-
GEBIET DER
ERFINDUNG
-
Diese
Erfindung betrifft Verfahren zur Isolation von Antagonisten und
Agonisten von Cytokinen.
-
HINTERGRUND
DER ERFINDUNG
-
Der
humane Granulocyten-Makrophagen-Kolonie stimulierende Faktor (GM-CSF),
die Interleukine IL-3 und IL-5 sind Cytokine, die beim hämopoietischen
Zellüberleben,
bei der Herstellung und Funktion (als Übersicht in Lopez et al., 1992)
beteiligt sind. Wegen dieser Eigenschaften werden IL-3 und GM-CSF
gegenwärtig
klinisch für
die Knochenmarkserneuerung nach einer Chemotherapie und Radiotherapie
(Groopman et al., 1987) verwendet. Allerdings ist ebenfalls offensichtlich
geworden, dass eine übermäßige oder
gegen die Regeln verstoßende
Produktion von GM-CSF, IL-3 und IL-5 zu Krankheitszuständen führen kann.
Beispielsweise sind erhöhte
Mengen an GM-CSF in den Lungen von allergischen Individuen (Kato
et al., 1992) und in den Gelenken von Patienten mit rheumatoider
Arthritis (Williamson et al., 1988) gefunden worden. Erhöhte mRNA
für IL-3,
GM-CSF und IL-5 sind in der Haut von allergischen Individuen (Kay
et al., 1991) gefunden worden. GM-CSF kann die Proliferation von
Leukämiezellen
(Young et al., 1986) stimulieren, und es ist gezeigt worden, dass
IL-3 durch Follikel-B-Zellen-Lymphomas in autokriner Weise produziert wird,
was zu einer Proliferation dieser Zellen in einer IL-3-abhängigen Weise
(Clayberger et al., 1991) führt.
-
Es
ist aus diesen klinischen Situationen deutlich, dass die Antagonisierung
von GM-CSF, IL-3 und IL-5 von therapeutischem Wert sein kann, in
Abhängigkeit
der in Frage stehenden Erkrankung, wobei die Antagonisierung eines
dieser Cytokine ausreichend sein kann.
-
Eine
Zahl von Vorschlägen
für Antagonisten
ist bereits gemacht worden, beispielsweise in der Spezifikation
WO 89 10403, und in der Spezifikation WO 95 04075 sind Varianten
von GPM-CSF als Antagonisten zur GM-CSF-Wirkung identifiziert, allerdings
gibt es keinen Hinweis darauf, dass diese Antagonisten für mehr als
nur die GM-CSF-Wirkung effektiv sind.
-
Allerdings
kann in anderen Situationen der simultane Antagonismus von allen
drei GM-CSF, IL-3 und IL-5 erwünscht
oder tatsächlich
notwendig sein. Beispielsweise können
Eosinophile, die der Hauptzelltyp, der bei Allergien beteiligt ist,
sein sollen, zahlenmäßig erhalten
werden und durch entweder IL-3, GM-CSF oder IL-5 (Lopez et al.,
1989) stimuliert werden. Der Antagonismus aller drei Cytokine kann
daher notwendig sein, um die Wirkungen von Eosinophilen und Basophilen
zu inhibieren. In ähnlicher
Weise können
Basophile, von denen man ebenfalls annimmt, dass sie eine Effektorrolle
bei der Allergie spielen, durch entweder IL-3, GM-CSF oder IL-5 (Lopez et al.,
1990) stimuliert werden. Der Antagonismus von GM-CSF, IL-3 und IL-3
kann durch die gleichzeitige Verabreichung spezifischer Antagonisten
für jedes
verschiedene Cytokin bewerkstelligt werden. Obwohl dieses machbar
ist, hat dieser Lösungsansatz
den Nachteil, dass man bis zu drei verschiedene Proteine verabreichen
muss, was nicht nur unbequem ist, sondern allerdings auch das Risiko
der Immogenizität
und anderen Nebeneffekten erhöht.
-
Ein
Krankheitszustand, der hier herausragt und der durch erhöhte Gehalte
dieser drei Cytokine bekämpft
werden kann, ist das Asthma. Die Rollen vom GM-CSF, IL-3 und IL-5
bei Asthma und Allergien sind bereits intensiv untersucht worden
und werden weiterhin intensiv untersucht. Verschiedene Studien,
durch in situ Hybridisierung, haben erhöhte Gehalte von IL-5-mRNA in mononuklearen
Lungenzellen (Fukuda et al., 1994; Robinson et al., 1992; Marini
et al., 1992; Hamid et al., 1991) und in Eosinophilen (Broide et
al., 1992) bei astmatischen Patienten gezeigt. Die Immunchemie hat
ebenfalls erhöhte
Mengen an IL-5 Protein in diesen Geweben (Ackerman et al., 1994)
gezeigt. Bei Allergen induzierten Hautreaktionen in der späten Phase
bei atopischen Individuen, ist erhöhte mRNA für IL-5, IL-3 und GM-CSF verzeichnet
worden (Kay et al., 1991).
-
Bronchoalveolare
Spüllflüssigkeiten
(BAL) von symptomatischen Asthmapatienten zeigten größere IL-5-Gehalte
als Patienten mit symptomatischen Asthma (Sur et al., 1995). Wenn
weiterhin symptomatische Asthmapatienten mit einem Antigen in einem
Lungensegment angegriffen wurden, verzeichnete man signifikante
Gehalte von IL-5 (980 pg/ml) im Vergleich mit einem fingiert angegriffenen
Segment des gleichen Patienten (2,8 pg/ml) (Sur et al., 1995). In
einer anderen Studie mit ähnlichem
Muster erhöhten
sich die IL-5-Gehalte von nicht nachweisbar bis 2.800 pg/ml bei
sowohl allergischen als auch nicht allergischen Asthmatikern (Zangrilli
et al., 1995). Ein kausaler Zusammenhang zwischen den IL-5-Mengen
und Asthma leitet sich aus Studien her, wobei die Behandlung von
mittelschwerkranken asthmatischen Patienten mit Corticosteroiden
für zwei
Wochen zu einer Reduktion der Anzahl der Zellen, die IL-5-mRNA (Bentley
et al., 1996) exprimieren, führte.
Diese Reduktion korrelierte mit der klinischen Verbesserung und
ebenfalls mit dem Verlust von CD3+-T-Zellen
und aktivierten (EG2+)-Eosinophilen.
-
GM-CSF
ist ebenfalls in den Lungen von Asthmatikern nachgewiesen worden.
Tatsächlich
schien in einer Studie von Sputum-Cytokinen GM-CSF einen dominanten
Effekt auf das Überleben
von Eosinophilen zu haben (Adachi et al., 1995).
-
IL-5
aktiviert ebenfalls Eosinophile, um das EG2-Epitop zu exprimieren.
In einigen Studien waren erhöhte
IL-5-Gehalte mit EG2+-Eosinophilen in der
Lunge assoziiert (Fukuda et al., 1994; Bentley et al., 1996). Des
Weiteren gibt es eine Evidenz der gewebespezifischen Aktivierung
von Eosinophilen. In einem Experiment wurde die Aktivierung von
Eosinophilen in der Lunge mit denjenigen im Blut des gleichen Patienten
verglichen. Die Aktivierung wurde durch die Zelloberflächenexpression
von CR-3, p150/95,
CD67, CD63 und den Verlust von L-Selektin festgestellt. Bei Patienten,
die mit dem endobronchialen Antigen angegriffen wurden, wurden sowohl
periphere Eosinophile als auch Lungeneosinophile bei 24 h gesehen,
jedoch haben nur die Lungeneosinophile erhöhte Werte von GM-CSF-mRNA, was eine
lokale Aktivierung dieser Zellen nahe legt. Außerdem gab es eine Evidenz
für die
Aktivierung von Eosinophilen in der Lunge, allerdings nicht von
denen aus Blut. Dieses Ergebnis stimmt mit einem spezifischen Effekt
von IL-5 auf Lungen auf Eosinophile und ihre Beteiligung bei Asthma überein.
-
Tiermodelle
legen ebenfalls eine Rolle für
IL-5 bei Asthma nahe. Die am meisten signifikanten Daten gibt es
mit einem Antikörper
(TRFK-5), gegen IL-5 in Affenmodellen von Asthma (Mauser et al.,
1995). Ascaris-empfindliche Cynomolgusaffen wurden mit aerosolisierten
Ascaris suum Extrakten angegriffen. 0,3 mg/kg TRFK beseitigte die
erhöhte
Luftwegreaktivität
und verminderte die Zahl der Eosinophile in den BAL-Flüssigkeiten.
Interessanterweise hielt diese Inhibierung für 3 Monate an (Mauser et al.,
1995). Studien mit dem Guineaschwein haben diese Schlussfolgerung
unterstützt
(Mauser et al., 1993). Weiterhin hatten Mäuse, bei denen das IL-5-Gen
genetisch entfernt war, nicht nur nicht nachweisbares IL-5 und eine
signifikante Reduktion der Anzahl der Eosinophile, sondern sie entwickelten
ebenfalls signifikant weniger schweres Asthma (wie durch Luftwegehyperreaktivität und Lungenschädigung festgestellt
wurde) als IL-5-Gen positive Artgenossen (Foster et al., 1996).
Ein deutliches Beispiel für
die Wiederherstellung von Luftwegereaktionen gegenüber Methacholin
wurde verzeichnet, nachdem IL-5-defiziente Mäuse IL-5-exprimierenden, allerdings
nicht kontrollierenden, Vaccinvirusinfektionen ausgesetzt wurden
(Foster et al., 1996).
-
Eine
mögliche
Rolle für
das GM-SCF beim Asthma ist ebenfalls durch die Überexpression von GM-CSF in
der Rattenlunge, was zu Eosinophilie, Makrophagengranulomae und
fibrotischer Reaktion führt, ein
Dreierelement, das ebenfalls bei Asthma verzeichnet wird, nahegelegt
worden (Xing et al., 1996).
-
Humanes
Interleukin (IL)-3, Granulocyten-Makrophagen-Kolonie stimulierender
Faktor (GM-CSF) und IL-5 üben
ihren biologischen Effekt dadurch aus, dass sie an spezifische Oberflächenrezeptoren
auf der Oberfläche
von Zellen (Bagley et al., 1995; Elliott et al., 1989; Park et al.,
1989; Lopez et al., 1991) binden. Die Rezeptoren sind Heterodimere,
die eine α-Kette,
die für
jeden Liganden spezifisch ist, und eine β-Kette (βc),
die zwischen den drei Rezeptoren geteilt wird, umfasst (Lopez, et
al., 1992), Kitamura et al., 1991). Während jeder Ligand an die jeweilige α-Kette bindet,
gibt es für βc eine
höhere
Affinität,
was eine Signalgebung ermöglicht (Miyajima
et al., 1992 und US-Patent 5112961 von Hayashida et al.). Die Erfinder
des US-Patents Nr. 5112961 legen nahe, dass der Rezeptor hoher Affinität ein wertvolles
Werkzeug dafür
ist, Kandidaten für
GM-CF-Agonisten und Antagonisten zu screenen.
-
Weil
alle drei dieser Cytokine über
eine gemeinsame Rezeptoruntereinheit (βc) agieren,
was wir hierzu vor als Hypothese aufgestellt haben (Bagley et al.,
1995 diese Referenz ist hiermit eingeschlossen), kann es möglich sein,
die Wirkung von GM-CSF, IL-3 und IL-5 mit einer einzigen Verbindung
zu inhibieren. Es gibt allerdings bisher noch kein Mittel, sich
dem Problem des Findens eines Effektors anzunähern, der in koordinativer
Weise auf alle drei Cytonkine wirkt. Es ist beispielsweise nicht
klar, welche Region der gemeinsamen Rezeptoruntereinheit (βc)
durch diese Cytokine gebunden wird und ob die gleiche Region für alle drei
Cytokine bindet oder ob drei verschiedene Regionen beteiligt sind.
-
Die
Analyse von βc hat gezeigt, dass sie vier extrazelluläre Domänen (Goodall
et al., 1993) enthält,
mit einer Domäne
1, die das meiste der N-terminalen Domäne darstellt und einer Domäne 4, die
das meiste der membrannahen Domäne repräsentiert.
Jede Domäne
besteht aus sieben β-Strängen, die
zusammen durch ineinander verschlungenen Schleifen verknüpft sind.
Die hier interessierende Schleife, die F'-G'-Schleife
befindet sich in der Domäne
4 von βc. Die hier interessierenden Reste werden
vom N-Terminus am Primärtranslationsterminus
mit dem Initiationsmethionin, das MetI ist,
nummeriert.
-
Die
molekulare Basis für
die Affinitätsumwandlung
von βc an jeden Liganden ist noch nicht vollständig verstanden,
weil der Liganden-Rezeptor-Komplex noch nicht kristallisiert worden
ist. Allerdings ist es wahrscheinlich, durch Analogie zur Wechselwirkung
des Wachstumshormons (GH) mit seinem homodimären Rezeptor (DeVos et al.,
1992), dass es eine direkte Wechselwirkung zwischen dem Liganden
und den Rezeptor- α- und β-Ketten gibt.
In dem GH : GH-Rezeptorsystem sind die Kontaktpunkte zwischen GH
und GHbp2 aus der Röntgenstrahlkristallographie
des GH : GHhp1 und 2-Komplexes (DeVos et al., 1992) bestimmt worden. Durch
Analogie mit diesem System haben wir (Woodcock et al., 1994) und
andere (Lock et al., 1994) kürzlich gezeigt,
dass die B'-C'-Schleife von βc eine
gewisse Rolle bei der hohen Affinitätsbindung von GM-CSF, IL-3 und
IL-5 spielt. Insbesondere
ist gezeigt worden, dass drei Reste, Tyr365,
His367 und Ile368 wichtig
für die GM-CSF-
und IL-5-Hochaffinitätsbindung
sind, wobei sie allerdings nur marginal an der IL-3-Hochaffinitätsbindung
beteiligt sind (Woodcock et al., 1994). Diese Ergebnisse implizieren,
dass ein gezieltes Zusammenspiel dieser drei Aminosäuren mit
geeigneten Verbindungen die GM-CSF- und IL-5-vermittelte Aktivität, allerdings nicht
die IL-3-vermittelte Aktivität,
beeinflussen kann.
-
Zusammenfassung
der Erfindung
-
Unter
Verwendung molekularer Gestaltungstechniken haben wir kürzlich postuliert,
dass die F'-G'-Schleife bei der
Ligandenbindung (Bagley et al., 1995) beteiligt sein kann. Die Erfindung
ist aus einer Untersuchung über
diese vorausgesagte F'-G'-Schleife von βc und
aus der Feststellung, dass diese Schleife wesentlich für die Hochaffinitätsbindung
und für
die Signalgebung aller drei Cytokine GM-CSF, IL-3 und Il-5 ist, entstanden.
-
Ein
weiterer Aspekt dieser Erfindung ist als Ergebnis der überraschenden
Identifikation einer einzelnen Aminosäure in der Rezeptor-βc-Kette
für GM-CSF,
IL-3 und IL-5 entstanden, die notwendig für die Hochaffinitätsbindung
aller dieser drei Liganden an ihre Oberflächenrezeptoren und für die Signalgebung
und für
die Tatsache, dass alle drei dieser Cytokine die F'-G'-Schleife für die Hochaffinitätsumwandlung
benötigen,
ist. Es wird angenommen, dass Verbindungen, die an Tyr421 binden
oder die Bindung des Liganden an Tyr421 inhibieren,
sich wie generische Antagonisten von IL-3, GM-CSF und IL-5 verhalten.
Es wird angenommen, dass Verbindungen, die an F'-G'-Schleife
binden, sterisch diese Bindung inhibieren.
-
Zusätzlich haben
möglicherweise
diese Feststellungen auch Einflüsse
auf andere Mitglieder der Cytokinrezeptorsuperfamilie, einige davon
teilen sich Untereinheiten in einer gegeben Subfamilie (das heißt, sie binden
einige Cytokine), und einige davon sind Liganden spezifisch und
binden an nur ein Cytokin. Die Rezeptor-α-Ketten für GM-CSF, IL-3 und IL-5 und βc gehören zu der
sich schnell vergrößernden
Cytokinrezeptorsuperfamilie. Innerhalb dieser Superfamilie haben
sich mittlerweile verschiedene Subfamilien hervorgetan, die sich
durch das Teilen einer gemeinsamen Rezeptoruntereinheit durch multiple
Liganden auszeichnen: gp130 wirkt als Affinitätsumwandler und Signalumwandler
für IL-6
(Hibi et al., 1990; Taga et al., 1992), IL-11 (Hilton et al., 1994),
Oncostatin M (Liu et al., 1992), ziliarer neutrophischer Faktor,
Leukämieinhibierungsfaktor
(LIF) (Ip et al., 1992) und Cardiotrophin-1 (Pennica et al., 1995);
der LIF-Rezeptor (LIFR) bindet ebenfalls an den ziliaren neutrophischen
Faktor (Davis et al., 1993), Cardiotrophin-1 (Pennica et al., 1995)
und Oncostatin M zusätzlich
zu LIF (Gearing et al., 1994); IL-2R β unterstützt die
Affinitätsumwandlung
und die Signalgebung von IL-2 und IL-15 (Giri et al., 1994); die
IL-2R γ-Kettenaffinität wandelt
IL-2 (Takeshita et al., 1992), IL-4 (Russell et al., 1993), IL-7
(Noguchi et al., 1993), IL-9 (Kimura et al., 1995) und IL-15 (Giri
et al., 1994) um; die Evidenz legt ebenfalls nahe, dass IL-4 und
IL-13 eine Rezeptorkomponente teilen (Zurawski et al., 1993) und
diese Untereinheit ist erst kürzlich
kloniert worden (Hilton et al., 1996). Es ist nicht bekannt, welche
Reste in gp130, LIFR und den IL-2R β- und γ-Ketten für die Ligandenbindung wichtig
sind oder welche verschiedenen Liganden tatsächlich einmalige Sets von Bindungsdeterminanten
auf diesen gemeinsamen Rezeptoruntereinheiten teilen oder besitzen.
Da diese gemeinsamen Untereinheiten für die Übertragung von Signalen durch verschiedene
Liganden wichtig sind, tritt die Möglichkeit zutage, dass ein
Eingreifen in die Fähigkeit
dieser gemeinsamen Untereinheiten, den Liganden zu binden oder Homodimere
zu bilden, die Aktion von mehr als einem Liganden beeinträchtigen
kann.
-
Ein
Vergleich der vorhergesagten F'-G'-Schleife von βc mit
derjenigen von verschiedenen herkömmlichen Untereinheiten und
Liganden spezifischen Rezeptoruntereinheiten haben die Gegenwart
von Tyrosin und anderen ähnlichen
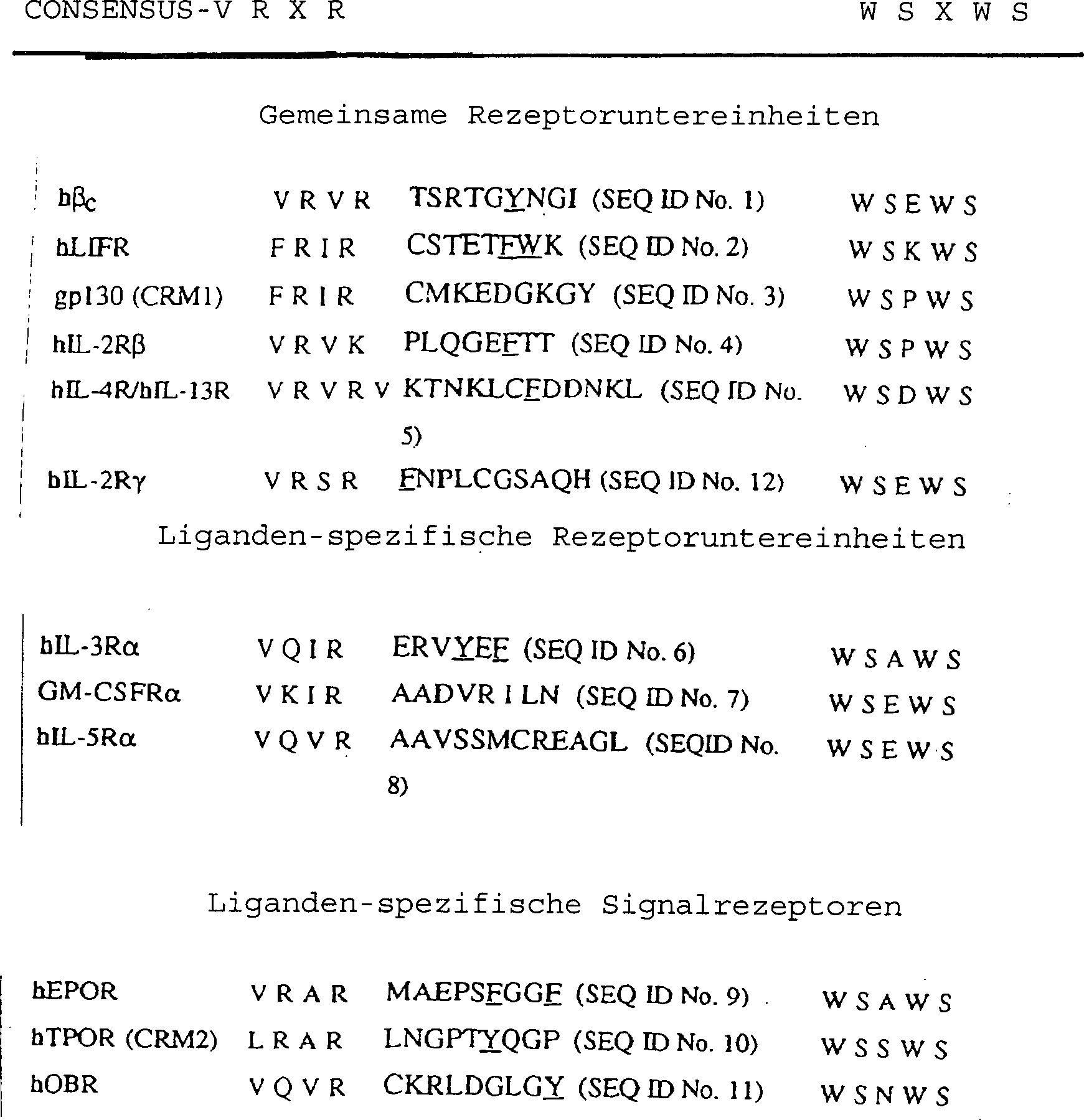
hydrophoben Resten in diesem Bereich (Tabelle 3) gezeigt. Obwohl
die genaue Länge
dieser putativen Schleife zwischen 7 bis 14 Aminosäuren unter
den verschiedenen Rezeptoren variiert, ist sie auf 2 streng konservierte
Sequenzen eingegrenzt; der VRVR-Konsensus am N-Terminus und der WSXWS-Konsensus am
C-Terminus. Es ist nahegelegt, dass, obwohl die Natur der Liganden-Rezeptor-Wechselwirkung spezifisch
für das
beteiligte System ist, die hydrophoben aromatischen Reste im Rezeptor
starke Kandidaten für
die Ligandenwechselwirkung darstellen. Daher können Mitglieder der Cytokinrezeptorfamilie
ein strukturell konserviertes Rezeptorgerüst verwenden, um eine Serie
von Schleifen auszubilden, die durch vornehmlich hydrophobe Wechselwirkungen
die Bindung der spezifischen Liganden unterstützen. Eine Unterstützung für diese
Idee stammt ebenfalls aus der Mutationsanalyse von IL-6 R, die gezeigt
hat, dass eine Mutante, bei der sowohl Glu297 als
auch Phe298 in der F'-G'-Schleife
substituiert waren, einen Verlust der IL-6-Bindung zeigte (Yawata et al., 1993).
-
Eine
neuere Analyse der Reste, die bei der Bindung des Wachstumshormons
in GHR beteiligt sind, hat gezeigt, dass der größte Beitrag zur Affinität von hydrophoben
Wechselwirkungen kommt (Clackson and Wells, 1995). Die systematische
Alaninsubstitution der 33-Reste in GHR, die bekannterweise einer
Zersetzung bei der Wachstumshormonbindung nicht zugänglich sind,
zeigte, dass die Substitution von nur 11 dieser Reste einen signifikanten
Effekt auf die Affinität
hatte, und davon waren sechs hydrophob. Des Weiteren sind diese hydrophoben
Reste auf der Oberfläche
von GHR in einer Region geclustert, die die Grenzfläche für die Wechselwirkung
mit dem Wachstumshormon bildet. Außerdem trat bei GHR die größte Affinitätsreduktion
bei der Substitution von zwei Tryptophanresten auf (Clackson and
Wells, 1995; Bass et al., 1991), was impliziert, dass hydrophopbe
aromatische Reste sehr wichtig für
die Ligandenwechselwirkung sind. Aufgrund unserer vorliegenden Studie
und früheren
Arbeiten (Woodstock et al., 1994) haben wir nun eine Gesamtheit
von drei hydrophoben Resten in vorhergesagten Schleifen identifiziert,
die eine Rolle bei der Ligandenwechselwirkung in βc spielen,
Tyr365, Ile368 und
Typr421. Daher scheint es, dass die Bindung
von βc an GM-CSF, IL-3 und Il-5 ebenfalls vornehmlich
durch die hydrophobe Wechselwirkung unterstützt wird.
-
Es
ist interessant, diese Erkenntnisse im Hinblick auf βc der
GM-CSF-, IL-3- und IL-5-Rezeptoren mit der Kristallstruktur- und Mutationsanalyse
von GH-GHR zu vergleichen. Im Gegensatz zu heterodimeren Natur der
GM-CSF-, IL-3- und IL-5-Rezeptoren ist GHR ein Homodimer. Die beiden
identischen Untereinheiten von GHR binden das Wachstumshormon nacheinander
und auf nicht äquivalente
Weise. In der Kristallstruktur werden die beiden Wachstumshormonbindungsproteine
entsprechend als GHbpI und GHbpII identifiziert. Die Anfangswechselwirkung
von GHbpI nutzt die Kontakte der Stellen I im Wachstumshormon und
ist relativ stark mit einer Dissoziationskonstante von 600 pM (Cunningham
et al., 1991). Die nachfolgende Wechselwirkung mit GHbpI über die
Kontakte der Stelle II erniedrigt die Dissoziationskonstante auf
200 pM. Deswegen verleiht die Wechselwirkung von GHbpII mit dem
Wachstumshormon, das an GHpbI gebunden ist, eine dreifache Erhöhung der
Affinität
auf dem Rezeptorkomplex. Demzufolge ist funktionell der humane βc analog
GHbpII, wobei nur nach der Anfangsassoziation des Liganden mit der
Rezeptor-α-Kette
gebunden wird und eine 1.000-fache Erhöhung der Affinität auf dem
Rezeptorkomplex verliehen wird (wie es der Fall bei IL-3 ist). Die
hier präsentierten
Ergebnisse zeigen allerdings drei signifikante Unterschiede zur
GH-GHR-Wechselwirkung. Als erstes ist die F'-G'-Schleife in GHbpI
und nicht in GHpbII (analog zu βc) bei der Ligandenbindung beteiligt. Als
zweites, obwohl die Reste in der F'-G'-Schleife
von GHbpI effektiv bei der Bindung des Wachstumshormons, Lösungsmittel
ausgeschlossen bei der Bindung des Wachstumshormons, sind, was einen
engen Kontakt zwischen Ligand und Rezeptor indiziert, gibt es offenbar
eine sehr geringe produktive Wechselwirkung zwischen den beiden
Proteinen in dieser Region. Die Alaninüberprüfungsmutagenese über die
Reste dieser Schleife in GHR hatte einen sehr geringen Effekt auf
die Wachstumshormonbindungsaffinität in Relation zum Wildtyp-Rezeptor (Clackson
and Wells, 1995; Bass et al., 1991). Daher trägt der Kontakt zwischen der
F'-G'-Schleife von GHbpI und dem Wachstumshormon
wenig zur Bindungsaffinität
des Rezeptors bei. Dieses ist im Kontrast zu unseren Feststellen
hier, wo wir einen vollständigen
Verlust der Affinitätsumwandlung
für alle
Liganden bei der Substitution von Tyr421 beobachten,
was eine sehr starke Wechselwirkung zwischen diesem Rest und Ligand
im Wildtypmolekül
indiziert.
-
Drittens
und konsistent mit dem Mangel einer produktiven Wechselwirkung zwischen
der F'-G'-Schleife von GHR
und GH, gibt es keine hydrophoben Reste in dieser Schleife. Deswegen,
obwohl GHR Kontaktstellen einer deutlich hydrophoben Natur benutzt,
liegen diese irgendwo in diesem Rezeptor, was anzeigt, dass der
Beitrag zur Affinität
der verschiedenen Kontaktstellen unterschiedlich in βc relativ
zu GHR ist.
-
Der
Erfindung liegt ein Verfahren zugrunde, bei dem ein therapeutisches
Mittel isoliert wird, das an die F'-G'-Schleife von Domäne 4 der
gemeinsamen βc-Kette des Rezeptors für GM-CSF, IL-3 und IL-5 oder
an eine analoge Schleife eines anderen Cytokinrezeptors binden kann,
wobei dieses Verfahren die Stufen umfasst, bei denen Kandidatenmoleküle auf ihre
Fähigkeit überprüft werden,
an diese Schleife zu binden.
-
In
einer Form dieses zweiten Aspekts der Erfindung umfasst die Stufe
der Überprüfung von
Kandidatenmolekülen
auf ihre Fähigkeit
an diese Schleife zu binden, die Überprüfung auf die Fähigkeit,
an eine Sequenz zu binden, die aus der Gruppe gewählt ist,
die SEQ ID Nr. 1, SEQ ID Nr. 2, SEQ ID Nr. 3, SEQ ID Nr. 4; SEQ
ID Nr. 5, SEQ ID Nr. 6, SEQ ID Nr. 7, SEQ ID Nr. 8, SEQ ID Nr. 9,
SEQ ID Nr. 10, SEQ ID Nr. 11 und SEQ ID Nr. 12 umfasst. Es sollte
ebenfalls verstanden werden, dass als Alternative nur die zu überprüfende Schleife aus
solchen gewählt
sein kann, die von einem gemeinsamen Signalübertragungsrezeptor stammt,
oder als zweite Alternative von solchen Rezeptoren mit einem hydrophoben
Aminosäurerest
oder als dritte Alternative von einem solchen Rezeptor, die Signalübertragungsrezeptoren
sind.
-
Alternativ
könnte
gesagt werden, dass die Erfindung in einem Verfahren zu sehen ist,
bei dem ein therapeutisches Mittel isoliert wird, durch Überprüfen auf
die Fähigkeit
der Verbindungen, an das Tyr421 der gemeinsamen βc des
Rezeptors für
GM-CF 6, IL-3 und IL-5, oder den entsprechenden aromatischen hydrophoben
Rest von gp130 oder an einen analogen aromatischen hydrophoben Rest
eines anderen gemeinsamen Signalübertragungsrezeptors
oder an einen spezifischen Rezeptor, der auf andere Cytokine antwortet,
zu binden.
-
Das
Mittel kann ein Antikörper
oder Fragment davon sein, und das Verfahren zur Isolierung dieses
Antikörpers
sollte so verstanden werden, dass weiterhin die Stufen der Impfung
eines Tieres mit einem Peptidmolekül mit der F'-G'-Schleife,
der Fusion der Antikörper
produzierenden Zellen mit einer Myelomzelllinie und die Überprüfung auf
eine Zelllinie, die einen Antikörper
reaktiv mit dieser F'-G'-Schleife und das
Ernten von Antikörpern
aus der Zellline, das Austesten auf die Inhibierung der Hochaffinitätsbindung
und das Testen auf die Inhibierung oder Anregung der Funktion umfasst.
Das kann weiterhin die Herstellung kleiner Fragmente der Antikörper, die
durch die Zelllinie, die an die Schleife bilden kann, hergestellt
werden, umfassen. Die Zelllinie kann in geeigneter Weise eine Mauszellenlinie
sein, und das Verfahren kann weiterhin den weiteren Schritt der
Humanisierung der Antikörperfragmente
durch Ersatz der Maussequenzen mit humanen Sequenzen in den nicht
bindenden Regionen umfassen.
-
Mittel,
die durch das erfindungsgemäße Verfahren
identifiziert werden können,
könnten
in einem Verfahren zur Behandlung eines Menschen oder eines Tiers
einer Krankheit verwendet werden, wobei das Verfahren die Stufe
der Verabreichung eines wie oben identifizierten therapeutischen
Mittels in einem pharmazeutisch annehmbaren Träger und in einer therapeutisch
effektiven Dosis umfassen.
-
Es
könnte
erwünscht
sein, die Erkrankung mit einem oder mehreren der hier identifizierten
oder definierten therapeutischen Mitteln in Kombination zu behandeln,
und vielleicht in Kombination mit anderen therapeutischen Mitteln.
-
Die
Behandlung könnte
als Ziel die Prävention
durch Reduktion des Risikos der Erkrankung abzielen, oder die Behandlung
kann dafür
verwendet werden, die Erkrankung zu lindern oder zu beseitigen.
Die Verabreichung des therapeutischen Mittels kann jede pharmazeutisch
annehmbare Form sein und in einem geeigneten Träger vorliegen.
-
Es
wird angenommen, dass die Konstruktion der Verbindungen, die die
F'-G'-Schleife von βc binden, therapeutisch
nützlich
für Behandlungen
von Krankheiten sind, wo IL-3, GM-CSF und IL-5 eine pathogene Rolle spielen, hauptsächlich die
Allergie, Asthma, Leukämie,
Lymphom und Entzündungen,
einschließlich
Arthritis.
-
Ähnlich wie
bei anderen Cytokinrezeptoren wird angenommen, dass Antagonisten
oder Agonisten therapeutisch nützlich
sind. Somit auch für
gemeinsame Signalübertragungsrezeptoren.
Da gp130 funktionell analog zu βc in dem GM-CSF/IL-3/IL-5-Rezeptorsystem in der Weise ist, dass
es eine gemeinsame Bindungsuntereinheit und Signalüberträger für IL-6,
Onkostatin M (OSM), ziliarer neurotrophischer Faktor (CNTF), Leukämieinhibitionsfaktor
(LIF) und IL-11 ist, ist nahegelegt, dass die Einnahme/Blockierung
dieses Tyrosins zu einem Antagonismus von IL-6, LIF, OSM, CNTF und
IL-11 führt.
Der Antagonismus dieses Rezeptorsystems ist bei Entzündungen,
Leukämie
und Lymphomen nützlich.
Die Antagonisten von IL-2Rβ/γ können als
Immununterdrücker
nützlich
sein. Der Antagonismus von LIFR kann für die Verhinderung der Implantation
von Embryos im Uterus nützlich
sein. Der Antagonist von IL-4/IL-13 inhibiert die IgE-Produktion
und kann bei der Behandlung von Asthma und Allergien nützlich sein.
-
Für spezifische
Untereinheiten. Der Antagonist von IL-3 kann für die Behandlung von Allergien
und Follikel-B-Zellen-Lymphome
geeignet sein. Der Antagonist von IL-4 kann die IgE-Produktion inhibieren
und für die
Behandlung von Asthma und Allergien nützlich sein. Der Antagonist
von IL-6R kann als entzündungshemmendes
Mittel nützlich
sein und kann dafür
verwendet werden, das Myelomwachstum zu inhibieren. Antagonisten
gegen IL-7 können
als Immunsuppressoren nützlich
sein. Antagonisten des Leptinrezeptors (OBR) können bei der Behandlung von
Cachexie, Gewichtsverlust bei Erkrankungen, wie AIDS, Krebs und
Parasitenerkrankungen, nützlich
sein.
-
Der
identifizierte Rest spielt deutlich eine vorherrschende Rolle bei
der Wirkung des Systems und während
einige Moleküle,
die mit der F'-G'-Schleife von βc oder
mit entsprechenden Resten in anderen Rezeptoren, (solche wie in
Tabelle 3 gezeigt) Wechselwirken, können erwartungsgemäß einen
Antagonisteneffekt aufweisen können.
Es ist allerdings auch möglich,
dass sie einen Agonisteneffekt insbesondere bei Rezeptoren, die
durch Homodimerisierung aktiviert werden können, aufweisen.
-
Als
erstes die allgemeinen Signalübertragungsrezeptoren.
Agonistenmittel, die an βc binden, können dafür verwendet werden, um die
Hämopoiesis
zu stimulieren und die Immunantwort gegenüber Mikroorganismen zu verstärken. Agonistenmittel,
die an LIFR binden, können
für die
Unterdrückung
der embryonalen Stammzellendifferenzierung nützlich sein. Agonisten, die
an IL-2Rβ binden,
können
bei der Immunstimulation eingesetzt werden. Agonistenmittel, die
an IL-4R/IL-13 binden, können
eine Antitumoraktivität
aufweisen.
-
Nun
zu den spezifischen Untereinheiten. Agonistenmittel, die IL-3 binden,
können
bei der in vivo und ex vivo Ausbreitung von frühen hämopoetischen Zellen verwendet
werden. Agonistenmittel, die an IL-4 binden, können eine nützliche Antitumoraktivität aufweisen.
Agonisten, die an IL-7R binden, können eine nützliche Antitumorimmunität aufweisen.
Agonistenmittel, die IL-11 binden, können ein nützliches Mittel in der Krebstherapie
darstellen. Agonistenmittel, die an EPOR binden, können dafür verwendet
werden, Anämie
und chronische Nierenleiden oder chronische Entzündungserkrankungen oder maligne
Erkrankungen zu korrigieren. Agonistenmittel, die an TPOR binden,
können
für die
Korrektur von Thrombocytopenie (kann mit chronischen Entzündungserkrankungen,
malignen Erkrankungen, Chemo- und Radiotherapie assoziiert sein)
nützlich
sein.
-
Beispiele
für geeignete
Agonisten sind solche für
Erythropoietin und Thrombopoietin, um die Zahl der Erythrocyten
und Blättchen
im Blut nach einem Blutzellenverlust, der Chemotherapie, Radiotherapie,
Immumsuppression oder Knochenmarktransplantation zu erhöhen. Agonisten
für OBR
können
dafür verwendet
werden, einen Gewichtsverlust zu induzieren, und insbesondere und
nicht-Insulin abhängigen
Diabetes mellitus beiträgt.
Die Moleküle,
sei es Agonist oder Antagonist, können auf der Basis ihrer Fähigkeit,
mit dem identifizierten Rest in Wechselwirkung zu treten, isoliert
werden.
-
Als
kurze Anmerkung, die folgenden Drei- und Einbuchstabenabkürzungen
für Aminosäurereste
werden in der Spezifikation, wie in Tabelle 1 definiert, verwendet.
-
Wenn
der spezifische Aminosäurerest
durch seine Position im Polypeptid eines Proteins angegeben wird,
wird die Aminosäureabkürzung mit
der Nr. des Restes, angegeben als Superscript (das heißt Xaan) verwendet.
-
-
Kurze Beschreibung der
Zeichnungen
-
1A Ist eine schematische
Darstellung der extrazellulären
Domäne
von menschlichem βc, die zwei Cytokin-Rezeptor-Module (CRM1 & 2) (Goodall et
al., 1993) und konservierte Merkmale der Cytokinrezeptorsuperfamilie
(Bazan 1990) zeigt.
-
1B Stellt die Aminosäuresequenzen
dar, die der putativen F'-G' Schleife in der
Membran proximal zu CRM2 von βc (Hayashida et al., 1990) entsprechen, ausgerichtet
mit den F'-G' Schleifen des Wachstumshormonrezeptors
(GHR) (Leung et al., 1987) und Prolactinrezeptors (PRLR) (Boutin
et al., 1988). Konservierte Strukturmotive sind als Box dargestellt,
und zuvor identifizierte Reste, die am Ligandenkontakt (GH) beteiligt sind
(De Vos et al., 1992, und Somers et al., 1994) sind fettgedruckt
gezeigt. Die mutanten Formen von βc, die hergestellt wurden, um die Ligandenbindungsdeterminaten
zu identifizieren, sind mit mutierten Resten fettgezeigt.
-
2 Zeigt graphische Darstellungen,
die zeigen, dass die Alaninsubstitution der Reste 418–422 in βc die
Hochaffinität
der GM-CSF und IL-3-Bindung abschaltet. Es wurde eine Scatchard-Transformation
in Sättigungsbindungsexperimenten
unter Verwendung von 125J-GM-CSF (linkes
Diagramm) und 125J-IL-3 (rechtes Diagramm)
mit COS-Zellen durchgeführt,
die sowohl GM-CSFR und IL-3R-α-Ketten
zusammen mit dem Wildtyp (o) oder der Mutante 418AAAAA422(•)βc exprimieren.
Der radioiodinierte Ligandenkonzentrationsbereich für GM-CSF-
und IL-3 betrug 10 pM–10
nM, und die nicht spezifische Bindung wurde in Gegenwart von 1 μM nicht markiertem
Liganden bestimmt. Die gestrichelte Linie zeigt die hohe und niedrige Affinitätsbindung
der Komponenten unter Verwendung von Wildtyp βc, und
die durchgezogene Linie stellt die Linie der besten Übereinstimmung
für die 418AAAAA422-Mutante βc,
dar, was unter Verwendung des Ligand-Programms bestimmt wurde. Ein
repräsentatives
Experiment ist gezeigt, und die Kd-Werte,
die daraus resultieren und verschiedene andere Experimente sind
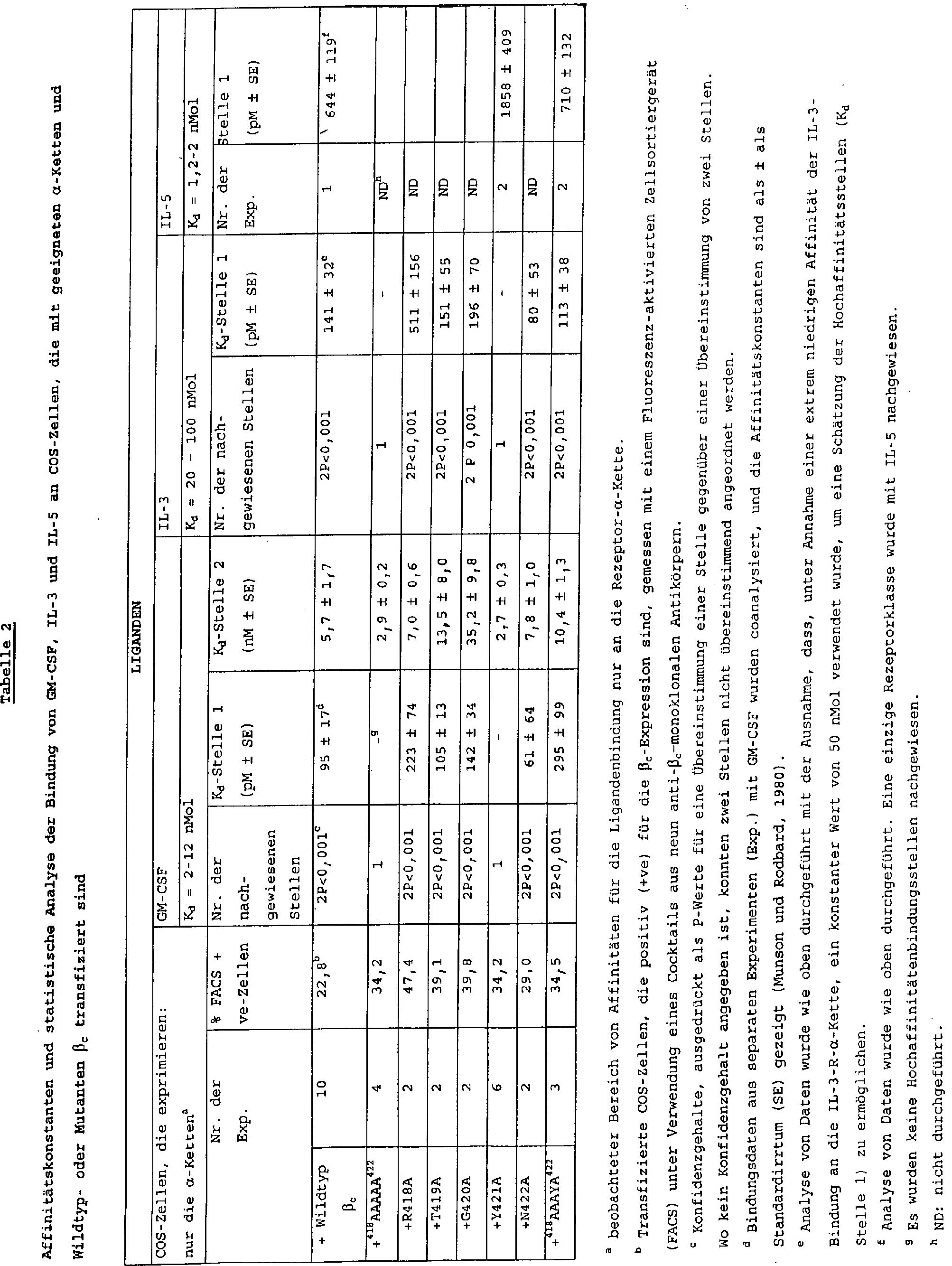
in Tabelle gezeigt.
-
3 Zeigt den Effekt einer
einzelnen Alaninsubstitution der Reste Arg418,
Thr419, Gly420,
Tyr421 und Asp422 von
humanem βc auf die Hochaffinitäts-GM-CSF- und Il-3-Bindung. Die Scatchard-Transformation
der Sättigungsbindungsstudien
mit 125J-GM-CSF (oberer Teil) und 125J-IL-3 (unterer Teil) wurden wie in 2 beschrieben durchgeführt. Die
gestrichelte Linie zeigt die Hoch- und Niedrig-Affinitätsbindungskomponenten
unter Verwendung von Wildtyp βc (o) und die nicht unterbrochene Linie zeigt
die Linie der besten Übereinstimmung
für jede
Mutante βc (•),
was unter Verwendung des Ligand-Programms bestimmt wurde. Ein repräsentatives
Experiment ist gezeigt, und die Kd-Werte
aus diesen und ähnlichen
Studien sind in Tabelle 2 gezeigt.
-
4 Zeigt den Effekt der Wiedereinführung von
Tyr421 in ein polyanalinsubstituiertes βc (418AAAYA422) auf
die Hochaffinitäts-
GMB-CSF und IL-3-Bindung. Die Scatchard-Transformation von Sättigungsbindungsstudien
mit 125J-GM-CSF (linke Seite) und 125I-IL-3 (rechte Seite) wurden wie in 2 beschrieben durchgeführt. Die
unterbrochene Linie zeigt die Hoch- und Niedrigaffinitätsbindungskomponenten
unter Verwendung von Wildtyp βc (o), und die durchgehende zeigt die Linie
der besten Übereinstimmung
für die 418AAAYA422-Mutante βc (•), was unter
Verwendung des Ligand-Programms bestimmt wurde. Ein repräsentatives
Experiment ist gezeigt, und die Kd-Werte
aus diesen und ähnlichen
Studien sind in Tabelle 2 gezeigt.
-
5A Ist die Darstellung eines
Autoradiogramms, das zeigt, dass STAT5-ähnliche (STAT5-L) Aktivität durch
GM-CSF und IL-3 nur in Jurkat-Zellen, die geeignete α-Ketten zusammen
mit dem Wildtyp (Wt) βc exprimieren, induziert wird. Nukleare Extrakte
wurden aus transfizierten Jurkat-Zellen, die für eine Stunde mit oder ohne
300 nM Cytokin stimuliert waren, und über einen elektrophoretischen
Mobilitätsverschiebungsassay unter
Verwendung einer STAT-5 spezifischen Sonde analysiert.
-
5B Ist eine Darstellung
eines Autoradiogramms ähnlich
zu dem von 5A, das zeigt,
dass das DNA-Bindungsprotein,
das durch Cytokin in Rezeptor transfizierten Jurkat-Zellen induziert
wird, sich wie STAT5 verhält.
Ein nuklearer Extrakt, der aus GM-CSF-stimulierten transfizierten
Jurkat-Zellen hergestellt war, wurde einem elektrophoretischen Mobilitätsverschiebungsassay
in Gegenwart eines 50-fachen
molaren Überschusses
von konkurrierenden nicht radioaktiven Oligonucleotiden unterworfen.
Die Oligonucleotide für
die Kompetition waren: das β-Caseinpromotorelement
(β-cas),
eine Mutante für
das β-Caseinpromotorelement (mut β-cas), eine
AP-1- Konsensusbindungsstelle
(AP-1) und das Haemopoeitinrezeptorantwortelement (HRRE).
-
6A Ist eine graphische Darstellung,
dass Y421A βc STAT5-L
mit reduzierter Potenz als Reaktion auf die GM-CSF-Stimulierung aktiviert. Eine Dosis/Antwort-Studie der cytokininduzierten
STAT5-L-Aktivität wurde
mit Jurkat-Zellen durchgeführt,
die mit α-Ketten und entweder
Wild-Ty- βc (o) oder Y421A (•) transfiziert waren. Die STAT5-L-Aktivität wurde
durch einen elektrophoretischen Mobilitätsverschiebungsassay unter
Verwendung einer STAT5-spezifischen
Sonde bestimmt und anschließend
durch Phosphorimager-Analyse quantifiziert. Die Ergebnisse sind
als Prozentzahl der maximalen Aktivität ausgedrückt, die nach der Korrektur
für den
Hintergrund und die Proteinkonzentration erhalten wurde.
-
6B Ist eine graphische Darstellung,
dass Y421A βc STAT5-L
mit reduzierter Potenz als Antwort zur IL-3-Stimulation aktiviert. Die Experimente
und die Prozeduren sind wie in 6A.
-
7 Ein Model der räumlichen
Anordnung zwischen den putativen B'-C' und
F'-G'-Schleifen von βc und
der ersten α-Helix
von GM-CSF, IL-3 oder IL-5 auf der Basis der GH-GHR-Kristallstruktur.
Nur die membrannahen Domänen
von βc (CRM2) und die erste α-Helix (Helix A) von GM-CSF, IL-3 oder
IL-5 sind gezeigt, mit Tyr365 und Tyr421 von βc und dem konservierten Glu von GM-CSF, IL-3
oder IL-5, die in der raumausfüllenden Darstellung
gezeigt sind.
-
DETAILLIERTE
BESCHREIBUNG DER BEISPIELE
-
BEISPIEL 1
-
Darstellung der Wirkung
von Tyr421 bei der βc-Untereinheitbindung
-
Grundprinzip
für die
Mutagenese von putativen Schleifen in der humanen β-Kette
-
Die
Region von βc, die wir für die Mutagenese gewählt haben,
wird durch Regionen flankiert, die eine Konservierung durch die
Cytokin-Rezeptorsuperfamilie zeigt: der zuvor beschriebene VRXR-Konsensus (Patthy
1990), worin X am meisten vorkommmend einen aliphatischen Rest darstellt
und das WSXWS-Motiv (Cosman et al., 1990; Bazan 1990) (1). Das WSXWS-Motiv ist
bereits seit langem als charakteristisches Merkmal dieser Rezeptorfamilie
erkannt, und verschiedene Studien sind auf das Verständnis seiner
Rolle in der Rezeptorfunktion gerichtet worden (Miyazaki et al.,
1991; Yoshimura et al., 1992; Quelle et al., 1992; Rozakis-Adcock
and Kelly 1992; Baumgartner et al., 1994). Die GH-GHR-Kristallstruktur
(De Vos et al., 1992) und seit kurzem die Kristallstruktur von GH,
gebunden an den Prolactinrezeptor (Somers et al., 1994) geben uns etwas
Einsicht in die Struktur der VRXR-WSXWS-Region in Cytokinrezeptoren
im Allgemeinen. Die beiden betreffenden β-Stränge stehen miteinander in Wechselwirkung
und bilden einen Stapel von aromatischen Seitenketten, die durch
hydrophobe Bereiche der basischen Seitenketten unterbrochen werden,
und in der Konsequenz bilden die Aminosäuren, die diese beiden Motive
trennen, eine Schleife. Die Reste in dieser Schleife sind für ein Lösungsmittel
in dem Liganden gebundenen Komplex in beiden gelösten Rezeptoren nicht zugänglich (De
Vos et al., 1992; Somers et al., 1994). Wir haben daher unser Augenmerk
hier auf den analogen F'-G'-Schleifenbereich
in der Membran proximal zum Cytokinrezeptormodul (CRM) von βc gerichtet,
um seine Rolle bei der GM-CSF-, Il-3- und Il-5-Bindung zu bestimmen.
-
Identifikation einer Region,
die bei der GM-CSF-, IL-3- und IL-5-Hochaffinitätsbindung beteiligt ist
-
Zur
Untersuchung der möglichen
Beteiligung der putativen F'-G'-Schleife von βc bei
der Hochaffinitäts-rezeptorkomplexbildung,
wurde die Region durch Substitution von Alaninen über die
Reste 418–422 (1) unterbrochen. Die Fähigkeit
dieser n-Alanin substituierten Mutante βc (418AAAAA422), die
Hochaffinitätsligandenbindung
zu unterstützen,
wurde durch Co-Expression
dieser mit sowohl GM-CSFR- und IL-3R- α-Ketten an COS-Zellen studiert,
wobei Radioliganden-Sättigungsbindungsstudien
durchgeführt
wurden. COS-Zellen wurden routinemäßig mit α-Ketten-cDNAs zusammen mit mutanter βc-cDNA
elektroporatisiert, so dass die gleiche Transfekte auf die GM-CSF- und IL-3-Bindung
analysiert werden konnte. In Abwesenheit von transfiziertem βc,
haben GM-CSFR und die IL-3R- α-Kette
transfizierten COS-Zellen GM-CSF und IL-3 mit nur geringer Aktivität gebunden
(Tabelle 2). Die Co-Expression von Wildtyp βc verlieh
GM-CSF eine hohe Affinitätsbindung,
und IL-3 wie die α-Ketten
wurden im Überschuss
von βc exprimiert, die geringe Affinitätsbindung
war immer noch evident, wie durch die Krummlinigkeit der Scratchard-Plots
(2, Tabelle 2) dargestellt
ist. Die COS-Zellen, die mit dem mutanten Penta-Alanin βc (418AAAAA422), GM-CSFR
und IL-3R- α-Kette
transfiziert waren, zeigten eine Zelloberflächenexpression aller Rezeptorketten
im Vergleich zu dem Wildtyp Transfektanten (Tabelle 2), sie zeigten
allerdings keine hohe Affinitätsbindung
von GM-CSF oder IL-3 (2,
Tabelle 2). Dieses zeigt, dass die Substitution der Region 418–422 in βc die
Fähigkeit
von βc beeinträchtigt,
eine Hochaffinitäts-GM-CSF- und IL-3-Bindung
zu unterstützen
und legt nahe, dass die Reste in dieser Region bei der Ligandenwechselwirkung
beteiligt sind.
-
Tyr421 ist
notwendig und ausreichend für
die GM-CSF-, IL-3- und IL-5-Hochaffinitätsbindung
-
Zur
Untersuchung des jeweiligen Beitrags der Reste 418–422 von βc auf
die Hochaffinitäts-
GM-CSF-, IL-3- und IL-5-Bindung, wurde jeder Rest in dieser Region
separat mit Alanin substituiert (1).
Wie oben wurde die Fähigkeit
der einzelnen Mutante, eine Hochaffinitätsligandenbindung zu vermitteln,
bestimmt, indem Radioligandensättigungsbindungsstudien
bei COS-Zellen, die die Mutante βc zusammen mit GM-CSFR und IL-3R α-Ketten exprimieren,
durchgeführt
wurden. Die Ergebnisse zeigten, dass alle Alaninsubstitutionsmutanten ähnlich wie
der Wildtyp βc auf der Zelloberfläche exprimiert wurden (Tabelle
2), und die Alaninsubstitution der Reste Thr419,
Glu420 und Asn422 hatte
keinen Einfluss auf die Hochaffinitätsligandenbindung (3, Tabelle 2). Die Alaninsubstitution
von Arg418 reduzierte die GM-CSF- und IL-3-Hochaffinitätsbindung
um das 2–3-fache,
was ein relativ kleiner Effekt ist (Tabelle 2). Interessanterweise
erzeugte allerdings die Alaninsubstitution von Tyr421 eine βc-Mutante,
die nicht in der Lage war, die Hochaffinitätsbindung von jeweils GM-CSF
oder IL-3 zu unterstützen
(2, Tabelle 2). Der
Effekt dieser Mutation auf die Hochaffinitäts-IL-5-Bindung wurde ebenfalls bestimmt, indem
COS-Zellen mit der Mutante Y421A βc zusammen mit der IL-5R-α-Kette transfiziert wurde. Die
mit radioaktiv iodiertem IL-5 erhaltenen Sättigungsbindungsdaten zeigten
eine einzige Klasse eines Rezeptors bei der Scatchard-Analyse. Obwohl βc einen
geringen Effekt auf die Affinitätsumwandlung
der IL-5-Bindungsstelle relativ zu IL-3 und GM-CSF hat, zeigten
Studien mit der Mutante Y421A βc einen Verlust der Affinitätsgröße mit dem
Verlust der Hochaffinitäts-IL-5-Bindung
(Tabelle 2). Dieses zeigt, dass Tyr421 eine ähnliche
Rolle bei der Unterstützung
der Hochaffinitätsbindung
von GM-CSF, IL-3 und IL-5 aufweist.
-
Die
Identifikation eines einzigen Restes in einer vorbestimmten Schleife
der Membran proximal zu CRM von βc, die für
die Hochaffinitäts-
GM-CSF-, IL-3- und IL-5-Bindung kritisch ist, hat die Möglichkeit
eröffnet,
dass dieser Rest allein im Zusammenhang einer Schleife ausreichend
sein könnte,
um die Hochaffinitätsbindung
zu unterstützen.
Um diese Feststellung zu testen, haben wir einen Tyrosinrest wieder
in eine polyalaninsubstituierte βc-Mutante (418AAAYA422) (1)
eingeführt. Überraschenderweise
war diese Mutante in der Lage, die Hochaffinitäts-GM-CSF- und IL-3-Bindung
(4) zu unterstützen, obgleich
mit einer dreifach reduzierten Affinität relativ zu Wildtyp GM-CSF-
und IL-3-Hochaffinitätsbindung
(Tabelle 2). Diese Mutante stellte ebenfalls die Hochaffintätsbindung
an IL-5 wieder her (Tabelle 2). Diese Feststellungen zeigen, dass
die Reste neben Tyr421, keine direkte Rolle
bei der Ligandenwechselwirkung spielen, und sie implizieren Tyr421 als Schlüsselrest bei der GM-CSF-, Il-3-
und IL-5-Hochaffinitätsbindung.
-
Die Substitution von Tyr421 unterbricht die funktionelle Rezeptoraktivierung
-
Zur
Bestimmung der funktionelle Signifikanz der Substitution von Tyr421 von βc durch Alanin, haben wir die Induktion der
Signalübertragung
und die Transkriptionsaktivitoren (STAT) gemessen. Die produktive
Wechselwirkung von GM-CSF, IL-3 und IL-5 mit ihren Hochaffinitätsrezeptoren
führt zu
einer von βc vermittelten Signalübertragung (Kitamura et al.,
1991b; Kitamura et al., 1992; Takaki et al., 1993). Es ist nun gut
dokumentiert, dass die Signalgebung durch βc die
JAK2-Protein-Tyrosinkinase
(Silvennoinen et al., 1993; Quelle et al., 1994) aktiviert, und
anschließend
der Transkriptionsfaktor STAT5 schnell phosphoryliert wird, was
diesen in die Lage versetzt, an seine DNA-Bindungsstelle im Kern
zu binden (Mui et al., 1995; Azam et al., 1995; Gouilleux et al., 1995;
Pallard et al., 1995). Die STAT5-Aktivierung stellt daher eine relativ
sofortige Antwort auf die βc vermittelte Rezeptorsignalgebung dar. STAT5
ist in vielen hämopoietischen
Zelllinien, einschließlich
Zellen von myeloider und lymphoider Abstammungen beschrieben worden
(Mui et al., 1995; Azam et al., 1995; Gouilleux et al., 1995; Pallard
et al., 1995). In vorläufigen
Experimenten haben wir Jurkat-T-Zellen, die mit GM-CSFR-, IL-3R-
oder IL-5R-Rezeptor-α-Ketten
zusammen mit Wildtyp βc transfiziert sind, untersucht. Ein Cytokin
induzierbares nukleares DNA-Bindungsprotein wurde nachgewiesen,
das eine Oligonucleotidsonde, die die STAT5-Bindungskonsensussequenz (TTc-N3-GAA) enthält, in einem elektrophoretischen
Mobilitätsverschiebungsassay
verzögerte.
-
Dieses
DNA-Bindungsprotein war nach der Cytokinbehandlung in den Zellen,
die mit der α-Kette
und βc transfiziert waren, induzierbar, allerdings
nicht in Abwesenheit von βc, was anzeigt, dass βc diesen
ligandeninduzierten Prozess vermittelt (5A). Die nicht markierte Sonde wie auch
das verwandte Hämopoiteinrezeptorantwortelement
(HRRE) (Morella et al., 1995) konkurrierten um die Bindung an die
radioaktiv markierte β-Casein-Promotorsonde,
wobei beide die STAT5-Bindungskonsenssusequenz enthalten. Allerdings
konkurrierten eine β-Casein-Promotermutantensonde,
die Mutationen im STAT5-Bindungskonsensus
und eine kanonische Sonde der AP-1-Stelle enthielt, nicht um die
Bindung (5B). Aus diesem
Grund bezeichnen wir dieses Protein als STAT5-ähnlich (STAT5-L), weil, obwohl
sie sich in ähnlicher
Weise wie es STAT5 verhält,
seine wahre Identität
noch nicht bestimmt worden ist.
-
Die
Fähigkeit
von mutanten Rezeptoren, das STAT5-L-Protein zu aktivieren, wurde
bestimmt, indem Jurkat-Zellen mit der Mutante βc- und
die GM-CSFR- oder IL-3R-α-Kette
cDNAs cotransfiziert wurden. Die Zelloberflächenexpression von Rezeptoruntereinheiten
wurde durch Fliesscytometrie unter Verwendung von kettenspezifischen
Antikörpern
bestätigt,
und es ist festgestellt worden, dass die Expression von Mutanten- und Wildtyp βc vergleichbar
war (Daten nicht gezeigt). Die Transfektanten wurden mit entweder
GM-CSF oder IL-3 über
einen Konzentrationsbereich stimuliert, und es wurden nukleare Extrakte
hergestellt. Die Induktion des STAT5-L-Proteins wurde mit einem
elektrophoretischen Mobilitätsverschiebungsassay
(EMSA) unter Verwendung der kanonischen STAT5-DNA-Bindungsstelle vom β-Caseinpromotor
als Sonde bestimmt.
-
STAT5-L
wurde in Reaktion auf entweder GM-CSF oder IL-3 in dosisabhängiger Weise
in Gegenwart der geeigneten α-Kette
und des Wildtyp βc aktiviert (6A & B).
Die Alaninsubstitution von Tyr421 produzierte ein βc,
das die STAT5-L-Aktivierung als Reaktion auf GM-CSF mit einer etwa
100–1000-fachen
Reduktion der Potenz unterstützte
(6A). Diese Reduktion
der GM-CSF-Antwort
mit dieser Mutanten-βc zeigt, dass Tyr421 eine
Rolle bei der GM-CSF induzierten Rezeptoraktivierung aufweist. Signifikanterweise
hatte Y421A βc einen größeren Effekt auf die IL-3-induzierte
STAT5-L-Aktivierung, die nur geringe Gehalte von nachweisbarem STAT5-L
(6B) produzierte, und
selbst bei 3 μM
IL-3 wurde eine halbe maximale Aktivierung nicht erreicht. Dieses
liegt nahe, dass der Tyrosinwert ebenfalls für die Wechselwirkung von IL-3
mit βc kritisch ist.
-
BEISPIEL 2
-
Herstellung
von Antikörperantagonisten
-
Monoclonale
Antikörper
können
hergestellt werden, indem mit βc oder mit einem Fragment von βc,
das die F'-G'-Schleife enthält, oder
mit einem Peptid, das die F'-G'-Sequenz enthält, immunisiert
wird. Nach Spezifizitätskontrollen,
die die spezifische Bindung an die F'-G'-Schleife
zeigen, können
die Antikörper
für die
Blockierung der GM-CSF-, IL-3- und IL-5-Hochaffinitätsbindung und für die Blockierung
der GM-CSF-, IL-3-
und IL-5-Funktionsstimulation gewählt werden. Wenn einmal ein
geeigneter monoclonaler Antikörper
identifiziert worden ist, der die GM-CSF-/, IL-3-/ und IL-5-Hochaffinitätsbindung
und -funktion blockiert, können
kleinere Fragmente hergestellt werden, z. B. F(ab)2,
Fab und schließlich
Fv. Unter Anwendung von Techniken der Molekularbiologie kann eine
einzelne Kette des Fv-Fragments konstruiert werden (Hv-Lv). Das
wäre ein
Inhibtionspeptid.
-
BEISPIEL 3
-
Herstellung
von Peptidantagonisten
-
Kurze
Peptide mit ähnlichen
Sequenzen zur F'-G'-Schleife können synthesiert
werden, die die Cytokinwechselwirkung mit der F'-G'-Schleife
von βc blockieren. Das Umgekehrte ist ebenfalls
möglich:
kurze Peptide mit ähnlichen
Sequenzen zur Helix A von GM-CSF/IL-3/IL-5 (dieses ist die Region
in den Cytokinen, die mit der F'-G'-Schleife wechselwirken
soll) können
ebenfalls die Cytokinwechselwirkung blockieren.
-
BEISPIEL 4
-
Herstellung
von Oligonucleotidantagonisten
-
Ein
großer
Pool von wahllos synthesierten Oligonucleotiden kann durch ein F'-G'-Peptid, das auf
einer festen Matrix (Bock et al., 1992, – diese Referenz ist hiermit
eingeschlossen) immobilisiert ist, gegeben werden. Nach dem Waschen
verbleiben die starkbindenden Oligonucleotide, die dann bei verschiedenen
Bedingungen (Salz, pH, etc.) eluiert werden können. Die Sequenz kann dann
durch PCR bestimmt werden und auf ihre Inhibition der βc vermittelten
Funktion auf einem realen Zellsystem getestet werden.
-
BEISPIEL 5
-
Ausrichtung von Aminosäuren der
F'-G'-Schleife von βc und
gp130
-
Die
Ausrichtung von βc und gp130 ist publiziert worden. Gp130
und βc enthalten Domänen, die mit der Fibronectindomäne vom Typ
III, die ein charakteristisches Merkmal der meisten Cytokinenrezeptoren
ist, verwandt sind. Gp130 hat 6 dieser Domänen, βc hat
4 dieser Domänen.
-
Jede
Domäne
enthält
reine eigene F-G-Schleife, somit hat gp130 6 und βc hat
4. Nur eine dieser F-G-Schleifen ist die wichtige, soweit die vorliegende
Erfindung betroffen ist. Die wichtige Domäne ist durch die Gegenwart
von i) einem konservierten Muster von Cys-Resten, ii) verschiedenen
Trp-Resten, iii) einem YXXRV/IR Motiv und iv) einem WSXWS Motiv
(wo X eine Aminosäure
ist) charakterisiert. Es ist postuliert, dass Tyr421 innerhalb
der F-G-Schleife der vierten Domänschleife
in βc (F'-G') liegt. Es ist postuliert, dass das äquivalente
funktionell wichtige Tyr in gp130 innerhalb der F-G-Schleife der
zweiten Domäne
liegt.
-
-
Tabelle
3 Ausrichtung der putativen F'-G'-Schleifen von Mitgliedern
der Cytokinrezeptorsuperfamilie
-
Die
aromatischen hydrophoben Reste sind unterstrichen.
- Ackerman
et al., (1994) Chest, 105; 687
- Adachi et al., (1995) Am J Respir Crit Care Med., 151, 618
- Akir et al (1993) Adv Immunol 54, 1–78.
- Azam, M. et al. (1995) EMBO J., 14, 1402–1411.
- Bagley, C. J., et al. (1995) J. Leuk. Biol., 57, 739–746.
- Barry, S. C., et al (1994) J. Biol. Chem., 269, 8488–8492.
- Bass, S. H. et al (1991) Proc. Natl. Acad. Sci. USA, 88, 4498–4502.
- Baumgartner, J. W. et al (1994) J. Biol. Chem., 269, 29094–29101.
- Bazan, J. F. (1990) Proc. Natl. Acad. Sci. USA, 87, 6934–6938.
- Bentley et al., (1996) Am J Respir Crit Care Med 153, 551
- Bock et al., (1992) Nature 355, 564–566
- Boutin, J. M., et al (1988) Cell, 53, 69–77.
- Broide et al (1992) J Clin Invest, 90, 1414
- Clackson, T and Wells, J. A. (1995) Science, 267, 383–386.
- Clayberger et al (1991), J Exp Med 175, 371
- Contreras, M. A. et al (1983) Methods Enzymol., 92, 277–292.
- Cosman, D. et al (1990) Trends Biochem. Sci., 17, 265–270.
- Cunningham, B. C. et al (1991) Science, 254, 821–825.
- Davis, S. et al (1993) Science, 260, 1805–1810.
- De Vos, A. et al (1992) Science, 255, 306–312.
- Elliott MJ, et al (1989) Blood 74, 2349–2359.
- Foster et al (1996) J Exp Med 183, 195
- Fukuda et al., (1994) J Allergy Clin Immunol. 94, 584
- Gearing, D. P. et al (1991) EMBO J., 10, 2839–2848.
- Gearing, D. P. et al (1994) Proc. Natl. Acad. Sci. USA, 91,
1119–1123.
- Giri, J. G. et al (1994) EMBO J., 13, 2822–2830.
- Goodall, G. J. et al (1993) Growth Factors, 8, 87–97.
- Goodwin, R. G. et al (1990) Cell, 60, 941–951.
- Gouilleux, F., et al (1995) EMBO J., 14, 2005–2013.
- Groopman JE, et al (1987) New Engl J Med, 317, 593–598.
- Hamid et al., (1991) J Clin Invest, 87, 1541
- Hatakeyama, M. et al (1989) Science, 244, 551–556.
- Hayashida, K. et al (1990) Proc. Natl. Acad. USA, 87, 9655–9659.
- Hercus, T. R. et al (1994) Proc. Natl. Acad. Sci. USA, 91, 5838–5842.
- Hibi, M. et al (1990) Cell, 63, 1149–1157.
- Hilton, D. J. et al (1994) EMBO J., 13, 4765–4775.
- Hilton, D. J. et al (1996) Proc. Natl. Acad. Sci. USA, 93, 497–501.
- Idzerda, R. L. et al (1990) J. Exp. Med., 171, 861–873.
- Ip, N. Y. et al (1992) Cell, 69, 1121–1132.
- Kato M, et al (1992) Lymphokine Cytokine Res 11, 287–292.
- Kay AB et al (1991) J Exp Med 173, 775–778.
- Kimura, Y. et al (1995) Int. Immunol, 7, 115–120.
- Kitamura, T. et al (1991a) Cell, 66, 1165–1174.
- Kitamura, T. et al. (1991b) Proc. Natl. Acad. Sci. USA, 88,
5082–5086.
- Kitamura, T. and Miyajima, A. (1992) Blood, 80, 84–90.
- Leung, D. W. et al (1987) Nature, 330, 537–543.
- Liu, J. et al (1992) J Biol. Chem., 267, 16763–16766.
- Lock, P. et al (1994) Proc. Natl. Acan. Sci. USA, 91, 252–256.
- Lopez AF et al (1989). Proc Natl Acad Sci USA 86, 7022–7026.
- Lopez et al (1990) J Cell Physiol, 145, 69–77
- Lopez AF et al (1991) J Biol Chem 266, 24741–24747.
- Lopez AF et al (1992) Immunol Today 13, 495–500.
- Lopez, A. F. et al EMBO J., 11, 909–916.
- Marini et al., (1992), Chest 102, 661
- Mauser et al., (1993) Am Rev Respir Dis, 148, 1623
- Mauser et al (1995) Am J Respir Crit Care Med 152, 467
- Miyajima, Mol Cell Biol. (1992) Trends Biochem Sci 17, 38–382
- Miyazaki, M. et al (1991) EMBO J., 10, 3191–3197.
- Morella. K. K. et al (1995) J. Biol. Chem., 270, 8298–8310.
- Mui, A. L.–F.
et al (1995) EMBO J., 14, 1166–1175.
- Munson, P. J. and Rodbard. D. (1980) Anal. Biochem., 107, 220–239.
- Nakamura. Y. et al. (1992) Science, 257, 1138–1141.
- Noguchi, M. et al (1993) Science, 262, 1877–1880.
- Pallard, C. et al (1995) J. Biol. Chem., 27, 15942–15945.
- Park LS, et al (1989) Blood, 74, 56–65.
- Patthy, L. (1990) Cell, 61, 13–14.
- Pennica, D. et al (1995) J. Biol. Chem., 270, 10915–10922.
- Quelle, D. E. et al (1992) Mol. Cell. Biol., 12, 4553–4561.
- Quelle, F. W. et al (1994) Mol. Cell Biol., 14, 4335–4341.
- Robinson et al., (1992) N Engl J Med 326, 298
- Rozakis-Adcock, M. and Kelly, P. A. (1992) J. Biol. Chem., 267,
7428–7433.
- Russell, S. M. et al (1993) Science, 262, 1880–1883.
- Shanafelt, A. B. and Kastelein, R. A. (1992) J. Biol. Chem.,
267, 25466–25472.
- Silvennoinen, O. et al (1993) Proc. Natl. Acad. Sci. USA, 90,
8429–8433.
- Somers, W. et al (1994) Nature, 372, 478–481.
- Sur et al., (1995) J Allergy Clin Immunol, 96, 661
- Sur et al., (1996) J Allergy Clin Immunol, 97, 1272
- Taga, T. et al (1992) Proc. Natl. Acad. Sci. USA, 89, 10998–11001.
- Takaki, S. et al (1993) J. Exp. Med., 177, 1523–1529.
- Takeshita, T. et al (1992) Science, 257, 379–382.
- Tartaglia et al., (1995) Cell 83, 1263–1271
- Tavernier, J. et al (1991) Cell, 66, 1175–1184.
- Vigon, I. et al (1992) Proc. Natl. Acad. Sci. USa, 89, 5640–5644.
- Williamson DJ et al (1988) Clin Exp Immunol 72, 67–73.
- Woodcock, J. M. et al (1994) EMBO J., 13, 5176–5185.
- Xing et al., (1996) J Clin Invest 97, 1102–1110
- Yamasaki, K. et al (1988) Science, 241, 825–828.
- Yawata, H. et al (1993) EMBO J., 12, 1705–1712. Yoshimura, A. et al
(1992) J. Biol. Chem., 267, 11619–11625.
- Young DC, Griffin JD. (1986) Blood 68, 1178–1181.
- Zangrilli et al., (1995) Am. J Respir Crit Care Med 151, 1346
- Zurawski, S. M. et al (1993) EMBO J, 12, 2663–2670.
-
-
-
-
-
-
-