WO2000034478A1 - Modification de betacelluline - Google Patents
Modification de betacelluline Download PDFInfo
- Publication number
- WO2000034478A1 WO2000034478A1 PCT/JP1999/006873 JP9906873W WO0034478A1 WO 2000034478 A1 WO2000034478 A1 WO 2000034478A1 JP 9906873 W JP9906873 W JP 9906873W WO 0034478 A1 WO0034478 A1 WO 0034478A1
- Authority
- WO
- WIPO (PCT)
- Prior art keywords
- amino acid
- seq
- cellulin
- acid sequence
- mutein
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Ceased
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/435—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from animals; from humans
- C07K14/475—Growth factors; Growth regulators
- C07K14/485—Epidermal growth factor [EGF], i.e. urogastrone
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K38/00—Medicinal preparations containing peptides
Definitions
- the present invention relates to a mutin having reduced proliferation promoting activity (EGF activity) of epithelial cells such as smooth muscle cells while retaining the activity of promoting cellulin (BTC) differentiation into three kidney cells (BTC activity).
- EGF activity proliferation promoting activity
- BTC activity cellulin
- Cellulin is a protein factor consisting of 80 amino acids that has been excised from a precursor consisting of 178 amino acids, and its entire amino acid sequence has been determined.
- baserulin acts on undifferentiated hepatocytes and has the effect of promoting the differentiation of insulin into hepatic 3 cells (BTC activity) [Mashima et al .; Journal of Clinical Invistigue. Jilin (J. Clin. Invest.), 97: 1647 (1996)] is useful as a preventive or therapeutic agent for diabetes (eg, insulin-dependent diabetes) and kidney dysfunction in diabetes.
- diabetes eg, insulin-dependent diabetes
- cellulin has a growth promoting activity on vascular smooth muscle cells and retinal pigment epithelial cells as described above, it is not suitable for use as a therapeutic agent for diabetes. Their proliferative activity is a problem.
- the inventors of the present invention have conducted various studies on a modified form of cellulin, and found that the modification of cellulin reduced the proliferation promoting activity of smooth muscle cells and the like while maintaining the activity of promoting differentiation into tunnel i3 cells. As a result, the present inventors have found for the first time a modified version of a bay cellulin which can be caused to cause no antigenicity problem when administered to a living body, and as a result of further study, completed the present invention. That is, the present invention relates to a variant of cellulin and a method for obtaining the variant.
- a cell line mutein or a salt thereof which retains the differentiation promoting activity of Teng 0 cells and has a reduced epithelial cell growth promoting activity
- the ratio of the activity of promoting the differentiation of splenic cells to the activity of promoting the proliferation of epithelial cells to the epithelial cell is at least twice as high as that of the cellulin, and the salt of the cellulin mutein or salt thereof according to the above (1);
- betacellulin 1 to 30 amino acid residues at the N-terminus of betacellulin may be deleted, and 1 to 30 amino acid residues from the C-terminus to the 23rd amino acid residue.
- a transformant transformed with a recombinant vector containing DNA encoding the cellulite mutein according to (1) above is cultured to produce the cellulite mutein.
- composition according to the above (9), wherein the composition is an agent for preventing and treating diabetes;
- a method for preventing and treating diabetes which comprises administering to a mammal the above-mentioned celerin mutein or salt thereof according to (1);
- FIG. 1 shows the construction of a 77-residue (3-terminal deletion type) C-cellulin expression plasmid.
- FIG. 2 shows the construction of a 76-residue (4-terminal C-terminal deletion) betacellulin expression plasmid.
- FIG. 3 shows a diagram showing the results of betacellulin mutein electrophoresis performed in Example 7.
- FIG. 4 shows a diagram illustrating the results of the incorporation of 3- P-thymidine into cells performed in Example 8.
- FIG. 5 shows a fluorescence micrograph of the cells differentiated into ⁇ cells described in Example 9.
- FIG. 6 is a diagram showing the results of the activity of promoting 3-cell differentiation performed in Example 12.
- FIG. 7 shows a construction diagram of an expression plasmid for 2- to 76-residue type (1 terminal at the ⁇ -terminal and 4 residues at the C-terminal deleted).
- FIG. 8 shows a construction diagram of an expression plasmid for 24- to 76-residue (23 residues at the ⁇ -terminal and 4 residues at the C-terminus) -based cellulin.
- FIG. 9 is a diagram showing the results of gel staining after electrophoresis performed in Example 19.
- FIG. 10 shows an outline of the construction of plasmid pTB1976 described in Reference Example 1.
- the level at which EGF activity is reduced while BTC activity is maintained is attenuated.
- cellulin refers to Sasada et al .; as described in Biochemical and Biophysical Reseach Communications (Biochem. Biophys. Res. Commun.), 190: 1173 (1993).
- Glu Gly Tyr lie Gly Ala Arg Cys Glu Arg Val Asp Leu Pe Tyr (SEQ ID NO: 35) means a polypeptide containing an amino acid sequence represented by SEQ ID NO: 35.
- Examples of the whole cellulin mutin having BTC activity include betacellulin mutein having an activity of 20% or more, preferably 50% or more of the BTC activity of whole cellulin.
- the cellulin mutin having reduced EGF activity there is a cellulin mutin having an activity of about 20% or less, preferably 15% or less, of the GF activity of the cellulin.
- the cellulin mutin of the present invention in which the EGF activity is reduced while retaining the BTC activity, has a ratio of the differentiation promoting activity of the pancreatic 3 cells to the epithelial cell growth promoting activity which is 2 times smaller than that of the cellulin.
- the cellulin mutin or a salt thereof is more than twice.
- 1 to 40 amino acid residues may be deleted from the ⁇ -terminal of the cellulin.
- one to four amino acid residues including the third amino acid residue Leu from the C-terminus or the fourth amino acid residue Asp from the C-terminus are deleted Or other amino acid residues or other peptide chains, such as sulin mutein or a salt thereof, and more specifically, 1 to 40 or 1 to 20 from the N-terminus. Examples of the above-mentioned cellulin mutein in which an amino acid residue is deleted and the like are mentioned.
- substitution with other amino acid residues or other peptide chains refers to the “replacement with other amino acid residues or other peptide chains” while maintaining BTC activity. If the substitution is within a range that does not impair the feature that “the EGF is attenuated while maintaining the BTC activity”, the amino acid residue or peptide chain of the overnight cellulin mutein of the present invention in which the EGF activity is attenuated, particularly Does not matter.
- other amino acids include non-polar (hydrophobic) amino acids (eg, alanine, leucine, isoleucine, palin, proline, fenylalanine, tributofan, methionine), polar (neutral) amino acids ( Eg, glycine, serine, threonine, cystine, tyrosine, asparagine, glutamine etc.), with a positive charge (basic)
- examples include amino acids (eg, arginine, lysine, histidine, etc.) and negatively charged (acidic) amino acids (eg, aspartic acid, glutamic acid, etc.).
- other peptide chain refers to a peptide chain in which two or more of the above-mentioned “other amino acids” are bound, and are preferably composed of 2 to 4 amino acid residues. Peptide chains and the like.
- amino acids 1 to 4 from the C-terminus may be deleted.
- amino acids 1 to 4 from the C-terminus the third amino acid residue from the C-terminus Leu
- one or four amino acid residues including the fourth amino acid residue Asp from the C-terminus are deleted or substituted with another amino acid residue or another peptide chain.
- a good example is
- amino acid residues 1 to 23 from the N-terminus of basel cellulin (iii) the amino acid residues 1 to 23 from the N-terminus of basel cellulin (AspGlyAsn Ser Thr Arg Ser Pro Glu Thr Asn Gly Leu Leu Cys Gly Asp Pro Glu Glu Asn Cys Ala) may be deleted,
- a cellulin mutein containing an amino acid sequence represented by Tyr lie Gly Ala Arg Cys Glu Arg Val (SEQ ID NO: 4);
- a cellulin mutein containing an amino acid sequence represented by Tyr He Gly Ala Arg Cys Glu Arg Val Asp Phe (SEQ ID NO: 14);
- succin mutein containing the amino acid sequence represented by SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 37 or SEQ ID NO: 38, more preferably, SEQ ID NO: It is a cellulin mutein containing the amino acid sequence represented by 37 or SEQ ID NO: 38. Most preferably, it is a cellulin mutein containing the amino acid sequence represented by SEQ ID NO: 38.
- amino acid sequence represented by 1SEQ ID NO: 1 is particularly preferred.
- amino acid sequence represented by 2SEQ ID NO: 1 An amino acid sequence in which 1 to 40 amino acids have been deleted from the N-terminal of the amino acid sequence, 3 an amino acid sequence represented by SEQ ID NO: 2, 4 an amino acid sequence represented by SEQ ID NO: 2 Or a cellulin mutein containing the amino acid sequence represented by SEQ ID NO: 37 or the amino acid sequence represented by SEQ ID NO: 38;
- 1 amino acid sequence represented by SEQ ID NO: 1 1 amino acid sequence represented by SEQ ID NO: 1, 2 amino acid sequence represented by SEQ ID NO: 2, 3 amino acid sequence represented by SEQ ID NO: 3, ⁇ ⁇ amino acid represented by SEQ ID NO: 4
- a cellulite mutein containing the amino acid sequence represented by the sequence, 5SEQ ID NO: 37 or 6SEQ ID NO: 38, may be mentioned. Particularly preferred is cell lymutein containing the amino acid sequence represented by SEQ ID NO: 38.
- the cellulosic mutin of the present invention in which EGF activity is reduced while retaining BTC activity is as follows: (1) One to 30 amino acid residues at the N-terminus of cellulin are deleted. Base cell with 1 to 5 amino acid residues inserted between the 2nd and 2nd amino acid residues from the C-terminus and the 2nd and 3rd amino acid residues Linmutein or a salt thereof; (2) Cellulin mutein or a salt thereof having an amino acid sequence represented by SEQ ID NO: 45;
- amino acid in the “cellulin mutein (modified molecule) having an amino acid residue inserted therein”
- non-polar (hydrophobic) amino acids eg, alanine, oral isin, isoleucine, norin
- Proline phenylalanine, tryptophan, methionine, etc.
- polar (neutral) amino acids eg, glycine, serine, threonine, cystine, tyrosine, asparagine, glutamine, etc.
- positively charged (basic) amino acids eg, , Arginine, lysine, histidine, etc.
- negatively charged (acidic) amino acids eg, aspartic acid, glutamic acid, etc.
- Asparagine, proline and serine are preferred.
- the “peptide chain” refers to a peptide chain formed by bonding 2 to 5 “amino acids”, preferably a peptide chain consisting of 2 to 4 amino acid residue
- N-terminal 1 to 30 amino acid residues which may be deleted include, for example, (1) a polypeptide having the amino acid sequence represented by the above SEQ ID NO: 35 (2) a modified amino acid sequence of a cellulase which has 1 to 30 amino acid residues at the N-terminus deleted; (3) an amino acid sequence of the N-terminal 1st to 30th amino acid of the cellulase Includes amino acid residues (eg, 1 to 5 amino acid residues) in which the amino acid residues are modified.
- the “Bayselin modified molecule in which 1 to 30 amino acid residues at the N-terminus have been deleted” includes: (1) 1 to 30 amino acid residues at the N-terminal side of cellulose (1) Betacellulin with 1 to 30 amino acid residues deleted at the N-terminal side and amino acid residues (eg, 1 to 5 amino acid residues) added And so on.
- One to thirty amino acid residues at the N-terminus of cellulose may be deleted, and the second to second amino acid residues from the C-terminus (ie, represented by SEQ ID NO: 35
- the 58th and 8th amino acid residues from the N-terminus of the amino acid sequence: G1n) and the 2nd and 3rd amino acid residues that is, 59th to 59th amino acids from the N-terminus of the amino acid sequence represented by SEQ ID NO: 35
- the 1st to 5th amino acid residue between the 3rd amino acid residue: T hr) and the “cellulin-modified molecule” is the N-terminal 1 to Any portion of the 30 amino acid residues may be deleted, between 1 and 5 amino acids between the 2nd and 2nd amino acid residues from the C-terminus As long as the residue has an inserted cellulin-modified molecule, a specific example may be used.
- the nightly cellulin-modified molecule of the present invention, preferably, a nightly cellulin-modified molecule containing the amino acid sequence represented by SEQ ID NO: 44 or SEQ ID NO: 45 and the like are exemplified.
- the cellulin mutein has an N-terminal (amino terminal) at the left end and a C-terminal (carboxyl terminal) at the right end in accordance with the convention of peptide notation.
- these cellulite muteins usually have a carboxyl group at the C-terminus (one C ⁇ H) or a hydroxyl group (one COO-), but have an amide (—C ⁇ NH 2 ) or ester (one C-terminus) at the C-terminus. COOR).
- R of the ester examples include a 6-alkyl group such as methyl, ethyl, n-propyl, isopropyl or n-butyl, Kuropenchiru, C 3 _ 8 cycloalkyl group such as cyclohexyl, phenyl, C 6 _ 1 2 Ariru group such Hiichina Fuchiru, benzyl, phenethyl, full of such benzhydryl Eniru C Bok 2 alkyl or ⁇ - naphthylmethyl, ⁇ - Nafuchiru CJ _ 2 C 7 _ i 4 Ararukiru group such as an alkyl, such as, such as the Ru mentioned Viva port Iruokishimechiru group (5
- cellulin mutin of the present invention also includes those having Met added to the N-terminal.
- a salt with a physiologically acceptable base for example, an alkali metal or the like
- an acid organic acid or inorganic acid
- Acid addition salts are preferred.
- Such salts include, for example, salts with inorganic acids (eg, hydrochloric acid, phosphoric acid, hydrobromic acid, sulfuric acid) or organic acids (eg, acetic acid, formic acid, propionic acid, fumaric acid, maleic acid, succinic acid, Salts with tartaric acid, cunic acid, malic acid, oxalic acid, benzoic acid, methanesulfonic acid, benzenesulfonic acid) are used.
- inorganic acids eg, hydrochloric acid, phosphoric acid, hydrobromic acid, sulfuric acid
- organic acids eg, acetic acid, formic acid, propionic acid, fumaric acid, maleic acid, succinic acid, Salts with tartaric acid, cunic acid, malic acid, oxalic acid
- the cellulase mutein of the present invention can be obtained, for example, by treating the cellulin obtained by genetic engineering or cell culture of a tumor cell described in JP-A-6-87894 with a protease known per se.
- it can also be prepared by treating with carboxypeptidase A, and more preferably by treating carcinomatous carboxypeptidase A.
- it can be produced according to the peptide synthesis method described later.
- it can be produced by culturing a transformant containing DNA encoding beycellin mutin described below.
- tissue or cells of the human warm-blooded animal are homogenized and then extracted with an acid or the like.
- the extract is subjected to salting out, dialysis, Purification and isolation can be performed by combining chromatography such as gel filtration, reverse phase chromatography, ion exchange chromatography, and affinity chromatography.
- a method for synthesizing a peptide for example, either a solid phase synthesis method or a liquid phase synthesis method may be used. That is, the partial peptide or amino acid capable of constituting the betacellulin mutein of the present invention is condensed with the remaining portion, and when the product has a protecting group, the protecting group is removed. By separating, the desired basel cellulin mutein can be produced.
- Known condensation methods and elimination of protecting groups include, for example, the methods described in the following 1 to 5.
- the polypeptide of the present invention can be purified and isolated by a combination of ordinary purification methods such as solvent extraction, distillation, column chromatography, liquid chromatography, and recrystallization.
- the polypeptide obtained by the above method is a free form, it can be converted to an appropriate salt by a known method, and conversely, when the polypeptide is obtained as a salt, it is converted to a free form by a known method be able to.
- amide form of betacellulin mutein a commercially available resin for peptide synthesis suitable for amide formation can be used.
- resins include chloromethyl resin, hydroxymethyl resin, benzhydrylamine resin, aminomethyl resin, 4-benzyloxybenzyl alcohol resin, 4-methylbenzhydrylamine resin, PAM resin, 4 -Hydroxymethylmethylphenylacetamidomethyl resin, polyacrylamide resin, 4- (2 ', 4'-dimethoxyphenylhydroxymethyl) phenoxy resin, 4- (2', 4'-Dimethoxyphenyl-1 Fmoc aminoethyl) phenoxy resin and the like.
- amino acids having a suitably protected amino group and side chain functional group are condensed on the resin in accordance with the sequence of the target peptide in accordance with various known condensation methods.
- the peptide is cleaved from the resin and at the same time, various protecting groups are removed.
- an intramolecular disulfide bond formation reaction is carried out in a highly diluted solution to obtain the desired amide.
- An activating reagent can be used, but carbodiimides are particularly preferable.
- carbodiimides include DCC, N, N'-diisopropylcarboimide, and N-ethyl- ⁇ ⁇ '-(3-dimethylaminopropyl) carbodiimide.
- Activation by these methods involves adding the protected amino acid directly to the resin along with a racemization inhibitor additive (eg, HOBt, H ⁇ Bt, etc.) or using a symmetrical acid anhydride or ⁇ Bt.
- the amino acid protected beforehand as an ester or a HOOBT ester can be added to the resin after activation.
- the solvent used for activation of the protected amino acid and condensation with the resin can be appropriately selected from solvents known to be usable for peptide condensation reactions.
- acid amides such as N-dimethylformamide, N, N-dimethylacetamide and N-methylpyrrolidone
- halogenated hydrocarbons such as methylene chloride and chloroform
- alcohols such as trifluoroethanol
- Sulfoxides such as dimethylsulfoxide, tertiary amines such as pyridine
- ethers such as dioxane and tetrahydrofuran
- nitriles such as acetonitrile and propionitrile
- esters such as methyl acetate and ethyl acetate or the like.
- the reaction temperature is appropriately selected from the range known to be usable for the peptide bond formation reaction, and is usually appropriately selected from the range of about 120 ° C to 50 ° C.
- the activated amino acid derivative is usually used in a 1.5 to 4 times excess.
- Examples of the protecting group for the amino group of the starting amino acid include Z, B oc, Yuichi Shari-pentyloxycarbonyl, isobornyloxycarbonyl, 4-methoxybenzyloxycarbonyl, CI—Z, Br—Z Adamantyloxycarbonyl, trifluoroacetyl, phthaloyl, formyl, 2-nitrophenylsulfenyl, diphenylphosphinothioyl, Fmoc and the like.
- the coercive Mamorumoto carboxyl group for example ⁇ Bok 6 alkyl group described above as R, C 3 _ 8 cycloalkyl Le group, C 7 - other Ararukiru group, 2-Adamanchiru, 4 twelve Torobenjiru, 4-main Examples include toxic benzyl, 4-cyclohexyl benzyl, phenacyl group and benzyloxycarbonyl hydrazide, butyroxybutoxycarbonyl hydrazide, and trityl hydrazide.
- the hydroxyl groups of serine and threonine can be protected, for example, by esterification or etherification.
- Suitable groups for this esterification include, for example, lower alkanoyl groups such as acetyl group, aroyl groups such as benzoyl group, and groups derived from carbon such as benzyloxycarbonyl group and ethoxycarbonyl group.
- groups suitable for etherification include, for example, a benzyl group, a tetrahydrovinyl group, and a short-butyl group.
- the protecting group of the phenolic hydroxyl group of tyrosine include B zl, C l 2 - B zl, 2- nitrobenzyl, B r- Z, evening like one tert-butyl.
- Histidine imidazole protecting groups include Tos, 4-methoxy-2,3,6-trimethylbenzenesulfonyl, DNP, benzyloxymethyl, Bum, Boc, Trt, Fmoc and the like.
- the activated carboxyl groups of the raw materials include, for example, the corresponding acid anhydrides, azides, active esters [alcohols (eg, pentachlorophenol, 2,4,5-trichloromouth phenol, 2,4-dinitro Phenol, cyanomethyl alcohol, paranitrophenol, H ⁇ NB, N-hydroxysuccinimide, N-hydroxyfurimide, ester with HOB t)].
- the activated amino group of the raw material includes, for example, the corresponding phosphoric amide.
- Methods for removing (eliminating) the protecting group include catalytic reduction in a hydrogen stream in the presence of a catalyst such as Pd black or Pd carbon, or anhydrous hydrogen fluoride, methanesulfonate, trifluorosulfone, or the like.
- a catalyst such as Pd black or Pd carbon
- anhydrous hydrogen fluoride, methanesulfonate, trifluorosulfone, or the like include acid treatment with an acid, trifluoroacetic acid or a mixture thereof, base treatment with diisopropylethylamine, triethylamine, piberidine, piperazine, and the like, and reduction with sodium in liquid ammonia.
- the elimination reaction by the above acid treatment is generally carried out at a temperature of -20 ° C to 40 ° C.
- a cation scavenger such as 1,4-butanedithiol or 1,2-ethanedithiol
- Histi The 2,4-dinitrophenyl group used as the imidazole protecting group of gin is removed by thiophenol treatment, and the formyl group used as the indole protecting group of tributofan is 1,2-ethanedithiol, 1,4-butane described above.
- alkali treatment with dilute sodium hydroxide and dilute ammonia.
- a method for introducing a protecting group to a functional group that should not be involved in the reaction, a method for removing the protecting group, and a method for activating a functional group involved in the reaction can be performed according to a method known per se or a method analogous thereto. Good.
- Another method for obtaining an amide form of cellulin mutein is to first amidate the ⁇ -carboxyl group of the carboxyl-terminal amino acid and then extend the peptide chain to the desired length on the amino group side. Thereafter, a peptide (or amino acid) from which only the protecting group for the ⁇ -amino group at the ⁇ -terminal of the peptide chain has been removed and a peptide (or amino acid) from which only the protecting group for the carboxyl group at the C-terminal has been removed are prepared.
- the peptide is condensed in a mixed solvent as described above. Details of the condensation reaction are the same as described above.
- the crude polypeptide is purified by various known purification means, and the main fraction is freeze-dried to obtain an amide of the desired polypeptide.
- the DNA encoding the betacellulin mutein of the present invention may include (1) 1 to 40 amino acid residues from the C-terminus of the cellulin, and 1 to 4 amino acids from the C-terminus.
- any DNA may be used as long as it contains a DNA encoding a betacellulin mutein having the amino acid sequence represented by SEQ ID NO: 45. Specifically, SEQ ID NO: 1 to SEQ ID NO: 14, SEQ ID NO: 37, SEQ ID NO:
- DNA containing DNA having the nucleotide sequence represented by 43 (3) DNA containing DNA having the nucleotide sequence represented by SEQ ID NO: 47 to SEQ ID NO: 49, (4) Stringent conditions
- Non-hybridizing ability DNA that encodes a polypeptide having the same amino acid sequence is used.
- Hybridization can be performed according to a method known per se or a method analogous thereto.
- the cellulite mutein of the present invention may be a DN encoding the cellulite mutein.
- the expression vector of the beta-cellulin mutein of the present invention used in the production by culturing the transformant containing A is, for example, (1) a DNA encoding the beta-cellulin mutein of the present invention. (2) ligating the DNA fragment downstream of the promoter in a suitable expression vector; Can be manufactured.
- vectors include Escherichia coli-derived plasmids (eg, pBR322, pBR325, pUC12, pUC13), Bacillus subtilis-derived plasmids (eg, pUB110, pTP5, pC194), yeast Derived plasmids (eg, pSH19, pSH15), bacteriophages such as ⁇ phage, and animal viruses such as retrovirus, vaccinia virus, and baculovirus are used.
- Escherichia coli-derived plasmids eg, pBR322, pBR325, pUC12, pUC13
- Bacillus subtilis-derived plasmids eg, pUB110, pTP5, pC194
- yeast Derived plasmids eg, pSH19, pSH15
- bacteriophages such as ⁇ phage
- animal viruses such as retrovirus, vaccinia virus
- the promoter used in the present invention may be any suitable promoter corresponding to the host used for gene expression.
- the promoter derived from SV40, the retrovirus promoter, the metamouth thionine promoter, the heat shock promoter, the cytomegalovirus promoter, and the SR hypromo One can be used.
- the host is a bacterium belonging to the genus Escherichia, the trp promoter, T7 promoter, lac promoter, recA promoter, ⁇ PL promoter, 1 pp promoter, etc.
- the host is a yeast, SP ⁇ 1 promoter, SP ⁇ 2 promoter, pen P promoter, etc.If the host is yeast, PH05 promoter, PGK promoter, GAP promoter, ADH 1 promoter
- the GAL promoter is preferable.
- a polyhedrin promoter, a P10 promoter and the like are preferable.
- the expression vector may contain, in addition to the above, an enhancer, a splicing signal, a poly-A addition signal, a selection marker, and an SV40 replication origin (hereinafter sometimes abbreviated as SV40 ori), if desired.
- an enhancer e.g., a splicing signal
- a poly-A addition signal e.g., a selection marker
- an SV40 replication origin e.g., SV40 ori
- the selectable marker include a dihydrofolate reductase (hereinafter sometimes abbreviated as dh fr) gene [methotrexate (MTX) resistance] and an ampicillin resistance gene (hereinafter abbreviated as Amp) ), The neomycin resistance gene (hereinafter sometimes abbreviated as Neo, G4 18 resistance), etc.
- dh fr dihydrofolate reductase
- Amp ampicillin resistance gene
- Neo neomycin resistance gene
- a signal sequence suitable for the host to the N-terminal side of the cellulin mutein.
- Pho A signal if host is Escherichia Sequence, ⁇ mp A, signal sequence, etc. If the host is a Bacillus bacterium, the hairlamine signal sequence, subtilisin, the signal sequence, etc., if the host is yeast, the mating factor o;
- the host is an animal cell, such as (MF ⁇ ) signal sequence, Inversease 'signal sequence, etc., for example, insulin signal sequence, ⁇ -inulin ferron signal sequence, antibody molecule, signal sequence Etc. can be used respectively.
- a transformant can be produced using the thus-constructed vector containing DNA encoding succin mutein.
- bacteria of the genus Escherichia bacteria of the genus Bacillus, yeast, insect or insect cells, animal cells, and the like are used.
- Escherichia examples include Escherichia coli K12 K12 DH1 [Procedings of the National Academy of Sciences, Obc. Natl. Acad. Sci. USA), Volume 60, 160 (1968)], JM 103 [Nucleic Acids Research ', (Nucleic Acids Research), Volume 9, 309 (1981)], J J221 [Journal' ⁇ Moleki Yura-I 'Biology (Journal of Molecular Biology)], 120, 517 (1 978)], HB 101 [Journal' ob '' Molecular 'Biology, 41, 459 (1969)], C 600 [ Genetics, 39, 440 (1954)], MM294 (Pro atl. Acad. Prossings of the National Affairs Demichi'ob'Sciences of the U.S.A. Sci. USA), 73, 4174 (1976)].
- Bacillus bacteria examples include, for example, Bacillus subtilis M11114 [Gene, 24, 255 (1983)], 207-21 [Journal of Biochemistry, 95] , 87 (1984)].
- yeast for example, Saccharomyces cerevisiae AH22, AH22R—, NA87-11A, DKD-5D, 20B-12 and the like are used.
- insects for example, silkworm larvae are used [Maeda et al. PC J
- Insect cells include, for example, when the virus is Ac NPV, a cell line derived from the larvae of night moth (Spodoptera frugiperda cell; S f cell), MG1 cells derived from the midgut of Trichoplusia ni, and eggs derived from eggs of Trichoplusia ni High Five TM cells, cells derived from Mamestra brassicae or cells derived from Estigmena acrea are used. If the virus is BmNPV, a silkworm-derived cell line (Bombyx mori N; BmN cell) is used. Examples of the Sf cell include Sf9 cell (ATCC CRL1711), Sf21 cell [Vaughn, JL et al., In Vitro, Vol. 13, pp. 213-217 ( 1977) :) is used.
- animal cells examples include monkey COS-7 cells, Vero cells, Chinese hamster cells CHO, DHFR gene-deficient Chinese hamster cells CHO
- mouse L cells mouse 3T3 cells, mouse myeoma cells, human HEK293 cells, human FL cells, 293 cells, C127 cells, BAL B 3T3 cells, Sp-2 / 0 cells are used.
- Transformation of animal cells is performed, for example, according to the method described in Virology, 52, 456 (1973).
- Examples of a method for introducing an expression vector into cells include the lipofection method.
- the above-mentioned method of selecting cells in which the expression vector introduced into the animal cells is integrated into the chromosome by clone selection is used. is there. Specifically, transformants are selected using the above selection marker as an index. Further, by repeatedly performing clone selection on the animal cells obtained using the selection marker in this manner, a stable animal cell line having high expression ability such as bersel mutein of the present invention is obtained. be able to.
- the dh fr gene was used as a selective agent, the MTX concentration was gradually increased, and culturing was carried out to select resistant strains.
- the DNA to be transfected can be amplified intracellularly to obtain a more highly expressed animal cell line.
- the above transformant is cultured under conditions that allow expression of the DNA encoding the cellulin mutein of the present invention, and the beta cellulin mutein of the present invention is produced and accumulated, whereby the vector of the present invention is produced. Evening cellulin mutin can be manufactured.
- a liquid medium is suitable as a medium used for the cultivation, and includes a carbon source necessary for growth of the transformant.
- Nitrogen sources, inorganic substances, etc. are included.
- Carbon sources include, for example, glucose, dextrin, soluble starch, sucrose, etc.
- Nonrogen sources include, for example, ammonium salts, nitrates, corn chips, liquor, peptone, casein, meat extract, soybean meal, potato extract, etc.
- Inorganic or organic substances For example, calcium chloride, sodium dihydrogen phosphate, magnesium chloride and the like can be mentioned.
- yeast, vitamins, growth promoting factors and the like may be added.
- the pH of the medium is preferably about 5-8.
- a culture medium for culturing a bacterium belonging to the genus Escherichia for example, an M9 medium containing glucose and casamino acid [Miller, Journal of Experiments in Molecular Journal (Journal of Experiments in Molecular) Genetics), 431-433, Cold Spring Harbor Laboratory, New York 1972].
- an agent such as 3) 3-indolylacrylic acid can be added to make the promoter work efficiently.
- the cultivation is usually carried out at about 15 to 43 ° C for about 3 to 24 hours, and if necessary, aeration and stirring can be applied.
- the cultivation is usually carried out at about 30 to 40 ° for about 6 to 24 hours, and if necessary, aeration and stirring can be applied.
- Culture is usually carried out at about 20 ° C to 35 ° C for about 24 to 72 hours, and aeration and stirring are added as necessary.
- the medium used is Grace's Insect.
- the pH of the medium is about 6.2 to 8.
- Culture is usually performed at about 27 ° C for about 3 to 5 days, and aeration and stirring are added as necessary.
- the culture medium When culturing a transformant in which the host is an animal cell, the culture medium may be, for example, about 5 to
- MEM medium containing 20% fetal bovine serum [Science, 122, 50 1 (1952)], DMEM medium (Virology, 8, 396 (1959)), RPMI 1640 medium [Journal of the American Medical Intermediary Association (The Journal of the American Medical) Association, 1999, 519 (1967)], 199 medium [Proceding of the Society for the Biological Medicine], 73, 1 (Proceding of the Society for the Biological Medicine), 73, 1 ( 1950)].
- the pH is about 6-8. Culture is usually performed for about 30 to 40 to about 15 to 60 hours, and aeration and stirring are added as necessary.
- CHO (dhfr-) cells and the dhfr gene are used as selection agents, it is preferable to use a DMEM medium containing dialyzed fetal serum containing almost no thymidine.
- the betacellulin mutein of the present invention can be separated and purified from the above culture by, for example, the following method.
- the cells or cells are collected by a known method, and the cells are suspended in an appropriate buffer, and then subjected to ultrasound, lysozyme and After destroying the cells or cells by freezing and thawing, a method of obtaining a crude polypeptide extract by centrifugation or filtration may be appropriately used.
- the buffer may contain a protein denaturing agent such as urea or guanidine hydrochloride, or a surfactant such as Triton X-100 (registered trademark, hereinafter sometimes abbreviated as TM).
- betacellulin mutein When betacellulin mutein is secreted into the culture solution, after the culture is completed, the supernatant is separated from the cells or cells by a method known per se, and the supernatant is collected.
- Purification of the polypeptide of the present invention contained in the thus obtained culture supernatant or extract can be carried out by appropriately combining known separation and purification methods.
- These known separation and purification methods include methods utilizing solubility such as salting-out and solvent precipitation, dialysis, ultrafiltration, gel filtration, and SDS-polyacrylamide gel electrophoresis, mainly molecular weight.
- Method using difference in charge such as ion-exchange chromatography, method using specific affinity such as affinity chromatography, reversed-phase high-speed chromatography, etc. Hydrophobic
- a method utilizing the difference, a method utilizing the difference between the isoelectric points such as isoelectric focusing and chromatofocusing are used.
- the thus-obtained celerin mutin of the present invention when obtained in a free form, it can be converted to a salt by a method known per se or a method analogous thereto. It can be converted to a free form or another salt by a known method or a method analogous thereto.
- the recombinant cell-produced cellulin mutein of the present invention may be arbitrarily modified or partially removed by the action of a suitable protein-modifying enzyme. You can also.
- a suitable protein-modifying enzyme for example, tribusin, chymotrypsin, arginyl endopeptidase, protein kinase, glycosidase and the like are used.
- the obtained cellulin mutein may be subjected to a refolding step described in, for example, Japanese Patent Application Laid-Open No. 10-1991-1989. Further, by adding cellulite mutein having Met added to the N-terminus to the N-terminal Met removal reaction described in Japanese Patent Application Laid-Open No. 10-1989 / 1989, the N-terminus can be obtained. Met can also be removed.
- DNA encoding the cellulin mutein of the present invention or the cellulin mutein or a salt thereof of the present invention may be used for diabetes (eg, insulin-dependent diabetes mellitus (type I diabetes), etc.), dysfunction of the kidney in diabetes, senility
- diabetes eg, insulin-dependent diabetes mellitus (type I diabetes), etc.
- dysfunction of the kidney in diabetes e.g.
- senility e.g, diabetes-dependent diabetes mellitus (type I diabetes), etc.
- drugs for undifferentiated renal cancer especially for the prevention and treatment of diabetes (eg, insulin-dependent diabetes, etc.) be able to.
- the cellulin mutin of the present invention or a salt thereof or a DNA encoding the same can be used as a safe and low toxic drug because EGF activity is attenuated and there is no problem of antigenicity.
- the cellulin mutein or salt thereof or the DNA encoding the same according to the present invention may be used for diabetes (for example, insulin-dependent sugar). It can be used as a remedy for diabetes mellitus dysfunction in diabetes mellitus, dysfunction of senile dysfunction associated with senile decrease in insulin secretion, and a therapeutic / prophylactic agent for diseases such as undifferentiated kidney cancer.
- the cellulin mutin of the present invention or a salt thereof or DNA encoding the same as the above-mentioned medicament can be carried out in a conventional manner.
- aseptic solution orally as tablets, capsules, elixirs, microcapsules, etc. coated with sugar coating or enteric coating as needed, or with water or other pharmaceutically acceptable liquids It can be used parenterally in the form of injections, or suspensions.
- the compound or a salt thereof is mixed with a physiologically acceptable carrier, flavoring agent, excipient, vehicle, preservative, stabilizer, binder and the like in a unit dosage form required for a generally accepted formulation. By doing so, it can be manufactured.
- the amount of active ingredient in these preparations is such that a suitable dosage in the specified range can be obtained.
- Additives that can be incorporated into tablets, capsules, etc. include binders such as gelatin, corn starch, tragacanth gum, gum arabic, excipients such as crystalline cell mouth, corn starch, gelatin, alginic acid, etc.
- binders such as gelatin, corn starch, tragacanth gum, gum arabic, excipients such as crystalline cell mouth, corn starch, gelatin, alginic acid, etc.
- a bulking agent such as magnesium stearate, a sweetener such as sucrose, lactose or saccharin, and a flavoring agent such as peppermint, cocoa oil or cherry may be used.
- the unit dosage form is a capsule, the above type of material can further contain a liquid carrier such as an oil or fat.
- Sterile compositions for injection are formulated according to standard pharmaceutical manufacturing methods, such as by dissolving or suspending the active substance in a pan-vehicle, such as water for injection, or naturally occurring vegetable oils such as sesame oil or coconut oil. be able to.
- aqueous liquids for injection include physiological saline, isotonic solutions containing glucose and other adjuvants (eg, D-sorbitol, D-mannitol, sodium chloride, etc.).
- Agents such as alcohols (eg, ethanol), polyalcohols (eg, propylene glycol, polyethylene glycol), and nonionic surfactants (eg, polysorbate 80 (TM), HCO-50) may be used in combination.
- oily liquid examples include sesame oil and soybean oil, and may be used in combination with benzyl benzoate, benzyl alcohol and the like as a solubilizing agent.
- buffers for example, phosphate buffer, sodium acetate buffer
- soothing agents for example, benzalkonium chloride, proforce hydrochloride, etc.
- stabilizers for example, human serum albumin, polyethylene glycol, etc.
- a preservative eg, benzyl alcohol, phenol, etc.
- an antioxidant and the like.
- the prepared injection solution is usually filled in an appropriate ampoule.
- the preparations obtained in this way are safe and low toxic, and can be used, for example, in humans and mammals (e.g., Etc.).
- the dosage of the beessellin mutein or a salt thereof according to the present invention varies depending on the symptoms and the like. However, in the case of oral administration, generally, in an adult diabetic patient (with a body weight of 60 kg), the dose is about 0 per day. .1 to 100 mg, preferably about 1.0 to 50 mg, more preferably about 1.0 to 20 mg.
- the single dose varies depending on the administration target, target organ, symptoms, administration method, etc., but it is used as an injection for adult diabetic patients (with a body weight of 60 kg).
- a 1 a Alanine
- Th r Threonine
- NMP N-methylpyrrolidone A substituent commonly used in the present specification.
- Trt trityl group The sequence numbers in the sequence listing in the present specification show the following sequences.
- Fig. 2 shows the amino acid sequence of Bessellin mutein (BTC 1-76) of the present invention. [SEQ ID NO: 3]
- FIG. 1 shows the amino acid sequence of succin mutein (BTC 1-76, 78-80) of the present invention.
- FIG. 1 shows the amino acid sequence of celite mutein (BTC 1-76, 78, 79) of the present invention.
- FIG. 1 shows the amino acid sequence of succin mutein (BTC 1_76, 78) of the present invention.
- Fig. 2 shows the amino acid sequence of succin mutein (BTCl-77, 79, 80) of the present invention.
- FIG. 1 shows the amino acid sequence of succin mutein (BTC 1-77, 80) of the present invention.
- FIG. 1 shows the amino acid sequence of cellulin mutin (BTC 31-76, 78-80) of the present invention.
- Fig. 2 shows the amino acid sequence of beta-cellulin mutein (BTC 31-76, 78, 79) of the present invention.
- FIG. 1 shows the amino acid sequence of succin mutein (BTC31-76, 78) of the present invention.
- FIG. 1 shows the amino acid sequence of succin mutein (BTC 31-77, 79, 80) of the present invention.
- the amino acid sequence of the cellulin mutin (BTC 31-77, 79) of the present invention is Show.
- SEQ ID NO: 1 shows the nucleotide sequence of cDNA encoding the amino acid sequence represented by amino acid sequence represented by SEQ ID NO: 1.
- SEQ ID NO: 2 shows the nucleotide sequence of cDNA encoding the amino acid sequence represented by amino acid sequence represented by SEQ ID NO: 2.
- SEQ ID NO: 3 shows the nucleotide sequence of cDNA encoding the amino acid sequence represented by amino acid sequence represented by amino acid sequence.
- SEQ ID NO: 5 shows the nucleotide sequence of cDNA encoding the cellulin mutein represented by the amino acid sequence represented by SEQ ID NO: 5.
- SEQ ID NO: 6 shows the nucleotide sequence of cDNA encoding the cell line mutin represented by the amino acid sequence represented by SEQ ID NO: 6.
- SEQ ID NO: 24 This shows the nucleotide sequence of cDNA encoding the cellulite mutein represented by the amino acid sequence represented by SEQ ID NO: 10.
- SEQ ID NO: 13 shows the nucleotide sequence of cDNA encoding the cellulin mutein represented by the amino acid sequence represented by SEQ ID NO: 13.
- Primer RI-3Xho used in Examples 10 and 27 described below Shows the base sequence.
- FIG 13 shows the nucleotide sequence of primer 4 used in Examples 13 and 16 described later.
- SEQ ID NO: 38 shows the nucleotide sequence of cDNA encoding the cellulin mutein represented by the amino acid sequence represented by SEQ ID NO: 38.
- 1 shows the amino acid sequence of the modified amino acid molecule of the present invention (BTC31-58, Asn, Pro, Ser, 59-80).
- 1 shows the amino acid sequence of the betacellulin-modified molecule (Asn, Ser, Asp, Ser, G1u, BTC 38-80) of the present invention.
- [SEQ ID NO: 46] j shows the amino acid sequence of a cellulin-modified molecule (BTC 1-58, Asn, Pro, Ser, -80).
- SEQ ID NO: 46 shows the nucleotide sequence of cDNA encoding the cellulase-modified molecule represented by the amino acid sequence shown in SEQ ID NO: 46.
- FIG. 14 shows the nucleotide sequence of Primer BT—95 h used in Reference Example 1 and Example 22 described later.
- Example 15 shows the nucleotide sequence of primer BTC-7 used in Example 23 described later.
- erchiacoli)] MM 294 (DE 3) / p TC II BTC77 has the accession number FERM BP-6584 as of January 24, 1998 on January 24, 1998 by the National Institute of Advanced Industrial Science and Technology (NI BH; Ibaraki, Japan) It has been deposited with Tsukuba East Co., Ltd. It has been deposited with the Fermentation Research Institute (IFO; 2-17-85, Jusanhoncho, Yodogawa-ku, Osaka-shi, Japan) under the accession number IFO16214 on January 1, 1998.
- IFO Fermentation Research Institute
- E. coli [Escherichiacoli] MM294 (DE 3) ZpTC II BTC76 obtained in Example 4 described below was obtained under the accession number FERM BP-6583 on January 24, 1998 by the Ministry of International Trade and Industry. Deposited at the Institute of Biotechnology and Industrial Technology. It has been deposited with the Fermentation Research Institute (IF ⁇ ) under the accession number IF ⁇ 16213 on January 2, 1998.
- IF ⁇ Fermentation Research Institute
- E. coli [Escherichiacoli] MM 294 (DE 3) / p TC II BTC2-76 obtained in Example 13 described below was obtained under the accession number FERM BP-6948 on January 24, 1999. It has been deposited with the Institute of Biotechnology and Industrial Technology of the Ministry of International Trade and Industry. It has been deposited with the Fermentation Research Institute (IF ⁇ ) under the accession number IF 0 16334 on January 9, 1999.
- IF ⁇ Fermentation Research Institute
- E. coli [Escherichiacoli] MM 294 (DE3) / pTC II BTC24-76 obtained in Example 16 described below was designated as Accession No. FERM BP-6949 on January 24, 1999 by the Ministry of International Trade and Industry. It is deposited at the Institute of Biotechnology and Industrial Technology, the National Institute of Advanced Industrial Science and Technology. It was deposited with the Fermentation Research Institute (IFO) under the accession number IFO 16335 on January 9, 1999.
- IFO Fermentation Research Institute
- Example 1 The Escherichiaco 1 i MM 294 (DE 3) / p Lys S, p TB 151 carrying the plasmid pTB 1516 used in Reference Example 1 described below was obtained from the Institute of Biotechnology and Industrial Technology, NI BH) and deposited under the accession number FERM BP-3836 from April 21, 1992 (Heisei Era 4), and accession number IF ⁇ from the fermentation research institute on April 16, 1992 (Heisei Era 4). Deposited as 15 282.
- NI BH Institute of Biotechnology and Industrial Technology
- a 77-residue (3-terminal C-terminal deletion) vector cellulin expression plasmid was constructed as follows (Fig. 1).
- 77-residue type C-terminal three-residue deletion type
- the structural gene for cellulin cellulase was converted to a cellulase expression plasmid ⁇ ⁇ 41 [Se ⁇ et al .; Growth Factors, 13: 181 (1996)] shows that a primer 1 (5′—CATATGGATGGGAATT CCACCAGAAGTCCTG) having an NdeI cleavage site and an initiation codon adjacent to the upstream of the structural gene, and a stop codon and a BamHI after the 77th aspartate.
- Amplification was performed by PCR using Primer 2 having a cleavage site (5'-GG AT CCCTA GTCAACTCTCTCACACCTTGCTCC).
- the gene amplified by PCR was ligated to the pCR2.1 vector using TAo rig i n a c l in ng ng kit (manufactured by Invitrogen) to prepare pCR 2.1 / BTC77.
- This was introduced into E. coli JM109, and transformants were selected using ampicillin resistance and 3-galactosidase activity as indices.
- a transformant having pCR2.1 / BTC77 was cultured, and pCR2.1 / BTC77 was prepared using QIArep8Miniprepkit (Qiagen).
- PBR322 was cut with NdeI, the ends were blunt-ended with T4 DNA polymerase (DNA B1 antigen kit, manufactured by Takara Shuzo Co., Ltd.), and religated to delete the NdeI recognition site.
- P BRdes Nde was prepared. After cutting pET3c with BglII-EcoRV and recovering a fragment of about 0.26 kbp, the ends were blunt-ended with T4 DNA polymerase, and the ScaI fragment of pBRdes nes Nde was used. Ligation was performed to produce PBRZT7 de sNde. In addition, pBR32 was introduced by site-directed mutagenesis (Quick Change, STRATA GENE).
- PBR322 desBam was prepared from which the BamHI recognition site was deleted.
- the SphI-EcoRV fragment of pBR322 desBam was ligated to the SphI-EcoRV fragment of pBRZT7desNde to produce a tetracycline resistance expression vector pTCII.
- PCR2.1 / BTC77 was digested with NdeI and BamHI and subjected to agarose gel electrophoresis, and a 77-residue-type overnight serine structural gene of about 240 bp was QI Aq uick S pin Purification on Kit. (Manufactured by Qiagen).
- the expression vector pTCII was digested with NdeI and BamHI and subjected to agarose gel electrophoresis. Similarly, a band of about 4.6 kbp was recovered. After ligating the 77-residue type cellulin structural gene to the NdeI-BamHI fragment of the expression vector pTCII, it was introduced into Escherichia coli JM109 to select transformants with tetracycline resistance. Then, the plasmid was recovered from the strain again to obtain an expression plasmid pTC II BTC 77.
- LB medium containing 1 OmgZL of tetracycline 1% peptone, 0.5% yeast extract, 0.5% sodium chloride
- the cells were cultured in 1 liter at 30 ° C for 16 hours.
- the obtained culture solution (15 OmL) was added to the main fermentation medium (1.68% sodium monohydrogen phosphate, 0.3% sodium dihydrogen phosphate, 0.1% ammonium chloride, 0.05% sodium chloride, 0.04% magnesium sulfate, 0.02% Newpol LB— 625, 0.0005% thiamine hydrochloride, 1.5% glucose, 1.0% casamino acid, 1.0% yeast extract) 1.5 liters
- the mixture was transplanted to the charged 2 L jar mentor, and aeration and agitation culture was started at 37 ° C, aeration rate of 2 L / min, and agitation speed of 500 rpm.
- Centrifugation (10000 rpm, 20 minutes) was performed to obtain 26 OmL of the centrifuged supernatant.
- the centrifuged supernatant was concentrated using a YM3 membrane (fraction molecular weight: 3000, Millipore).
- the pH was adjusted to 5.0 with hydrochloric acid.
- centrifugation 10,000 rpm, 20 minutes
- the obtained centrifuged supernatant was equilibrated with 5 OmM sodium acetate buffer (H5.0), SP-Toyopearl 650M column (2.2 cm x 12 cm).
- 76-residue type (C-terminal 4 residues deleted) Structural gene of baseline cellulin was constructed from pTC II / BTC 77 constructed in Example 1 adjacent to the upstream of the structural gene at NdeI cleavage site and start The primer 1 having a codon (5'-CATATGGATGGG AATTCCACCAGAAGTCCTG) and the primer 1 having a termination codon and a BamHI cleavage site after the 76th valine (5'-GG AT CCCTAA.
- the gene amplified by PCR was It was ligated to the pCR2.1 vector using the originalcl on ng kit (manufactured by Invitrogen) to produce pCR2.1ZBTC 76. This was introduced into E.
- a transformant having pCR2.1 / BTC76 was cultured, and pCR2.1 / BTC76 was selected using QI Aprep 8 Miniprepkit (Qiagen). Was prepared.
- pCR2.1ZBTC76 was digested with NdeI and BamHI and subjected to agarose gel electrophoresis, and the approximately 240 bp 76-residue cellulin structural gene was ligated with a QIA quick Spin Purification Kit ( (Qiagen).
- the expression vector pTCIII prepared in Example 1 was digested with NdeI and BamHI and subjected to agarose gel electrophoresis. Similarly, a band of about 4.6 kbp was recovered.
- After ligating the 76-residue type cellulin structural gene to the NdeI-BamHI fragment of the expression vector pTCII it was introduced into E. coli JM109, and a transformant was selected with tetracycline resistance. The plasmid was recovered again from the resulting plasmid to give the expression plasmid pTC II BTC76.
- LB medium containing 1 Omg / L tetracycline 1% peptone, 0.5% yeast extract, 0.5% sodium chloride
- a 76 residue type overnight cellulin expressing strain MM294 (DE3) / pTC II BTC76 The cells were cultured in 1 liter at 30 ° C. for 16 hours. The obtained culture solution was used for the main fermentation medium (1.68% sodium monohydrogen phosphate, 0.3% sodium dihydrogen phosphate, 0.1% ammonium chloride, 0.05% sodium chloride, 0.024%).
- centrifugation (10000 rpm, 20 minutes) was performed to obtain 20 L of centrifuged supernatant.
- the centrifuged supernatant was concentrated using a Pellicon cassette system (molecular weight cut-off: 5000, Millipore).
- Concentrated desalted solution 3 After adding 13.2 L of 2M urea to 3 L, the pH was adjusted to 5.0 with hydrochloric acid. It is adsorbed on a POROS 50HS column (2.2 cm x 12 cm, Nippon Perceptive) equilibrated with 50 mM sodium acetate buffer (pH 5.0) at a flow rate of 3 OmL / min, and the buffer used for equilibration is sufficient.
- the characteristics of the Met-77 residue type cellulin obtained in Example 3 and the Met-76 residue type cellulin obtained in Example 6 were determined as follows.
- a modified overnight cellulin and a Me-80 residue type overnight cellulose prepared according to the method described in JP-A-10-191989 were added to a sample buffer (125 mM Tris-HC 1% sodium dodecyl sulfate, 1%).
- the cells were suspended in 5% glycerol, 5% 2-mercaptoethanol, 0.005% bromphenol and electrophoresed on Multigel 15-25 (Daiichi Pure Chemicals). The gel after electrophoresis was stained with Rapid CBBBB (Kanto Chemical Co., Ltd.). It was a single band [Figure 3].
- the modified betacellulin is subjected to gas phase hydrolysis with 6 N hydrochloric acid containing 4% thioglycolic acid at 110 ° (:, 24, and 48 hours, and the amino acid analyzer (Hitachi L-1 8500AAmino Acid Ana 1 yzer As a result, all the variants contained methionine derived from the initiation codon ATG, and were consistent with the amino acid composition expected from the cDNA base sequence [Table 1, Table 2].
- N-terminal amino acid sequence analysis was determined using a gas-phase protein sequencer (Applied Biosystems Model 477A). As a result, all the variants were in agreement with the amino acid composition predicted from the cDNA base sequence. All variants had a methionine derived from the initiation codon ATG at the N-terminus as in the 80 residue type [Tables 3 and 4]. [Table 3] N-terminal amino acid sequence analysis of 77-residue type betacellulin
- the cells are washed once with PBS, fixed with 10% formaldehyde, treated with 0.1% Triton X-100 for 5 minutes, and then added with Block Ace (Snow Brand, Japan) and incubated at room temperature for 40 minutes. Blocking was performed for minutes.
- An anti-insulin antibody (Advanced 'Immuno Chemical) diluted with 10% Block Ace was added and reacted at room temperature for 40 minutes. After 0.1% Triton X-100 was added and left at room temperature for 5 minutes, it was washed three times with PBS.
- the activity of promoting differentiation into three cells was also performed using AR42J cells transformed with alkaline phosphatase ligated as a repo overnight downstream of the insulin promoter.
- cells differentiated into / 3 cells by cellulase produce alkaline phosphatase, and by measuring the activity of this alkaline phosphatase, the activity of promoting differentiation into 3 cells is quantitatively measured. Is possible You.
- Genomic DNA was prepared from the rat tail according to a conventional method. Using this DNA as a template, a primer synthesized based on the previously reported base sequence of rat insulin II gene promoter (GenBank: Accession No. J 00748).
- the amplified 0.75 Kb DNA fragment was isolated, and the nucleotide sequence of the cloned fragment was determined using the plasmid pTB1881 obtained by incorporating it into the pT7 Blue vector (Novagen 69820-1). I confirmed that it was overnight. Plasmid pTB1881 was digested with XhoI-ClaI to obtain a 0.73 kb DNA fragment, which was rat insulin promoter overnight. Next, the expression plasmid PTB 1330 of 2.0 kbc DNA encoding human placental alkaline phosphatase (PLAP) (J. Berger et al., Gene 66, 1 (1988)) was transformed into XhoI-Hind.
- PLAP placental alkaline phosphatase
- AR42 J cells plasmid containing ne o r gene PL AP expression plasmid pTB 1898 and Tn 5 pMCn e opo ly A STRATAGENE
- STRATAGENE a transflector Ekushi Yon reagent T rans IT TM - LT 1 ( M irus, P e nVe ra Coporation).
- the transfected cells were cultured in DMEM supplemented with 10% fetal bovine serum for 2 days, and then added to a selective medium supplemented with 800 g / mL of G418 (genetecin, GI BCO BRL), followed by culturing. Clones were isolated from cells grown to G418 resistance by the limiting dilution method.
- the cells of each clone are seeded on a 24-well plate, cultivated with Me-80 residue type cellulin overnight with or without 20 ng ZmL, and the culture supernatant is collected and heat-treated at 65 ° C for 30 minutes. After performing, the alkaline phosphatase activity in the medium was measured. A clone showing an increase in alkaline phosphatase activity when 80-residue type cellulin was added was selected. The results of some clones are shown in [Table 7] below.
- PLAP-expressing AR42 J cells Measurement of 3-cell differentiation promoting activity
- the PLAP-expressing AR42J cells constructed in Example 11 were combined with various concentrations of the met-77 residue type cellulin prepared in Example 3 and the Met-76 residue type prepared in Example 6 for the cell line.
- Dulbecco's Modified Eagle containing Me-80 residue type Cellulin and 10% fetal serum prepared by the method described in JP-A No. 109-191989. with MEM medium and suspended at 1 0 5 cells ZML, sown 1 0 0 L to 96 ⁇ El plate, carbon dioxide incubator base Isseki within one (5% carbon dioxide gas, 95% air) in The cells were cultured at 37 ° C for 5 days.
- BTC 76 2-76 residue type (N-terminal 1 residue, C-terminal 3 residue deleted type) Betacellulin structural gene, 76-residue type (C-terminal 3 residue deleted type) II
- BTC 76 [Example 4] Primer 3 (5′-CAGCATATGG GGAATTC CACCAGAAGTCCT) having an NdeI cleavage site and an initiation codon immediately upstream of the second glycine, and terminated after C-terminal valine. Primer 4 having a stop codon and a BamHI cleavage site (5, —Amplified with GGATCCC. The gene amplified by PCR was ligated to pCR2.1 vector using TA originalcloning kit (manufactured by Invitrogen).
- pCR2.1 BTC2-76 was digested with NdeI and BamHI, followed by agarose gel electrophoresis, and the approximately 230 bp 2-76 residue type cellulin structural gene was ligated with QI AGEN gelextraction kit. (Manufactured by Qiagen).
- the expression vector PTC II [Example 1] was cut with Nde I and Bam HI and subjected to agarose gel electrophoresis. Similarly, a band of about 4.6 kbp was recovered.
- C2-76 was cultured in LB medium (1% peptone, 0.5% yeast extract, 0.5% sodium chloride) 3 0111 containing 1 OmgZL of tetracycline at 30 ° for 16 hours.
- the obtained culture solution is used as a main fermentation medium (1.68% sodium monohydrogen phosphate, 0.3% sodium dihydrogen phosphate, 0.1% ammonium chloride, 0.05% sodium chloride, 0.012% sulfuric acid).
- ImM EDTA, ImM APMSF and 7 g of the cells obtained in Example 14 were added to 5 g of the cells.
- Betacellulin expression plasmid pTC II BTC 76 upstream of the 24th alanine Primer 5 (5'-1 GG) with an adjacent NdeI cleavage site and initiation codon followed by a termination codon and a BamHI cleavage site (5'-1 CAGC phosphorus) was amplified by PCR.
- the gene amplified by PCR was ligated to pCR2.1 vector using TA original clone kit (manufactured by Invitrogen) to produce pCR2.1 BTC 24_76.
- transformants having pCR2.1 BTC 24-76 were cultured, and pCR2 was prepared using QI Aprep 8 Miniprepkit (Qiagen). 1 BTC 24-76 was prepared.
- pCR2.1 BTC 24-76 was digested with NdeI and BamHI, and subjected to agarose gel electrophoresis.
- the approximately 160 bp 24--76 residue type cellulite structural gene was ligated with QIAGEN geletraction. It was collected using a kit (Qiagen).
- the expression vector pTCIII prepared in Example 1 was digested with NdeI and BamHI and subjected to agarose gel electrophoresis. Similarly, a band of about 4.6 kbp was recovered.
- the gene was introduced into Escherichia coli JM109, and transformed strains were selected with tetracycline resistance.
- the plasmid was recovered again from the strain and used as an expression plasmid pTC II BTC 24-76.
- This pTC II BTC 24—76 was introduced into E. coli MM294 (DE 3), and a transformant was selected based on tetracycline resistance, and a 24—76 residue cellulin-expressing strain MM294 (DE3) p TCIIBTC 24—76 I got Example 17
- LB medium containing 1 Omg / L tetracycline 1% Peptone, 0.5% yeast extract, 0.5% yeast extract) 5% sodium chloride
- the obtained culture solution was used as a main fermentation medium (1.68% sodium monohydrogen phosphate, 0.3% sodium dihydrogen phosphate, 0.1% ammonium chloride, 0.05% sodium chloride, 0.1% sodium chloride).
- the fractions containing 24-76 residues of cellulin were collected, diluted three-fold with 5 OmM sodium acetate buffer (pH 4.5), and then equilibrated with this buffer to form a TSKgel CM-5PW column ( 0.75 cm X 7.5 cm, Tosoh Corporation). TSKg e 1 adsorbed with 24--cell residue cellulite The CM-5 PW column was eluted with a linear gradient of sodium chloride from 0.3 ⁇ 1 to 0.5 M.
- the characteristics of the 2- to 76-residue cellulin obtained in Example 3 and the 24- to 4-residue cellulin obtained in Example 6 were determined as follows.
- the modified cellulinated cellulin and the 76-residue type cellulin prepared in Example 6 were combined with sample buffer (125 mM Tris-HC and 1% sodium dodecyl sulfate, 15% glycerol, 5% 2-mercaptoethanol, The mixture was suspended in 0.0005% bromphenol blue and electrophoresed on Multigel 15/25 (Daiichi Pure Chemicals). When the gel after electrophoresis was stained with Rabbit CBB KANTO (Kanto Chemical Co., Ltd.), almost all bands were single bands (Fig. 9).
- the modified Bayerin cellulin was subjected to gas phase hydrolysis with 6 N hydrochloric acid containing 4% thioglycolic acid at 110 ° C for 24 and 48 hours, and an amino acid analyzer (Hitachi L-1 8500A Amino Acid Ana 1 Amino acid composition was determined using yzer). As a result, all of the variants matched the amino acid composition predicted from the cDNA nucleotide sequence [Table 8, Table 9]. [Table 8] Analysis of amino acid composition of 2-76 residue betacellulin Number of residues per mole of amino acid Theoretical value
- Met 0.0.0 0 lie 2.02 then eu 0.0.0 0
- N-terminal amino acid sequence analysis was determined using a gas phase protein sequencer (Applied Biosystems Model 477A). As a result, all the variants were in agreement with the amino acid sequence predicted from the cDNA nucleotide sequence [Table 10, Table 11].
- the C-terminal amino acid was determined by a gas-phase hydrazine decomposition method (100 ° C, 6 hours) using an amino acid analyzer (Hitachi L-1800A Amino Acid AnaliZer). Palin was detected in all variants [Table 12, Table 13].
- A431 cells is an highly expressed strain. That is, using a Dulbecco's Modified Eagle's MEM (DMEM) medium containing 10% fetal serum, seed the A431 cells suspended in a volume of 105 cells to 100 mL each in a 96-well plate, and place in a CO2 incubator. (5% CO 2, 95% air) at 37 ° C for 2 days.
- DMEM Dulbecco's Modified Eagle's MEM
- the cells were washed three times with 200 L of binding medium, and the cells were lysed with 200 aL of 0.1 N aqueous sodium hydroxide solution containing 1% SDS. Cell lysate The mixture was transferred to a scintillation vial, 1 mL of scintillator A (Wako Pure Chemical Industries, Ltd.) was added, and the binding inhibitory activity for 125 I-EGF was measured using a scintillation counter.
- scintillation vial 1 mL of scintillator A (Wako Pure Chemical Industries, Ltd.) was added, and the binding inhibitory activity for 125 I-EGF was measured using a scintillation counter.
- PLAP-expressing AR42 J cells were prepared at various concentrations using 2-76 residue type cellulin prepared in Example 15 and 24-18 residue type cellulin prepared in Example 18 and Example 6 Contains the prepared Met-76 residue type cellulin, or the Met-80 residue type cellulin prepared by the method described in JP-A No. 10-191989 and 10% fetal serum. was suspended at 10 5 cells / mL using Dulbecco's modified Eagle's MEM medium, 1 00 L were seeded 96 Uerupuredo, a carbon dioxide gas incubator (5% carbon dioxide, 95% air) 5 days at 37 ° C for at The culture was continued for a while.
- 2-76 residue type cellulin prepared in Example 15 and 24-18 residue type cellulin prepared in Example 18 and Example 6 Contains the prepared Met-76 residue type cellulin, or the Met-80 residue type cellulin prepared by the method described in JP-A No. 10-191989 and 10% fetal serum. was suspended at 10 5 cells / mL using Dulbec
- 5'-primer PET-1 5 and 3'-primer BTC-1: 5'-AGGAGGGCGTCGAGGGGTTTTGCTCGGCCA- 3 '
- 5'-primer BTC-2 PCR was performed using 5'-TGG primer-BTC_3: 5'-TCTATGCGCACCCGTTCTCGGAGC ACTGTC-3 ', respectively.
- PCR was performed using the 5′-primer BT—95 h and the 3′-primer BT—94 h, and the hB TC 50
- a DNA fragment encoding a modified molecule A was obtained in which three amino acids (A sn, Pro, Ser) in the sequence of Her were inserted between Cys 3 and Cys 4 of the DNA sequence.
- NdeI and BamHI After digestion of this DNA fragment with NdeI and BamHI, it was incorporated into the NdeI-BamHI site of pET-3c to generate pTB1985.
- PCR was performed using the 'and 3' primers BT-94h to obtain a DNA fragment encoding the modified molecule B in which the N-terminal 7 residues of hBTC50 were substituted with the corresponding 5 residues of EGF. .
- This DNA fragment was digested with NdeI and BamHI, and then incorporated into the NdeI-BamHI site of pET3c to produce pTB1987.
- E. coli BL21 DE3 ZpLysSS (Novagen) for expression.
- Each E. coli recombinant was cultured at 37 ° C. in an LB medium containing 100 gZm1 ampicillin and 100 g / m1 chloramphenicol.
- Cretutunite 160 isopropyl ⁇ -D_thiogalactovyranoside was added to a final concentration of 0.4 mM, and the culture was continued at 37 ° C for 4 hours. After the culture, the cells were collected by centrifugation and stored at 180 ° C until the extraction operation.
- Example 24 Example 24
- the cells collected from 400 ml of the culture were suspended in 20 ml of 10 mM Tris-HC1 (H8.0), 10% sucrose, 1 OmM EDTA, frozen and thawed. The cells were left on ice for 30 minutes to completely lyse the cells. After sonication (10 sec, 3 times) under ice-cooling, the precipitate was collected again by centrifugation (900 rpm, 15 min, 4 ° C: Bee km an). This washing operation was repeated three times to prepare an inclusion body.
- the inclusion bodies obtained in 4-1) were suspended in 8 ml of 7 M guanidine-HC and 0.1 MT ris-CH 3 C ⁇ H (pH 8.0), suspended in ImM EDTA at 4 C for 1 hour.
- the recombinant protein was extracted by gentle stirring with a stirrer.
- reduced glutathione was added to a concentration of 0.1 M, the pH was adjusted to 8.4 with a NaOH solution, and nitrogen gas was blown into the extract to replace the air.
- the 20-fold volume l OmM T ris- CH 3 C_ ⁇ _OH (pH8. 0), 0.
- 3 H-thymidine [Amersham-Pharmacia Biotech] was added at 0.25 Ci / we 1 I.
- the cells were washed three times with PBS, and 100 L of 5% SDS was added. Added cells were lysed. The cell lysate was transferred to a scintillation vial, 1 mL of Scintillation A (Wako Pure Chemical Industries) was added, and the uptake of 3 H-thymidine into the cells was measured with a scintillation counter.
- hBTC 80 80-residue cellulin prepared by the method described in JP-A No. 10-191989, and 10%
- FITC Fluorescein Isothiocyanate
- Block Ace diluted with 10% Block Ace was added and reacted at room temperature for 40 minutes. After 0.1% Triton X-100 was added and left at room temperature for 5 minutes, it was washed three times with PBS and observed with a fluorescence microscope.
- the activity of promoting differentiation into 3 cells was also carried out using AR42J cells transformed with an alkaline phosphatase gene ligated as a repo sequence downstream of the insulin promoter.
- cells differentiated into i3 cells by overnight cellulin produce alkaline phosphatase, and by measuring the activity of this alkaline phosphatase, the activity of promoting differentiation into three cells is quantitatively measured. It is possible to do so.
- Genomic DNA was prepared from the rat tail according to a conventional method. Using this DNA as a template, primers RI-1: 5'-AGAGTCAAGGATCCCCCAACCACT-3 'and primers synthesized based on the previously reported base sequence of rat insulin II gene promoter (GenBank: Accession No.] 00748) One
- Plasmid pTB1881 was digested with XhoI-C1aI to obtain a 0.73 kb DNA fragment which was rat insulin promoter overnight.
- the expression plasmid pTB1330 of 2.0 kbc DNA encoding human placental alkaline phosphatase (PLAP) J.
- the culture supernatant was collected and heat-treated at 65 ° C for 30 minutes, and then the alkaline phosphatase activity in the medium was measured.
- a clone showing an increase in alkaline phosphatase activity upon addition of 80-residue cellulin was selected, and one of the clones, AR71-104, was used in the following studies.
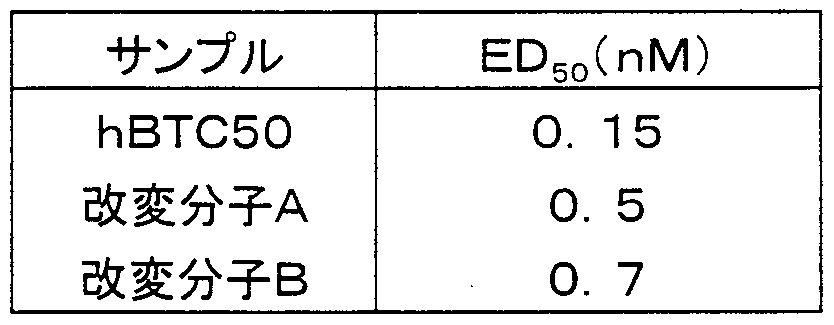
- the protein concentration (ED 50 ) required to show 50% activity when the maximum absorbance when hBTC 50 is added is 100% is shown in [Table 16].
- primer BT—95h: 5′- AGCAT PCR was performed using '.
- the overnight cellulinmutin or a salt thereof according to the present invention is useful as an excellent therapeutic agent for diabetes, since EGF activity is attenuated while BTC activity is maintained and there is no problem of antigenicity.
Landscapes
- Health & Medical Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Gastroenterology & Hepatology (AREA)
- General Health & Medical Sciences (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biophysics (AREA)
- Zoology (AREA)
- Genetics & Genomics (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biochemistry (AREA)
- Toxicology (AREA)
- Medicines That Contain Protein Lipid Enzymes And Other Medicines (AREA)
- Peptides Or Proteins (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Description





Claims
Priority Applications (6)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| DE69936666T DE69936666T2 (de) | 1998-12-09 | 1999-12-08 | Betacellulin-veränderung |
| CA002353909A CA2353909C (en) | 1998-12-09 | 1999-12-08 | Betacellulin muteins |
| AU16811/00A AU1681100A (en) | 1998-12-09 | 1999-12-08 | Betacellulin modification |
| DK99959698T DK1148129T3 (da) | 1998-12-09 | 1999-12-08 | Modifikation af betacellulin |
| EP99959698A EP1148129B1 (en) | 1998-12-09 | 1999-12-08 | Betacellulin modification |
| US09/857,815 US6825165B1 (en) | 1998-12-09 | 1999-12-08 | Betacellulin modification |
Applications Claiming Priority (4)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP10/350377 | 1998-12-09 | ||
| JP35037798 | 1998-12-09 | ||
| JP5532699 | 1999-03-03 | ||
| JP11/55326 | 1999-03-03 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| WO2000034478A1 true WO2000034478A1 (fr) | 2000-06-15 |
Family
ID=26396224
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| PCT/JP1999/006873 Ceased WO2000034478A1 (fr) | 1998-12-09 | 1999-12-08 | Modification de betacelluline |
Country Status (11)
| Country | Link |
|---|---|
| US (1) | US6825165B1 (ja) |
| EP (1) | EP1148129B1 (ja) |
| AT (1) | ATE368111T1 (ja) |
| AU (1) | AU1681100A (ja) |
| CA (1) | CA2353909C (ja) |
| CY (1) | CY1106882T1 (ja) |
| DE (1) | DE69936666T2 (ja) |
| DK (1) | DK1148129T3 (ja) |
| ES (1) | ES2288034T3 (ja) |
| PT (1) | PT1148129E (ja) |
| WO (1) | WO2000034478A1 (ja) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2006012707A1 (en) * | 2004-08-06 | 2006-02-09 | Gropep Limited | Method for treating diabetes |
| US20070054851A1 (en) * | 2005-05-27 | 2007-03-08 | Junyu Lin | Methods of and compositions for stimulation of glucose uptake into muscle cells and treatment of diseases |
| TW202216761A (zh) | 2020-07-16 | 2022-05-01 | 瑞士商諾華公司 | 抗β細胞素抗體、其片段及多特異性結合分子 |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH0687894A (ja) * | 1992-02-10 | 1994-03-29 | Takeda Chem Ind Ltd | 平滑筋細胞増殖因子およびそれをコードする単離されたdna |
| JPH10191989A (ja) * | 1996-11-12 | 1998-07-28 | Takeda Chem Ind Ltd | ベータセルリン類の製造方法 |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6410506B1 (en) * | 1995-05-19 | 2002-06-25 | Human Genome Sciences, Inc. | Transforming growth factor α HII |
| US6232288B1 (en) | 1995-11-09 | 2001-05-15 | Takeda Chemical Industries, Ltd. | Composition for improving pancreatic function |
-
1999
- 1999-12-08 PT PT99959698T patent/PT1148129E/pt unknown
- 1999-12-08 AU AU16811/00A patent/AU1681100A/en not_active Abandoned
- 1999-12-08 US US09/857,815 patent/US6825165B1/en not_active Expired - Fee Related
- 1999-12-08 CA CA002353909A patent/CA2353909C/en not_active Expired - Fee Related
- 1999-12-08 WO PCT/JP1999/006873 patent/WO2000034478A1/ja not_active Ceased
- 1999-12-08 DK DK99959698T patent/DK1148129T3/da active
- 1999-12-08 ES ES99959698T patent/ES2288034T3/es not_active Expired - Lifetime
- 1999-12-08 DE DE69936666T patent/DE69936666T2/de not_active Expired - Lifetime
- 1999-12-08 AT AT99959698T patent/ATE368111T1/de not_active IP Right Cessation
- 1999-12-08 EP EP99959698A patent/EP1148129B1/en not_active Expired - Lifetime
-
2007
- 2007-09-19 CY CY20071101216T patent/CY1106882T1/el unknown
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JPH0687894A (ja) * | 1992-02-10 | 1994-03-29 | Takeda Chem Ind Ltd | 平滑筋細胞増殖因子およびそれをコードする単離されたdna |
| JPH10191989A (ja) * | 1996-11-12 | 1998-07-28 | Takeda Chem Ind Ltd | ベータセルリン類の製造方法 |
Non-Patent Citations (10)
| Title |
|---|
| COOK P.W. ET AL.: "Carboxyl-terminal truncation of leucine 76 converts heparin-binding EGF-like-growth factor from a heparin-enhancible to a heparin-suppressible growth factor", J. CELL. PHYSIOL.,, vol. 163, no. 2, 1995, pages 407 - 417, XP002924355 * |
| JOHNSON G.R. ET AL.: "Characterization of high and low molecular weight forms of amphiregulin that differ in glycosylation and peptide core length. Evidence that the NH2-terminal region is not critical for bioactivity", J. BIOL. CHEM.,, vol. 268, no. 25, 1993, pages 18835 - 18843, XP002924356 * |
| K. MIYASONO ET AL.: "Jikken Igaku Bessatsu, BioScience, Yogo Library Saitikain, Zoshoku Inshi", KABUSHIKI KAISHA YODOSHA,, 1 May 1997 (1997-05-01), pages 92 - 93, XP002929902 * |
| KOJIMA I. ET AL.: "Differentiation of pancreatic endocrine cells", DIABETES J.,, vol. 26, no. 3, September 1998 (1998-09-01), pages 97 - 103, XP002924357 * |
| MIXAN B. ET AL.: "Betacellulin-Pseudomonas toxin fusion proteins bind but are not cytotoxic to cells expressing HER4; correletion of EGFR for cytotoxic activity", ONCOGENE,, vol. 16, no. 9, March 1998 (1998-03-01), pages 1209 - 1215, XP002924360 * |
| REIKO SASADA ET AL.: "Betacellulin: Atarashii Kekkan Heikatsukin Saibo Seicho Inshi (Betacellulin: a new growth factor for vascular smooth muscle cells)", NIPPON RINSHO,, vol. 51, no. 12, 1993, pages 3308 - 3317, XP002929901 * |
| SASADA R. ET AL.: "Cloning and expression of cDNA encoding human betacellulin, a new member of the EGF family", BIOCHEM. BIOPHYS. RES. COMMUN.,, vol. 190, no. 3, 1993, pages 1173 - 1179, XP002924353 * |
| SHIN S.Y. ET AL.: "The chemical synthesis and biological activity of EGF-like domain of betacellulin, a new member of EGF family", PEPTIDE CHEMISTRY,, vol. 1993, 1994, pages 225 - 228, XP002924359 * |
| SHING Y. ET AL.: "Betacellulin: a mitogen from pancreatic beta cell tumors", SCIENCE,, vol. 259, no. 5101, 1993, pages 1604 - 1607, XP002924354 * |
| WATANABE T. ET AL.: "Recombinant Human Betacellulin", J. BIOL. CHEM.,, vol. 269, no. 13, 1994, pages 9966 - 9973, XP002924358 * |
Also Published As
| Publication number | Publication date |
|---|---|
| PT1148129E (pt) | 2007-09-05 |
| EP1148129B1 (en) | 2007-07-25 |
| US6825165B1 (en) | 2004-11-30 |
| DE69936666T2 (de) | 2008-05-21 |
| CA2353909C (en) | 2009-07-07 |
| DK1148129T3 (da) | 2007-10-15 |
| CA2353909A1 (en) | 2000-06-15 |
| DE69936666D1 (de) | 2007-09-06 |
| EP1148129A1 (en) | 2001-10-24 |
| EP1148129A4 (en) | 2005-06-22 |
| CY1106882T1 (el) | 2012-09-26 |
| AU1681100A (en) | 2000-06-26 |
| ATE368111T1 (de) | 2007-08-15 |
| ES2288034T3 (es) | 2007-12-16 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3045398B2 (ja) | 蛋白質、dnaおよびその用途 | |
| US6025467A (en) | Parathyroid hormone derivatives and their use | |
| EP0477885B1 (en) | Parathyroid hormone derivatives | |
| US6020163A (en) | Lipocalin homolog | |
| US20090111742A1 (en) | Use of fgf-21 and thiazolidinedione for treating type 2 diabetes | |
| US5622932A (en) | IGF-1 superagonists | |
| CN110546161A (zh) | 对胰岛素受体具有降低的结合力的胰岛素类似物及其用途 | |
| KR20200140878A (ko) | 성장 분화 인자 15 융합 단백질 | |
| WO2010016069A1 (en) | A process for preparation of insulin compounds | |
| EP3137494A2 (en) | Islet amyloid polypeptides with improved solubility | |
| WO2000034478A1 (fr) | Modification de betacelluline | |
| RU2344172C2 (ru) | Мутеины плацентарного фактора роста 1 типа, способ их получения и применение | |
| WO1998022505A1 (fr) | Polypeptides solubles | |
| WO2001089582A1 (en) | Preventives and remedies for pulmonary hypertension | |
| US7132397B1 (en) | Bone morphogenetic protein antagonist based on the mature protein | |
| JP4512222B2 (ja) | ベータセルリン改変体 | |
| US5670342A (en) | NDF peptides | |
| CN113527506A (zh) | 融合蛋白及其应用 | |
| HUT61339A (en) | Process for producing des(64,64)-proinsulin | |
| JPWO2000034478A1 (ja) | ベータセルリン改変体 | |
| US20110237509A1 (en) | hGH AND METHODS FOR PREPARING hGH | |
| KR102646845B1 (ko) | 클로스트리파인을 이용한 지속형 인슐린 아날로그 복합체의 활성형 제조방법 | |
| JPH09503646A (ja) | アンチトロンビンポリペプチド類似体及びその製造法 | |
| EP1142908A1 (en) | Process for producing activated protein | |
| JP2003033189A (ja) | Zaqリガンドの製造方法 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| ENP | Entry into the national phase |
Ref country code: AU Ref document number: 2000 16811 Kind code of ref document: A Format of ref document f/p: F |
|
| AK | Designated states |
Kind code of ref document: A1 Designated state(s): AE AL AM AU AZ BA BB BG BR BY CA CN CR CU CZ DM EE GD GE HR HU ID IL IN IS JP KG KR KZ LC LK LR LT LV MA MD MG MK MN MX NO NZ PL RO RU SG SI SK SL TJ TM TR TT TZ UA US UZ VN YU ZA |
|
| AL | Designated countries for regional patents |
Kind code of ref document: A1 Designated state(s): GH GM KE LS MW SD SL SZ TZ UG ZW AM AZ BY KG KZ MD RU TJ TM AT BE CH CY DE DK ES FI FR GB GR IE IT LU MC NL PT SE BF BJ CF CG CI CM GA GN GW ML MR NE SN TD TG |
|
| DFPE | Request for preliminary examination filed prior to expiration of 19th month from priority date (pct application filed before 20040101) | ||
| 121 | Ep: the epo has been informed by wipo that ep was designated in this application | ||
| WWE | Wipo information: entry into national phase |
Ref document number: 1999959698 Country of ref document: EP |
|
| ENP | Entry into the national phase |
Ref document number: 2353909 Country of ref document: CA Ref country code: CA Ref document number: 2353909 Kind code of ref document: A Format of ref document f/p: F |
|
| WWE | Wipo information: entry into national phase |
Ref document number: 09857815 Country of ref document: US |
|
| WWP | Wipo information: published in national office |
Ref document number: 1999959698 Country of ref document: EP |
|
| NENP | Non-entry into the national phase |
Ref country code: JP Ref document number: 2000586912 Format of ref document f/p: F |
|
| WWG | Wipo information: grant in national office |
Ref document number: 1999959698 Country of ref document: EP |