-peptide-vesikel dat eveneens de voorkeur verdient is een SPV dat in het saccharidedeel een B-cel activerend deel omvat dat ten minste kruisreactie vertoont met meningococcus immunotypen L: en L3.
Een saccharide-peptide-vesikel dat kruisreactie vertoont met ten minste immunotypen Llt L2 en L3 omvat als B-cel activerend deel in het saccharidedeel ten minste de vertakte oligosaccharide 6-D-Glcp(l-*i0-[L-a-D-Heppil-^)]-L-a-D-Hepp). Een geschikt voorbeeld van een dergelijk saccharide-peptide-vesikel volgens de uitvinding omvat tevens op de vertakte oligosaccharide B-D-Glcp(l-*4)-[L-a-D-Hepp(l-*3)]~L-a-D-Hepp) een PEA-groep op de 0-3 van de heptosyleenheid.
Een saccharide-peptide-vesikel volgens de uitvinding, dat kruisreactie vertoont met meer immunotypen dan Llt L2 en L3) omvat als B-cel activerend deel in het saccharidedeel ten minste de vertakte oligosaccharide 6-D-Glcp(l-*4)-[L-a-D-Hepp(l-*3) ]-L-a-D-Hepp) die voorzien is van een spacer die verbinding geeft met ten minste een ander immunotype specifiek B-cel activerend deel met ten minste een epitoop.
Een SPV volgens de uitvinding kan als B-cel activerend deel een saccharidedeel omvatten dat gesynthetiseerd en/of gemodificeerd is ten opzichte van een natuurlijk B-cel activerend deel voor een bepaald immunotype. Een dergelijk gesynthetiseerd en/of gemodificeerd B-cel activerend deel zal met voorkeur kruisreactie geven met diverse immuno-typen en/of een verbeterde immuunreactie geven dan het overeenkomstige deel van het natuurlijke LPS. In ieder geval zal de terminale galactose (figuur 2) verwijderd dienen te zijn, hetzij via enzymatische weg, hetzij via genetische manipulatie, door mutatie.
Een dergelijk gesynthetiseerd en/of gemodificeerd B-cel activerend deel kan ontstaan door selectieve additie en/of deletie van bepaalde groepen en/of suikereenheden.
Een saccharide-peptide-vesikel volgens de uitvinding zal bij voorkeur als peptidedeel een buitenmembraaneiwit (OMP) of een van een OMP afgeleid fragment omvatten. Bij voorkeur zal een SPV volgens de uitvinding een OMP van meningococcus Klasse I omvatten omdat dit OMP beschermende aantistoffen kan opwekken, niet de neiging vertoont om antistoffen gericht tegen andere eiwitten te blokkeren en een niet al te grote anti-gene variatie vertoont. Op het Klasse I buitenmembraaneiwit van meningococcus /76 zijn ook vele herkenningsplaatsen onderzocht voor menselij-• ke T-cellen, waardoor het een geschikte uitgangsstam voor toe te passen OMP is.
De onderhavige octrooiaanvrage heeft ook betrekking op een werkwijze voor het bereiden van een buitenmembraanvesikel volgens de uitvinding.
Deze werkwijze omvat de volgende stappen 1) men brengt een nucleotidesequentie coderend voor het polypeptide tot expressie in een bacterie; 2) men kweekt de bacterie onder op zich voor het produceren van buiten-membraanvesikels bekende omstandigheden, zoals in NIPH ANNALS, Volume 14, nummer 2, December 1991 door Frederiksen J.H. et al op p6?~ 79. in het bijzonder opp 69-71 en de aldus ontstane membraanstukken met polypeptide kan men eventueel isoleren.
Voor het bereiden van een SPV volgens de uitvinding kan men vervolgens 3) de in stap 2 ontstane membraanstukken voorzien van saccharide-pep-tide-conjugaten door het polypeptide te koppelen aan het gewenste saccharidedeel.
Men kan voor het bereiden van een OMV of een SPV volgens de uitvinding als de voor stap 1) benodigde nucleotidesequentie een recombinant nucleotidesequentie toepassen die voor OMP of een fragment daarvan met ten minste de immuunwerking‘van OMP codeert, waarbij in vergelijking tot de overeenkomstige natuurlijke sequentie een mutatie is aangebracht in een van de aan het oppervlak gelegen lussen. De voorkeur gaat uit naar een mutatie in een van de lussen 2, 3. 5. 6, 7 of 8, in het bijzonder naar lussen 6 en 7* De mutatie resulteert in een codon voor ten minste een aminozuur met een specifiek reactieve zijketen in een van de oppervlakte lussen. In het bijzonder resulteert de mutatie in een codon voor cysteine. Men kan bij de werkwijze voor het bereiden van een SPV volgens de uitvinding een via recombinant DNA-technieken verkregen saccharidedeel toepassen of een langs synthetische weg verkregen saccharidedeel.
Voor het koppelen van een saccharidedeel aan een OMV volgens de uitvinding in het bijzonder aan een OMV-SH die voorzien is van een cysteine-groep kan men de principe van het volgende reactieschema volgen:
COre-KD0-C00H + H2N-X-Br e°re-KDO-CO-NH-X-Br (EDO 1-ethyl-3-dime thylaminopropylearbodi imide) core-KDO-CO-NH-X-Br + OMV-SH------ core-KDO-CO-NH-X-S-OMV
Meer specifiek kan de reactie als volgt worden uitgevoerd:
BrCH2-C0-NH-(CH2)ή-ΝΗ2 (stof 1, overmaat) + OS-KDO + EDC + sulfo-NHS ^ · os co-cH2Br (st0f 2). Stof 2 wordt onderworpen aan gelfiltratie en vervolgens wordt stof 2 + OMV-SH —-* conjugaat.
Een andere mogelijkheid de koppeling uit te voeren is met BrCH2-C0-NH-(CH2)/,-NH-C0-CH2Br (stof 3. overmaat) + OS-SH (zoals beschreven op pagina 34 en 35 van het proefschrift van Andre Verheul, Meningococcal LPS derived oligosaccharide protein conjugate vaccines, te verdedigen op 29 oktober 1991. Utrecht). Dit resulteert in OS- - -C0-CH2Br (stof 2*). Deze wordt aan gelfiltratie of eventueel aan een simpele ET20-was onderworpen. Stof 2* kan vervolgens met OMV-SH worden omgezet tot het conjugaat. Aangezien stof 3 lipofiel is geschiedt de modificatie van OS-SH in een mengsel van een waterige buffer en een organisch oplosmiddel, zoals dioxaan. Om deze reden is de reactie van OMV-SH met stof 3» na zuivering gevolgd door incubatie met OS-SH minder aantrekkelijk.
Voor het specifiek koppelen van koolhydraten is de aanwezigheid van geschikte groepen zoals een amino (-NH2), carbonzuur (-C00H), thiol (-SH) of een aldehyd (CHO) noodzakelijk binnen het koolhydraatantigeen (Dick, W.E. et al, 1989. Glycoconjugates of bacterial carbohydrate antigens. In Cruse J.M. en R.E. Lewis, Conjugate vaccines, Contrib. Microbiol. Immunol., Basel, Krager, 10:48-114). Deze groepen kunnen aanwezig zijn in het antigeen maar moeten vaak via chemische of enzymatische werkwijzen worden opgenomen. Het verdient de voorkeur een koppelingswijze toe te passen die resulteert in zo min mogelijke modificatie van het koolhydraa- en eiwitantigeen. Het gebruik van een specifiek aan te brengen spacer is hiervoor bevorderlijk. Het verdient de voorkeur zo min mogelijk de aanwezige epitoop of epitopen te vernietigen of ongewenste immuno-dominante neo-antigenen te genereren. De invloed van de koppelingswerkwijze op de immunologische eigenschappen van een oligosaccharide-peptidevesikel is groot wanneer kleine oligosacchariden worden toegepast (Hoppner, W., et al , 1985, Study on the carbohydrate specificity of antibodies formed in rabbits to synthetic glycoproteins with the carbohydrate structure of asialoglycophorin A. Mol Immunol. 12:13^1-13^8).
De meeste bestaande vaccins zijn gebaseerd op kapselpolysacchariden of O-antigenen die bestaan uit zich herhalende eenheden van 1 tot 8 mono-sacchariden zodat de invloed van de koppeling kan worden geminimaliseerd door het gebruik van grotere oligosacchariden. Meningococcus LPS bevat geen herhalende eenheden en derhalve zal met name de keus van de koppelingswerkwijze bij het gebruik van meningococcus LPS van belang zijn voor de immunologische en immunochemische eigenschappen van het resulterende conjugaat.
Bij voorkeur koppelt men dus de saccharide aan het peptide via een spacer die aan de uiteinden reactieve groepen zoals NH2 en COOH bevat. Het gebruik van een spacer bij saccharide-peptide-vesikelen volgens de uitvinding heeft als voordeel dat de tertiaire structuur van het saccha-ridedeel niet gewijzigd wordt, hetgeen van belang is voor de immuun-werking van de saccharide.
Bij de meningococcus oligosacchariden zijn er twee groepen beschikbaar die kunnen worden gebruikt voor het koppelen van een dergelijke oligosaccharide aan dragerpeptide: de vrije aminogroep van de fos-forethanolaminegroep (PEA-groep) en de carbonzuurgroep van de KDO-eenheid (figuur 1). De PEA-groep dient bij voorkeur niet te worden gebruikt voor de koppeling omdat deze groep waarschijnlijk deel uitmaakt van een aantal van de immunotype specifieke epitopen. Derhalve verdient het de voorkeur in een saccharide-peptide-vesikel volgens de uitvinding fosforethanol-aminegroepen te behouden met vrije aminogroepen. Gewoonlijk kunnen saccharide-peptide-conjugaten worden gemaakt op basis van het koppelen van de carbonzuurfunctie van de oligosaccharide aan de vrije aminogroepen van het peptide. Wanneer deze werkwijze wordt toegepast bij de meningococcus oligosaccharide kan dit resulteren in koppeling van oligosaccharide aan oligosaccharide of van oligosaccharide aan carbon-zuurgroepen van het peptide door de PEA-groep van de oligosaccharide. Jennings et al. (Infect. Immun. 43;^07“4l2) hebben dit probleem opgelost door de PEA-groepen te verwijderen door behandeling met waterstof-fluoride. De gedefosforyleerde oligosacchariden werden vervolgens gekoppeld aan tetanustoxoïde door β-(ij-aminofenyl)ethylamine als spacer aan het reducerende uiteinde op te nemen door middel van reductieve aminering (hetgeen leidde tot verlies van de ringstructuür van KDO), gevolgd door activering van de aminofunctie met thiofosgeen gevolgd door koppeling aan tetanustoxoïde. Dergelijke conjugaten met immunotype L2, L5 en L10 waren zeer immunogeen in konijnen terwijl die van L3 slecht immunogeen waren. Voor deze laatste groep blijkt het verlies van PEA-groepen en/of ringstructuur van de KDO-groep van belang te zijn voor de immuniteit en verdient het de voorkeur een saccharide-peptide-vesikel met L3 oligosaccharide van meningococcus te maken, waarbij de PEA-groepen en de KDO-ringstructuur van de meningococcus oligosaccharide zo min mogelijk zijn gemodificeerd.
Voor het bereiden van een saccharide-peptide-vesikel volgens de uitvinding waarvan de B-cel activerende werking van het saccharidedeel afhangt van de aanwezigheid van een of meer fosforethanol (PEA) groepen voldoen bovengenoemde werkwijzen niet. Voor het bereiden van dergelijke saccharide-peptide-vesikelen zijn additionele maatregelen nodig.
Een mogelijkheid voor het bereiden van een saccharide-peptide-vesikel volgens de uitvinding dat het bovengenoemde probleem oplost is het gebruiken van een saccharidedeel dat synthetisch verkregen is.
Met de huidige technieken is het mogelijk oligosacchariden te synthetiseren. Bovendien, zoals eerder beschreven is in deze octrooiaanvrage, kan het, in het geval van saccharide-peptide-vesikelen die van LPS afgeleide oligosacchariden omvatten, voordelig zijn het B-cel activerende deel dat zich in de "inner core" van LPS bevindt te synthetiseren. Een ander voordeel van een synthetisch verkregen sacccharidedeel is gelegen in de mogelijkheid de minimale saccharide die B-cel activerend werkt in het SPC volgens de uitvinding op te nemen. Bovendien is het ook mogelijk een saccharidedeel te synthetiseren dat een minimale oligosaccharide omvat met kruisreactieve immuniteit verlenende werking. Het is ook mogelijk B-cel activerende delen te synthetiseren die een betere immuunwer-king geven dan de natuurlijke oligosacchariden.
Men kan met een synthetisch verkregen saccharidedeel eenvoudiger de spacer op een specifieke plaats aanbrengen. Men kan tevens een of meer fosforethanol groepen inbouwen in het B-cel activerende deel van het saccharidedeel ter verkrijging van een verbeterde immuunwerking.
Teneinde de spacer op een specifieke plaats in de saccharide aan te brengen verdient het de voorkeur de reactieve groepen van de saccharide van beschermende groepen te voorzien voordat men de spacer koppelt. De reactieve groep waar de koppeling met de spacer plaatsvindt voorziet men niet van een beschermende groep. Door deze werkwijze wordt voorkomen dat groepen die essentieel zijn voor de immuniteit verlenende werking tijdens het koppelen van de spacer worden verwijderd of gewijzigd.
De uitvinding heeft tevens betrekking op een nucleotidesequentie die voor recombinant-OMP of voor een recombinant-OMP-fragment codeert welke zich onderscheidt van niet-recombinant-OMP of een niet-OMP-fragment door ten minste een mutatie in een van de lussen 2, 3. 6 „ 7 of 8 van het OMP. In het bijzonder heeft de uitvinding betrekking op een dergelijke nucleotidesequentie met een mutatie in een van de lussen 6 of 7· de voorkeur gaat uit naar een sequentie die tevens codeert voor de belangrijkste antigene determinanten die in lussen 1 en 4 liggen. De nucleotidesequentie kan als mutatie een insertie, deletie of wijziging van ten minste een codon omvatten, waarbij de voorkeur uitgaat naar een mutatie, waarbij ten minste een codon dat voor een additioneel aminozuur met een reactieve zijgroep codeert in een van de lussen 2, 3* 6, 7 of 8 ontstaat in vergelijking tot de aminozuursequentie van het overeenkomstige OMP of OMP-fragment. De nucleotidesequentie volgens de uitvinding omvat verder bij voorkeur een codon dat voor cysteine codeert.
De uitvinding heeft eveneens betrekking op een expressievector die de nucleotidesequentie volgens de uitvinding omvat, alsmede op een mi-croorganisme dat de nucleotidesequentie en/of de expressievector volgens de uitvinding omvat.
Ook is de uitvinding gericht op een polypeptide met ten minste de immuunwerking van OMP welk polypeptide wordt gekenmerkt door aanwezigheid van een additioneel aminozuur met een reactieve zijketen in een van de oppervlaklussen, in het bijzonder lussen 2, 3« 5. 6, 7 of 8, in vergelijking met overeenkomstige natieve OMP of een overeenkomstig fragment van het natieve OMP. Een polypeptide met een aminozuur met een specifiek reactieve zijketen in lus 6 of 7 heeft de voorkeur omdat in de tertiare struktuur van OMP deze lussen. de minste hindering zullen geven van lussen 1 en 4 wanneer een saccharidedeel wordt gekoppeld. Bij voorkeur omvat een polypeptide volgens de uitvinding een cysteine als aminozuur. Aangezien natief OMP geen cysteine omvat kan zeer specifiek aan cysteine worden gekoppeld in een polypeptide volgens de uitvinding. Het aanbrengen van een additionele lysine behoort ook tot de mogelijkheden, maar verdient niet de voorkeur vanwege de aanwezigheid van meer dan een lysine in na-tief OMP en dus ook in het polypeptide. De koppeling zal meer dan een conjugaat opleveren en er zal gescreened moeten worden naar de juiste conjugaat.
Een saccharide-peptide-conjugaat dat ten minste een epitoop van een OMP omvat dat gekenmerkt wordt door aanwezigheid van een polypeptide volgens de uitvinding valt ook onder de strekking van de uitvinding, evenals een vaccin dat een buitenmembraanvesikel, een SPV, een polypeptide of een saccharide-peptide-conjugaat volgens de uitvinding in een werkzame hoeveelheid omvat.
VOORBEELD I Samenvatting
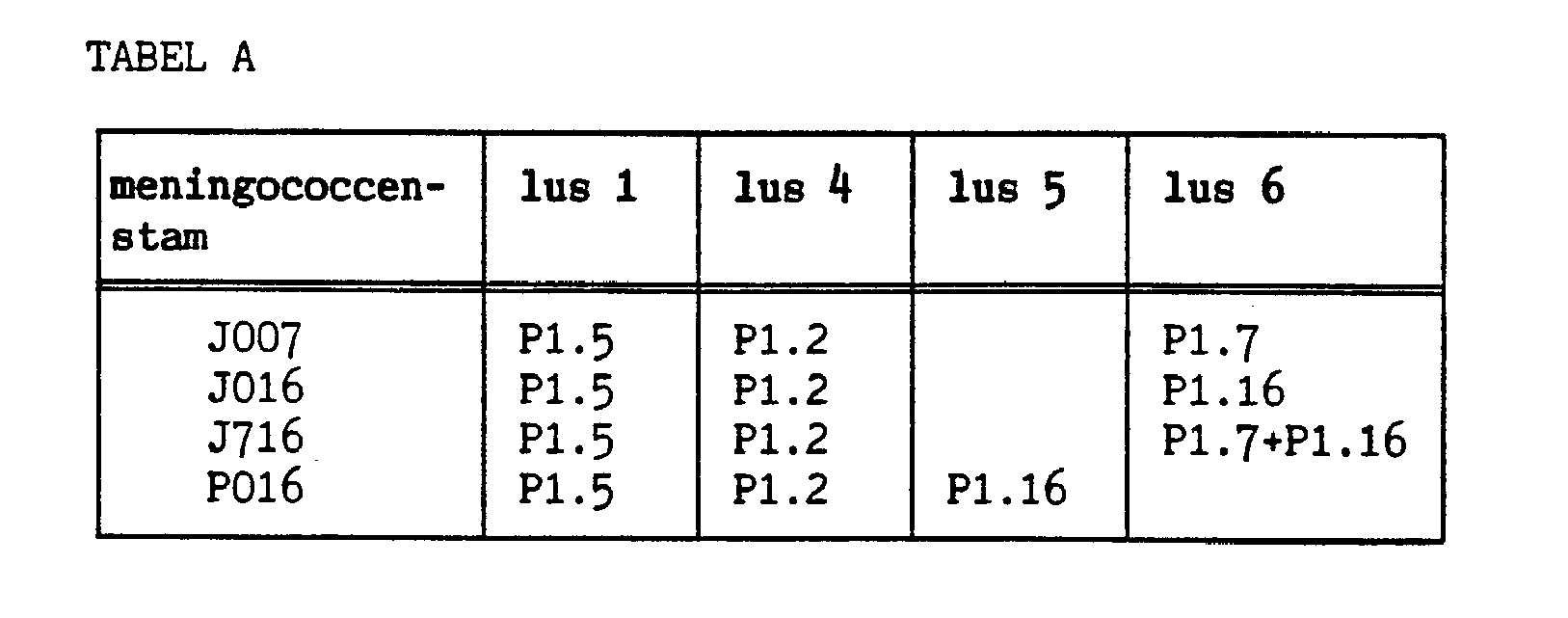
Voor de vaccin-ontwikkeling tegen B meningococcen wordt gebruik gemaakt van klasse 1 buitenmembraaneiwitten. Deze eiwitten bevatten van nature twee variabele gebieden (=epitopen), welke bactericide antilichamen binden. Deze epitopen bevinden zich in twee aan de oppervlakte blootgestelde lussen 1 en 4. Er zijn vier meningococcen-stammmen gemaakt met extra epitopen in lussen 5 en 6 van het klasse-l-eiwit (zie tabel A). De stammen hebben allen de natuurlijke epitopen van de ouderstam Αδ-6* in lussen 1 en 4, namelijk de PI.5 en PI.2 epitopen. Verder heeft stam J007 één extra PI.7 en stam J016 één extra P1.16 epitoop, terwijl stam J7l6 twee extra epitopen, de PI.7 én de P1.16 in lus 6 heeft. Als laatste heeft stam P0l6 in lus 5 één extra P1.16 epitoop. Al deze stammen zijn nog in staat om monoklonale antilichamen aan het celoppervlak te binden.
Met behulp van deze informatie kunnen nieuwe meningococcenstemmen worden geproduceerd, die mutaties in OMP bevatten, welke stammen kunnen worden toegepast om buitenmembraanvesikelen (Outer Membrane Vesicles (=0MV's)), die uit stukken buitenmembraan bestaan en door N. meningitidis onder bepaalde omstandigheden worden uitgescheiden te produceren. Deze stukken kunnen vervolgens worden gekoppeld aan LPS. De OMVs kunnen als vaccin tegen B-meningococcen dienen.
n
1.1 De structuur van het klasse-l-OMP
De klasse 1 OMPs zijn cation-selectieve porie-eiwitten [15]· Het eiwit is ca. 374 aminozuren lang en wordt vooraf gegaan door een signaalpeptide van 19 aminozuren [1]. Een model voor de topologie van het eiwit in het buitenmembraan voorspelt acht aan de oppervlakte blootgestelde lussen (zie figuur 4).
Tussen de verscheidene Neisseria porines zijn de transmembraanse-quenties behouden en zij vormen een β-sheet structuur. De aan de oppervlakte blootgestelde hydrofiele lussen vertonen grote variatie in lengte en sequentie. Wanneer de verschillende klasse-1-OMPs worden vergeleken, vertonen de sequenties een homologie van $0%. De variatie in de sequenties blijkt zich in de eerste en de vierde lus te bevinden. Deze worden aangeduid met variabele gebieden of epi-topen. Bactericide antilichamen blijken zich aan de epitopen in lussen 1 en 4 te binden, hetgeen overeenkomt met het feit dat deze lussen het langst zijn (zie fig. 4). Deze twee epitopen bepalen de subtype-specificiteit van het klasse-l-OMP [6].
1.2 Inbouw van extra epitopen in het meningococcen klasse-l-OMP
Wanneer de verschillende epitopen van de klasse-l-OMPs worden vergeleken, zijn er in totaal 10 verschillende subtype-specifieke epitopen te onderscheiden. Van twee epitopen, de PI.7 en P1.16, zijn oligonucleotiden met "sticky ends" gesynthetiseerd. Deze oligonucle-otiden zijn vervolgens in lus 5 en lus 6 van het gebruikte klasse 1-OMP-gen geplaatst.
De inbouw van de PI.7 en P1.16 oligonucleotiden in het klasse-l-OMP-gen leverde nieuwe stammen op met extra epitopen in het klasse-l-OMP. Deze stammen hebben reeds in lus 1 de PI.5 epitoop en in lus 4 de PI.2 epitoop. De nieuw ingebouwde epitopen, PI.7 en P1.16, zijn in lus 5 of 6 geplaatst. De expressie van de ingebouwde epitopen in de nieuwe stammen is vervolgens met whole cell ELISA bekeken en vergeleken met de expressie van de ouderstam. Vervolgens zijn van zowel de nieuwe stammen als de ouderstam de outer membrane complexes (OMCs) geïsoleerd, welke gebruikt worden voor de immunisatie van muizen.
1.3 Materialen
Het gebruikte DNA & plasmide: - Het plasmide pTZ19R is een plasmide afkomstig van Pharmacia, welke codeert voor o.a. Amp-resistentie (Ampr) en het lac Z -gen. Het gebruikte plasmide 2-2*SE is een afgeleide van pTZ19R. Dit plasmide bevat een 1.9 kB insert met een deel van het klasse-l-OMP-gen uit meningococcenstam 2996 (PI.5.2), vanaf aminozuur 100. Dit insert bevindt zich in de EcoRI site van de Multiple Cloning Site (=MCS) van pTZ19R. Door een deletie in de Multiple Cloning Site, van de Sail site tot de EcoRI site (waar het insert begint), is het klasse-l-gen in hetzelfde reading frame gekomen als het lac Z' van pTZ19R. Het klasse-l-OMP-gen in dit plasmide bevat een Kpnl-restric-tie-site in lus 5·
Een ander gebruikt plasmide pPH2CA, is weer een afgeleide van 2-2*SE. Het klasse-l-OMP-gen in dit plasmide bevat een nieuwe Kpnl-restrictie-site, namelijk in lus 6 i.p.v. lus 5· De oorspronkelijke Kpnl site in lus 5 is vervangen door een BamHI-restrictiesite en vervolgens is door PCR-mutagenese een Kpnl-site in lus 6 gemaakt [17]·
De gebruikte enzymen:
De gebruikte enzymen zijn allen afkomstig van Boehringer Mannheim, tesamen met de bijgeleverde buffers.
Kpnl (vol. act.: 10 U/μΙ), met incubatie-buffer L.
Spel (vol. act.: 10 U/μΙ), met incubatie-buffer H.
Sna BI (vol. act.: 8 U/μΙ), met incubatie-buffer M. T4-DNA-ligase (vol. act.: 1 U/μΙ), met T4-DNA-ligase-buffer.
Met bijbehorende incubatie-buffers: M-buffer (lx): 10 mM Tris-HCl; 10 mM MgCl2; 50 mM NaCl; 1 mM dithioerythritol (DTE), pH 7-5 (bij 37’C) - L-buffer (lx): 10 mM Tris-HCl; 10 mM MgCl2; ImM dithi oerythritol (DTE), pH 7.5 (bij 37'C)
H-buffer (lx): 50 mM Tris-HCl; 10 mM MgCl2; 100 mM
NaCl; 1 mM dithioerythritol (DTE) pH 7-5 (bij 37ec) T4-DNA-ligase-buffer (10x):660 mM Tris-HCl; 50 mM MgCl2; 50 mM dithioerythritol; lOmM ATP; pH 7*5 (bij 37°C)
De gebruikte monoklonale antistoffen gericht tegen de epitopen en LPS-typen:

alle monoklonalen afkomstig van RIVM
De overige materialen staan vermeld bij de methoden (l.k).
De gebruikte bacteriestammen: E. colt K12 NM522: Deze bacteriestam is Hsd' (=bevat geen restrictie-modificatie systeem) - meningococcenstam H44/76 B" (Pl.7.16): Deze bacteriestam is een kapselloze, klasse 3 deficïente mutant van H44/76 (PI.7.16).
- meningococcenstam Α8-6* B~ (PI.5.2): Deze bacteriestam is een kapselloze mutant H44/76 (Pl.7.16), welke getransformeerd is met het klasse-l-eiwit van 2996 (PI.5»2) [17]· 1.4 Methoden 1.4.1 Controle van pPH204 door transformatie naar meningococcen
Het geconstrueerde plasmide pPH204 (PI.2) is naar meningococcenstam H44/76 B' (Pl.7.16) getransformeerd. Hier werd de P1.16 epitoop in lus 4 van H44/76 B" uitgewisseld met de PI.2 epitoop van plasmide pPH204. Met behulp van kolonieblotting en immunoblotting (zie verder) zijn PI.2* kolonies geïsoleerd, die na controle met SDS-PAGE, Western- en immunoblotting de stam J072 oplevert, zie ook tabel B bij Resultaten (1.5)·
1.4.2 Inbouw van extra epitopen in meningococcen klasse 1 OMP
1.4.2.1 Inbouw van oligonucleotiden in de plasmiden pPH204 en 2-2*SE
Het gebruikte klasse-l-OMP-gen in plasmide pPH204 bevat in het gedeelte coderend voor lus 6 een restrictie site voor KpnI. Hierin is een oligonucleotide met KpnI sticky ends geplaatst, welke codeert voor de PI.7 epitoop zoals hieronder weergegeven en bij bereiding & insertie van oligonucleotiden (1.4.3). Door inbouw van de oligonucleotide verdween de KpnI restrictie site en is een unieke Spel restrictie site gecreëerd. Dit plasmide wordt construct pJB007 genoemd .
2.1 oligonucleotide van de PI.7 epitoop met Kpnl sticky ends:
NGGASGQVKVT
5’-g|aac ggc ggc gcc tct ggc CAA GTT AAA GTT ACT|AGTAC-3'
3'-catgc|ttg ccg ccg cgg aga ccg GTT CAA ΊΤΓ caa tga|t-5’ VAAGRALNLNS
restrictie enzym: restrictie enzym:
A/CTAGT GGTAC/C
TGATC/A C/CATGG
Spel Kpnl
Bij insertie van de oligonucleotide in het plasmide, verdwijnt de Kpnl site en verschijnt een unieke Spel site: links: rechts: 5’-ggtacg|aac ggc-3’ 5'-gtt act|agtacc-3’ 3’-ccatgc|ttg ccg-5’ 3’-caa tga|tcatgg-5'
Op dezelfde wijze is een oligonucleotide met Kpnl sticky ends in lus 6 geplaatst, welke codeert voor de P1.16 epitoop zoals hieronder weergegeven en bij bereiding & insertie van oligonucleotiden (1.4.3). Ook hier verdween door inbouw van de oligonucleotide de Kpnl restrictie site, maar is er een unieke SnaBI restrictie site gecreëerd. Dit plasmide wordt construct pJB0l6 genoemd.
2.2 oligonucleotide van de P1.16 epitoop met Kpnl sticky ends: ytkdtnnnltl 5’-g|tat acc aaa gac acc aac aac aac ttg acc ttg|AGTAC-3*
3'-catgc|ata tgg ttt ctg ttg ttg ttg ttg aac ttg AAC|T-5’ IGFVGVVVQGQ
restrictie enzym: restrictie enzym:
TAC/GTA GGTAC/C
ATG/CAT C/CATGG
SnaBI Kpnl
Bij insertie van de oligonucleotide in het plasmide, verdwijnt de Kpnl site en verschijnt een unieke SnaBI site: links: rechts: 5’-ggtacg|tat acc-3’ 5’-acc ttg|agtacc-3’ 3*-ccatgc|ata TGG-5' 3’-TGG aac|tcatgg»5'
In het construct pJB007 is vervolgens een oligonucleotide met Spel sticky ends achter de oligonucleotide PI.7 in lus 6 geplaatst» waarbij de oligo codeert voor de P1.16 epitoop zoals hieronder weergegeven. Hierbij verdween door inbouw van de oligonucleotide de Spel restrictie site. Dit plasmide wordt construct pJB7l6 genoemd.
2.3 oligonucleotide van de Pl.l6 epitoop met Spel sticky ends:

restrictie enzym:
A/CTAGT
TGATC/A
Spel
Door nu de ingebouwde oligonucleotide van de PI.7 epitoop te knippen met Spel, gevolgd door een insertie met de oligonucleotide van de PI.16 epitoop (met Spel sticky ends), verdwijnt de Spel site: links: rechts: 5’-actagc|tat acc-3’ 5'-acc ttg|tctagt-3’ 3*-tgatcg|ata TGG-5' 3’-TGG AACiAGATCA-5*
Het gebruikte klasse-l-OMP-gen in plasmide 2-2*SE bevat in het gedeelte coderend voor lus 5 een Kpnl restrictie-site. Hier is een PI. 16 oligonucleotide in geplaatst. Door inbouw verdween de Kpnl restrictie site. Dit construct wordt pPH0l6 genoemd.

1.4.2.2 Transformatie van constructen naar E. coli
Vervolgens is er met de vier constructen naar E. coli K12 NM522 getransformeerd (zie verder). De transformanten zijn onder selectie-druk van 100 pg/ml Ampicilline gegroeid. Van de ontstane kolonies is plasmide geïsoleerd (zie verder) en gecontroleerd op aanwezigheid van de juiste constructen. Construct PJB007 is gecontroleerd op de verdwenen Kpnl en de ontstane Spel site, het construct pJB0l6 is gecontroleerd op de verdwenen Kpnl en de ontstane SnaBI site en het construct pJB7l6 is gecontro-leerd op de verdwenen Spel site. Mbv. SDS-PAGE, Western blotting & immunoblotting (zie verder) met de geschikte monoklonalen is bekeken of de inserties van de PI.7 en P1.16 epitoop gelukt zijn, dus expressie vertonen en antilichamen binden.
1.4.2.3 Transformatie van constructen naar meningococcen
Vervolgens zijn de constructen (pJB007, pJB0l6, pJB7l6) gebruikt om meningococcenstam Αδ-β* B' (PI.5,2) te transformeren, waarbij recom-binatie van het meningococcen chromosomaal DNA en het construct optrad. Met behulp van kolonieblotting en iraminoblotting (zie verder) zijn PI.7* en de P1.16* kolonies geïsoleerd, die na controle met SDS-PAGE, Western & immunoblotting de gewenste stammen opleverden, zie ook tabel C bij Resultaten.
1.4.2.4 Controle van nieuwe stammen met whole cell Elisa
Met whole cell Elisa (zie verder) is gekeken naar de mate waarin de ingebouwde epitopen in de stammen J007, J016, J716, J072 en P0l6 tot expressie worden gebracht en deze expressie is vergeleken met die van de ouderstammen.
1.4.3 Bereiding & insertie van oligonucleotiden
De complementaire niet-gefosforyleerde oligonucleotiden zoals hierboven zijn weergegeven, zijn gesynthetiseerd op de Applied Biosys-tems 3814 DNA synthesiser. Hybridisatie vond plaats door de complementaire oligonücleotiden in gelijke concentratie bij elkaar te voegen, te verwarmen tot 95 °C en vervolgens af te koelen naar kamertemperatuur. Hierbij ontstonden de oligo's met de juiste sticky ends. Het plasmide is met het juiste enzym gedigesteerd, waarna oligonucleotiden o/n bij 15®C mbv. T4 ligase in het plasmide geli-geerd zijn (in de verhouding 200 ng oligo/1 pg plasmide). Het liga-tiemengsel is vervolgens tot êO°C verwarmd en langzaam afgekoeld tot kamertem-peratuur om er voor te zorgen dat slechts 1 oligonucleotide werd ingebouwd. Door inbouw van de oligo’s in het plasmide verdween de gebruikte restrictie site. Het ligatie-mengsel is met dit enzym nageknipt, waardoor de zelfsluiters onder de plasmiden lineair zijn geworden. Bij de transformatie naar E. colt K12 NM522 (zie verder) zijn deze lineaire stukken afgebroken, zodat alle transformanten een insertie moesten hebben. Mbv. SDS-PAGE, Western blotting L Inmiuno-blotting (zie verder) met monoklonalen tegen de nieuwe epitopen werden de transformanten geselecteerd op de juiste oriëntatie van de ingebouwde oligonucleotide.
1.4.4 Transformatie naar E. coli K12 NM522
Transformatie van plasmide DNA naar E. coli werd uit-gevoerd volgens [13]. Hierbij is gebruik gemaakt van E. coli K12 NM522, welke in Luria Broth medium (Trypton 10 g/1; gistextract 5 g/1; NaCl 10 g/1, pH 7.0) bij 37eC is opgegroeid. Na transformatie is de E. coli uit-geplaat op een vaste voedingsbodem, welke uit LB-medium met 1.5% (m/v) agar (Noble, Difco) bestaat. Voor de selectie op Ampicilline-resistentie werd aan het medium 100 pg/ml Ampicilline (Sigma) toegevoegd, waarna bij 30°C werd gegroeid om satelietvorming te voorkomen.
1.4.5 Plasmide-isolatie
Plasmide-isolatie uit E. coli is uitgevoerd volgens alkalische lysis methode [13]· 1.4.6 Agarose-gel-electroforese
Agarose-gel-electroforese is uitgevoerd volgens [13]* Hierbij is gebruik gemaakt van een 1% (m/v) agarose (Sigma No A-9539) voor een normale gel en een 1.2% (m/v) low melting agarose (Nusieve GTG, FMC Bioproducts) voor een preperatieve gel. Als buffer bij deze gelen is TBE-buffer (RIVM), lx geconcentreerd gebruikt: 0.089 M Tris, 0.089 M boorzuur, 0.002 M EDTA (pH 8.3)· De gebruikte molecuulmassa-marker is hier de lambda-Hindlll-digest (Boehringer Mannheim) met fragment-grootten van 125, 564, 2027, 2322, 4361, 6557, 9416, 23130 bp. Als loading-buffer (5x) diende een 30JÜ (m/v) sucrose-oplossing met 0.25% (m/v) broomfenolblauw.
De electroforese werd bij 10 V/cm uitgevoerd. Daarna werden deze met 0.5 pg/ml ethidiumbromide gekleurd en gefotografeerd.
lA.J Transformatie naar meningococcen
Een plaat met meningococcen werd o/n met 5% C02 en vochtig tissue bij 37°C gegroeid. De cellen van de volgegroeide plaat -zijn vervolgens in 10 ml Müller Hinton medium (RIVM), met 10 mM MgCl2 van 37°C geresuspendeerd. De bacteriesuspensie werd vervolgens 1:5 in Müller Hinton medium verdund, waarna 1 pg/ml plasmide DNA werd toe-gevoegd. Na 3 uur incubatie bij 37*C, werd 10* verdund in steriele PBS (met 0.01 M fosfaat, gebufferd fysiologisch zout pH 7.2; RIVM) en uitge-plaat op een Gonococcen agarplaat (* bijlage 3) · Na gedurende de nacht te zijn gekweekt werden de juiste transformanten geselecteerd mbv. kolonieblotting. Zie verder, bij kolonieblotting.
L.4.8 Kolonieblotting
De cellen van een volgegroeide meningococcen-plaat werden op een 0.^5 pm nitrocellulosemembraan (Schleicher & Schuell) overgebracht. Vervolgens werden de bacterie-cellen op de nitrocellulosemembraan in PBS (fosfaat gebufferd fys. zout, pH 7-2; RIVM) met 0.1% Tween 80 (Polyoxyethyleensorbiteanmonooleaat, Merck) gebracht en 1 uur bij 56°C geïnactiveerd. Daarna werden de overtollige cellen er afgeveegd en werd de blot behandeld als bij Immmoblotting (zie verder).
1.4.9 SDS-PAGE
SDS-PAGE is uitgevoerd volgens [8]. De gel bestond uit een 5# (m/v) acrylamide stacking gel en een 11% (m/v) acrylamide resolving gel. De gebruikte referentie-eiwitten (RIVM) hebben de grootten van 14, 20, 30, 43, 67 en 94 kD. Alle monsters voor de SDS-PAGE bestonden uit gehele cellen.
Voor de E. Coli-ceHen werden deze o/n bij 37*C opgegroeid in LB-medium met 100 pg/ml Ampicilline, zowel met als zonder 1 mM IPTG (=-Isopropyl-B-D-thiogalactopyranoside, Sigma). Van de suspensie is vervolgens 1.5 ml af gecentrifugeerd. De pellet is in 70 μΐ H20 opgenomen, waarna er 130 μΐ loading buffer (Tris/HCl 0.625 M pH 6.8; 10# SDS; 50# glycerol; 0.01# broomfenolblauw; B-mercapto-ethanol =2:4:4:2:1) aan toegevoegd is. Het mengsel is vervolgens 10 min. bij 95°C geïncubeerd.
Voor de meningococcen-cellen zijn cellen van een vol-gegroeide plaat (o/n, met 5# C02 en vochtige tissue bij 37eC) in 10 ml PBS ( met 0. 01 M fosfaat, gebufferd fysiologisch zout, pH 7*2; RIVM) opgelost, 30 min. bij 56 °C geïnactiveerd en af gecentrifugeerd (10 min. bij 4000 rpm). De pellet is opgenomen in 500 μΐ H20. Aan 35 tot 70 μΐ bacteriesuspensie werd 170 μΐ loadingbuffer en 0 tot 35 μΐ H20 toegevoegd, totale volume van 200 μΐ. Dit mengsel werd 10 min. bij 95°C verwarmd en op gel gezet.
De gebruikte electroforese-buffer bestond uit 50 mM Tris/HCl (pH 8.3); 38Ο mM glycine; 0.1% SDS. De electro-forese werd bij constante stroom van 40 mA uitgevoerd. Na electroforese werden de gelen: 1. óf 1 uur bij 56°C met Coomassie brilliant blue (0.1% (w/v) in 10# (v/v) azijnzuur en 30# (v/v) methanol) gekleurd en minimaal 1 uur bij -56eC ontkleurd in een 10# (v/v) azijnzuur-/5# (v/v) methanol-oplossing, 2. óf geblot op een 0.2 pm nitrocellulose-membraan (Schleicher & Schuell), zie verder bij Western blotting.
1.4.10 Western- en immmoblotting
De eiwitten welke op de SDS-PAG gescheiden zijn, werden overgebracht op een 0.2 pm nitrocellulose-membraan (Schleicher & Schuell) met behulp van de Ancos Semi Dry Electroblotter A, 1 uur lang bij 0.8 mA/cm2 gel, met als blotbuffer 25 mM TRIS/HC1 (pH8.3); 192 mM glycerine; 20# methanol; 0.0375# SDS. Vervolgens werd de blot gedurende een kwartier in PBS (fosfaat gebufferd fysiologisch zout, pH 7.2) met 0.1# Tween 80 (Merck) gespoeld. Daarna werden de blots een half uur in PBS met 0.1# Tween en 0.3# caseïne (hydrolysate, N-Z-AmineA, ICN Biochemicals) gespoeld. De blots zijn vervolgens minimaal 1 uur lang in PBS/0.1# Tween/0.3# caseïne met de monoklonale antilichamen tegen de aan te tonen epitoop geïncubeerd. Na incubatie werd 3x 10 min. gespoeld met PBS/0.1# Tween en vervolgens een half uur geïncubeerd met proteïne A-peroxidase-conjugaat (1:10.000 verdund) [9].
Na nogmaals 3X 10 min. met PBS/0.1# Tween en lx met H20 spoelen, werd het substraat/waterstofperoxidemengsel: 30 ml substraat A
(substr. A: fosfaat/citraatbuffer, 0.02 M Na2HP04 en 0.01 M citroenzuur 1:1, pH5.0) met 10 ml substraat B (substr. B: 24 mg Tetra-me-thyl-benzidine en 80 mg DONS (Dioctyl-sulfosuccinate) in 10 ml 96# ethanol) en 20 pl waterstofperoxide 30# (Merck) toegevoegd. Na enkele minuten werden er groen/blauwe bandjes zichtbaar op de blots waar antilichamen met de eiwitten binden. Daarna werden de blots 3x nagewassen met H20 en gefotografeérd.
1.4.11 Whole cell Elisa
Een plaat met meningococcen werd o/n met 5% C02 en vochtige tissue bij 37°C gegroeid. De cellen zijn vervolgens in 5 PBS (fosfaat gebufferd fys. zout, pH 7.2; RIVM) geresuspendeerd en 30 min, bij 56'C geïnactiveerd. Vervolgens is de suspensie met PBS tot een ODé20 van 0.1 verdund. Microtiterplaatjes (titertek PVC microassay plates U bottom) zijn met 100 μΐ suspensie /putje gevuld (bekleding). De suspensie is o/n bij 37°C of 2 dagen bij kamertemperatuur ingedroogd. De plaatjes zijn vlak voor gebruik 3X net PBS met 0.1% Tween (Merck) gewassen. Per putje is 100 μΐ oplossing met de mono-klonale-antilichamen-verdunning in PBS/0.1# Tween/0.5# Protifar (Nutricia) toegevoegd en is 1 uur bij 37°C geïncubeerd. De plaatjes zijn 3x gewassen met PBS/0.12 Tween. Vervolgens zijn de putjes met 100 μΐ proteïne A-peroxidase-conjugaat (RIVM, l:100000x verdund) of anti-IgM*-conjugaat (RIVM, l:2000x verdund in PBS/0.1% Tween/0.5# Protifar) suspensie/putje gevuld en is 1 uur bij 37°C geïncubeerd. Na 3X wassen met PBS/O.lJi Tween is er per putje 100 μΐ substraat C (substr. C: 1 ml Tetra-methyl-benzidine (6mg/ml alcohol 96%) en 22 μΐ H202 30% (Merck) in 60 ml 0.11 M NaAc buffer) toegevoegd, waarna 10 minuten is geïncubeerd. De reactie is gestopt door te blokkeren met 100 μΐ 2M H2S0/,/putje. De adsorpties zijn afgelezen met een ELI-SA-reader (Biokinetics Reader EL312e van Bio-Tek Instruments) bij 45Ο nm.
1.4.12 OMP-lsolatie
Een plaat met meningococcen werd o/n bij 37°C, met 5# C02 en vochtige tissue gegroeid. De cellen zijn vervolgens in 5 ml vloeibaar meningococcenmedium (RIVM) geresuspendeerd en verdeeld over twee flessen met 200 ml meningococcen-medium (RIVM). Deze flessen zijn o/n bij 37*0 geschud. De cellen zijn 30 min. bij 56*0 geïnactiveerd, waarna 10 min. bij 10.000 rpm is gecentrifugeerd (Beekman model J-21B, rotor JA-14). De bovenstaande vloeistof is afgegoten en aan iedere pellet is 10 ml 0.01 M Tris/HCl pH 8.0 toegevoegd. Na resus-penderen van de pellets zijn de suspensies samen in één 50 ml buis gevoegd. De buis is in ijswater gezet en is vervolgens 15 min. aan ultrasone trilling onderworpen (Branson Sonifier 250, stand 4, 50#) * Hierna is 30 min. bij 4000 rpm gecentrifugeerd (Heraeus Christ Mini-fuge GL met vaste rotor). De bovenstaande vloeistof werd 1 uur bij 30.000 rpm en 10eC gecentrifugeerd (Sorvall ARC-1 ultracentrifuge Oil Turbine Drive (OTD)-65, rotor 70 TA). Vervolgens is de pellet in 4 ml 1# sarcosyl in Tris/HCl pH 8.0 geresuspendeerd en 15 min. bij 5000 rpm (Heraeus minifuge) gecentrifugeerd. De bovenstaande vloeistof is 1 uur bij 30.000 rpm en 10°C gecentrifugeerd (Sorvall ultra-centrifuge, rotor T865.I). Na het afgieten van de bovenstaande vloeistof is de pellet in 2 ml 0.01 M Tris/HCl pH 8.0 opgelost. Vervolgens is de opbrengst gecontroleerd door het eiwitgehalte te bepalen (met het BCA* Protein Assay Reagent van Pierce, voorschrift volgens fabrikant). Hierna zijn de OMPs op een eiwitgehalte van 1 mg/ml in Tris-buffer gebracht en in 50 μΐ/buisje uit-gevuld. De zuiverheid is m.b.v. een 11# SDS-PAA-gel en een ELISA bepaald.
1·5· ReswUatgfl 1.5.1 Controle van pPH204 door transformatie naar meningococcen
Om te controleren of het plasmide pPH204 nog intact is gebleven, is dit plasmid naar meningococcenstam H44/76 B‘ getransformeerd. Hier werd de PI.2 epitoop van plasmide pPH204 uitgewisseld met de P1.16 epitoop van stam H44/76. Selectie op de juiste transformanten vond plaats m.b.v. kolonie- en immunoblotting, waarbij P1.2+ kolonies zijn geïsoleerd. De kolonies leverden na controle mbv. SDS-PAGE, Western-immunoblotting (zie fig. 5 en 6, 7. 8 en 9) de stam J072 op. Deze stam heeft in lus 1 de P1.7- en in lus 4 de P1.2-epitoop.
1.5.2 Insertie van PI.7 oligonucleotide in de Kpnl site van pPH204
De oligonucleotide voor de P1.7~epitoop is in de Kpnl site van pPH204 geplaatst, waarna het ontstane construct pJB007 naar E. coli K12 NM522 is getransformeerd. Van de ontstane kolonies is plasmide geïsoleerd. Het geïsoleerde plasmide-materiaal is gecontroleerd op het verdwijnen van de Kpnl restrictie site en de aanwezigheid van de nieuwe Spel restrictie site, zie fig.10. Bij alle gecontroleerde transformanten was de Kpnl restrictie site verdwenen èn was de nieuwe Spel restrictie site aanwezig. De transformanten zijn op SDS-PAGE gezet. Na Western-immunoblotting met monoklonale antistoffen gericht tegen de PI.7 epitoop, zijn 3 transformanten geselecteerd waar de oligonucleotide in de juiste oriëntatie zit, zie fig. 11 en 12, 13. 14 en 15.
1.5·3 Insertie van P1.16 oligonucleotide in de Kpnl site van pPH204
De oligonucleotide voor de Pl.l6-epitoop is in de Kpnl site van pPH204 geplaatst (construct pJB0l6) en vervolgens getransformeerd. Plasmiden van transformanten zijn gecontroleerd op de verdwenen Kpnl site en de nieuwe SnaBI site, zie fig.10. Met Westem-immunoblotting zijn 3 trans formanten geselecteerd met de Pl.l6-epitoop in de juiste oriëntatie, zie fig. 11 en 12, 13, 14 en 15.
1.5.4 Insertie van P1.16 oligonucleotide in de Spel site van pJB007
De oligonucleotide voor de PI. 16-epitoop is in de Spel site van pJB007 geplaatst (construct pJB7l6) en getransformeerd. Plasmiden van transformanten zijn gecontroleerd op de verdwenen Spel site, zie fig.10. Met westem-immunoblotting is 1 transformant geselecteerd met de oligonucleotide in de juiste oriëntatie, zie fig.11 en 12, 13, 14 en 15.
1.5.5 Insertie van P1.16 oligonucleotide in de Kpnl site van 2-2*SE
De oligonucleotide voor de Pl.l6-epitoop is in de Kpnl site van plasmide 2-2*SE geplaatst (construct ρΡΗΟΙβ) en getransformeerd. Plasmiden van transformanten zijn gecontroleerd op de juiste inbouw van de oligonucleotide (gegevens niet opgenomen) [17]· 1.5.6 Transformatie van de constructen naar meningococcen
Vervolgens zijn de vier constructen pJB007, pJB0l6, pJB7l6 en ρΡΗΟΙβ naar meningococcenstam A8-6* B' (PI.5.2) getransformeerd.·. Selectie op de juiste transformanten vond plaats mbv. kolonie- en immunoblot-ting, waarbij PI.7* en Pl.l6+ kolonies zijn geïsoleerd. Dit leverde na controle de stammen volgens tabel D op, zie ook fig. 5 en 6, 7» 8 en 9·

1.5.7 Whole cell ELISA
Met whole cell ELISA is de expressie van de ingebouwde epitopen in de nieuwe stammen bekeken en vergeleken met de ouderstammen A8-6l (PI.5,2) en HM/76 (PI.7.16), zie fig. 16 en tabel D.
1.6. Conclusie L discussie
Bij deze proefreeks zijn vier nieuwe meningococcen-stammen gemaakt met extra epitopen in lus 5 en 6. Stammen J007 en J016 bevatten de 1.7 resp. de P1.16 epitoop in lus 6, terwijl J716 zowel' de PI.7 als de PI.16 epitoop draagt. Stam P016 bevat uitsluitend de PI.16 epitoop in lus 5. De uitgevoerde whole cell ELISA laat zien dat de monoklonale antilichamen, gericht tegen de ingebouwde epitopen goed tot redelijk aan de hele cellen binden, wanneer vergeleken wordt met de ouderstammen H44/76 en A8-6*. Ook laat de ELISA zien dat de nieuwe klasse-l-OMPs nog geheel intact naar het buitenmembraan getransporteerd kunnen worden, anders vond er geen binding aan de monoklo-nalen antilichamen plaats. Als stam H44/76 vergeleken wordt met de vier stammen met extra epitopen in lussen 5 en 6, blijkt in alle gevallen de PI.7 epitoop ongeveer even goed te binden. Het maakt niet uit of de P1.7-epitoop zich in lussen 1, 5 of 6 bevindt.
1.7 Literatuur bij Voorbeeld I
[1] Barlow A.K., Heckels J.E. and Clarke I.N., The class 1 outer membrane protein of Neisseria meningitidis: gene sequence and structural immunological similarities to gonococcal porins, Molecular Biology 1989, 3(2), p. I3I-I39.
[2] Frasch E.F., Zollinger W.D. and Poolman J.T., Serotyping antigens of Neisseria meningitidis and a proposed scheme for designation of serotypes, Reviews of infectious diseases, vol. 7» no. 4, july-au-gust 1985, P. 504-510.
[3] Gotschlich E.C., Meningococcal meningitis. In: Bacterial Vaccins (Ed. Germanier, R.), Acedemice Press, Inc, 1984, ch.8, p. 237-255 [4] Maiden M.C.J., Suker J., McKenna A.J., Bygraves J.A. and Feavers I.M., Comparison of the class 1 outer membrane proteins of eight serological reference strains of Neisseria meningitis, Molecular Microbiology(1991).5(3), p. 727-736 [5] Klugman K.P., Gotschlich E.C. and Blake M.S., Sequence of the structural gene (rpmM) for the class 4 outer membrane protein of Neisseria menigitidis, homology of the protein to gonococcal protein III and Escherichia coli Omp A and construction of meningococcal strains that lack class 4 protein, Infection and Immunity, July 1989. P· 2066-2071.
[6] Ley P. van der, Heckels J.E., Virji M., Hoogerhout P. and Poolman J.T., Topology of outer membrane porins in pathogenic Neisseria, in press.
[7] Lifely M.R., Moreno C. and Lindon J.C., An intergrated molecular and immunological approach toward a meningococcal group B vaccine. Vaccine, vol. 5, march 1987. p. 11-26 [8] Lugtenberg B., Meijers J., Peters R., Hoek P. van de, Alphen L. van, Electrophoretic resolution of the major outer membrane protein of Escherichia coli K12 into four bands, FEBS letters, 1975* vol. 58, no.l, P.254-258.
[9] Nakane P.K., Kawaoi A., Perosidase-labeled antibody. A new method of conjugation, J. Histochem. and Cytochem.,1974, no. 22, p. 1084-1091.
[10] Nester E.W., Evans Roberts C., Pearsall N.N. and McCarthy B.J., Microbiology, 2nd edition, Eastbourne, Sussex, Holt Rinehart and Winston, 1978, p. 433. 443, 474, 540-543. 591.
[11] Peltola H., Safary A., Kaythy H., Karanko V. and André F.E., Evaluation of two tetravalent (ACYW-135) meingococcal vaccins in infants and small children: a clinical study comparing immunogenity of 0-acetyl-negative and 0- acetyl-positive grou C polysaccharides.
[12] Poolman J.T., Marie S. and Zanen H.C., Variability of low molecular weight, heat modifiable outer membrane proteins of Neisseria Meningitidis, Infection and Immunity, dec. 1980, p. 642-648 [13] Sambrook J.t Fritsch E.F. and Maniatis T., Molecular cloning: a laboratory manual, 2nd edition, 1989, Cold Spring Harbour Laboratory, Cold Spring Harbour, N.Y. p.1.25 -1.28, 1.82-1.84, 6.3-6.16, 6.I8-6.I9.
[14] Saukonen K., Leinonen M., Abdillahi H. and Poolman J.T., Comparative evaluation of potentional components for group B meningococcal vaccin by passive protection in the infant rat and in vitro bacterial assay, Vaccin, vol. 7. august 1989, Ρ· 325“328.
[15] Tomassen J., Vermeij P., Struyvé M., Benz R. and Poolman J.T., Isolation of Neisseria meningitidis mutants deficient in class 1 (PorA) and class 3 (PorB) outer membrane proteins, Infection and Immunity, may 1990, p. 1355-1359· [16] Tsai C.M., Frasch C.E. and Mocca L.F., Five structural classes of major outer membrane proteins in Neisseria meningitidis, J. Bacteri-01.,1981, no. 146, P.69-78.
[17] P. van der Ley persoonlijke informatie.
VOORBEELD II SAMENVATTING:
In dit voorbeeld wordt het creëren van een gemuteerd buitenmembraaneiwit beschreven, welk OMP een koppelingsmogelijkheid biedt voor het koppelen van het klasse-l-OMP aan het oligosaccharide deel van het lipopolysaccha-ride. Dit is gedaan door een cvsteïne in lus 6 van het klasse-l-OMP in te bouwen. Hiertoe is een oligonucleotide coderend voor een eysteïne ingebouwd in een bestaande restrictie site van het klasse-l-eiwitgen, welke oligo vervolgens is getransformeerd naar een kapselloze mutant van menin-gococcenstam H44/76, H44/76-B'. Na transformatie bleek cysteïne-inbouw geen invloed te hebben op epitoopexpressie of op produktie van het klas-se-l-OMP.
De keuze van lussen 5 en 6 werd bepaald door het feit dat deze geen belangrijke klasse-l-epitopen bevatten en door het in Voorbeeld I aangetoonde behoud van immuunwerking na het aanbrengen van mutaties in deze lussen. Controle van het gevolg van inbouw vond plaats aan de hand van behoud van epitoopexpressie van de ontstane transformanten.
2.1 MATERIAAL:
Het gebruikte plasmied:
Het gebruikte plasmied, pPH204, is afgeleid van het pTZ19R plasmied van Pharmacia
OVERZICHT VAN HET ONTSTAAN VAN DE GEBRUIKTE PLASMIEDEN

Gebruikte enzymen: -Kpnl (vol. act.: 10 U/μΙ); met incubatie-buffer L.
-Pstl (vol. act.: 10 U/μΙ); met incubatie-buffer H.
-T4 DNA Ligase (vol. act.: 1 U/μΙ); met T4-DNA-Ligase-buffer.
De samenstelling van de bijbehorende incubatie buffer,zie tabel E.

Zowel de enzymen als de buffers zijn afkomstig van B0EHR1NGER MANNHEIM. Gebruikte bacteriën:
Escherichia coli K12 NM522: deze stam bevat geen restrictie modifi-catie-systeem.
Meningococcen stam: H^4/76-B'. Dit is de kapselloze mutant van H44/76.
Gebruikte monoklonalen: zie tabel F

Alle gebruikte monoklonalen zijn afkomstig van het RIVM.
Gebruikte membranen: VOOR WESTERN BLOT: BIO RAD Trans Blot Transfer medium pure Nitrocellulose membrane Blotting Filter Paper, 0.45 micron.
Lot.No.: 4072/87020 Cat.No.: 162-0113 VOOR KOLONIE BLOT: SCHLEICHER & SCHUELL BA 85/22, 0,45 micron, diameter: 82 mm Ref.No.: 406 216
2.2 METHODES
2.2.1 HYBRIDISERING VAN 0LIG0NUCLE0TIDEN:
De complementaire niet-gefosforyleerde oligonucleotiden (OLIGO'S) (zie hieronder) werden samengevoegd in een concentratie van 1 pg/100 μΐ elk, en tot 95°C verwarmd. Na verwarming kon het mengsel langzaam afkoelen tot kamertemperatuur. Doordat het afkoelen langzaam gebeurt kan hybridisatie van de complementaire oligonucleotiden plaatsvinden [5]. Hierdoor ontstaan oligo’s met de juiste sticky ends, nl Kpnl sticky ends.
INFORMATIE C1-C2 OLIGONUCLEOTIDE:
Oligonucleotide coderend voor cysteine met Kpnl sticky ends:

Restrictie site voor Kpnl: 5'“GGTACC-3' 3'-CCATGG-5’
Restrictie site voor Pstl: S^CTGCAG^' 3’-GACGTC-5'
Het oligonucleotide is samengesteld uit twee enkelstrengs oligonucleo-tiden:
Oligo Cl : 5’-GGG CTG CAG CCT AAG TAC-3’
Oligo C2 : 5'“TTA GGC TGC AGC CCG TAC-3’ 2.2.2 LIGERING VAN GEHYBRIDISEERDE OLIGONUCLEOTIDEN IN PLASMIEDEN:
De plasmieden werden met het juiste enzym aan digestie onderworpen en daarna gescheiden van de ongeknipte plasmieden op een 1,2# low melting gel. Daarna werd de gel gekleurd met ethidiumbromide en werden de geknipte plasmieden uit de gel gesneden. Waarna d.m.v. fenolextractie en etha-nolprecipitatie het geknipte plasmied DNA weer uit de agarose gel werd geïsoleerd [8].
Van de geïsoleerde plasmieden werd de helft genomen om de oligo's in te ligeren. Het ligatiemengsel werd als volgt samengesteld:50μ1 geknipt plasmied-DNA, 20μ1 gehybridiseerd oligomengsel, 10μ1 ligatiebuffer, 4μ1 T^-ligase, Ι6μ1 dH20. Na overnacht (o/n) incuberen van het ligatiemengsel bij 16°C werd het mengsel geconcentreerd door ethanolprecipitatie. Vervolgens werden de ligatieprodukten gescheiden op een 1,2# low melting gel en werden de geligeerde produkten uitgesneden waarna deze uit de gel geïsoleerd werden door fenolextractie en ethanolprecipitatie.
2.2.3 UITSMELTEN OVERMAAT OLIGO:
Na isolatie van de ligatieprodukten uit de gel, werden deze tot 65°C verwarmd zodat de overmaat oligo werd uitgesmolten. Door wederom het mengsel langzaam af te laten koelen kon tussen de complementaire delen van de aan het plasmied geligeerde oligo weer hybridisatie optreden.
2.2 Λ VERWIJDEREN VAN ZELFSLUITERS:
De oplossingen werden vervolgens nageknipt met het oorspronkelijke restrictie enzym, zodat de zelfsluiters weer opengeknipt werden. Dit naknippen kon plaatsvinden omdat door inbouw van de oligo de oorspronkelijke restrictie site was veranderd in een andere restrictie site. Na knippen ontstonden dus lineaire plasmieden zonder oligo en circulaire plasmieden met een ingebouwd oligonucleotide. Omdat alleen circulair plasmied DNA door E.coli kan worden opgenomen, kunnen in principe alleen transformanten ontstaan die het oligo bevatten.
2.2.5 TRANSFORMATIE NAAR E.COLI:
Transformatie procedure zie: [8].
Na transformatie werden de transformanten op Luria Broth (L.B.) medium voedingsbodems uitgespateld waaraan 100pg/ml Ampicilline was toegevoegd. De platen werden o/n bebroed bij 37°C, waarna de transformanten werden reingestreken op L.B. voedingsbodems met ampicilline en wederom o/n wer- den bebroed waarna enkele kolonies werden overgeënt in vloeibaar L.B. medium met ampicilline, welk mengsel na o/n schuddend incuberen bij 37°C werd gebruikt voor plasmied isolatie (zie verder).
2.2.6 PLASMIED ISOLATIE:
Plasmied isolatie uit de transformanten werd uitgevoerd volgens de alkalische lysis methode[8].
2.2.7 DIGESTIE VAN DE RECOMBINANT-PLASMIEDEN:
Om te controleren of het oligonucleotide in de plasmieden aanwezig was, werd een digestie uitgevoerd met het restrictie enzym coderend voor de nieuwe restrictie site. Ook werd een digestie uitgevoerd met het restrictie enzym coderend voor de restrictie site welke verdwenen zou moeten zijn, zodat aan de hand van het restrictie patroon op een agarose gel kon worden gezien of de oude restrictie site verdwenen was en de nieuwe site ook echt aanwezig was. Van de positieve trans formanten werd de eiwitsa-menstelling gecontroleerd met Sodium-Dodecyl-Sulfaat-PolyAcrylAmide-Gel-Electroforese (S.D.S.-PAAGE).
2.2.8 S.D.S.-PAAGE:
De S.D.S.-PAAGE werd uitgevoerd volgens voorschrift[8]. Als scheidingsgel werd een 11# acrylamide-gel gebruikt, als concentrerings-gel een 5# acrylamide-gel .
Procedure bij E.coli.
Voor de electroforese werden de positieve transformanten o/n opgegroeid in vloeibaar LB-medium met (100pg/ml) ampicilline en zowel met als zonder (ImM) IPTG {=Isoprpyl-B-D-thiogalactopyranoside (sigma)}. Van de o/n cultuur werd l,5ml afgecentrifugeerd, waarna de pellet werd geresuspen-deerd in 4θμ1 dH20, hieraan werd 10μ1 loading buffer (250mM Tris-HCl pH6,8 , 10% S.D.S., 10% dithiotreitol, $0% glycerol, 0,05# bromophenol blue } toegevoegd waarna de suspensie 10 minuten bij 95°C werd verwarmd en op gel werd gezet.
Procedure bij meningococcen.
Voor de meningococcen werden de cellen, van een o/n in een vochtige atmosfeer met 5# C02 begroeide meningococcen plaat, verrijkt met isovitalex, opgenomen in 10ml PBS (0,01M fosfaat gebufferd fysiologisch zout, pH7, 2:RIVM). De verkregen suspensie werd vervolgens 30 min. bij 56°C verwarmd, zodat de meningococcen geïnactiveerd werden, hierna werd de suspensie afgecentrifugeerd (10 minuten bij 4000rpm). De pellet werd opgenomen in 4θμ1 dH20 en hieraan werd weer 10μ1 loading buffer toegevoegd waarna de suspensie 10 min. bij 95°C werd verwarmd en op gel werd gezet.
De gebruikte electroforese buffer had de volgende samenstelling: 50mM Tris/HCl (pH8,3). 380mM glycine, 0,1# S.D.S.. De electroforese werd bij 20mA per gel uitgevoerd.
Na de electroforese werd de gel: 1=> gekleurd met coomassie brilliant blue (1 uur bij 56°C), en daarna ontkleurd met 10# HAc/5 (v/v)# methanol (3 maal 30 min. bij 56°C).
2=> geblot op een 0,^5 micron nitrocellulosemembraan (Biorad).
2.2.9 WESTERN- OF IMMUNOBLOTTING:
Met behulp van een Ancos Semi Dry Electroblotter A werden de eiwitten van de S.D.S.-PAAgel overgeblot op een 0,^5 micron nitrocellulose membraan (Biorad). Het eiwittransport, van de gel naar de membraan,. vond plaats door gedurende 1 uur te blotten bij 0,8 mA/cm2 gel waarbij als blotbuffer 25mM Tris-HCl (pH8,3). 192mM glycine, 20# methanol, 0.03755* S.D.S. werd gebruikt.
Na het blotten werd de blot een kwartier gespoeld in PBS met 0,1# Tween 80 (polyoxyethylensorbitanmonooleate, Merck). Vervolgens werd de blot een half uur gespoeld met PBS waaraan 0,1# tween 80 en 0,3# caseïne hydrolysate (N-Z-Amine A, ICN Biochemicals) was toegevoegd, waarna gedurende een uur geïncubeerd werd met monoklonalen, opgelost in PBS/0,1# tween 80/0,3# caseïne, tegen de aan te tonen epitoop. Hierna werd de blot 3 maal 10 min. gespoeld met PBS/0,1# Tween 80 waarna een half uur geïncubeerd werd met proteïne A-peroxidase conjugaat,1:10.000 verdund met PBS/ 0,1# Tween 80/ 0,3# caseïne. Vervolgens werd wederom 3 maal 10 min. met PBS/ 0,1# Tween 80 en 1 maal met dH20 gespoeld waarna de waterstof-peroxide-sub-straat mix (20yl 30# waterstof-peroxide, Merck; 30ml substraat A:fos-faat/citraat buffer, 0,02 M Na2HP0/, en 0.01M citroenzuur (1:1) pH=5,0; 10ml substraat B:80mg Dioctyl-Sulfosuccinaat{DONS}, 24mg Tetra-Methyl-Benzidine{TMB} in 10 ml 96# ethanol) werd toegevoegd.
Na enkele minuten werden blauwe bandjes zichtbaar op de plaats waar de monoklonale antistoffen aan de bijbehorende eiwitten hadden gebonden. Daarna werden de blots nog 3 maal gespoeld met water waarna ze gefotografeerd werden of in het donker werden bewaard totdat ze gefotografeerd werden.
2.2.10 TRANSFORMATIE NAAR MENINGOCOCCEN:
Meningococcen werden o/n gegroeid op GC agar (Difco), verrijkt met iso-vitalex, bij 37°C in een vochtige atmosfeer met 5% C02(3). De cellen van de volgegroeide plaat werden geresuspendeerd in 10ml Müller Hinton medium (RIVM), met lOmM MgCl2, van 37°C. Vervolgens werd de suspensie 1:5 verdund in Müller Hinton medium met lOmM MgCl2 en werd lpg/ml plasmied DNA toegevoegd. Hierna werd 3 uur bij 37°C geïncubeerd waarna de bacterie suspensie 10^ maal met steriele PBS werd verdund en op een GC plaat uitgespa-teld. Na o/n gegroeid te zijn werden de juiste transformanten geselecteerd m.b.v. een kolonie blot.
2.2.11 KOLONIE BLOT:
De kolonies van een o/n bij 37°C in een vochtige atmosfeer met 3% C02 begroeide plaat, werden overgeblot op een 0,45 micron nitocellulose membraan (Schleicher L Schuell). Vervolgens werd de blot 30 min. in PBS/0,1# Tween 80 verhit bij 56°C zodat de meningococcen geïnactiveerd werden. Na verhitten werd de overmaat bacterïen van de blot af geveegd. Zie voor de verdere procedure Western- of Immunoblot techniek.
2.2.12 OMP ISOLATIE:
Een plaat met meningococcen werd o/n bij 37°C. in een vochtige atmosfeer met 5% C02. begroeid waarna de cellen in 5ml meningococcen medium gere-suspendeerd werden. Met 2,5nil hiervan werd 200ml meningococcen medium beënt. Deze 200ml werd o/n geschud bij 37°C, waarna de meningococcen geïnactiveerd werden door ze een half uur bij 56°C te incuberen. Vervolgens werden de meningococcen gepelleteerd door het medium 10 min. af te draai en bij lO.OOOrpm (centrifuge: centrikon T-324,rotor A6.9)· Het supernatant werd af gegoten en de pellet werd geresuspendeerd in 10ml 0.01M Tris/HCl pH 8.0. Vervolgens werd deze oplossing 15 min. ultrasoon getrild (Branson Sonifier 250, stand 4, 50%), waarbij de oplossing in een ijsbad stond. Vervolgens werd de gesonificeerde oplossing 10 min. bij 5000 rpm. afgedraaid (centrifuge: centrikon T-324, rotor A8.20). Het supernatant werd vervolgens een uur bij 20.000 rpm. en 10°C afgedraaid (centrifuge: centrikon T-324, rotor A8.20). De zo ontstane pellet werd geresuspendeerd in 4ml 1% sarcosyl in 0.01M Tris/HCl pH 8.0, waarna 5 min. bij 5000 rpm. (centrikon T-324 centrifuge, rotor A8.20) gecentrifugeerd werd. Het supernatant werd vervolgens een uur bij 20.000 rpm. en 10°C afgedraaid (centrikon T-324 centrifuge, rotor A8.20). De zo ontstane pellet werd geresuspendeerd in 1ml 0.01M Tris/HCl pH 8.0. De opbrengst werd bepaald met de Microassay Procedure (BI0RAD). De zuiverheid van de OMPs werd gecontroleerd met S.D.S.-PAAGE.
2.2.13 TRANSFORMANTCONTROLE m.b.v. POLYMERASE CHAIN REACTION:
Dagl:
De te PCRen meningococcen werden afgeënt op een G.C.plaat, verrijkt met isovitalex, en o/n in een vochtige atmosfeer met 5% C02 bij 37°C bebroed.
Dae2:
Van elk van de te PCRen stammen werd een celsuspensie gemaakt, door een "plukje” bacteriën te resuspenderen in 1 mL steriel aqua dest, en deze suspensie vervolgens 10 min. bij 95°C te verhitten. De ontstane oplossing werd vervolgens kort afgedraaid en daarna op ijs bewaard. Het reactie-mengsel was als volgt samengesteld: 10pL buffer (500mM KCl.lOOmM Tris/HCl pH8,3,15mM MgCl2, 0,ljï(w/v) gelatine) 200μΜ van elk van de dNTP's lOOng van elk van de primers 25pL celsuspensie aangevuld tot 100pL met steriel aqua dest.
- 100pL minerale olie.
De PCR condities waren als volgt: zie tabel G Tabel G: PCR condities

Na de 30e cyclus werden de monsters nog 8 minuten op r]2°C gehouden om al het in de monsters aanwezige DNA dubbelstrengs te maken.
Na het PCRen werden op de monsters een fenolextraktie en een ethanolpre-cipitatie uitgevoerd om het geproduceerde DNA zuiver in handen te krijgen.
Van het gezuiverde DNA werd 10# op een 1,0# agarose gel geëlèctroforeerd om het resultaat te bekijken. Hier uit moet blijken of het ingebouwde nucleotide in het DNA aanwezig is.
2.2 RESULTATEN: 2.2.1 TRANSFORMATIE VAN DE RECOMBINANT PLASMIEDEN NAAR E.coli:
Het gehybridiseerde oligonucleotide is in met Kpnl geknipt pPH204 geli-geerd waarna het geheel is getransformeerd naar E.coli K12 NM522. Uit de ontstane transformanten zijn de plasmieden geïsoleerd. De geïsoleerde plasmieden zijn met Kpnl en Pstl geknipt en vervolgens op een 0,8# Agarose gel geëlectroforeerd, zie fig 16.

Uit het ontstaan van een fragment van ongeveer 600 baseparen bij de Pstl digesten en het verdwijnen van de Kpnl site blijkt dat de oligo in pPH204 is ingebouwd.
De transformanten zijn vervolgens o/n bij 37°C opgekweekt in vloeibaar L.B. medium met ImM IPTG en 100pg/ml ampiciline. De bacteriële eiwitten zijn vervolgens m.b.v. S.D.S.-PAGE gescheiden waarna met immunoblotting is gekeken of de oligo in de juiste oriëntatie was ingebouwd. Bij de blotting is gekeken naar de grootte van het klasse-l-eiwit door met het monoklonaal MN16C13F4 de PI.2 epitoop aan te tonen, zie fig 18 en 19.
De transformanten in de lanen n° 1,3.5.7.9.11 geven een eiwitband te zien die minder ver is gelopen dan die van de trans formanten in de overige lanen. Dit geeft aan dat het geproduceerde eiwit groter is dan bij de transformanten in de lanen n° 2,4,6,8,10. De transformanten in de oneven lanen hebben het oligonucleotide in de juiste oriëntering en die van de even lanen in de onjuiste oriëntering, waardoor een stopcodon ontstaat, hetgeen een kleiner eiwit tot gevolg heeft.
2.3. TRANSFORMATIE NAAR H44/76 B':
De plasmieden van de positieve transformanten zijn vervolgens getransformeerd naar H44/76 B‘, een kapselloze mutant van H44/76. De transformanten zijn geselecteerd met kolonie blotting op de PI.2 epitoop met de monoklonaal MN16C13F4. Van de zuivere transformanten zijn de buitenmembraan eiwitten geïsoleerd. De zuiverheid is gecontroleerd d.m.v. S.D.S.-PAAGE en immunoblotting, zie fig 20 en 21. Er is ook gekeken of de verhouding tussen het klasse-l-eiwit en de andere eiwitten bij de transformanten was veranderd was. Dit bleek niet het geval te zijn (resultaten niet verwerkt) .
Vervolgens is bekeken of de P1.16 epitoop uit de transformanten verdwenen is, als controle zijn de monoklonalen tegen de PI.2 en de PI.7 epitoop meegenomen en de stam H44/76 B' is als controle stam meegenomen fig 22.
Uit de figuur blijkt dat de monoklonaal MN5C11G, gericht tegen de PI.I6 epitoop, alleen met de ouderstam reageert. De monoklonaal MN14C11.6, gericht tegen de PI.7 epitoop reageert met zowel de ouderstam als de transformanten. MN16C13F4, gericht tegen de PI.2 epitoop reageert alleen met de transformanten.
Claims (39)
- 2.4 CONCLUSIE: Het succesvol plaatsen van een cysteine residue in het klasse 1 eiwit van H44/76-B' blijkt uit de volgende resultaten: 1. bij inbouw van het oligonucleotide is de Kpnl site verdwenen en is een nieuwe Pstl site ontstaan. 2. na transformatie naar H44/76-B' is deze veranderd van PI.7.16 naar PI.7,2 hetgeen aangeeft dat het gen coderend voor het klasse-l-eiwit dat in plasmied pPH204 zat, in welk gen de oligo was ingebouwd door de meningococ is opgenomen. Cysteïne inbouw lijkt geen invloed te hebben op de produktie van het klasse 1 eiwit gezien de hoeveelheid die door de meningococ wordt geproduceerd. De epitoopexpressie door de bacterie lijkt ook normaal te blijven. 2.5 LITERATUURLIJST bij VOORBEELD 2:1. Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmem-braaneiwitten (OMP's) hebben, met het kenmerk, dat een polypeptide uit de groep een in het membraan verankerd OMP of OMP-fragment is met een mutatie in een van de oppervlaktelussen.
- 2. Buitenmembraanvesikel dat volgens conclusie 1, net het kenmerk, dat een polypeptide uit de groep een in het membraan verankerd OMP of OMP-fragment van Klasse-l-OMP is.
- 3. Buitenmembraanvesikel volgens conclusie 1 of 2, met het kenmerk, dat een polypeptide uit de groep een in het membraan verankerd OMP of OMP-fragment is met een mutatie in een van de oppervlaktelussen 2, 3» 5* 6, 7 of 8 is. k. Buitenmembraanvesikel volgens een van de voorgaande conclusies met het kenmerk, dat een polypeptide uit de groep een in het membraan verankerd OMP of OMP-fragment is met een mutatie in een van de lussen 6 of 7-
- 5. Buitenmembraanvesikel volgens een van de voorgaande conclusies met het kenmerk, dat het polypeptide recombinant-OMP of een fragment daarvan met ten minste de immuunwerking van OMP is.
- 6. Buitenmembraanvesikel volgens een van de voorgaande conclusies met het kenmerk, dat de mutatie de aanwezigheid van ten minste een aminozuur met een specifiek reactieve zijketen omvat. 7« Buitenmembraanvesikel volgens een van de voorgaande conclusies met het kenmerk, dat de mutatie ten minste de aanwezigheid van cysteine in het polypeptide omvat.
- 8. Buitenmembraanvesikel volgens een van de vooorgaande conclusies met het kenmerk, dat het polypeptide voor ten minste twee HLA-fenotypes herkenbaar is.
- 9. Buitenmembraanvesikel volgens een van de voorgaande conclusies met het kenmerk, dat het voorzien is van saccharide-peptide-conjugaten die als peptidedeel het polypeptide omvatten en op de plaats van de mutatie in het polypeptide zijn geconjugeerd met het saccharidedeel, welk saccharidedeel een van een gramnegatieve bacterie afgeleid B-cel activerend deel met ten minste een epitoop omvat.
- 10. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat het saccharidedeel ten minste een gesynthetiseerd B-cel activerend deel met ten minste één epitoop omvat.
- 11. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, eet het kenmerk, dat het saccharidedeel ten minste een B-cel activerend deel met ten minste één epitoop omvat, welk B-cel activerend deel van het oligosaccharidedeel (inner core) van een LPS van Meningococcus is afgeleid.
- 12. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat het saccharidedeel via een spacer is gekoppeld aan het peptidedeel.
- 13. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat het saccharidedeel met het peptidedeel is geconjugeerd met behoud van de vrije aminogroepen van in het B-cel activerende deel van het saccharidedeel aanwezige fosforethanolamine (PEA)-groepen.
- 14. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat bij het saccharidedeel bij het B-cel activerende deel met ten minste één epitoop voor de conjugatie ten minste één PEA-groep is aangebracht.
- 15. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat het saccharidedeel een B-cel activerend deel omvat dat is afgeleid van een oligosaccharidedeel van een LPS van Meningococcus met immunotype L3.
- 16. Buitenmembraanvesikel (OMV) volgens een van de voorgaande conclusies, met het kenmerk, dat het saccharidedeel een B-cel activerend deel omvat dat is afgeleid van

- 17. Buitenmembraanvesikel volgens een van de conclusies 1-14, met het kenmerk, dat het B-cel activerende deel van het saccharidedeel is afgeleid van een oligosaccharidedeel van een LPS van Meningococcus met immunotype L2.
- 18. Buitenmembraanvesikel (OMV) volgens conclusie 17, met het kenmerk, dat het B-cel activerende deel van het saccharidedeel een afgeleide is van de vertakte oligosaccharide
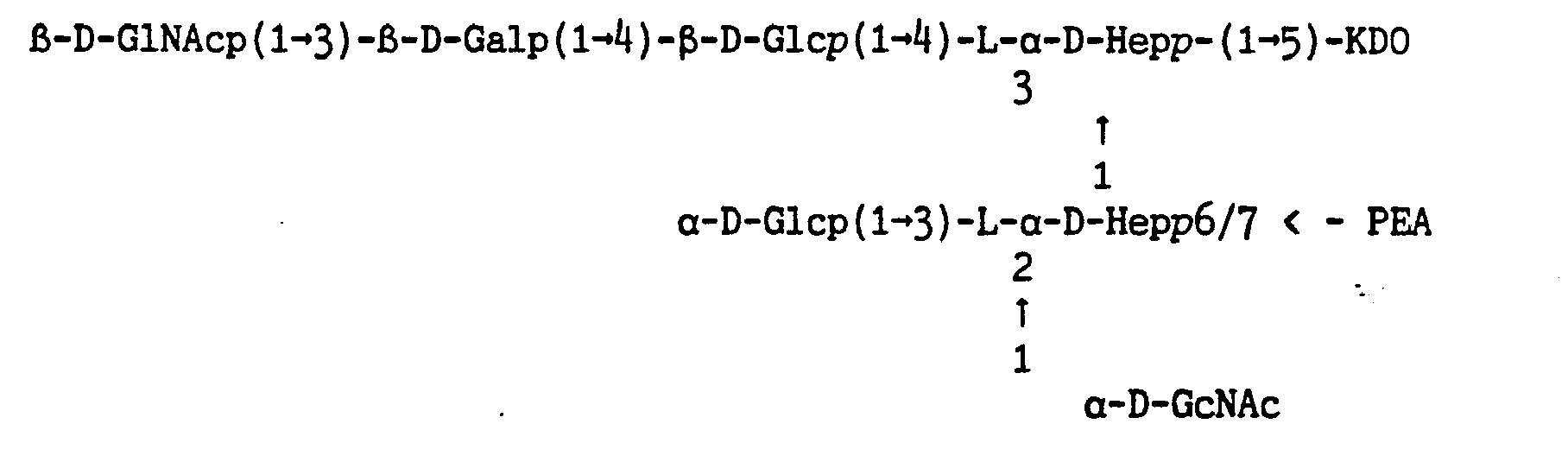
- 19· Werkwijze voor het bereiden van een buitenmembraanvesikel volgens een van de voorgaande conclusies, «et het kenmerk, dat men ten minste 1. een nucleotidesequentie coderend voor het polypeptide tot expressie brengt in een bacterie; 2. de bacterie kweekt onder op zich bekende omstandigheden waarbij deze buitenmembraanvesikels produceert en de aldus ontstane buitenmem-braanvesikels met polypeptide eventueel isoleert.
- 20. Werkwijze voor het bereiden van een buitenmembraanvesikel volgens conclusie 19, met het kenmerk, dat men 3. de in stap 2 ontstane buitenmembraanvesikels voorziet van sacchari-de-peptide-conjugaten door het polypeptide te koppelen aan het gewenste saccharidedeel.
- 21. Werkwijze volgens conclusie 20, waarbij men een via recombinant DNA-technieken verkregen saccharidedeel toepast, welk deel afkomstig is uit een gemutageniseerde of geselecteerde productiestam zodat ten minste de terminale galactose niet wordt aangemaakt.
- 22. Werkwijze volgens een van de conclusies 20 of 21, waarbij men een langs synthetische weg verkregen saccharidedeel toepast.
- 23· Werkwijze volgens een van de conclusies 20-22, waarbij men de saccharide van het saccharidedeel via een spacer die aan de uiteinden reactieve groepen zoals NH2, C00H, SH en C0CH2Br bevat, koppelt aan het peptidedeel van het OMV dat uit stap 2) resulteert.
- 24. Werkwijze volgens een van de conclusies 20-23, waarbij men het saccharide modificeert met een met sulfhydryl reactieve groep.
- 25. Werkwijze volgens conclusie 24, waarbij de met sulfhydryl reactieve groep broomacetyl is.
- 26. Werkwijze volgens conclusie 25, waarbij men de broomacetylgroep heeft verkregen met N-succinimidyl-broomacetaat.
- 27. Nucleotidesequentie die voor recombinant-OMP of voor een recom-binant-OMP-fragment codeert met het kenmerk, het recombinant -0MP of -fragment zich onderscheidt van niet recombinant-OMP of niet-recombinant- OMP-fragment door ten minste een mutatie in een van de oppervlaktelussen van het OMP.
- 28. Nucleotidesequentie volgens conclusie 27 met het kenmerk, dat het OMP een Klasse-l-OMP is
- 29· Nucleotidesequentie volgens conclusie 27 of 28 met het kenmerk, dat het recombinant -OMP of -fragment zich onderscheidt van niet recombi-nant-OMP of niet-recombinant-OMP-fragment door ten minste een mutatie in een van de oppervlaktelussen 2, 3. 5* 6, 7 of 8 van het OMP.
- 30. Nucleotidesequentie volgens een van de conclusies 27-29 met het kenmerk, dat het recombinant -OMP of -fragment zich onderscheidt van niet recombinant-OMP of niet-recombinant-OMP-fragment door ten minste een mutatie in een van de oppervlaktelussen 6 of 7 van het OMP.
- 31. Nucleotidesequentie volgens een van de conclusies 27”30 met het kenmerk, dat de mutatie ten minste resulteert in een codon dat voor een aminozuur met een specifiek reactieve zijketen codeert.
- 32. Nucleotidesequentie volgens een van de conclusies 27”31 met het kenmerk, dat de mutatie ten minste resulteert in een codon dat voor cysteine codeert.
- 33. Expressievector met het kenmerk, dat deze een nucleotidesequentie volgens een van de conclusies 27-32 omvat.
- 34. Microorganisme met het kenmerk, dat het een nucleotidesequentie volgens een van de conclusies 27-32 of een expressievector volgens conclusie 33 omvat.
- 35. Polypeptide dat van OMP of een OMP-fragment verkrijgbaar is en ten minste de immuunwerking van aan een membraan gebonden OMP heeft met het kenmerk, dat het polypeptide in vergelijking tot het OMP of het OMP-fragment een mutatie in een van de oppervlakte lussen omvat.
- 36. Polypeptide volgens conclusie 35 met het kenmerk, dat het OMP een klasse-l-OMP is.
- 37. Polypeptide volgens conclusie 35 of 36 met het kenmerk, dat het polypeptide in vergelijking tot het OMP of het OMP-fragment een mutatie in een van de oppervlakte lussen 2, 3· 5· 6, 7 of 8 omvat.
- 38. Polypeptide volgens een van de conclusies 35“37 met het kenmerk, dat het polypeptide in vergelijking tot het OMP of het OMP-fragment een mutatie in een van de oppervlakte lussen 6 of 7 omvat.
- 39. Polypeptide volgens een van de conclusies 35“3δ met het kenmerk, dat de mutatie ten minste resulteert in aanwezigheid van een aminozuur met een specifiek reactieve zijketen in een van de oppervlakt lussen. 4θ. Polypeptide volgens een van de conclusies 35~39 wet het kenmerk, dat de mutatie ten minste resulteert in aanwezigheid van cysteine.
- 41. Saccharide-peptide-conjugaat dat ten minste een epitoop van een OMP omvat met het kenmerk, dat het een polypeptide volgens een van de conclusies 35“40 als peptidedeel omvat.
- 42. Vaccin gekenmerkt door een werkzaam gehalte aan buitenmembraan-vesikel volgens een van de conclusies 1-18, een volgens de werkwijze van een van de conclusies 19“26 verkregen buitenmembraanvesikel of een polypeptide volgens een van de conclusies 35~40 of een saccharide-peptide-conjugaat volgens conclusie 4l.
Priority Applications (14)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| NL9201716A NL9201716A (nl) | 1992-10-02 | 1992-10-02 | Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. |
| EP93921120A EP0680512B1 (en) | 1992-10-02 | 1993-07-30 | Immunogenic meningococcal lps and outer membrane vesicles and vaccine therefrom |
| US08/411,727 US5705161A (en) | 1992-10-02 | 1993-07-30 | Immunogenic meningococcal LPS and other membrane vesicles and vaccine therefrom |
| AT93921120T ATE364702T1 (de) | 1992-10-02 | 1993-07-30 | Immunogene meningococcale lps und aussenmembranvesikel und impstoff daraus |
| DK93921120T DK0680512T3 (da) | 1992-10-02 | 1993-07-30 | Immunogent meningokok LPS og ydermembranvesikler og vaccine deraf |
| ES93921120T ES2289739T3 (es) | 1992-10-02 | 1993-07-30 | Liposacarido meningococico inmunogenico y vesiculas de membrana externa y vacunas de los mismos. |
| DE69334147T DE69334147T2 (de) | 1992-10-02 | 1993-07-30 | Immunogene meningococcale lps und aussenmembranvesikel und impstoff daraus |
| JP6508917A JPH08501940A (ja) | 1992-10-02 | 1993-07-30 | 免疫原性髄膜炎菌のlpsと外膜小胞とそれからのワクチン |
| FI951535A FI951535L (fi) | 1992-10-02 | 1993-07-30 | Immunogeeninen meningokokki -LPS ja siitä saadut ulkomembraanirakkulat ja rokote |
| PCT/NL1993/000163 WO1994008021A1 (en) | 1992-10-02 | 1993-07-30 | Immunogenic meningococcal lps and outer membrane vesicles and vaccine therefrom |
| PT93921120T PT680512E (pt) | 1992-10-02 | 1993-07-30 | Uma molécula provida de imunidade activadora das células e derivada de um lipopolissacarídeo meningocócico, métodos para a sua preparação, uma vesícula de membrana externa provida com grupo de polipepetídeos |
| AU48351/93A AU684720B2 (en) | 1992-10-02 | 1993-07-30 | Immunogenic meningococcal LPS and outer membrane vesicles and vaccine therefrom |
| CA002146145A CA2146145C (en) | 1992-10-02 | 1993-07-30 | Immunogenic meningococcal lps and outer membrane vesicles and vaccine therefrom |
| NO19951181A NO321651B1 (no) | 1992-10-02 | 1995-03-28 | Immunogent meningokokk-lipopoly-sakkarid, ytter-membranvesikler, samt vaksine fremstilt derav. |
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| NL9201716 | 1992-10-02 | ||
| NL9201716A NL9201716A (nl) | 1992-10-02 | 1992-10-02 | Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| NL9201716A true NL9201716A (nl) | 1994-05-02 |
Family
ID=19861338
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| NL9201716A NL9201716A (nl) | 1992-10-02 | 1992-10-02 | Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. |
Country Status (14)
| Country | Link |
|---|---|
| US (1) | US5705161A (nl) |
| EP (1) | EP0680512B1 (nl) |
| JP (1) | JPH08501940A (nl) |
| AT (1) | ATE364702T1 (nl) |
| AU (1) | AU684720B2 (nl) |
| CA (1) | CA2146145C (nl) |
| DE (1) | DE69334147T2 (nl) |
| DK (1) | DK0680512T3 (nl) |
| ES (1) | ES2289739T3 (nl) |
| FI (1) | FI951535L (nl) |
| NL (1) | NL9201716A (nl) |
| NO (1) | NO321651B1 (nl) |
| PT (1) | PT680512E (nl) |
| WO (1) | WO1994008021A1 (nl) |
Families Citing this family (43)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7118757B1 (en) * | 1988-12-19 | 2006-10-10 | Wyeth Holdings Corporation | Meningococcal class 1 outer-membrane protein vaccine |
| AU707131B2 (en) * | 1995-08-04 | 1999-07-01 | University Of Guelph | Novel vaccines and pharmaceutical compositions using membrane vesicles of microorganisms, and methods for preparing same |
| US20020028215A1 (en) | 1999-08-09 | 2002-03-07 | Jagath L. Kadurugamuwa | Novel vaccines and pharmaceutical compositions using membrane vesicles of microorganisms, and methods for preparing same |
| US6872398B2 (en) * | 1995-12-22 | 2005-03-29 | The United States Of America As Represented By The Secretary Of The Army | Conjugate vaccine against gram-negative bacterial infections |
| US6645503B1 (en) * | 1998-03-10 | 2003-11-11 | Wyeth Holdings Corporation | Antigenic conjugates of conserved lipopolysaccharides of gram negative bacteria |
| GB9823978D0 (en) * | 1998-11-02 | 1998-12-30 | Microbiological Res Authority | Multicomponent meningococcal vaccine |
| US10967045B2 (en) | 1998-11-02 | 2021-04-06 | Secretary of State for Health and Social Care | Multicomponent meningococcal vaccine |
| US6514948B1 (en) * | 1999-07-02 | 2003-02-04 | The Regents Of The University Of California | Method for enhancing an immune response |
| US6531131B1 (en) | 1999-08-10 | 2003-03-11 | The United States Of America As Represented By The Department Of Health And Human Services | Conjugate vaccine for Neisseria meningitidis |
| US7585510B1 (en) | 1999-09-30 | 2009-09-08 | Isis Innovation Limited | Vaccine |
| US6936261B2 (en) * | 2000-07-27 | 2005-08-30 | Children's Hospital & Research Center At Oakland | Vaccines for broad spectrum protection against diseases caused by Neisseria meningitidis |
| EP1495039A4 (en) * | 2002-03-20 | 2006-07-26 | Univ Emory | MUTANTS OF NEISSERIA, LIPOOLIGOSACCHARIDES AND IMMUNOGENIC COMPOSITIONS |
| EP1524991A1 (en) * | 2002-08-02 | 2005-04-27 | GlaxoSmithKline Biologicals S.A. | Vaccine composition comprising transferrin binding protein and hsf from gram negative bacteria |
| RU2317106C2 (ru) * | 2002-08-02 | 2008-02-20 | ГлаксоСмитКлайн Байолоджикалз с.а. | Нейссериальные вакцинные композиции, содержащие комбинацию антигенов |
| GB0220194D0 (en) | 2002-08-30 | 2002-10-09 | Chiron Spa | Improved vesicles |
| HUE031886T2 (en) | 2002-10-11 | 2017-08-28 | Glaxosmithkline Biologicals Sa | Polypeptide vaccines for extensive protection against hypervirulent meningococcal lines |
| EP1562982B1 (en) | 2002-11-15 | 2010-05-05 | Novartis Vaccines and Diagnostics S.r.l. | Unexpected surface proteins in neisseria meningitidis |
| GB0227346D0 (en) | 2002-11-22 | 2002-12-31 | Chiron Spa | 741 |
| ES2461350T3 (es) | 2003-01-30 | 2014-05-19 | Novartis Ag | Vacunas inyectables contra múltiples serogrupos de meningococos |
| GB0323103D0 (en) | 2003-10-02 | 2003-11-05 | Chiron Srl | De-acetylated saccharides |
| GB0408977D0 (en) | 2004-04-22 | 2004-05-26 | Chiron Srl | Immunising against meningococcal serogroup Y using proteins |
| US7537766B2 (en) * | 2004-08-30 | 2009-05-26 | Wyeth | Neisseria meningitidis inner core lipo-oligosaccharide epitopes, multivalent conjugates thereof and immunogenic compositions thereof |
| GB0424092D0 (en) | 2004-10-29 | 2004-12-01 | Chiron Srl | Immunogenic bacterial vesicles with outer membrane proteins |
| ES2616294T3 (es) | 2005-01-27 | 2017-06-12 | Children's Hospital & Research Center At Oakland | Vacunas de vesículas basadas en GNA1870 para protección de amplio espectro contra enfermedades provocadas por Neisseria meningitidis |
| GB0524066D0 (en) | 2005-11-25 | 2006-01-04 | Chiron Srl | 741 ii |
| DE602007003596D1 (de) | 2006-06-12 | 2010-01-14 | Glaxosmithkline Biolog Sa | Impfstoff |
| CN102356089B (zh) | 2008-02-21 | 2014-02-19 | 诺华股份有限公司 | 脑膜炎球菌fHBP多肽 |
| JP5723768B2 (ja) * | 2008-05-30 | 2015-05-27 | ザ ユナイテッド ステイツ オブ アメリカ, アズ レプレゼンテッド バイ ザ セクレタリー オブ ジ アーミー, オン ビハーフ オブ ウォルター リード アーミー インスティチュート オブ リサーチ | 髄膜炎菌性多価未変性外膜小胞ワクチン、その作製方法およびその使用 |
| EP3017826A1 (en) | 2009-03-24 | 2016-05-11 | Novartis AG | Combinations of meningococcal factor h binding protein and pneumococcal saccharide conjugates |
| HUE049695T2 (hu) | 2009-03-24 | 2020-10-28 | Glaxosmithkline Biologicals Sa | Adjuváló H meningococcus faktort kötõ fehérje |
| DK2430151T3 (da) * | 2009-05-14 | 2014-09-15 | Sanofi Pasteur | Meningokokvaccine på basis af lipooligosaccharid (los) fra modificerede stammer af neisseria meningitidis med immuntype l6 |
| JP5536765B2 (ja) * | 2009-05-20 | 2014-07-02 | 国立大学法人鳥取大学 | 部分糖鎖エピトープを用いた、病原性ナイセリア属細菌感染の検出方法およびそれら細菌に対するワクチン |
| CN102740882A (zh) | 2009-08-27 | 2012-10-17 | 诺华有限公司 | 含有铝、寡核苷酸和聚阳离子的佐剂 |
| US20110076300A1 (en) | 2009-08-27 | 2011-03-31 | Mariagrazia Pizza | Hybrid Polypeptides Including Meningococcal fHBP Sequences |
| CN102724988B (zh) | 2009-09-30 | 2014-09-10 | 诺华股份有限公司 | 脑膜炎球菌fHBP多肽的表达 |
| CA2779816A1 (en) | 2009-10-27 | 2011-05-05 | Novartis Ag | Modified meningococcal fhbp polypeptides |
| CN102802662A (zh) | 2010-03-18 | 2012-11-28 | 诺华有限公司 | 用于脑膜炎球菌血清组b的含佐剂疫苗 |
| CA2803239A1 (en) | 2010-06-25 | 2011-12-29 | Novartis Ag | Combinations of meningococcal factor h binding proteins |
| EP2797624A1 (en) | 2011-12-29 | 2014-11-05 | Novartis AG | Adjuvanted combinations of meningococcal factor h binding proteins |
| JP6786394B2 (ja) | 2014-02-28 | 2020-11-18 | グラクソスミスクライン バイオロジカルズ ソシエテ アノニム | 改変髄膜炎菌fHbpポリペプチド |
| GB201712824D0 (en) | 2017-08-10 | 2017-09-27 | Glaxosmithkline Biologicals Sa | Multi-functionalized nOMV Conjugates |
| EP3836957A1 (en) * | 2018-08-14 | 2021-06-23 | Evercyte GmbH | Target-specific extracellular vesicles |
| WO2022152939A1 (en) | 2021-01-18 | 2022-07-21 | Conserv Bioscience Limited | Coronavirus immunogenic compositions, methods and uses thereof |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP3436756B2 (ja) * | 1988-12-19 | 2003-08-18 | アメリカン・サイアナミド・カンパニー | 髄膜炎菌のクラスiの外膜タンパク質のワクチン |
| NL9101359A (nl) * | 1991-08-07 | 1993-03-01 | Nederlanden Staat | Nieuw saccharide-peptide-conjugaat, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk saccharide-peptide-conjugaat omvat. |
-
1992
- 1992-10-02 NL NL9201716A patent/NL9201716A/nl not_active Application Discontinuation
-
1993
- 1993-07-30 DE DE69334147T patent/DE69334147T2/de not_active Expired - Lifetime
- 1993-07-30 ES ES93921120T patent/ES2289739T3/es not_active Expired - Lifetime
- 1993-07-30 JP JP6508917A patent/JPH08501940A/ja active Pending
- 1993-07-30 WO PCT/NL1993/000163 patent/WO1994008021A1/en not_active Ceased
- 1993-07-30 FI FI951535A patent/FI951535L/fi unknown
- 1993-07-30 DK DK93921120T patent/DK0680512T3/da active
- 1993-07-30 AU AU48351/93A patent/AU684720B2/en not_active Ceased
- 1993-07-30 CA CA002146145A patent/CA2146145C/en not_active Expired - Lifetime
- 1993-07-30 PT PT93921120T patent/PT680512E/pt unknown
- 1993-07-30 US US08/411,727 patent/US5705161A/en not_active Expired - Lifetime
- 1993-07-30 AT AT93921120T patent/ATE364702T1/de active
- 1993-07-30 EP EP93921120A patent/EP0680512B1/en not_active Expired - Lifetime
-
1995
- 1995-03-28 NO NO19951181A patent/NO321651B1/no not_active IP Right Cessation
Also Published As
| Publication number | Publication date |
|---|---|
| NO951181L (no) | 1995-06-01 |
| PT680512E (pt) | 2007-09-25 |
| AU4835193A (en) | 1994-04-26 |
| US5705161A (en) | 1998-01-06 |
| WO1994008021A1 (en) | 1994-04-14 |
| CA2146145C (en) | 2009-10-27 |
| DE69334147T2 (de) | 2008-02-14 |
| EP0680512B1 (en) | 2007-06-13 |
| DE69334147D1 (de) | 2007-07-26 |
| DK0680512T3 (da) | 2007-10-01 |
| ES2289739T3 (es) | 2008-02-01 |
| NO321651B1 (no) | 2006-06-19 |
| AU684720B2 (en) | 1998-01-08 |
| CA2146145A1 (en) | 1994-04-14 |
| NO951181D0 (no) | 1995-03-28 |
| FI951535A7 (fi) | 1995-06-01 |
| FI951535A0 (fi) | 1995-03-31 |
| FI951535L (fi) | 1995-06-01 |
| JPH08501940A (ja) | 1996-03-05 |
| ATE364702T1 (de) | 2007-07-15 |
| EP0680512A1 (en) | 1995-11-08 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| NL9201716A (nl) | Buitenmembraanvesikel dat voorzien is van een groep polypeptiden welke ten minste de immuunwerking van aan membraan gebonden buitenmembraaneiwitten (OMP's) hebben, werkwijze ter bereiding ervan alsmede een vaccin dat een dergelijk buitenmembraanvesikel omvat. | |
| JP3436756B2 (ja) | 髄膜炎菌のクラスiの外膜タンパク質のワクチン | |
| Masignani et al. | Vaccination against Neisseria meningitidis using three variants of the lipoprotein GNA1870 | |
| RU2181378C2 (ru) | Способ высокоуровневой экспрессии, способ очистки и способ переукладки поринового протеина внешней мембраны менингококка группы в или слитого с ним протеина, высокоочищенный пориновый протеин (варианты), высокоочищенный пориновый слитый протеин (варианты), вакцина для предупреждения бактериального менингита группы в, способ получения конъюгата протеина менингококка группы в и полисахарида, способ предупреждения бактериального менингита, штамм е.coli (варианты) | |
| JP4827196B2 (ja) | 髄膜炎菌類タンパク質nmb1870のドメインおよびエピトープ | |
| US7118757B1 (en) | Meningococcal class 1 outer-membrane protein vaccine | |
| US6613336B2 (en) | Immunogenic conjugates comprising a group B meningococcal porin and an H. influenzae polysaccharide | |
| US6645503B1 (en) | Antigenic conjugates of conserved lipopolysaccharides of gram negative bacteria | |
| KR102626831B1 (ko) | 수막구균 백신 | |
| KR20160127104A (ko) | 변형된 수막구균 fhbp 폴리펩티드 | |
| US20230137914A1 (en) | nOMV-ANTIGEN CONJUGATED COMPOUNDS AND USE THEREOF | |
| EP1572729A2 (en) | Mutant protein and refolding method | |
| CA2007248C (en) | Meningococcal class 1 outer-membrane protein vaccine | |
| de Jonge | Opacity Proteins of Neisseria Meningitidis: Structure-function Relationship and Vaccine Potential | |
| CZ200087A3 (cs) | Imunogenní konjugáty obsahující peptin meningokoků skupiny B a polysacharid H. influenzae | |
| MXPA00000556A (en) | Immunogenic conjugates comprising a group b meningococcal porin and an h. influenzae |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A1B | A search report has been drawn up | ||
| BV | The patent application has lapsed |