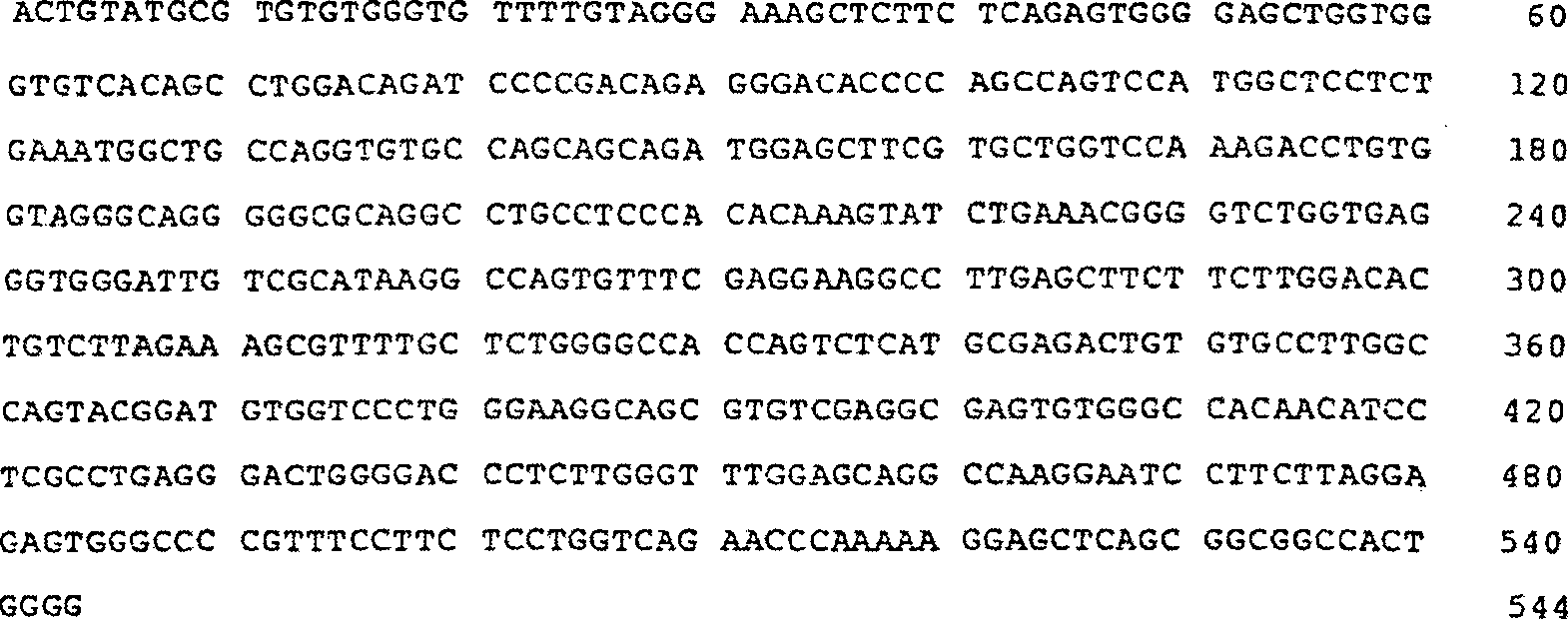-
Die
vorliegende Erfindung betrifft ein Verfahren zum Screenen einer
Verbindung, welche das Expressionsniveau der menschlichen induzierbaren
Stickstoffoxid-Synthase beeinflussen kann, genauer gesagt ein Verfahren
zum Screenen einer Verbindung, welche die Expression des Enzyms
kontrollieren kann.
-
Seitdem
Stickstoffoxid (NO) als gefäßerweiternder
Faktor identifiziert wurde, hat man herausgefunden, dass es eine
physiologisch aktive Substanz ist, die eine wichtige Rolle bei der
Regulierung der biologischen Funktionen spielt. Es wurde berichtet,
dass NO zusätzlich
zu den vorstehenden Funktionen eine thrombocytenaggregationssupprimierende
Wirkung, eine neurotransmitterfreisetzende Wirkung und eine Wirkung
besitzt, die dazu führt,
dass Makrophagen antitumorale und bakterizide Aktivitäten zeigen
(Moncada, S. und Higgs, A. (1993) N. Engl. J. Med. 329, 2002–2012).
-
NO
wird biologisch durch NO-Synthase (NOS) aus L-Arginin als Substrat
synthetisiert. Derzeit ist die Existenz dreier Isoenzyme von diesem
Enzym bestätigt
(der Gehirn-Typ, der Endothel-Typ und der induzierbare Typ). Ihre
chromosomale Lokalisierung ist ebenfalls bekannt (Knowles, R. G.
und Moncada, S. (1994) Biochem. J. 298, 249–258).
-
Von
diesen Typen kann der induzierbare Typ NOS (iNOS) exprimiert werden,
indem Endotoxine oder Cytotoxine für solche Zellen wie vaskuläre Zellen
der glatten Muskulatur (VSMC), Hepatocyten, Chondrocyten oder Gliazellen
verwendet werden, so dass die Gentranskription induziert wird. Kürzlich wurden
Mäuse,
denen dieses Gen fehlt, (Knockout-Mäuse) gezüchtet. Es wurde berichtet,
dass die Mäuse
eine schwächere
Abwehrfähigkeit
gegen Infektionen besitzen, aber im Vergleich zum Wildtyp leichtere
Symptome von Entzündung
und Sepsis zeigen (Wei, X. et al., (1995) Nature 375, 408–411; MacMicking,
J. D. et al., (1995) Cell 81, 641–650). Es gibt einen Bericht,
der zeigt, dass iNOS in jeder Spezies durch inflammatorische Zustände induziert
wird und dass die Suppression seiner enzymatischen Aktivitäten und
die Expression zur Erleichterung der Entzündungssymptome wirksam sind
(Moncada, S. und Higgs, E. A. (1995) FASEB J. 9, 1319–1330).
Ferner fand man in einem Sepsis-Modell heraus, dass die Verabreichung
von iNOS-Enzym-Inhibitoren
wirksam ist (Kerwin, J. F. et al., (1995) J. Med. Chem. 38, 4343–4362).
-
Es
wurde andererseits berichtet, dass der Endotheltyp-NOS (eNOS) an
der Homöostase,
insbesondere an der Suppression des Anstiegs des Blutdrucks, beteiligt
ist, und man zieht in Betracht, dass er eine wichtige Rolle bei
biologischen Funktionen spielt. Folglich war es wünschenswert,
eine Verbindung zu finden, die die eNOS-Aktivität nicht beeinflusst, sondern
die iNOS-Aktivität
spezifisch hemmt. Da jedoch die primären Strukturdomänen der
Proteine, die die Aktivitäten
dieser Isoenzyme hemmen, einander sehr stark ähneln, ist keiner der vorliegenden
NOS-Enzym-Inhibitoren im Hinblick auf seine Spezifitäten zufriedenstellend.
-
In
anderer Hinsicht wurde bestätigt,
dass Substanzen, die NO erzeugen, die vaskuläre Hypertrophie supprimieren
können,
so dass Arteriosklerose und postangioplastische vaskuläre Restenose
in Tiermodellen verhindert wird (Garg, U. C. und Hassid, A. J. (1989)
J. Clin. Invest. 83, 1774–1777,
Cooke, J. P. et al., (1992) J. Clin. Invest. 90, 1168–1172).
Es wurde auch berichtet, dass die erzwungene Expression des NOS-Gens
in VSMC zu einem Anstieg der NO-Produktion führt, die mit der Suppression
der Hypertrophie der inneren Membran von VSMC einhergeht (von der
Leyen, H. E. et al., (1995) Proc. Natl. Acad. Sci. USA 92, 1137–1141). Daher
erwartet man, dass die Erzeugung von NO an der Stelle der vaskulären Hypertrophie
bei der Behandlung oder Vermeidung der vaskulären Hypertrophie wirksam sein
kann.
-
Die
vorstehend beschriebenen Tatsachen legen nahe, dass, falls man iNOS-Isozym-spezifische
genexpressionsregulierende Verbindungen findet, die Inhibitoren
als antiinflammatorische Agentien nützlich sind, während die
Induktoren als Agentien nützlich
sind, die zu therapeutischen Verbesserungen insbesondere auf dem
cardiovaskulären
Gebiet führen.
-
Reportergene
werden zur Überwachung
der Expression eines bestimmten Gens einfach und leicht mit hoher
Sensitivität
eingesetzt (Yokota, T., und Arai, K. (1993) Biomanual Series 4,
Youdosha). Anstatt die Testgenexpression direkt nachzuweisen, werden
Reportergene verwendet, und derartige Gene, die derzeit häufig verwendet
werden, schließen
diejenigen ein, deren Expressionsprodukte wie Chloramphenicol-Acetyltransferase
(CAT), β-Galactosidase
(β-Gal)
und Luciferase leicht getestet werden können. Die Aktivität der Test-DNA, deren
regulatorische Aktivität
gemessen werden soll, kann mit hoher Sensitivität nachgewiesen werden, indem sie
entweder stromaufwärts
oder stromabwärts
des Reportergens in ein Plasmid eingebaut wird.
-
Die
Reportergene werden verwendet, um die Regulation der Maus-iNOS (miNOS)-Genexpression
zu untersuchen, und über
die Ergebnisse wurde berichtet. Beispielsweise fand man innerhalb
der stromaufwärts liegenden
Region der miNOS-Transkriptionsstartstelle
(hier nachstehend als „flankierende
5'-Region" bezeichnet) eine
Region etwa 1,7 kb stromaufwärts
von der Transkriptionsstartstelle, die an der Induktion des miNOS-Gens
als Reaktion auf Lipopolysaccharide (LPS) oder IFN-γ beteiligt
ist. Ferner wurde gezeigt, dass Consensussequenzregionen, von denen
man annimmt, dass die Transkriptionsfaktoren NF-κB und IRF-1 daran binden, bei
der Induktion der Genexpression wesentlich sind (Xie, Q. et al.,
(1993) J. Exp. Med. 177, 1779–1784,
Vodovotz, Y. et al., (1993) J. Exp. Med. 178, 605–613, Martin,
E. et al., (1994) J. Exp. Med. 180, 977–984, Xie, Q. et al., (1994)
J. Biol. Chem. 269, 4705–4708).
Nach einem weiteren Bericht fand man die Region innerhalb der flankierenden
5'-Region mit etwa
1,6 kb (Lowenstein, C. J. et al., (1993) Proc. Natl. Acad. Sci.
USA, 90, 9730–9734,
Kamijo, R. et al., (1994) Science 263, 1612–1615). In beiden Fällen betrug
die Induktionsfähigkeit
jedoch weniger als das 50-Fache, was nicht stark genug sein kann,
um die eigentliche hohe Induktionsfähigkeit des iNOS-Gens widerzuspiegeln.
-
Dagegen
war es schwierig, die Existenz der cDNA des menschlichen iNOS (hiNOS)-Gens
zu beweisen. Einer der Gründe
dafür liegt
darin, dass die Zellen, in denen die Induktion durch ein einzelnes
Cytokin oder eine Kombination von zwei Cytokinen erfolgte, im Gegensatz
zu Maus-Makrophagen oder Ratten-VSMC nicht nachgewiesen wurden.
Es gibt einen Bericht, dass hiNOS in menschlichen Makrophagen sogar
durch eine Kombination von drei oder mehr unterschiedlichen Arten
von Cytokinen nicht induziert werden kann (Weinberg, J. B. et al.,
(1995) Blood 86, 1184–1195).
Nach dem Bericht über
die erste erfolgreiche Clonierung von hiNOS-cDNA durch Stimulieren
menschlicher Hepatocyten mit drei oder mehr unterschiedlichen Arten
von Cytokinen, um das Gen zu induzieren (Geller, D. A. et al., (1993)
Proc. Natl. Acad. Sci. USA, 90, 3491–3495), tauchten jedoch viele
Berichte über
die Clonierung von hiNOS-cDNA auf, aber der regulatorische Mechanismus
der Induktion wurde nicht aufgeklärt (Sherman, P. A. et al.,
(1993) Biochemistry 32, 11600–11605,
Charles, I. G. et al., (1993) Proc. Natl. Acad. Sci. USA 90, 11419–11423,
Hokari, A. et al., (1994), J. Biochem. 116, 575–581). Gleichzeitig wurden
die vollständige
Struktur des hiNOS-Strukturgens (1) und dessen
Nucleotidsequenz der flankierenden 5'-Region mit etwa 0,4 kb aufgeklärt (Chartrain,
N. A. et al., (1994) J. Biol. Chem., 269, 6765–6772).
-
Andererseits
gelang es den hier genannten Erfindern, das iNOS-Gen aus der vaskulären glatten
Muskulatur von Ratten zu clonieren, und sie berichteten, dass dessen
cDNA vermutlich überall
in verschiedenen Geweben und Spezies einheitlich ist (Nunokawa,
Y. et al., (1993) Biochem. Biophys. Res. Commun., 191, 89–94). Ferner
clonierten die hier genannten Erfinder einen Bereich mit mehr als
3,2 kb DNA, von dem erwartet wird, dass er die iNOS-Genpromotor-Region
enthält,
bestimmten die Nucleotidsequenz der flankierenden 5'-Region von etwa
1,5 kb und berichteten, dass die Sequenz wie auch die miNOS-Gensequenz
eine Consensussequenz enthält,
von der man glaubt, dass sie von Interferon (IFN)-γ und dem
Transkriptionsfaktor NF-κB reguliert
wird (Nunokawa, Y., et al., (1994) Biochem. Biophys. Res. Commun.
200, 802–807).
-
Danach
wurden 16 kb der flankierenden 5'-Region
des hiNOS-Gens unter Verwendung eines Reportergens untersucht. Das
Ergebnis zeigte, dass eine Region existiert, die für die Induktion
des hiNOS-Gens zwischen 3,8 kb und 16 kb stromaufwärts von
der Transkriptionsstartstelle notwendig ist (de Vera, M. E. et al., (1996)
Proc. Natl. Acad. Sci. USA 93, 1054–1059). Jedoch betrug die Induktionsfähigkeit
immer noch ungefähr das
10-Fache, und die Expression ohne Stimulierung war ziemlich stark.
Daher liefert dieser Bericht eine ungenügende Erklärung für die tatsächliche und starke Induktion
des iNOS-Gens. Außerdem
wurde gezeigt, dass die Region bis zu 3,7 kb stromaufwärts von
der Transkriptionsstartstelle des hiNOS-Gens an der Induktion der
Genexpression nicht beteiligt ist (Laubach, V. E. et al., (1994)
Abstract Book of the 1st International Conference of Biochemistry
and Molecular Biology of Nitric Oxide, (UCLA Sunset Village, Los
Angeles, KA, USA), A16, Kleinert, H. et al., (1996) Mol. Pharmacol.
49, 15–21).
-
Die
hier genannten Erfinder erzeugten Zellen, die mit einem Plasmid
transfiziert wurden, das etwa 3,2 kb der flankierenden 5'-Region des hiNOS-Gens
enthält,
das früher
cloniert worden war und das an ein Reportergen gebunden ist. Die
Ergebnisse zeigten im Gegensatz zu denjenigen, über die unter Verwendung der
miNOS-Promoterregion berichtet wurden, aber ähnlich zu denjenigen, über die
von anderen Gruppen, die das hiNOS-Gen verwenden, berichtet wurden,
dass das Reportergen sogar unter nicht-induzierenden Bedingungen
exprimiert wurde, wenn die flankierende 5'-Region von etwa 3,2 kb verwendet wird,
und dass die Induzierbarkeit durch Cytokine entweder fehlt oder
sehr schwach ist (Nunokawa, Y. et al., (1996) Biochem. Biophys. Res.
Commun. 223, 347–352).
Außerdem
wurde die Reportergenexpression sogar in mit dem vorstehenden Plasmid
transfizierten Zellen nachgewiesen, die keine hiNOS-Induktion durch
verschiedene Cytokine zeigen (Nunokawa, Y. et al., (1996) Biochem.
Biophys. Res. Commun. 223, 347–352).
D.h., man fand heraus, dass die Kontrolle der hiNOS-Gen-Expression
durch Induktoren wie Cytokine nicht ausschließlich basierend auf der Promotorregion
erklärt
werden kann, die von den berichteten Ergebnissen über das
miNOS-Gen abgeleitet werden kann. Jedoch wurde der Grund dafür bisher überhaupt
nicht aufgeklärt.
-
JP 62 286 949 offenbart
ein Phenylmethylbenzochinonderivat der in der nachstehenden Beschreibung offenbarten
Formel als Beispiel einer Verbindung, die die Aktivität einer
Verbindung supprimieren kann, welche die hiNOS-Genexpression induziert.
Dieser Verbindung aus
JP 62
286 949 wurde eine schützende
Wirkung für
das Gehirn bei cerebraler Anoxie zugeschrieben.
-
Die
Aufgabe der vorliegenden Erfindung besteht darin, ein Verfahren
zum Screenen einer Verbindung bereitzustellen, welche die Expression
des hiNOS-Gens supprimieren
oder induzieren kann, indem der/die Bereiche) des hiNOS-Gens identifiziert
wird/werden, der/die für
die Kontrolle der Expression durch induzierende Faktoren notwendig
ist/sind, der Expressionsvektor, der den/die Bereiche) enthält, konstruiert
wird und der Vektor für
das Screenen genutzt wird.
-
Daher
betrifft die vorliegende Erfindung in einem Aspekt eine DNA, die
die flankierende 3'-Region
des menschlichen induzierbaren Stickstoffoxid-Synthase (hiNOS)-Gens umfasst, wobei
die flankierende 3'-Region hilft,
die hiNOS-Genexpression zu regulieren, und die Nucleotidsequenz
von SEQ ID NO: 16 beinhaltet, und in einem zweiten Aspekt einen
Expressionsvektor, der die DNA und die 3'-UTR des hiNOS-Gens umfasst, wobei die
DNA und die 3'-UTR
des hiNOS-Gens gemeinsam wirken, um die hiNOS-Genexpression zu regulieren.
-
Wenn,
wie vorstehend beschrieben, die stromaufwärts liegende Region von der
Transskriptionsstartstelle des hiNOS-Gens, d.h. die flankierende
5'-Region, stromaufwärts von
einem Reportergen eingebaut wird, wird die Reportergenexpression
unerwartet und unter nicht induzierenden Bedingungen beobachtet.
Basierend auf diesem Befund nahmen wir an, dass die Möglichkeit
bestand, dass Regionen, die nicht die flankierende 5'-Region sind, die
hiNOS-Genexpression kontrollieren könnten, und führten Experimente
durch.
-
Als
Ergebnis der Untersuchung der Sequenz der nicht-translatierten 3'-Region (UTR) des
hiNOS-Gens (SEQ ID NO:1) fanden die hier genannten Erfinder die „AUUUA-Sequenz", von der berichtet
wurde, dass sie eine Rolle bei der Destabilisierung der mRNA spielt,
an vier verschiedenen Stellen innerhalb der 3'-UTR des hiNOS-Gens (2,
unterstrichene Positionen). Daher zogen die hier genannten Erfinder
in Betracht, dass ein bestimmtes Element, das die Transkriptionsprodukte
des Gens destabilisiert, die Induktion der hiNOS-Genexpression kontrollieren
könnte.
-
Viele
Reportergene, die heute häufig
verwendet werden, werden durch Einbau einer von SV40 stammenden
Region mit einer Poly A-Addition
konstruiert, um die reife mRNA zu exprimieren. Die bisher verwendeten
Plasmide, um die hiNOS-Genexpression und das „pGL3- Grundplasmid" zu bestätigen, das die hier genannten
Erfinder in den früheren
Berichten verwendeten, werden durch Einbau der von SV40 stammenden Region
mit einer Poly A-Addition (SEQ ID NO:2) stromabwärts vom Reportergen konstruiert
(3). Falls die Induktion des Testgens nicht vollständig von
der Aktivierung der Transkription abhängt, sondern auch von der Instabilität der produzierten
mRNA verursacht wird, ist es daher notwendig, die Region, die an
der Instabilität der
produzierten mRNA beteiligt ist, in den Expressionsvektor einzubauen,
der das Reportergen enthält.
-
Da
in Betracht gezogen wurde, dass die hohe Induzierbarkeit, die inhärent beobachtet
werden musste, nicht umgesetzt werden konnte, indem das Reportersystem
eingeführt
wurde, das nur die flankierende 5'-Region
einschließlich
des Promotors enthält,
bauten die hier genannten Erfinder zusätzlich ein Genfragment mit etwa
1 kb, das die 3'-UTR
und die flankierende 3'-Region
des hiNOS-Gens enthält,
stromabwärts
des Reportergens ein. Folglich fand man heraus, dass diese Bereiche
gemeinsam mit der Promotoregion wirken, so dass eine starke Induktion
erzeugt wird. D.h., durch Einbauen dieser Regionen fand man heraus,
dass die Reportergenexpression unter nicht induzierenden Bedingungen
eliminiert wird und nur aktiviert wird, wenn man Cytokine wirken
lässt.
Diese Ergebnisse zeigten die Existenz der für die Induktion des hiNOS-Gens durch Cytokine
notwendigen Regionen, die unbekannt gewesen waren.
-
Ferner
konstruierten die hier genannten Erfinder ein Plasmid, das die genomische
Struktur des hiNOS-Gens nachahmt, mit der Ausnahme, dass der offene
Leserahmen (ORF) des hiNOS-Gens durch den ORF des Reportergens ersetzt
wurde. Genauer gesagt, wurde die hiNOS-Gen-Promotoregion stromaufwärts eingebaut,
und das Genfragment mit etwa 1 kb, das die 3'-UTR und die flankierende 3'-Region des hiNOS-Gens
enthält,
wurde stromabwärts
des Reportergens eingebaut. Durch Verwendung der menschlichen, mit
dem Plasmid transformierten Zelllinie entwickelten die hier genannten
Erfinder ein Verfahren zum Screenen von Verbindungen, welche die
hiNOS-Genexpression schnell mit hoher Sensitivität kontrollieren, so dass die
vorliegende Erfindung vollendet wird.
-
Genauer
gesagt, schließt
die vorliegende Erfindung jede Erfindung ein, die in den Ansprüchen dieser Beschreibung
beschrieben wird.
-
Die
DNAs, welche die flankierende 5'-Region,
die 5'-UTR, die
3'-UTR und die flankierende
3'-Region des hiNOS-Gens
ausmachen und in der vorliegenden Erfindung verwendet werden, schließen nicht
nur diejenigen ein, die aus menschlichen Zellen isoliert werden,
sondern auch diejenigen, die synthetisch hergestellt werden. Außerdem können diese
DNAs mit chemischen Modifizierungen oder mit durch Substitution,
Deletion oder Addition veränderten
Basen ebenfalls verwendet werden, solange ihre Fähigkeit zur Kontrolle der Expression
beibehalten wird. Die in der vorliegenden Erfindung verwendete DNA,
die die flankierende 3'-Region
des hiNOS-Gens ausmacht, umfasst diejenige, welche die Sequenz besitzt,
die in SEQ ID NO: 16 beschrieben wird (die Sequenz außerhalb
der Umrandung in 7).
-
Ein
Gen, das irgendein Peptid oder Protein codiert, kann als Reportergen
der vorliegenden Erfindung verwendet werden, solange ein Fachmann
die Aktivität
oder die produzierte Menge seines Expressionsprodukts (einschließlich der
Menge der produzierten mRNA) messen kann. Beispielsweise können Chloramphenicol-Acetyltransferase
(CAT), β-Galactosidase
(β-Gal)
und Luciferase verwendet werden, indem ihre enzymatischen Aktivitäten gemessen
werden. Auch kann das sekretierte Wachstumshormon oder dergleichen
verwendet werden, indem seine produzierte Menge durch solche Mittel
wie die immunologische Antikörperreaktion
gemessen wird.
-
Der
Vektor in der vorliegenden Erfindung, der die hiNOS-Genexpressionskontrollsequenz
enthält, kann
erhalten werden, indem die Expressionskontrollsequenz in einen replizierbaren
Vektor eingebaut wird. Beispiele des replizierbaren Vektors schließen pUC18
und pGEM-3Z ein, von denen bekannt ist, dass sie in E. coli replizierbar
sind.
-
Beim
Screenen von Substanzen, welche die hiNOS-Genexpression nach der
vorliegenden Erfindung beeinflussen können, kann man die Zellen,
die mit dem Vektor transformiert sind, der die hiNOS-Genexpressions-Kontrollsequenzen
der vorliegenden Erfindung enthält,
oder die natürlich
vorkommenden Zellen verwenden, die die in der vorliegenden Erfindung
beschriebenen Expressionskontrollsequenzen besitzen und die hiNOS-Genexpression kontrollieren
können.
Säuger-Zellen
werden vorzugsweise als derartige Zellen verwendet. Die Transformation
kann nach dem herkömmlichen
Verfahren erfolgen. Die transformierten Zellen der vorliegenden
Erfindung können
entweder diejenigen sein, die den Vektor permanent in die Wirtschromosomen
integriert haben, oder diejenigen, bei denen der Vektor transient
in den Wirt eingebaut ist. Die Zellen mit dem permanent in die Wirtschromosomen
integrierten Vektor können
selektiert werden, indem die Wirtszellen mit dem einzuführenden
Vektor, der ein Selektionsmarkergen enthält, oder mit dem einzuführenden
Vektor zusammen mit einem anderen Vektor transfiziert werden, der
einen Selektionsmarker enthält,
und die Zellen in einem Medium gezüchtet werden, das zulässt, dass
nur jene Zellen überleben,
in die der Selektionsmarker eingebaut wird.
-
Die
Substanzen, welche die hiNOS-Genexpression nach der vorliegenden
Erfindung beeinflussen können,
können
beispielsweise durchmustert werden, indem den transformierten, für einen
spezifizierten Zeitraum gezüchteten
Zellen eine beliebige Menge der Testsubstanz zugegeben wird und
die Menge des Reportergenexpressionsprodukts nach einem spezifizierten
Zeitraum als Enzymaktivität
oder der Menge des exprimierten Proteins gemessen wird. Die Testsubstanz
kann natürlich
oder synthetisch sein. Sie kann auch eine einzelne Substanz oder
ein Gemisch von Substanzen sein. Es ist beispielsweise möglich, eine
einzelne Kandidatensubstanz unabhängig oder ein Gemisch mehrerer
Kandidatensubstanzen zu testen. Außerdem ist es auch möglich, kombinatorische
Genbanken zu testen. Ferner können
auch Fraktionen eines Gemisches wie eines Zellextrakts getestet
werden und wiederholt fraktioniert werden, um letztendlich die Substanz
zu isolieren, die für
die Beeinflussung der hiNOS-Genexpression verantwortlich ist.
-
Andererseits
können
nach der vorliegenden Erfindung Substanzen durchmustert werden,
welche die Aktivität
einer bestimmten Substanz ändern
können,
welche die hiNOS-Genexpression beeinflusst, indem beispielsweise
den transformierten, für
einen spezifizierten Zeitraum gezüchteten Zellen eine beliebige
Menge der Kandidatensubstanz zugegeben wird und die Änderungen
der Menge des Reportergenexpressionsprodukt nach einem spezifizierten
Zeitraum als die Änderungen
der Enzymaktivität
oder der exprimierten Proteinmenge gemessen werden. Die zu durchmusternden
Kandidatensubstanzen werden definiert, wie vorstehend für die Substanzen,
welche die hiNOS-Genexpression
beeinflussen, beschrieben.
-
Die
Wirksamkeit einer Substanz, die durch das Screening-Verfahren oder
den Screening-Kit der vorliegenden Erfindung erhältlich ist und die hiNOS-Expression
supprimieren kann, oder einer Verbindung, die durch das Screening-Verfahren
oder den Screening-Kit der vorliegenden Erfindung erhältlich ist
und die Aktivität
der Verbindung unterdrücken
kann, welche die hiNOS-Genexpression induziert, kann gemessen werden, indem
die Verringerung der NO-Exkretion in das Kulturmedium, in Blut oder
Urin bei der Zugabe oder Verabreichung der Substanz unter den Bedingungen
bestimmt wird, die den Zellen, die Säuger-iNOS exprimieren können, oder
ihren lebenden Geweben erlauben, NO zu produzieren. Die Menge des
produzierten NO kann durch die im Allgemeinen bekannten Verfahren
gemessen werden, die durch das Griess-Verfahren (Green, L. C. et
al., (1982) Anal. Biochem. 126, 131–138), das in Beispiel 5-5
beschrieben wird, repräsentiert
werden.
-
Die
Verbindungen, die durch das Screening-Verfahren oder den Screening-Kit
der vorliegenden Erfindung erhältlich
sind, wie vorstehend beschrieben, sind diejenigen, die die Menge
des exprimierten hiNOS-Gens beeinflussen können, beispielsweise diejenigen,
welche die hiNOS-Genexpression
supprimieren können,
oder diejenigen, die die Aktivität
einer bestimmten Verbindung verändern
können,
welche die Menge des exprimierten hiNOS-Gens beeinflusst, beispielsweise
diejenigen, welche die Aktivität
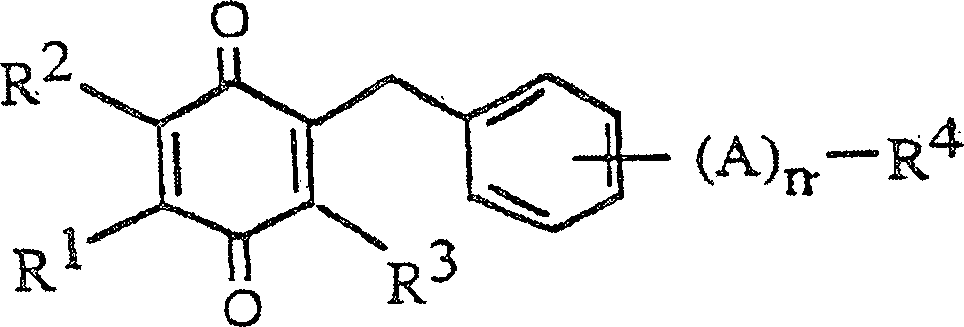
einer Verbindung supprimieren, welche die hiNOS-Genexpression induziert. Ein spezifisches
Beispiel der Verbindung, welche die Aktivität einer Verbindung supprimieren
kann, die die hiNOS-Genexpression
induziert, ist die Verbindung, die durch die folgende Strukturformel
repräsentiert
ist,
wobei, R
1,
R
2 und R
3 jeweils
unabhängig
ein Wasserstoffatom, eine Methylgruppe oder eine Methoxygruppe sind;
A eine Ethylengruppe oder eine Vinylengruppe ist; n 0 oder 1 ist
und R
4 ein Wasserstoffatom, eine Hydroxymethylgruppe
oder eine Carboxylgruppe ist, die verestert oder amidiert sein kann.
Genauer gesagt schließen
derartige Verbindungen die Verbindung ein, die in Beispiel 7 der
vorliegenden Beschreibung erhalten wurde und durch die chemische
Formel repräsentiert
ist, wie in
17 dargestellt, (3-[4-(3,4-Dimethoxy-6-methyl-2,5-benzochinonylmethyl)phenyl]-1-thiomorpholino-1-oxopropan),
Ethyl-3-[3-(2,5-benzochinonylmethyl)phenyl]acrylat, 3-(2,5-Benzochinonylmethyl)benzylalkohol,
3-(2,5-Benzochinonylmethyl)-benzosäure, 3-[3-(2,5-Benzochinonylmethyl)phenyl]propionsäure, 3-[3-(2,5-Benzochinonylmethyl)phenyl]propylalkohol, 3-[3-(2,5-Benzochinonylmethyl)phenyl]-acrylsäure, 3-[4-(3,4-Dimethoxy-6-methyl-2,5-benzochinonylmethyl)phenyl]-propionsäure2,3-Dimethoxy-5-benzyl-6-methyl-1,4-benzochinon,
3-[4-(3,4-Dimethoxy-6-methyl-2,5-benzochinonylmethyl)phenyl]propanol
und Ethyl-3-[3-(3,4-dimethoxy-6-methyl-2,5-benzochinonylmethyl)phenyl]-propionat,
wobei weder diese Verbindung noch das Arzneimittel, das die Verbindung
umfasst, noch die Verwendung der Verbindung durch die vorliegende
Erfindung umfasst sind. Der Einschluss in die vorliegende Beschreibung
dient illustrativen Zwecken.
-
Da
die Verbindungen, die durch das Screening-Verfahren oder den Screening-Kit
der vorliegenden Erfindung erhältlich
sind, weder direkt noch indirekt die hiNOS-Genexpression supprimieren
können,
sind sie als Arzneimittel zur Behandlung pathologischer Zustände oder
Erkrankungen nützlich,
die mit der Abnormalität
der hiNOS-Expression assoziiert sind, vorzugsweise die pathologischen
Zustände
oder Erkrankungen, die mit einer übermäßigen Expression des hiNOS-Gens
einhergehen. Beispielsweise sind die Verbindungen als Arzneistoffe
zur Behandlung von Herz- und cerebrovaskulären Erkrankungen, ischämischen
Herzerkrankungen, septischem Schock, akuten Schmerzen, Rheuma, Arthritis,
Asthma, Immunschwäche,
viraler oder nicht-viraler Infektionen, Autoimmunerkrankungen, Demenz
und Krebs nützlich
(Cattell, V. und Jensen, A., Histochem. J. Bd. 27, S. 777–784, 1995;
Ogden, J. E. und Moore, P. K., TIBTECH Bd. 13, S. 70–78, 1995).
-
Wenn
die Verbindungen, die durch das Screening-Verfahren oder den Screening-Kit
der vorliegenden Erfindung erhältlich
sind, als Arzneimittel verwendet werden, können sie entweder oral in der
Darreichungsform wie Tabletten, Kapseln, Elixiren, Mikrokapseln
oder parenteral als Injektionen in Form von Lösungen in Wasser oder in anderen
pharmazeutisch verträglichen
Flüssigkeiten
oder Suspensionen verabreicht werden. Beispielsweise können die
Verbindungen in die vorstehenden Darreichungsformen formuliert werden,
indem sie mit physiologisch verträglichen Trägern, Geschmacksstoffen, Excipienten,
Stabilisatoren etc. in einer allgemein zulässigen Form gemischt werden.
Zusatzstoffe, die in Tabletten gemischt werden können, schließen Bindemittel
wie Gelatine, Quellmittel wie Maisstärke, Excipienten wie kristalline
Cellulose und Schmiermittel wie Magnesiumstearat ein. Im Falle von
Kapseln können
zusätzlich
zu den vorstehend beschriebenen Bestandteilen flüssige Träger enthalten sein. Sterile
Zusammensetzungen zur Injektion können ebenfalls in der herkömmlichen
Weise formuliert werden.
-
Als
Beispiel einer wässrigen
Lösung
zur Injektion wird eine isotonische, Glucose enthaltende Lösung aufgeführt, die
auch mit geeigneten Lösungsvermittlern
wie Polyethylenglykol kombiniert werden kann. Sie kann ferner Puffer,
Stabilisatoren, Konservierungsstoffe, Antioxidantien und Linderungsmittel
enthalten. Die so hergestellten Arzneimittel können beispielsweise an Menschen
und andere Säuger
verabreicht werden. Auch wenn die Dosis abhängig von den Symptomen und
dergleichen variieren kann, wird einem Erwachsenen im Allgemeinen
die Zubereitung mit etwa 0,01 mg bis etwa 100 mg, vorzugsweise etwa
0,1 mg bis etwa 50 mg, besonders bevorzugt etwa 1,0 mg bis etwa
25 mg täglich
im Falle der oralen Verabreichung verabreicht. Im Falle der parenteralen
Verabreichung, beispielsweise in Form einer Injektion, werden einem
Erwachsenen im Allgemeinen intravenös etwa 0,001 mg bis etwa 50
mg, vorzugsweise etwa 0,01 mg bis etwa 25 mg, stärker bevorzugt etwa 0,1 mg
bis etwa 10 mg täglich
verabreicht.
-
1 zeigt
die vollständige
Struktur des hiNOS-Strukturgens.
-
2 zeigt
die Nucleotidsequenz der nicht-translatierten 3'-Region (UTR) innerhalb von Exon 26
des hiNOS-Gens. Die Zahlen an der linken Seite stellen die Nucleotidnummern,
wobei die erste Base der Translationstartstelle als 1 betrachtet
wird, dar. Die ATTTA-Sequenz ist unterstrichen, während die
erwartete Poly A-Additionsstelle mit „↑" angegeben ist. Die fettgedruckten Buchstaben
geben das Stopp-Codon im hiNOS-Gen an. Die Nucleotidsequenzen, die
die Basis für
die Primer darstellten, wurden mit ihren Bezeichnungen und Pfeilen
gekennzeichnet.
-
3 zeigt
die Struktur des pGL3-Grundplasmids und die Nucleotidsequenz der
von SV40 stammenden Poly A-Additionsregion. Das „Poly A-Signal" und das mit der
Poly A-Addition assoziierte „G/T-Cluster" sind unterstrichen.
-
4 zeigt
die Nucleotidsequenzen der zur Konstruktion der Plasmide verwendeten
Primen.
-
5 ist
das Foto eines Southern-Blots der Fragmente aus der Restriktionsenzymspaltung
(gespalten mit EcoRI, HindIII und XhoI) des gereinigten Phagengenoms,
das die flankierende 5'-Region
des hiNOS-Gens enthält.
-
6 zeigt
die Nucleotidsequenzsubstitution, um eine NcoI-Schnittstelle in der DNA der flankierenden 5'-Region des hiNOS-Gens
zu erzeugen. Die fettgedruckte Base „T" wurde durch „CA" substituiert, um das mutierte Plasmid
zu konstruieren.
-
7 zeigt
die Nucleotidsequenzen der nicht-translatierten 3'-Region (UTR) und der flankierenden 3'-Region des hiNOS-Gens.
Die Zahlen an der linken Seite stellen die Nucleotidnummern, wobei
die erste Base der Translationstartstelle als 1 betrachtet wird,
dar. Der eingekreiste Anteil ist die 3'-UTR von Exon 26. Die Consensussequenz
für die
Poly A-Addition (YGTGTTYY) innerhalb der flankierenden 3'-Region ist unterstrichen.
Die fettgedruckten Buchstaben geben das Stopp-Codon im hiNOS-Gen
an. Die Nucleotidsequenzen, die die Basis für die Primer darstellten, wurden
mit ihren Bezeichnungen und Pfeilen gekennzeichnet.
-
8 zeigt
die Strukturen der Expressionsvektoren, die die nicht-strukturellen Regionen
des hiNOS-Gens enthalten.
-
9 zeigt
die Reaktion der A549-Zellen, die transient mit den Expressionsvektoren
transfiziert wurden, auf die Cytokinstimulation.
-
10 ist
das Foto eines Gel-Shift-Tests unter Verwendung der Kernextrakte
aus den mit Cytokinen stimulierten A549-Zellen.
-
11 zeigt
die Reaktion der A549-Zellen, die stabil mit dem Expressionsvektor
(hiNOSLuc) transfiziert wurden, auf die Cytokinstimulation.
-
12 zeigt
die Reaktion der A549-Zellen auf die Cytokinstimulation, die mit
dem Griess-Verfahren gemessen wurde.
-
13 zeigt
die Wirkungen von LPS, IL-6, cAMP und TPA auf die Reaktion der A549-Zellen,
die stabil mit dem Expressionsvektor (hiNOSLuc) transfiziert wurden,
auf die Cytokinstimulation.
-
14 zeigt
die Wirkungen des Proteinkinase-Inhibitors Staurosporin auf die
induzierte Genexpression durch Cytokine.
-
15 zeigt
die Wirkungen von Dexamethason auf die induzierte Genexpression
durch Cytokine.
-
16 zeigt
die Wirkungen des Proteinkinase-Inhibitors TLCK auf die induzierte
Genexpression durch Cytokine.
-
17 zeigt
die Strukturformel von Verbindung I, die unter Verwendung des Screening-Systems
der vorliegenden Erfindung entdeckt wurde.
-
18 zeigt
die Wirkungen von Verbindung I auf das Screening-System der vorliegenden
Erfindung.
-
19 ist
das Foto, das zeigt, dass Verbindung I die hiNOS-Expression auf
der mRNA-Ebene supprimiert.
-
Das
folgende Beispiel veranschaulicht die vorliegende Erfindung detaillierter,
soll jedoch nicht den Umfang der vorliegenden Erfindung einschränken.
-
[Beispiel 1] Konstruktion
der Expressionsvektoren
-
1) Clonieren der flankierenden
5'-Region von hiNOS
-
Eine
Sonde, die für
das Durchmustern von Clonen verwendet werden soll, die die flankierende
5'-Region von hiNOS
enthalten, wurde durch Plaquehybridisierung aus einer menschlichen
Genbank hergestellt, die aus 2,5 × 106 Phagen
bestand (Clontech, USA; eine Phagen-Bank, die in den EMBL3-Vektor eingebaut
ist).
-
Die
verwendete Sonde war die durch Polymerasekettenreaktion (PCR) amplifizierte
cDNA, wobei als Matrize verwendet wurde: die von Ratten stammende
iNOS-cDNA (VSM-NOS), die früher
von den hier genannten Erfindern isoliert worden war (Nunokawa,
Y. et al., (1993) Biochem. Biophys. Res. Commun. 191, 89–94), sowie
der SU802-Primer (der den Basen -138 bis -117 der Ratten-VSM-NOS-cDNA/SEQ ID NO:3
entspricht, 4) und der MI103-Primer (der
den Basen 168 bis 188 der Ratten-VSM-NOS-cDNA/SEQ ID NO: 4 entspricht, 4)
verwendet wurden. Die PCR erfolgte unter Verwendung der Taq-DNA-Polymerase
(Takara Shuzo) und eines beiliegenden Puffers. Diese cDNA besitzt
offensichtlich eine hohe Homologie zu der Nucleotidsequenz des 5'-Endes des hiNOS-Strukturgens. Die
Plaquehybridiserung erfolgte unter Verwendung des direkten ECL-DNA-Markierungs-Nachweissystems
(Amersham, U.K.) nach der beiliegenden Versuchsvorschrift.
-
Positive
Plaques wurden unter Verwendung des Qiagen-Lambda-Kits (Qiagen,
Deutschland) gereinigt und mit EcoRI gespalten. Die Southern-Blot-Analyse
zeigte, dass das DNA-Fragment mit etwa 5 kb mit der vorstehenden
Sonde hybridisierte (5).
-
Das
mit EcoRI gespaltene Fragment mit etwa 5 kb wurde in die EcoRI-Schnittstelle in
das pUC118-Plasmid (Takara Shuzo) cloniert, und ein mutiertes Plasmid,
in dem eine Base substituiert war, wie in 6 gezeigt
(SEQ ID NO:13), wurde hergestellt, um eine NcoI-Schnittstelle zu
erzeugen. Dieses Verfahren wurde unter Verwendung des ortsgerichteten
Mutagenese-Kits von Clontech nach der beiliegenden Versuchsvorschrift
durchgeführt.
Das so hergestellte, mutierte Plasmid wurde mit KpnI und NcoI gespalten
und das sich ergebende DNA-Fragment wurde im Versuch 3) nachstehend
verwendet.
-
2) Clonieren der hiNOS-3'-UTR und eines Genfragments
mit etwa 1 kb stromabwärts
davon, das die Region enthält
-
Die
in 1) verwendete menschliche Genombank wurde in 60 Unterpoole aufgeteilt
(etwa 40.000 Clone pro Pool) und der die 3'-UTR enthaltende Pool wurde durch PCR
identifiziert, wobei zwei Arten sowohl von Vorwärtsprimern (SA101/SEQ ID NO:5,
SA102/SEQ ID NO:7) als auch Rückwärtsprimern
(KI101/SEQ ID NO:6, KI102/SEQ ID NO:8) verwendet wurden, die basierend
auf der hiNOS-cDNA-3'-UTR-Sequenz
(das 3'-Ende von
Exon 26 in 1 und innerhalb des eingekreisten
Anteils von 7) konstruiert wurden. Der positive
Pool wurde weiter in 30 Unterpoole aufgeteilt und der Pool, der
die positiven Clone enthielt, wurde genauso wie vorstehend durch
PCR identifiziert. An diesem Punkt schätzte man, dass ein Pool nicht
weniger als 1.000 Clone enthielt. Die Phagenpopulation, die alle
Clone in dem identifizierten positiven Unterpool enthielt, wurde
gezüchtet,
und die Phagen-DNA wurde unter Verwendung des Qiagen-Lambda-Kits
gereinigt. Anhand des Ergebnisses, dass die DNA mit SA101 and KI101
amplifiziert wurde, wurde bestätigt,
dass die Clone die hiNOS 3'-UTR
enthalten. Die Bestätigung
der PCR-Ergebnisse erfolgte auf dieselbe Weise wie in 1).
-
Dann
wurde die stromabwärts
liegende Region, die die 3'-UTR
enthält,
unter Verwendung der gereinigten DNA als Matrize und eines Vorwärtsprimers
(SA101) und eines Rückwärtsprimers
(KI103/SEQ ID NO: 9, 4), der basierend auf der Sequenz
des SP6-Promotoranteils des rechten EMBL3-Arms des Phagenvektors
konstruiert wurde, durch PCR amplifiziert. Die Sequenz der amplifizierten
DNA wurde mit dem Dye-Deoxyterminator-Kit (Applied Biosystems, USA)
durch das direkte Sequenzierverfahren unter Verwendung des Applied-Biosystems-A373-DNA-Sequenziergeräts bestimmt.
Die Ergebnisse sind als SEQ ID NO:14 und 7 dargestellt.
-
Es
wurde bestätigt,
dass die amplifizierte DNA ein Bereich ist, der die hiNOS-3'-UTR enthält. Basierend auf dieser Information
wurde die Poly A-Additionsstelle bestimmt, und ein Rückwärtsprimer,
der ein Ende des 3'-UTR
(KI104/SEQ ID NO: 11, 4) enthält, wurde hergestellt. Der
KI105-Rückwärtsprimer
(SEQ ID NO:12, 4 und 7) wurde
auch basierend auf der Sequenz der flankierenden 3'-Region hergestellt.
Die 3'-UTR DNA mit
etwa 0,5 kb wurde unter Verwendung der gereinigten DNA als Matrize
sowie von SA103 (SEQ ID NO: 10) und von KI104 durch PCR amplifiziert.
Da SA103 eine NheI-Erkennungsstelle an seinem 5'-Ende enthält und KI104 eine SalI-Erkennungsstelle
an seinem 5'-Ende
enthält,
wurde die vorstehende DNA mit SalI und NheI gespalten, und das sich
ergebende DNA-Fragment
(3'-UTR) wurde in
den Experimenten in 3) nachstehend verwendet.
-
Bei
Verwendung der gereinigten DNA als Matrize sowie von SA103 und von
KI105 wurde die DNA mit etwa 1 kb, die die 3'-UTR und die flankierende 3'-Region enthält, durch
PCR amplifiziert. Da SA103 eine NheI-Erkennungsstelle an seinem
5'-Ende enthält und KI105
eine BamHI-Erkennungsstelle an seinem 5'-Ende enthält, wurde das vorstehende DNA-Fragment
mit BamHI und NheI gespalten, und das sich ergebende Fragment (das
die 3'-UTR und die
flankierende 3'-Region
enthält)
wurde in den Experimenten in 3) nachstehend verwendet.
-
3) Konstruktion von Expressionsvektoren
die die nicht-strukturellen Regionen des hiNOS-Gens enthalten
-
- (3-A) Konstruktion des Plasmids (pGLNOS5 +
SV3) bei dem die flankierende 5'-Region und die 5'-UTR stromaufwärts des
Luciferase-Gens des pGL3-Grundplasmids
eingebaut ist.
Das pGL3-Grundplasmid (Promega, USA) wurde mit
den Restriktionsenzymen KpnI und NcoI gespalten, und das in 1) vorstehend
hergestellte DNA-Fragment wurde darin eingebaut (8A).
- (3-B) Konstruktion des Plasmids (pGLNOS3A), in dem die Region,
die sich unmittelbar stromabwärts
des Luciferase-Stoppcodons bis zur von SV40 stammenden PolyA-Additionssignalregion
des pGL3-Grundplasmid erstreckt, deletiert wurde und ein etwa 1
kb-Fragment, das die 3'-UTR
und die die flankierende 3'-Region
des hiNOS-Gens enthält,
dafür unmittelbar
stromabwärts
des Luciferase-Gens
in das pGL3-Grundplasmid eingebaut wurde Das pGL3-Grundplasmid wurde
mit BamHI und XbaI gespalten und das in 2) vorstehend hergestellte
DNA-Fragment (flankierende 3'-Region)
wurde darin eingebaut (8B).
- (3-C) Konstruktion des Plasmids (pGLNOS53A), in dem die flankierenden
5'-Region und die
5'-UTR des hiNOS-Gens
stromabwärts
des Luciferase-Gens in das pGL3-Grundplasmid
eingebaut wurden, die Region, die sich unmittelbar stromabwärts des Luciferase-Stoppcodons
bis zur von SV40-stammenden PolyA-Additionssignalregion des pGL3-Grundplasmid
erstreckt, deletiert wurde und ein etwa 1 kb-Fragment, das die 3'-UTR und die die
flankierende 3'-Region
des hiNOS-Gens enthält,
dafür eingebaut
wurde
-
Das
pGL3-Grundplasmid wurde mit BamHI und XbaI gespalten, und das in
2) vorstehend hergestellte DNA-Fragment (enthaltend die 3'-UTR und die flankierende
3'-Region) wurde
darin eingebaut. Das sich ergebende Plasmid wurde mit den Restriktionsenzymen
KpnI und NarI gespalten, und das in 1) vorstehend hergestellte DNA-Fragment
wurde darin eingebaut (8C).
- (3-D) Konstruktion des Plasmids (pGLNOS53 – SV3),
in dem die Region, die sich unmittelbar stromabwärts des Luciferase-Stoppcodons
bis zur von SV40 stammenden PolyA-Additionssignalregion des pGLNOS5
+ SV3-Plasmids erstreckt, das in (3-A) vorstehend hergestellt wurde,
deletiert wurde und die 3'-UTR
des hiNOS-Gens unmittelbar stromabwärts des Luciferase-Gens dafür eingebaut
wurde Das pGL3-Grundplasmid wurde mit den Restriktionsenzymen XbaI
und SalI gespalten, das DNA-Fragment (3'-UTR) wurde darin eingebaut und es wurde
mit KpnI und NarI gespalten. Das durch die Spaltung von pGLNOS5
+ SV3 mit KpnI und NarI erhaltene DNA-Fragment wurde dann in dieses
Plasmid eingebaut (8D).
- (3-E) Konstruktion des Plasmids (pGLNOS53 – SV3), in dem die 3'-UTR des hiNOS-Gens
unmittelbar stromabwärts
des Luciferase-Gens in das pGLNOS5 + SV3-Plasmid, das in (3-A) vorstehend hergestellt wurde,
eingebaut wurde Das in (3-D) vorstehend hergestellte pGLNOS53 – SV3-Plasmid
wurde mit dem Restriktionsenzym SalI gespalten, glatte Enden wurden
mit dem DNA-Blunting-Kit (Takara Shuzo) hergestellt, und es wurde
mit NarI gespalten. Das DNA-Fragment wurde durch Spaltung des pGLNOS5
+ SV3-Plasmids mit XbaI hergestellt, glatte Enden wurden mit dem
DNA-Blunting-Kit (Takara Shuzo) hergestellt, gefolgt von einer Spaltung
mit NarI. Das sich ergebende DNA-Fragment wurde in das vorstehende Plasmid
eingebaut (8E).
-
[Beispiel 2] Reaktion
der A549-Zellen, die transient mit den Expressionsvektoren transfiziert
wurden, auf die Cytokinstimulation
-
Eine
von einem menschlichen Lungenkrebs stammende Zelllinie, A549, von
der bekannt ist, dass sie hiNOS als Reaktion auf die Cytokinstimulation
exprimiert, wurde von der ATCC (Katalog #CCL185) erhalten und in
GIT-Medium (Wako Pure Chemical Industries, Ltd.) in einem Inkubator
in einer Atmosphäre
mit 5% CO2 gezüchtet. Die in 8 dargestellten
Expressionsvektoren wurden jeweils transient in die A549-Zellen
unter Verwendung von LipofectAMINE (Life Technologies Inc., USA)
transfiziert. Sechs Stunden nach Zugabe von entweder menschlichem
IL-1β (1
ng/ml) oder menschlichem IL-1β (1
ng/ml) + menschlichem IFN-γ (1.000 U/ml)
+ menschlichem TNF-α (500
ng/ml) (dieses Gemisch wird hier manchmal nachstehend als „CM" bezeichnet) wurde
die Reaktion untersucht (9). Die Luciferase-Aktivität wurde
nach der Vorschrift für
das Luciferase-Test-System (Promega) gemessen. Für den Nachweis wurde das ARGUS-50-Luminometer
(Hamamatsu Photonics) verwendet. Menschliches IL-1β wurde von
Genzyme (USA) bezogen. Menschliches IFN-γ und der menschliche TNF-α wurden am
Suntory Institute for Biomedical Research durch das herkömmliche Verfahren
hergestellt. Um den Test in diesem Experiment zu normalisieren,
wurde ein β-Gal-Expressionsvektor (pSV-β-gal, Promega)
als Kontrollvektor ebenfalls transfiziert, und die Ergebnisse wurden
extrapoliert.
-
Wenn
pGLNOS5 + SV3 eingeführt
wurde, wurde eine Expression in hohem Ausmaß ohne die Stimulierung beobachtet,
und die starke Induktion durch Cytokine wurde nicht beobachtet.
Wenn pGLNOS53 + SV3 eingeführt
wurde, war das Ergebnis dasselbe wie in dem Fall, in dem pGLNOS5
+ SV3 eingeführt
wurde. Außerdem
wurde kein Anstieg der Reporterexpression sowohl in dem Fall beobachtet,
wenn keine Stimulierung erfolgte, als auch in dem Fall, wenn eine
Stimulierung mit dem Cytokin erfolgte, wenn pGLNOS53 – SV3 eingeführt wurde.
Andererseits, wenn pGLNOS53A (das Plasmid, das die 3'-UTR und die flankierende
3'-Region des hiNOS-Gens
enthält)
verwendet wurde, trat ohne die Stimulierung fast keine Luciferase-Expression
auf, während
eine starke Expression bei Zugabe von IL-1β beobachtet
wurde und das Expressionsniveau durch die Zugabe von den drei Arten
von Cytokinen ein viel höheres
Ausmaß erreichte.
Man fand nämlich
heraus, das die Regulation der hiNOS-Genexpression die Gegenwart
von sowohl der 3'-UTR als auch der
flankierenden 3'-Region
erfordert.
-
[Beispiel 3] Die Gensequenz,
die wahrscheinlich an der hiNOS-Induktionsfähigkeit beteiligt ist
-
Mittels
des Gel-Shift-Verfahrens ist es möglich, die Bindung der Proteine
durch Cytokinstimulation an die doppelsträngige DNA (5'-AACTGTACACAAGCTGGGGACACTCCCTTTGGAAA-3'/SEQ ID NO:15) zu
beobachten, die den Nucleotiden -131 bis -97 stromaufwärts der
Transkriptionsstartseite des hiNOS-Gens entspricht. Die vorstehende
DNA-Sequenz enthält die Consensus-Sequenz,
von der man erwartet, dass NF-κB daran
bindet. Das Gel-Shift-Verfahren wurde durchgeführt, indem die vorstehende
doppelsträngige
DNA mit Digoxigenin (DIG) (unter Verwendung des Gel-Shift-Test-Kits, Boehringer
Mannheim, Deutschland) modifiziert wurde, mit den Kernextrakten
aus A549 inkubiert wurde und einer 7,5%-igen Polyacrylamidgelelektrophorese (Bio-Rad,
USA) bei 4°C
unterzogen wurde. Die Zellkernextrakte wurden von den Zellen, die
4 Stunden mit IL-1β (1
ng/ml) oder mit CM stimuliert wurden, oder aus den nicht-stimulierten
Zellen unter Verwendung des Verfahrens von Schreiber et al. (Schreiber,
E. et al., (1989) Nucleic Acids Res. 17, 6419) erhalten. Die von
dem Gel nach der Gelelektrophorese erhaltene DNA wurde durch das
Elektroblot-Verfahren auf die Zeta-Probe-Nylonmembran (Bio-Rad)
(unter Verwendung des von Atto hergestellten Gerätes) übertragen, und die DIG-markierte
DNA wurde mit einem chemilumineszierenden DIG-Erkennungsantikörper nachgewiesen.
-
10 zeigt
die Ergebnisse. Man fand heraus, dass ein Protein (A), das im A549-Kernextrakt
ohne Stimulierung vorliegt, an die vorstehende Sequenz bindet (was
der Bande A von 10 entspricht), und dass ein
anderes Bindeprotein (B), das im A549-Kernextrakt vorliegt, durch
die Stimulierung mit IL-1β oder CM
induziert wird, an die Sequenz zu binden (entsprechend der Bande
B von 10). Diese Ergebnisse zeigten, dass
die flankierende 5'-Region
des hiNOS-Gens sowie das miNOS-Gen die Region enthält, an die
NF-κB bei Cytokin-Stimulierung
bindet.
-
[Beispiel 4] Konstruktion
von A549-Zellen, die stabil mit Expressionsvektoren (A549/hiNOSLuc)
transfiziert wurden und ihre Reaktion auf die Cytokinstimulation
-
pGLNOS53A
und pSV2neo (Clontech) wurden gemeinsam in A549-Zellen unter Verwendung
von LipofectAMINE (Life Technologies Inc., USA) transfiziert, und
die Zellen, die stabil mit pGLNOS53A transfiziert waren (A549/hiNOSLuc), wurden
selektiert, indem G418-Sulfat (1 mg/ml, Life Technologies Inc.)
zum Medium zugegeben wurde.
-
Sechs
Clone aus mehreren G418-resistenten Zellen wurden beliebig ausgewählt. IL-1β (1 ng/ml)
oder CM wurde dazugegeben, und die Reaktion wurde nach 24 Stunden
mit den nicht-stimulierten Clonen verglichen. Als Ergebnis exprimierten
alle 6 Clone Luciferase nur, wenn sie mit Cytokinen stimuliert wurden.
Diese Daten zeigten, dass die Struktur des einzuführenden
Plasmids für
die Induktion der Genexpression wichtig ist, aber sie wird nicht
durch die Position des eingeführten
Gens auf dem Chromosom beeinflusst.
-
[Beispiel 5] Vergleich
der Reaktion von A549/hiNOSLuc auf die Cytokinstimulierung und des
hiNOS-Expressionsmusters
-
Ein
Clon (A5) wurde aus den in Beispiel 4 erhaltenen G418-resistenten
Zellen ausgewählt,
um die Induktion der Luciferase-Expression durch eine Vielfalt von
nachstehend aufgeführten
Substanzen zu untersuchen. (Es gibt keinen bestimmten Grund für die Wahl
von A5.) Die A5-Zellen wurden auf der Platte mit 96 Vertiefungen
bis etwa 10,000 Zellen/100μl
pro Vertiefung ausplattiert und 24 Stunden gezüchtet. Die folgenden Substanzen
wurden jeder Vertiefung zugegeben, und nach 6 oder 24 Stunden wurde
die Luciferaseaktivität nach
der Vorschrift des Luciferase-Testsystems
(Promega) gemessen. Das ARGUS-50-Luminometer (Hamamatsu Photonics)
wurde zum Nachweis verwendet.
-
1) IL-1β
-
Die
Luciferase-Expression wurde innerhalb einer mindestens 6-stündigen Stimulierung
mit IL-1β (1 ng/ml)
beobachtet. Jedoch änderte
dieselbe IL-1β-Menge
allein das Expressionsniveau sogar nach 24 Stunden Stimulierung
sowie nach 6 Stunden nicht (11).
-
2) IFN-γ
-
Die
Stimulierung mit IFN-γ (1.000
U/ml) verursachte keinen Anstieg der Expression (11).
-
3) TNF-α
-
Die
Stimulierung mit TNF-α (500
ng/ml) verursachte keinen Anstieg der Expression (11).
-
4) Kombination von Cytokinen
-
Wenn
die 3 in 1) bis 3) verwendeten Cytokinarten gemeinsam zugegeben
wurden (CM), wurde ein schwacher Anstieg der Expression innerhalb
von 6 Stunden beobachtet, während
eine sehr starke Expression nach 24 Stunden beobachtet wurde (11).
Das Expressionsniveau wird auf etwa 500-mal so hoch geschätzt wie
der Hintergrund (100 bis 200 Zählimpulse),
was mit dem Bericht übereinstimmt,
dass hiNOS von CM stark induziert wird. Außerdem induzierten IL-1β (1 ng/ml)
+ IFN-γ (1.000
U/ml) die Expression ebenfalls stark, und die 24-Stundenkultur in
Gegenwart der beiden Cytokine führte
zu einem stärkeren
Anstieg der Expression als von IL-1β (1 ng/ml) allein (11).
Sogar in Abwesenheit von IL-1β verursachte
die Stimulierung durch IFN-γ und
TNF-α ebenfalls
eine Induktion nach 24 Stunden.
-
5) Vergleich mit den unter
Verwendung des Griess-Verfahrens erhaltenen Ergebnissen
-
Das
Griess-Verfahren, das die Diazotierungsreaktion nutzt, ist als Verfahren
zur indirekten Überwachung
der Produktion von NO durch die Zelle bekannt (Green, L. C. et al.,
(1982) Anal. Biochem. 126, 131–138).
In dem Verfahren lässt
man das Griess-Reagenz, das ein Gemisch aus Naphthylethylendiamin (Kanto
Chemical, Tokio) und Sulfanilsäure
ist (Nacalai Tesque), mit dem NO2 –-Ion
im Kulturmedium reagieren, um die Farbentwicklung durch das Multititer-Plattenlesegerät (Molecular
Devices) durch Messung der Absorption bei 540 nm zu bestimmen. Wenn
das akkumulierte NO in der A549-Zellkultur 48 Stunden nach der Cytokinstimulierung
durch dieses Verfahren gemessen wurde, wurde die durch IL-1β (1 ng/ml)
oder CM stimulierte Produktion von NO beobachtet (12).
-
Diese
Ergebnisse entsprechen den Testsergebnissen von 1) bis 4), in denen
die Luciferase-Expression gemessen wurde. Daher wurde bestätigt, dass
die von hiNOS stammende NO-Produktion leicht durch den Nachweis
der Luciferase-Expression berechnet werden kann. (Die Sensitivität des auf
der Luciferase-Expression basierenden Verfahrens war um mehr als
das 100-Fache höher
als diejenige des Griess-Verfahrens.)
-
[Beispiel 6] Wirkungen
der Verbindungen, von denen erwartet wird, dass sie die iNOS-Expression
beeinflussen, auf dem Testsystem der vorliegenden Erfindung
-
1) Wirkungen der LPS (Lipopolysaccharide),
von IL-6, cAMP, TPA (Tetradecanoylphorbolacetat) und der Proteinkinase-Inhibitoren
-
Es
wurde berichtet, dass LPS (Sigma, USA), IL-6 (Genzyme), cAMP (Dibutyrylform,
Sigma) und TPA (Nacalai Tesque) iNOS in Versuchssystemen unter Verwendung
von Maus- oder Rattenzellen induzieren. Die Luciferase-Expression
wurde in Gegenwart dieser Substanzen auf dieselbe Weise gemessen
wie in Beispiel 5. Als Ergebnis beeinflussten diese Substanzen nicht
nur kaum die Kontrollspiegel, sondern auch kaum die Aktivierung
durch Cytokine (13). Diese Ergebnisse zeigten,
dass diese Substanzen das in Beispiel 5 beschriebene Testsystem
kaum beeinflussen. Andererseits fand man heraus, dass der Proteinkinase-Inhibitor Staurosporin
(Wako Pure Chemical Industries) die induzierte Genexpression durch
IL-1β (1
ng/ml) oder CM bei einer Konzentration von nicht mehr als 1 μM supprimierte
(14).
-
2) Wirkung von Glucocorticoid
-
Dexamethason
(Dex) (Wako Pure Chemical Industries) besitzt eine starke antiinflammatorische
Wirkung. Von einem seiner Mechanismen ist bekannt, dass er der Suppression
der Aktivierung des Transkriptionsfaktors NF-κB zugeschrieben werden kann
(Ray, A. und Prefontaine, K. E. (1994) Proc. Natl. Acad. Sci. USA
91, 752–756).
Es wurde auch berichtet, dass es die iNOS-Expression supprimiert
(Radomski, M.W. et al., (1990) Proc. Natl. Acad. Sci. USA 87, 10043–10047).
Die Luciferase-Expression wurde auf dieselbe Weise wie in Beispiel
5 in Gegenwart von Dexamethason gemessen, und man fand heraus, dass
die induzierte Genexpression durch IL-1β (1 ng/ml) oder CM bei einer
Konzentration von nicht mehr als 10 μM supprimiert wurde (15).
Zudem verschwand der supprimierende Effekt, wenn es nach 1 Stunde
der Cytokinstimulierung zugegeben wurde.
-
3) Wirkung der Protease-Inhibitoren
-
Es
wird in Betracht gezogen, dass Proteasen an der Aktivierung von
NF-κB durch
den Abbau von I-κB (ein
Protein, das die Aktivierung von NF-κB im Cytoplasma supprimiert;
Verma, I. M. et al., (1995) Genes & Dev. 9, 2723–2735) beteiligt sind. Von
einem der Protease-Inhibitoren TLCK (Nacalai Tesque) ist bekannt,
dass er die iNOS-Induktion sowie die NF-κB-Aktivierung hemmt (Griscavage,
J. M. et al., (1995) Biochem. Biophys. Res. Commun. 215, 721–729). Die
Luciferase-Expression wurde auf dieselbe Weise wie in Beispiel 5
in Gegenwart von TLCK gemessen. Man fand heraus, dass TLCK die durch
IL-1β (1
ng/ml) oder CM induzierte Genexpression bei einer Konzentration
von etwa 10 μM
supprimiert (16).
-
Diese
Versuchsergebnisse zeigten auch, dass ein System, wie in Beispiel
5 beschrieben, verwendet werden kann, um Verbindungen zu durchmustern,
die an der Kontrolle der hiNOS-Expression beteiligt sind.
-
[Beispiel 7] Screenen
nach Verbindungen, welche die hiNOS-Genexpression beinflussen
-
Wie
in Beispiel 5 wurden die A5-Zellen auf die Platte mit 96 Vertiefungen
mit etwa 10,000 Zellen/100 μl
pro Vertiefung ausplattiert und 24 Stunden gezüchtet. Anschließend wurden
spezifische Verbindungen, von denen nicht bekannt war, ob sie die
hiNOS-Genexpression beeinflussen oder nicht, entweder einzeln oder
als Gemische zugegeben, und die Zellen wurden eine weitere Stunde
gezüchtet.
Dann wurde dem Kulturmedium IL-1β oder
TNF-α zugegeben,
um die Luciferase-Aktivität
24 Stunden nach Zugabe von IL-1β oder
TNF-α auf dieselbe
Weise wie in Beispiel 6 zu messen.
-
Als
Ergebnis der Durchmusterung zahlreicher Verbindungen und Gemische
von Verbindungen wurde deutlich, dass die in 17 dargestellte
Verbindung I die Luciferase-Aktivität konzentrationsabhängig supprimieren
kann (18).
-
Ferner
wurde bestätigt,
dass die Verbindung I (20 μg/ml),
von der durch dieses Screening-Verfahren nachgewiesen wurde, dass
sie hiNOS-Genexpression supprimiert, die Expression der von CM stimulierten
hiNOS-mRNA in den Wildtyp-A549-Zellen
supprimiert (19). Andererseits beeinflusste
die Verbindung I mit der gleichen Konzentration nicht die in den
Wildtyp-A549-Zellen konstitutiv exprimierte β-Aktin-mRNA-Expression (19).
-
Es
ist auch möglich,
Verbindungen, die die hiNOS-Expression induzieren, durch ein ähnliches
wie das in Beispiel 7 beschriebene Verfahren zu durchmustern, indem
beispielsweise Testverbindungen zu A5-Zellen zugegeben werden und
die Verbindungen gewählt
werden, die die Luciferase-Aktivität erhöhen.
-
Daher
wurde gezeigt, dass es möglich
ist, mittels des Screening-Systems der vorliegenden Erfindung Verbindungen
zu durchmustern, die die hiNOS-Genexpression
beeinflussen.
-
Die
vorliegende Erfindung stellt ein Verfahren bereit, eine Verbindung
einfach und leicht zu durchmustern, die die Expression einer menschlichen
induzierbaren Stickstoffoxid (NO)-Synthase (hiNOS) mit hoher Sensitivität kontrollieren
kann.
-
Das
Verfahren der vorliegenden Erfindung ermöglicht auch das bequeme und
sensitive Screenen einer Verbindung, die zur Behandlung von Entzündungen
und Sepsis als nützlich
betrachtet wird, indem die hiNOS-Expression supprimiert wird, oder
einer Verbindung, die für
Antitumor-, antivirale und vaskuläre Restenose-Präventionsbehandlungen
als nützlich
betrachtet wird, indem die Expression induziert wird. Sequenzprotokoll
SEQUENZPROTOKOLL




 SEQUENZPROTOKOLL
SEQUENZPROTOKOLL