ES2720140T3 - Proteínas de fusión transportadoras de péptido como vacunas para alergias - Google Patents
Proteínas de fusión transportadoras de péptido como vacunas para alergias Download PDFInfo
- Publication number
- ES2720140T3 ES2720140T3 ES12729450T ES12729450T ES2720140T3 ES 2720140 T3 ES2720140 T3 ES 2720140T3 ES 12729450 T ES12729450 T ES 12729450T ES 12729450 T ES12729450 T ES 12729450T ES 2720140 T3 ES2720140 T3 ES 2720140T3
- Authority
- ES
- Spain
- Prior art keywords
- seq
- amino acids
- allergen
- der
- phl
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 229960005486 vaccine Drugs 0.000 title claims description 49
- 206010020751 Hypersensitivity Diseases 0.000 title claims description 42
- 230000007815 allergy Effects 0.000 title claims description 41
- 208000026935 allergic disease Diseases 0.000 title claims description 37
- 102000037865 fusion proteins Human genes 0.000 title description 97
- 108020001507 fusion proteins Proteins 0.000 title description 97
- 108010082406 peptide permease Proteins 0.000 title description 3
- 239000013566 allergen Substances 0.000 claims abstract description 211
- 108090000765 processed proteins & peptides Proteins 0.000 claims abstract description 191
- 102000004196 processed proteins & peptides Human genes 0.000 claims abstract description 159
- 230000027455 binding Effects 0.000 claims abstract description 96
- 229920001184 polypeptide Polymers 0.000 claims abstract description 73
- 101150115433 SLC26A5 gene Proteins 0.000 claims abstract description 69
- 210000001744 T-lymphocyte Anatomy 0.000 claims abstract description 65
- 230000000774 hypoallergenic effect Effects 0.000 claims abstract description 55
- 108010033276 Peptide Fragments Proteins 0.000 claims abstract description 36
- 102000007079 Peptide Fragments Human genes 0.000 claims abstract description 36
- 230000009257 reactivity Effects 0.000 claims abstract description 36
- 208000002672 hepatitis B Diseases 0.000 claims abstract description 23
- 230000002829 reductive effect Effects 0.000 claims abstract description 23
- 210000004899 c-terminal region Anatomy 0.000 claims abstract description 17
- 210000003719 b-lymphocyte Anatomy 0.000 claims abstract description 16
- 230000004044 response Effects 0.000 claims abstract description 16
- 238000004519 manufacturing process Methods 0.000 claims abstract description 12
- 125000000539 amino acid group Chemical group 0.000 claims abstract description 9
- 108010074328 Interferon-gamma Proteins 0.000 claims abstract description 7
- 102100037850 Interferon gamma Human genes 0.000 claims abstract description 6
- 230000001939 inductive effect Effects 0.000 claims abstract description 5
- 150000001413 amino acids Chemical class 0.000 claims description 76
- 239000000203 mixture Substances 0.000 claims description 57
- 229960004784 allergens Drugs 0.000 claims description 55
- 238000011282 treatment Methods 0.000 claims description 38
- 108010061608 Dermatophagoides pteronyssinus antigen p 2 Proteins 0.000 claims description 36
- 239000013598 vector Substances 0.000 claims description 28
- 229940046528 grass pollen Drugs 0.000 claims description 27
- 238000009472 formulation Methods 0.000 claims description 25
- 239000012634 fragment Substances 0.000 claims description 22
- 108010061629 Dermatophagoides pteronyssinus antigen p 1 Proteins 0.000 claims description 18
- 235000018185 Betula X alpestris Nutrition 0.000 claims description 17
- 235000018212 Betula X uliginosa Nutrition 0.000 claims description 17
- 108020004707 nucleic acids Proteins 0.000 claims description 14
- 102000039446 nucleic acids Human genes 0.000 claims description 14
- 150000007523 nucleic acids Chemical class 0.000 claims description 14
- 108010061638 Dermatophagoides pteronyssinus antigen p 7 Proteins 0.000 claims description 12
- 125000003275 alpha amino acid group Chemical group 0.000 claims description 12
- 206010048908 Seasonal allergy Diseases 0.000 claims description 10
- 239000000428 dust Substances 0.000 claims description 9
- 230000002265 prevention Effects 0.000 claims description 9
- 208000035285 Allergic Seasonal Rhinitis Diseases 0.000 claims description 8
- 241000282326 Felis catus Species 0.000 claims description 8
- 241001465754 Metazoa Species 0.000 claims description 8
- 239000013574 grass pollen allergen Substances 0.000 claims description 6
- -1 preferably Amb to 1 Substances 0.000 claims description 6
- 201000004338 pollen allergy Diseases 0.000 claims description 4
- 244000036975 Ambrosia artemisiifolia Species 0.000 claims description 2
- 235000003129 Ambrosia artemisiifolia var elatior Nutrition 0.000 claims description 2
- 240000005109 Cryptomeria japonica Species 0.000 claims description 2
- 208000035533 House dust allergy Diseases 0.000 claims description 2
- 102000011195 Profilin Human genes 0.000 claims description 2
- 108050001408 Profilin Proteins 0.000 claims description 2
- 241000238711 Pyroglyphidae Species 0.000 claims description 2
- 235000003484 annual ragweed Nutrition 0.000 claims description 2
- 235000006263 bur ragweed Nutrition 0.000 claims description 2
- 235000003488 common ragweed Nutrition 0.000 claims description 2
- 229940046533 house dust mites Drugs 0.000 claims description 2
- 235000009736 ragweed Nutrition 0.000 claims description 2
- 244000025254 Cannabis sativa Species 0.000 claims 1
- 240000007817 Olea europaea Species 0.000 claims 1
- 241001465379 Parietaria judaica Species 0.000 claims 1
- 101150108812 proC gene Proteins 0.000 claims 1
- 241000283973 Oryctolagus cuniculus Species 0.000 description 77
- 230000000172 allergic effect Effects 0.000 description 68
- 208000010668 atopic eczema Diseases 0.000 description 67
- 235000001014 amino acid Nutrition 0.000 description 54
- 108090000623 proteins and genes Proteins 0.000 description 43
- 102000004169 proteins and genes Human genes 0.000 description 38
- 235000018102 proteins Nutrition 0.000 description 36
- 210000002966 serum Anatomy 0.000 description 30
- 238000002965 ELISA Methods 0.000 description 27
- 239000000872 buffer Substances 0.000 description 26
- 210000004027 cell Anatomy 0.000 description 25
- 239000000427 antigen Substances 0.000 description 23
- 108091007433 antigens Proteins 0.000 description 23
- 102000036639 antigens Human genes 0.000 description 23
- 230000005764 inhibitory process Effects 0.000 description 23
- 230000000638 stimulation Effects 0.000 description 22
- 230000000694 effects Effects 0.000 description 20
- 239000002671 adjuvant Substances 0.000 description 19
- 210000003819 peripheral blood mononuclear cell Anatomy 0.000 description 19
- 238000012546 transfer Methods 0.000 description 19
- 238000005406 washing Methods 0.000 description 18
- 230000028993 immune response Effects 0.000 description 17
- 101100189913 Caenorhabditis elegans pept-1 gene Proteins 0.000 description 16
- 108010078791 Carrier Proteins Proteins 0.000 description 16
- 108010088535 Pep-1 peptide Proteins 0.000 description 16
- FAPWRFPIFSIZLT-UHFFFAOYSA-M Sodium chloride Chemical compound [Na+].[Cl-] FAPWRFPIFSIZLT-UHFFFAOYSA-M 0.000 description 16
- 230000002788 anti-peptide Effects 0.000 description 16
- 239000003814 drug Substances 0.000 description 16
- 238000003556 assay Methods 0.000 description 14
- 230000003053 immunization Effects 0.000 description 14
- 238000002649 immunization Methods 0.000 description 14
- 101100494726 Neurospora crassa (strain ATCC 24698 / 74-OR23-1A / CBS 708.71 / DSM 1257 / FGSC 987) pep-4 gene Proteins 0.000 description 13
- 230000006698 induction Effects 0.000 description 13
- 238000000034 method Methods 0.000 description 13
- 229910000162 sodium phosphate Inorganic materials 0.000 description 13
- 238000002255 vaccination Methods 0.000 description 13
- 208000037265 diseases, disorders, signs and symptoms Diseases 0.000 description 12
- 108010037248 lantibiotic Pep5 Proteins 0.000 description 12
- SRCAXTIBNLIRHU-JJKPAIEPSA-N lantibiotic pep5 Chemical compound N([C@@H](C)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](C)C(=O)N[C@H](CS)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCCN)C(=O)N\C(=C/C)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](C)C(=O)N\C(=C/C)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC=1C=CC=CC=1)C(=O)N\C(=C(/C)S)C(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CS)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(=O)NCC(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CC(N)=O)C(=O)NCC(=O)N[C@@H](CS)C(=O)N[C@@H](CCCCN)C(O)=O)C(=O)[C@@H]1CCCN1C(=O)CNC(=O)[C@H](C)NC(=O)C(=O)CC SRCAXTIBNLIRHU-JJKPAIEPSA-N 0.000 description 12
- 239000000047 product Substances 0.000 description 12
- 238000009169 immunotherapy Methods 0.000 description 11
- 238000011533 pre-incubation Methods 0.000 description 11
- 238000002415 sodium dodecyl sulfate polyacrylamide gel electrophoresis Methods 0.000 description 11
- IQFYYKKMVGJFEH-XLPZGREQSA-N Thymidine Chemical compound O=C1NC(=O)C(C)=CN1[C@@H]1O[C@H](CO)[C@@H](O)C1 IQFYYKKMVGJFEH-XLPZGREQSA-N 0.000 description 10
- 238000006243 chemical reaction Methods 0.000 description 10
- 101100476210 Caenorhabditis elegans rnt-1 gene Proteins 0.000 description 9
- 102000014914 Carrier Proteins Human genes 0.000 description 9
- KCXVZYZYPLLWCC-UHFFFAOYSA-N EDTA Chemical compound OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=O KCXVZYZYPLLWCC-UHFFFAOYSA-N 0.000 description 9
- 230000002009 allergenic effect Effects 0.000 description 9
- 238000002347 injection Methods 0.000 description 9
- 239000007924 injection Substances 0.000 description 9
- 230000003612 virological effect Effects 0.000 description 9
- 102000004127 Cytokines Human genes 0.000 description 8
- 108090000695 Cytokines Proteins 0.000 description 8
- 241000699666 Mus <mouse, genus> Species 0.000 description 8
- 241000699670 Mus sp. Species 0.000 description 8
- 241001494479 Pecora Species 0.000 description 8
- 102000007056 Recombinant Fusion Proteins Human genes 0.000 description 8
- 108010008281 Recombinant Fusion Proteins Proteins 0.000 description 8
- 230000037396 body weight Effects 0.000 description 8
- 201000010099 disease Diseases 0.000 description 8
- 239000011780 sodium chloride Substances 0.000 description 8
- 239000001488 sodium phosphate Substances 0.000 description 8
- 235000011008 sodium phosphates Nutrition 0.000 description 8
- 208000024891 symptom Diseases 0.000 description 8
- RYFMWSXOAZQYPI-UHFFFAOYSA-K trisodium phosphate Chemical compound [Na+].[Na+].[Na+].[O-]P([O-])([O-])=O RYFMWSXOAZQYPI-UHFFFAOYSA-K 0.000 description 8
- 241000700721 Hepatitis B virus Species 0.000 description 7
- 241000700605 Viruses Species 0.000 description 7
- 208000006673 asthma Diseases 0.000 description 7
- 230000000903 blocking effect Effects 0.000 description 7
- 239000006228 supernatant Substances 0.000 description 7
- 238000012360 testing method Methods 0.000 description 7
- 206010027654 Allergic conditions Diseases 0.000 description 6
- 108010061573 Dermatophagoides pteronyssinus antigen p 5 Proteins 0.000 description 6
- PEDCQBHIVMGVHV-UHFFFAOYSA-N Glycerine Chemical compound OCC(O)CO PEDCQBHIVMGVHV-UHFFFAOYSA-N 0.000 description 6
- 241000700739 Hepadnaviridae Species 0.000 description 6
- 239000000020 Nitrocellulose Substances 0.000 description 6
- WNROFYMDJYEPJX-UHFFFAOYSA-K aluminium hydroxide Chemical compound [OH-].[OH-].[OH-].[Al+3] WNROFYMDJYEPJX-UHFFFAOYSA-K 0.000 description 6
- IQFYYKKMVGJFEH-UHFFFAOYSA-N beta-L-thymidine Natural products O=C1NC(=O)C(C)=CN1C1OC(CO)C(O)C1 IQFYYKKMVGJFEH-UHFFFAOYSA-N 0.000 description 6
- 238000001516 cell proliferation assay Methods 0.000 description 6
- 238000001514 detection method Methods 0.000 description 6
- 238000010828 elution Methods 0.000 description 6
- 239000013604 expression vector Substances 0.000 description 6
- 230000004927 fusion Effects 0.000 description 6
- 230000002163 immunogen Effects 0.000 description 6
- 238000011534 incubation Methods 0.000 description 6
- 238000013411 master cell bank Methods 0.000 description 6
- 229920001220 nitrocellulos Polymers 0.000 description 6
- 230000003287 optical effect Effects 0.000 description 6
- 239000013612 plasmid Substances 0.000 description 6
- 230000035755 proliferation Effects 0.000 description 6
- 101000897042 Homo sapiens Nucleotide pyrophosphatase Proteins 0.000 description 5
- 102000008100 Human Serum Albumin Human genes 0.000 description 5
- 108091006905 Human Serum Albumin Proteins 0.000 description 5
- 102100021969 Nucleotide pyrophosphatase Human genes 0.000 description 5
- 230000008033 biological extinction Effects 0.000 description 5
- 210000004369 blood Anatomy 0.000 description 5
- 239000008280 blood Substances 0.000 description 5
- 238000005119 centrifugation Methods 0.000 description 5
- 238000000855 fermentation Methods 0.000 description 5
- 230000004151 fermentation Effects 0.000 description 5
- 239000000499 gel Substances 0.000 description 5
- 238000010348 incorporation Methods 0.000 description 5
- 230000001965 increasing effect Effects 0.000 description 5
- 230000001404 mediated effect Effects 0.000 description 5
- 239000002609 medium Substances 0.000 description 5
- 235000019799 monosodium phosphate Nutrition 0.000 description 5
- 238000002360 preparation method Methods 0.000 description 5
- AJPJDKMHJJGVTQ-UHFFFAOYSA-M sodium dihydrogen phosphate Chemical compound [Na+].OP(O)([O-])=O AJPJDKMHJJGVTQ-UHFFFAOYSA-M 0.000 description 5
- 230000001225 therapeutic effect Effects 0.000 description 5
- MTCFGRXMJLQNBG-REOHCLBHSA-N (2S)-2-Amino-3-hydroxypropansäure Chemical compound OC[C@H](N)C(O)=O MTCFGRXMJLQNBG-REOHCLBHSA-N 0.000 description 4
- QFVHZQCOUORWEI-UHFFFAOYSA-N 4-[(4-anilino-5-sulfonaphthalen-1-yl)diazenyl]-5-hydroxynaphthalene-2,7-disulfonic acid Chemical compound C=12C(O)=CC(S(O)(=O)=O)=CC2=CC(S(O)(=O)=O)=CC=1N=NC(C1=CC=CC(=C11)S(O)(=O)=O)=CC=C1NC1=CC=CC=C1 QFVHZQCOUORWEI-UHFFFAOYSA-N 0.000 description 4
- DWRXFEITVBNRMK-UHFFFAOYSA-N Beta-D-1-Arabinofuranosylthymine Natural products O=C1NC(=O)C(C)=CN1C1C(O)C(O)C(CO)O1 DWRXFEITVBNRMK-UHFFFAOYSA-N 0.000 description 4
- 241000238631 Hexapoda Species 0.000 description 4
- 241000282412 Homo Species 0.000 description 4
- 108010002350 Interleukin-2 Proteins 0.000 description 4
- 102000000588 Interleukin-2 Human genes 0.000 description 4
- 108091028043 Nucleic acid sequence Proteins 0.000 description 4
- XSQUKJJJFZCRTK-UHFFFAOYSA-N Urea Chemical compound NC(N)=O XSQUKJJJFZCRTK-UHFFFAOYSA-N 0.000 description 4
- 230000004913 activation Effects 0.000 description 4
- 238000007792 addition Methods 0.000 description 4
- 229940037003 alum Drugs 0.000 description 4
- 238000004458 analytical method Methods 0.000 description 4
- 230000003141 anti-fusion Effects 0.000 description 4
- 210000003651 basophil Anatomy 0.000 description 4
- 150000001875 compounds Chemical class 0.000 description 4
- 238000010276 construction Methods 0.000 description 4
- UQLDLKMNUJERMK-UHFFFAOYSA-L di(octadecanoyloxy)lead Chemical compound [Pb+2].CCCCCCCCCCCCCCCCCC([O-])=O.CCCCCCCCCCCCCCCCCC([O-])=O UQLDLKMNUJERMK-UHFFFAOYSA-L 0.000 description 4
- 238000010586 diagram Methods 0.000 description 4
- 238000010790 dilution Methods 0.000 description 4
- 239000012895 dilution Substances 0.000 description 4
- 239000000284 extract Substances 0.000 description 4
- 239000001963 growth medium Substances 0.000 description 4
- 229930027917 kanamycin Natural products 0.000 description 4
- 229960000318 kanamycin Drugs 0.000 description 4
- SBUJHOSQTJFQJX-NOAMYHISSA-N kanamycin Chemical compound O[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1N SBUJHOSQTJFQJX-NOAMYHISSA-N 0.000 description 4
- 229930182823 kanamycin A Natural products 0.000 description 4
- 230000001589 lymphoproliferative effect Effects 0.000 description 4
- 238000005259 measurement Methods 0.000 description 4
- 239000003147 molecular marker Substances 0.000 description 4
- 230000009467 reduction Effects 0.000 description 4
- 238000013207 serial dilution Methods 0.000 description 4
- 238000007920 subcutaneous administration Methods 0.000 description 4
- 238000002560 therapeutic procedure Methods 0.000 description 4
- 229940104230 thymidine Drugs 0.000 description 4
- QKNYBSVHEMOAJP-UHFFFAOYSA-N 2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chloride Chemical compound Cl.OCC(N)(CO)CO QKNYBSVHEMOAJP-UHFFFAOYSA-N 0.000 description 3
- 241000283707 Capra Species 0.000 description 3
- 108090000565 Capsid Proteins Proteins 0.000 description 3
- 102100023321 Ceruloplasmin Human genes 0.000 description 3
- 108020004414 DNA Proteins 0.000 description 3
- 238000012286 ELISA Assay Methods 0.000 description 3
- 241000588724 Escherichia coli Species 0.000 description 3
- 241000746983 Phleum pratense Species 0.000 description 3
- 229920002873 Polyethylenimine Polymers 0.000 description 3
- 229920001213 Polysorbate 20 Polymers 0.000 description 3
- 206010070834 Sensitisation Diseases 0.000 description 3
- 239000007983 Tris buffer Substances 0.000 description 3
- 229940074608 allergen extract Drugs 0.000 description 3
- 201000009961 allergic asthma Diseases 0.000 description 3
- 230000002547 anomalous effect Effects 0.000 description 3
- 238000000376 autoradiography Methods 0.000 description 3
- 230000015572 biosynthetic process Effects 0.000 description 3
- 238000010367 cloning Methods 0.000 description 3
- 235000018417 cysteine Nutrition 0.000 description 3
- 238000011161 development Methods 0.000 description 3
- 208000035475 disorder Diseases 0.000 description 3
- 238000011156 evaluation Methods 0.000 description 3
- 239000013613 expression plasmid Substances 0.000 description 3
- 238000010353 genetic engineering Methods 0.000 description 3
- 230000005847 immunogenicity Effects 0.000 description 3
- 239000007972 injectable composition Substances 0.000 description 3
- 239000000463 material Substances 0.000 description 3
- 239000002245 particle Substances 0.000 description 3
- 239000000546 pharmaceutical excipient Substances 0.000 description 3
- 239000000256 polyoxyethylene sorbitan monolaurate Substances 0.000 description 3
- 235000010486 polyoxyethylene sorbitan monolaurate Nutrition 0.000 description 3
- 239000013641 positive control Substances 0.000 description 3
- 238000001556 precipitation Methods 0.000 description 3
- 230000008313 sensitization Effects 0.000 description 3
- 238000012163 sequencing technique Methods 0.000 description 3
- 239000007929 subcutaneous injection Substances 0.000 description 3
- 238000010254 subcutaneous injection Methods 0.000 description 3
- PORMUFZNYQJOEI-UHFFFAOYSA-N sumatriptan succinate Chemical compound OC(=O)CCC(O)=O.CNS(=O)(=O)CC1=CC=C2NC=C(CCN(C)C)C2=C1 PORMUFZNYQJOEI-UHFFFAOYSA-N 0.000 description 3
- 239000000725 suspension Substances 0.000 description 3
- LENZDBCJOHFCAS-UHFFFAOYSA-N tris Chemical compound OCC(N)(CO)CO LENZDBCJOHFCAS-UHFFFAOYSA-N 0.000 description 3
- CQTGBCFGAAYOCY-ZCRNMIQFSA-N (3S)-3-[[(2S)-2-[[(2S)-2-[[(2S)-2-[[(2S,3R)-2-[[(2S)-2-[[(2S)-2-[[(2S)-2-acetamido-3-methylbutanoyl]amino]-3-methylbutanoyl]amino]-3-phenylpropanoyl]amino]-3-hydroxybutanoyl]amino]-3-hydroxypropanoyl]amino]-3-(1H-indol-3-yl)propanoyl]amino]-5-amino-5-oxopentanoyl]amino]-4-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-[[(2S)-1-amino-3-methyl-1-oxobutan-2-yl]amino]-3-(1H-indol-3-yl)-1-oxopropan-2-yl]amino]-1-oxo-3-phenylpropan-2-yl]amino]-4-methyl-1-oxopentan-2-yl]amino]-3-(4-hydroxyphenyl)-1-oxopropan-2-yl]amino]-4-oxobutanoic acid Chemical compound CC(C)C[C@H](NC(=O)[C@H](Cc1ccc(O)cc1)NC(=O)[C@H](CC(O)=O)NC(=O)[C@H](CCC(N)=O)NC(=O)[C@H](Cc1c[nH]c2ccccc12)NC(=O)[C@H](CO)NC(=O)[C@@H](NC(=O)[C@H](Cc1ccccc1)NC(=O)[C@@H](NC(=O)[C@@H](NC(C)=O)C(C)C)C(C)C)[C@@H](C)O)C(=O)N[C@@H](Cc1ccccc1)C(=O)N[C@@H](Cc1c[nH]c2ccccc12)C(=O)N[C@@H](C(C)C)C(N)=O CQTGBCFGAAYOCY-ZCRNMIQFSA-N 0.000 description 2
- 206010001742 Allergy to animal Diseases 0.000 description 2
- 241001225321 Aspergillus fumigatus Species 0.000 description 2
- 241000219495 Betulaceae Species 0.000 description 2
- 239000002028 Biomass Substances 0.000 description 2
- 241000282472 Canis lupus familiaris Species 0.000 description 2
- BVKZGUZCCUSVTD-UHFFFAOYSA-L Carbonate Chemical compound [O-]C([O-])=O BVKZGUZCCUSVTD-UHFFFAOYSA-L 0.000 description 2
- 240000009226 Corylus americana Species 0.000 description 2
- 235000001543 Corylus americana Nutrition 0.000 description 2
- 235000007466 Corylus avellana Nutrition 0.000 description 2
- 241000672609 Escherichia coli BL21 Species 0.000 description 2
- 241001198387 Escherichia coli BL21(DE3) Species 0.000 description 2
- 241001646716 Escherichia coli K-12 Species 0.000 description 2
- 229920001917 Ficoll Polymers 0.000 description 2
- WQZGKKKJIJFFOK-GASJEMHNSA-N Glucose Natural products OC[C@H]1OC(O)[C@H](O)[C@@H](O)[C@@H]1O WQZGKKKJIJFFOK-GASJEMHNSA-N 0.000 description 2
- NTYJJOPFIAHURM-UHFFFAOYSA-N Histamine Chemical compound NCCC1=CN=CN1 NTYJJOPFIAHURM-UHFFFAOYSA-N 0.000 description 2
- 108010002616 Interleukin-5 Proteins 0.000 description 2
- KFZMGEQAYNKOFK-UHFFFAOYSA-N Isopropanol Chemical compound CC(C)O KFZMGEQAYNKOFK-UHFFFAOYSA-N 0.000 description 2
- 239000006391 Luria-Bertani Medium Substances 0.000 description 2
- 244000141359 Malus pumila Species 0.000 description 2
- 235000011430 Malus pumila Nutrition 0.000 description 2
- 235000015103 Malus silvestris Nutrition 0.000 description 2
- 101100243377 Mus musculus Pepd gene Proteins 0.000 description 2
- 108091034117 Oligonucleotide Proteins 0.000 description 2
- 108010004729 Phycoerythrin Proteins 0.000 description 2
- 241000209504 Poaceae Species 0.000 description 2
- 230000006052 T cell proliferation Effects 0.000 description 2
- 239000002253 acid Substances 0.000 description 2
- 230000003092 anti-cytokine Effects 0.000 description 2
- 229940091771 aspergillus fumigatus Drugs 0.000 description 2
- 230000001580 bacterial effect Effects 0.000 description 2
- 230000008901 benefit Effects 0.000 description 2
- 229940041514 candida albicans extract Drugs 0.000 description 2
- 239000004202 carbamide Substances 0.000 description 2
- 229910052799 carbon Inorganic materials 0.000 description 2
- 230000001413 cellular effect Effects 0.000 description 2
- 230000008859 change Effects 0.000 description 2
- 238000012512 characterization method Methods 0.000 description 2
- 238000004587 chromatography analysis Methods 0.000 description 2
- 230000001684 chronic effect Effects 0.000 description 2
- 239000000562 conjugate Substances 0.000 description 2
- XUJNEKJLAYXESH-UHFFFAOYSA-N cysteine Natural products SCC(N)C(O)=O XUJNEKJLAYXESH-UHFFFAOYSA-N 0.000 description 2
- 125000000151 cysteine group Chemical group N[C@@H](CS)C(=O)* 0.000 description 2
- 230000016396 cytokine production Effects 0.000 description 2
- 238000000432 density-gradient centrifugation Methods 0.000 description 2
- 239000013024 dilution buffer Substances 0.000 description 2
- 229940079593 drug Drugs 0.000 description 2
- 230000008030 elimination Effects 0.000 description 2
- 238000003379 elimination reaction Methods 0.000 description 2
- 238000005516 engineering process Methods 0.000 description 2
- 230000002255 enzymatic effect Effects 0.000 description 2
- 238000002474 experimental method Methods 0.000 description 2
- 238000001914 filtration Methods 0.000 description 2
- 238000013467 fragmentation Methods 0.000 description 2
- 238000006062 fragmentation reaction Methods 0.000 description 2
- 239000008103 glucose Substances 0.000 description 2
- 208000006454 hepatitis Diseases 0.000 description 2
- 231100000283 hepatitis Toxicity 0.000 description 2
- 238000000265 homogenisation Methods 0.000 description 2
- 230000001900 immune effect Effects 0.000 description 2
- 230000006872 improvement Effects 0.000 description 2
- 210000003000 inclusion body Anatomy 0.000 description 2
- 229910052500 inorganic mineral Inorganic materials 0.000 description 2
- 238000003780 insertion Methods 0.000 description 2
- 230000037431 insertion Effects 0.000 description 2
- XMBWDFGMSWQBCA-YPZZEJLDSA-N iodane Chemical compound [125IH] XMBWDFGMSWQBCA-YPZZEJLDSA-N 0.000 description 2
- 229940044173 iodine-125 Drugs 0.000 description 2
- 210000004698 lymphocyte Anatomy 0.000 description 2
- 238000010907 mechanical stirring Methods 0.000 description 2
- 230000003278 mimic effect Effects 0.000 description 2
- 235000010755 mineral Nutrition 0.000 description 2
- 239000011707 mineral Substances 0.000 description 2
- 229940126619 mouse monoclonal antibody Drugs 0.000 description 2
- 230000035772 mutation Effects 0.000 description 2
- 239000013642 negative control Substances 0.000 description 2
- 230000003389 potentiating effect Effects 0.000 description 2
- 239000003755 preservative agent Substances 0.000 description 2
- 230000002335 preservative effect Effects 0.000 description 2
- 230000003449 preventive effect Effects 0.000 description 2
- 230000008569 process Effects 0.000 description 2
- 230000009696 proliferative response Effects 0.000 description 2
- 230000000069 prophylactic effect Effects 0.000 description 2
- 238000000746 purification Methods 0.000 description 2
- 230000001105 regulatory effect Effects 0.000 description 2
- 239000011347 resin Substances 0.000 description 2
- 229920005989 resin Polymers 0.000 description 2
- 108091008146 restriction endonucleases Proteins 0.000 description 2
- 150000003839 salts Chemical class 0.000 description 2
- 230000001932 seasonal effect Effects 0.000 description 2
- 230000028327 secretion Effects 0.000 description 2
- 235000004400 serine Nutrition 0.000 description 2
- 238000010181 skin prick test Methods 0.000 description 2
- 230000004936 stimulating effect Effects 0.000 description 2
- 239000000126 substance Substances 0.000 description 2
- 239000003053 toxin Substances 0.000 description 2
- 231100000765 toxin Toxicity 0.000 description 2
- 238000000870 ultraviolet spectroscopy Methods 0.000 description 2
- 230000003827 upregulation Effects 0.000 description 2
- 239000011534 wash buffer Substances 0.000 description 2
- 238000001262 western blot Methods 0.000 description 2
- 239000012138 yeast extract Substances 0.000 description 2
- YYGNTYWPHWGJRM-UHFFFAOYSA-N (6E,10E,14E,18E)-2,6,10,15,19,23-hexamethyltetracosa-2,6,10,14,18,22-hexaene Chemical compound CC(C)=CCCC(C)=CCCC(C)=CCCC=C(C)CCC=C(C)CCC=C(C)C YYGNTYWPHWGJRM-UHFFFAOYSA-N 0.000 description 1
- ZTOJFFHGPLIVKC-UHFFFAOYSA-N 3-ethyl-2-[(3-ethyl-6-sulfo-1,3-benzothiazol-2-ylidene)hydrazinylidene]-1,3-benzothiazole-6-sulfonic acid Chemical compound S1C2=CC(S(O)(=O)=O)=CC=C2N(CC)C1=NN=C1SC2=CC(S(O)(=O)=O)=CC=C2N1CC ZTOJFFHGPLIVKC-UHFFFAOYSA-N 0.000 description 1
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N 4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acid Chemical compound C([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1 FWMNVWWHGCHHJJ-SKKKGAJSSA-N 0.000 description 1
- 206010067484 Adverse reaction Diseases 0.000 description 1
- 229920001817 Agar Polymers 0.000 description 1
- 102000002260 Alkaline Phosphatase Human genes 0.000 description 1
- 108020004774 Alkaline Phosphatase Proteins 0.000 description 1
- 206010002198 Anaphylactic reaction Diseases 0.000 description 1
- 206010003645 Atopy Diseases 0.000 description 1
- 241000894006 Bacteria Species 0.000 description 1
- 206010066091 Bronchial Hyperreactivity Diseases 0.000 description 1
- 102100021943 C-C motif chemokine 2 Human genes 0.000 description 1
- 101710155857 C-C motif chemokine 2 Proteins 0.000 description 1
- 101710132601 Capsid protein Proteins 0.000 description 1
- 101710197658 Capsid protein VP1 Proteins 0.000 description 1
- OKTJSMMVPCPJKN-UHFFFAOYSA-N Carbon Chemical compound [C] OKTJSMMVPCPJKN-UHFFFAOYSA-N 0.000 description 1
- 108010012236 Chemokines Proteins 0.000 description 1
- 102000019034 Chemokines Human genes 0.000 description 1
- 108020004705 Codon Proteins 0.000 description 1
- 206010010741 Conjunctivitis Diseases 0.000 description 1
- 206010010744 Conjunctivitis allergic Diseases 0.000 description 1
- 102100031673 Corneodesmosin Human genes 0.000 description 1
- 101710139375 Corneodesmosin Proteins 0.000 description 1
- 208000020406 Creutzfeldt Jacob disease Diseases 0.000 description 1
- 208000003407 Creutzfeldt-Jakob Syndrome Diseases 0.000 description 1
- 208000010859 Creutzfeldt-Jakob disease Diseases 0.000 description 1
- 241001464975 Cutibacterium granulosum Species 0.000 description 1
- 208000001840 Dandruff Diseases 0.000 description 1
- 201000004624 Dermatitis Diseases 0.000 description 1
- 206010012438 Dermatitis atopic Diseases 0.000 description 1
- 241000238740 Dermatophagoides pteronyssinus Species 0.000 description 1
- 241000196324 Embryophyta Species 0.000 description 1
- 241000283074 Equus asinus Species 0.000 description 1
- 208000010201 Exanthema Diseases 0.000 description 1
- 108700024394 Exon Proteins 0.000 description 1
- 238000012413 Fluorescence activated cell sorting analysis Methods 0.000 description 1
- 241000233866 Fungi Species 0.000 description 1
- CEAZRRDELHUEMR-URQXQFDESA-N Gentamicin Chemical compound O1[C@H](C(C)NC)CC[C@@H](N)[C@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](NC)[C@@](C)(O)CO2)O)[C@H](N)C[C@@H]1N CEAZRRDELHUEMR-URQXQFDESA-N 0.000 description 1
- 229930182566 Gentamicin Natural products 0.000 description 1
- 244000068988 Glycine max Species 0.000 description 1
- 235000010469 Glycine max Nutrition 0.000 description 1
- 108090000288 Glycoproteins Proteins 0.000 description 1
- 102000003886 Glycoproteins Human genes 0.000 description 1
- 108010017080 Granulocyte Colony-Stimulating Factor Proteins 0.000 description 1
- 102000004269 Granulocyte Colony-Stimulating Factor Human genes 0.000 description 1
- 102000004457 Granulocyte-Macrophage Colony-Stimulating Factor Human genes 0.000 description 1
- 108010017213 Granulocyte-Macrophage Colony-Stimulating Factor Proteins 0.000 description 1
- 108010042653 IgA receptor Proteins 0.000 description 1
- 208000001718 Immediate Hypersensitivity Diseases 0.000 description 1
- 206010061218 Inflammation Diseases 0.000 description 1
- 102000008070 Interferon-gamma Human genes 0.000 description 1
- 108090000978 Interleukin-4 Proteins 0.000 description 1
- 108090001005 Interleukin-6 Proteins 0.000 description 1
- 108010002586 Interleukin-7 Proteins 0.000 description 1
- 108090001007 Interleukin-8 Proteins 0.000 description 1
- XUJNEKJLAYXESH-REOHCLBHSA-N L-Cysteine Chemical compound SC[C@H](N)C(O)=O XUJNEKJLAYXESH-REOHCLBHSA-N 0.000 description 1
- QNAYBMKLOCPYGJ-REOHCLBHSA-N L-alanine Chemical compound C[C@H](N)C(O)=O QNAYBMKLOCPYGJ-REOHCLBHSA-N 0.000 description 1
- ZDXPYRJPNDTMRX-VKHMYHEASA-N L-glutamine Chemical compound OC(=O)[C@@H](N)CCC(N)=O ZDXPYRJPNDTMRX-VKHMYHEASA-N 0.000 description 1
- 229930182816 L-glutamine Natural products 0.000 description 1
- 239000006142 Luria-Bertani Agar Substances 0.000 description 1
- 108010052285 Membrane Proteins Proteins 0.000 description 1
- 102000018697 Membrane Proteins Human genes 0.000 description 1
- 241001467578 Microbacterium Species 0.000 description 1
- 229910019142 PO4 Inorganic materials 0.000 description 1
- 101800005149 Peptide B Proteins 0.000 description 1
- 101800004191 Peptide P2 Proteins 0.000 description 1
- 239000001888 Peptone Substances 0.000 description 1
- 108010080698 Peptones Proteins 0.000 description 1
- 208000024777 Prion disease Diseases 0.000 description 1
- 102100034014 Prolyl 3-hydroxylase 3 Human genes 0.000 description 1
- 108010076504 Protein Sorting Signals Proteins 0.000 description 1
- 239000012614 Q-Sepharose Substances 0.000 description 1
- 101710118046 RNA-directed RNA polymerase Proteins 0.000 description 1
- 108020004511 Recombinant DNA Proteins 0.000 description 1
- 240000004808 Saccharomyces cerevisiae Species 0.000 description 1
- 229920002684 Sepharose Polymers 0.000 description 1
- MTCFGRXMJLQNBG-UHFFFAOYSA-N Serine Natural products OCC(N)C(O)=O MTCFGRXMJLQNBG-UHFFFAOYSA-N 0.000 description 1
- 102000012479 Serine Proteases Human genes 0.000 description 1
- 108010022999 Serine Proteases Proteins 0.000 description 1
- 229910000831 Steel Inorganic materials 0.000 description 1
- 108010090804 Streptavidin Proteins 0.000 description 1
- 230000024932 T cell mediated immunity Effects 0.000 description 1
- BHEOSNUKNHRBNM-UHFFFAOYSA-N Tetramethylsqualene Natural products CC(=C)C(C)CCC(=C)C(C)CCC(C)=CCCC=C(C)CCC(C)C(=C)CCC(C)C(C)=C BHEOSNUKNHRBNM-UHFFFAOYSA-N 0.000 description 1
- 239000013504 Triton X-100 Substances 0.000 description 1
- 229920004890 Triton X-100 Polymers 0.000 description 1
- 108060008682 Tumor Necrosis Factor Proteins 0.000 description 1
- 102100040247 Tumor necrosis factor Human genes 0.000 description 1
- 108010067390 Viral Proteins Proteins 0.000 description 1
- 208000036142 Viral infection Diseases 0.000 description 1
- 101710108545 Viral protein 1 Proteins 0.000 description 1
- JLCPHMBAVCMARE-UHFFFAOYSA-N [3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[3-[[3-[[3-[[3-[[3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-[[5-(2-amino-6-oxo-1H-purin-9-yl)-3-hydroxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxyoxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(5-methyl-2,4-dioxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(6-aminopurin-9-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl]oxy-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methyl [5-(6-aminopurin-9-yl)-2-(hydroxymethyl)oxolan-3-yl] hydrogen phosphate Polymers Cc1cn(C2CC(OP(O)(=O)OCC3OC(CC3OP(O)(=O)OCC3OC(CC3O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c3nc(N)[nH]c4=O)C(COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3COP(O)(=O)OC3CC(OC3CO)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3ccc(N)nc3=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cc(C)c(=O)[nH]c3=O)n3cc(C)c(=O)[nH]c3=O)n3ccc(N)nc3=O)n3cc(C)c(=O)[nH]c3=O)n3cnc4c3nc(N)[nH]c4=O)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)n3cnc4c(N)ncnc34)O2)c(=O)[nH]c1=O JLCPHMBAVCMARE-UHFFFAOYSA-N 0.000 description 1
- 150000007513 acids Chemical class 0.000 description 1
- 230000001154 acute effect Effects 0.000 description 1
- 230000006838 adverse reaction Effects 0.000 description 1
- 239000008272 agar Substances 0.000 description 1
- 238000005054 agglomeration Methods 0.000 description 1
- 230000002776 aggregation Effects 0.000 description 1
- 238000013019 agitation Methods 0.000 description 1
- 239000013572 airborne allergen Substances 0.000 description 1
- 235000004279 alanine Nutrition 0.000 description 1
- 230000009285 allergic inflammation Effects 0.000 description 1
- AZDRQVAHHNSJOQ-UHFFFAOYSA-N alumane Chemical class [AlH3] AZDRQVAHHNSJOQ-UHFFFAOYSA-N 0.000 description 1
- 150000001412 amines Chemical class 0.000 description 1
- 230000036783 anaphylactic response Effects 0.000 description 1
- 208000003455 anaphylaxis Diseases 0.000 description 1
- 238000010171 animal model Methods 0.000 description 1
- 235000021120 animal protein Nutrition 0.000 description 1
- 230000003266 anti-allergic effect Effects 0.000 description 1
- 239000012131 assay buffer Substances 0.000 description 1
- QVGXLLKOCUKJST-UHFFFAOYSA-N atomic oxygen Chemical compound [O] QVGXLLKOCUKJST-UHFFFAOYSA-N 0.000 description 1
- 201000008937 atopic dermatitis Diseases 0.000 description 1
- 239000012752 auxiliary agent Substances 0.000 description 1
- OHDRQQURAXLVGJ-HLVWOLMTSA-N azane;(2e)-3-ethyl-2-[(e)-(3-ethyl-6-sulfo-1,3-benzothiazol-2-ylidene)hydrazinylidene]-1,3-benzothiazole-6-sulfonic acid Chemical compound [NH4+].[NH4+].S/1C2=CC(S([O-])(=O)=O)=CC=C2N(CC)C\1=N/N=C1/SC2=CC(S([O-])(=O)=O)=CC=C2N1CC OHDRQQURAXLVGJ-HLVWOLMTSA-N 0.000 description 1
- 239000011324 bead Substances 0.000 description 1
- 230000009286 beneficial effect Effects 0.000 description 1
- 230000008827 biological function Effects 0.000 description 1
- 239000000090 biomarker Substances 0.000 description 1
- 230000001851 biosynthetic effect Effects 0.000 description 1
- 230000036427 bronchial hyperreactivity Effects 0.000 description 1
- 125000000484 butyl group Chemical group [H]C([*])([H])C([H])([H])C([H])([H])C([H])([H])[H] 0.000 description 1
- 239000001506 calcium phosphate Substances 0.000 description 1
- 229910000389 calcium phosphate Inorganic materials 0.000 description 1
- 235000011010 calcium phosphates Nutrition 0.000 description 1
- 239000000969 carrier Substances 0.000 description 1
- 210000002421 cell wall Anatomy 0.000 description 1
- 238000012412 chemical coupling Methods 0.000 description 1
- 238000011210 chromatographic step Methods 0.000 description 1
- 210000000349 chromosome Anatomy 0.000 description 1
- 238000005352 clarification Methods 0.000 description 1
- 238000003776 cleavage reaction Methods 0.000 description 1
- 238000004590 computer program Methods 0.000 description 1
- 230000021615 conjugation Effects 0.000 description 1
- 238000007796 conventional method Methods 0.000 description 1
- 238000012258 culturing Methods 0.000 description 1
- 230000009089 cytolysis Effects 0.000 description 1
- 210000000805 cytoplasm Anatomy 0.000 description 1
- 230000003247 decreasing effect Effects 0.000 description 1
- 238000012217 deletion Methods 0.000 description 1
- 230000037430 deletion Effects 0.000 description 1
- 230000001419 dependent effect Effects 0.000 description 1
- 230000000779 depleting effect Effects 0.000 description 1
- 238000013461 design Methods 0.000 description 1
- 238000011026 diafiltration Methods 0.000 description 1
- 230000029087 digestion Effects 0.000 description 1
- AIUDWMLXCFRVDR-UHFFFAOYSA-N dimethyl 2-(3-ethyl-3-methylpentyl)propanedioate Chemical compound CCC(C)(CC)CCC(C(=O)OC)C(=O)OC AIUDWMLXCFRVDR-UHFFFAOYSA-N 0.000 description 1
- 230000006806 disease prevention Effects 0.000 description 1
- PRAKJMSDJKAYCZ-UHFFFAOYSA-N dodecahydrosqualene Natural products CC(C)CCCC(C)CCCC(C)CCCCC(C)CCCC(C)CCCC(C)C PRAKJMSDJKAYCZ-UHFFFAOYSA-N 0.000 description 1
- 239000002552 dosage form Substances 0.000 description 1
- 231100000673 dose–response relationship Toxicity 0.000 description 1
- 239000012636 effector Substances 0.000 description 1
- 239000000839 emulsion Substances 0.000 description 1
- 239000003623 enhancer Substances 0.000 description 1
- 230000007613 environmental effect Effects 0.000 description 1
- 210000000981 epithelium Anatomy 0.000 description 1
- 210000003527 eukaryotic cell Anatomy 0.000 description 1
- 201000005884 exanthem Diseases 0.000 description 1
- 230000002349 favourable effect Effects 0.000 description 1
- 210000003754 fetus Anatomy 0.000 description 1
- 239000012467 final product Substances 0.000 description 1
- 239000013568 food allergen Substances 0.000 description 1
- 230000008014 freezing Effects 0.000 description 1
- 238000007710 freezing Methods 0.000 description 1
- 230000002538 fungal effect Effects 0.000 description 1
- 230000002068 genetic effect Effects 0.000 description 1
- 229960002518 gentamicin Drugs 0.000 description 1
- 108091005608 glycosylated proteins Proteins 0.000 description 1
- 102000035122 glycosylated proteins Human genes 0.000 description 1
- 108060003552 hemocyanin Proteins 0.000 description 1
- 229960001340 histamine Drugs 0.000 description 1
- 239000011539 homogenization buffer Substances 0.000 description 1
- 230000028996 humoral immune response Effects 0.000 description 1
- 230000002519 immonomodulatory effect Effects 0.000 description 1
- 210000004201 immune sera Anatomy 0.000 description 1
- 229940042743 immune sera Drugs 0.000 description 1
- 210000000987 immune system Anatomy 0.000 description 1
- 230000036039 immunity Effects 0.000 description 1
- 230000007233 immunological mechanism Effects 0.000 description 1
- 230000003308 immunostimulating effect Effects 0.000 description 1
- 230000001976 improved effect Effects 0.000 description 1
- 238000000338 in vitro Methods 0.000 description 1
- 238000011065 in-situ storage Methods 0.000 description 1
- 230000002757 inflammatory effect Effects 0.000 description 1
- 230000004054 inflammatory process Effects 0.000 description 1
- 230000000977 initiatory effect Effects 0.000 description 1
- 238000011081 inoculation Methods 0.000 description 1
- 229960003130 interferon gamma Drugs 0.000 description 1
- BPHPUYQFMNQIOC-NXRLNHOXSA-N isopropyl beta-D-thiogalactopyranoside Chemical compound CC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1O BPHPUYQFMNQIOC-NXRLNHOXSA-N 0.000 description 1
- 231100000518 lethal Toxicity 0.000 description 1
- 230000001665 lethal effect Effects 0.000 description 1
- 150000002617 leukotrienes Chemical class 0.000 description 1
- 150000002632 lipids Chemical class 0.000 description 1
- 238000005567 liquid scintillation counting Methods 0.000 description 1
- 210000004072 lung Anatomy 0.000 description 1
- 210000004962 mammalian cell Anatomy 0.000 description 1
- 230000007246 mechanism Effects 0.000 description 1
- 230000005541 medical transmission Effects 0.000 description 1
- 238000002483 medication Methods 0.000 description 1
- 239000012528 membrane Substances 0.000 description 1
- 125000001360 methionine group Chemical group N[C@@H](CCSC)C(=O)* 0.000 description 1
- 230000004048 modification Effects 0.000 description 1
- 238000012986 modification Methods 0.000 description 1
- 238000010369 molecular cloning Methods 0.000 description 1
- 229940035032 monophosphoryl lipid a Drugs 0.000 description 1
- 125000001446 muramyl group Chemical group N[C@@H](C=O)[C@@H](O[C@@H](C(=O)*)C)[C@H](O)[C@H](O)CO 0.000 description 1
- 102000006392 myotrophin Human genes 0.000 description 1
- 108010058605 myotrophin Proteins 0.000 description 1
- 238000011587 new zealand white rabbit Methods 0.000 description 1
- 229910052760 oxygen Inorganic materials 0.000 description 1
- 239000001301 oxygen Substances 0.000 description 1
- 230000001575 pathological effect Effects 0.000 description 1
- 230000007170 pathology Effects 0.000 description 1
- 108010091748 peptide A Proteins 0.000 description 1
- 239000000863 peptide conjugate Substances 0.000 description 1
- 229940023041 peptide vaccine Drugs 0.000 description 1
- 235000019319 peptone Nutrition 0.000 description 1
- 125000001997 phenyl group Chemical group [H]C1=C([H])C([H])=C(*)C([H])=C1[H] 0.000 description 1
- NBIIXXVUZAFLBC-UHFFFAOYSA-K phosphate Chemical compound [O-]P([O-])([O-])=O NBIIXXVUZAFLBC-UHFFFAOYSA-K 0.000 description 1
- 239000010452 phosphate Substances 0.000 description 1
- 229940068196 placebo Drugs 0.000 description 1
- 239000000902 placebo Substances 0.000 description 1
- 239000013573 pollen allergen Substances 0.000 description 1
- 229920002401 polyacrylamide Polymers 0.000 description 1
- 108091033319 polynucleotide Proteins 0.000 description 1
- 102000040430 polynucleotide Human genes 0.000 description 1
- 239000002157 polynucleotide Substances 0.000 description 1
- 210000001236 prokaryotic cell Anatomy 0.000 description 1
- 230000001012 protector Effects 0.000 description 1
- 238000000159 protein binding assay Methods 0.000 description 1
- 239000012460 protein solution Substances 0.000 description 1
- 230000012743 protein tagging Effects 0.000 description 1
- 239000001397 quillaja saponaria molina bark Substances 0.000 description 1
- 206010037844 rash Diseases 0.000 description 1
- 238000010188 recombinant method Methods 0.000 description 1
- 210000003289 regulatory T cell Anatomy 0.000 description 1
- 238000011160 research Methods 0.000 description 1
- 210000003296 saliva Anatomy 0.000 description 1
- 229930182490 saponin Natural products 0.000 description 1
- 150000007949 saponins Chemical class 0.000 description 1
- 230000007017 scission Effects 0.000 description 1
- 230000035945 sensitivity Effects 0.000 description 1
- 150000003355 serines Chemical class 0.000 description 1
- 230000035939 shock Effects 0.000 description 1
- 239000000243 solution Substances 0.000 description 1
- 238000000527 sonication Methods 0.000 description 1
- 229940043517 specific immunoglobulins Drugs 0.000 description 1
- 229940031439 squalene Drugs 0.000 description 1
- TUHBEKDERLKLEC-UHFFFAOYSA-N squalene Natural products CC(=CCCC(=CCCC(=CCCC=C(/C)CCC=C(/C)CC=C(C)C)C)C)C TUHBEKDERLKLEC-UHFFFAOYSA-N 0.000 description 1
- 239000012192 staining solution Substances 0.000 description 1
- 229910001220 stainless steel Inorganic materials 0.000 description 1
- 239000010935 stainless steel Substances 0.000 description 1
- 238000007619 statistical method Methods 0.000 description 1
- 239000010959 steel Substances 0.000 description 1
- 238000003756 stirring Methods 0.000 description 1
- 238000006467 substitution reaction Methods 0.000 description 1
- 238000003786 synthesis reaction Methods 0.000 description 1
- 230000009885 systemic effect Effects 0.000 description 1
- 238000010257 thawing Methods 0.000 description 1
- 230000004797 therapeutic response Effects 0.000 description 1
- 238000004448 titration Methods 0.000 description 1
- 230000003614 tolerogenic effect Effects 0.000 description 1
- 231100000331 toxic Toxicity 0.000 description 1
- 230000002588 toxic effect Effects 0.000 description 1
- 239000011573 trace mineral Substances 0.000 description 1
- 235000013619 trace mineral Nutrition 0.000 description 1
- 238000013518 transcription Methods 0.000 description 1
- 230000035897 transcription Effects 0.000 description 1
- 230000009466 transformation Effects 0.000 description 1
- 238000013519 translation Methods 0.000 description 1
- 230000032258 transport Effects 0.000 description 1
- QORWJWZARLRLPR-UHFFFAOYSA-H tricalcium bis(phosphate) Chemical compound [Ca+2].[Ca+2].[Ca+2].[O-]P([O-])([O-])=O.[O-]P([O-])([O-])=O QORWJWZARLRLPR-UHFFFAOYSA-H 0.000 description 1
- 238000000108 ultra-filtration Methods 0.000 description 1
- 241001515965 unidentified phage Species 0.000 description 1
- 210000002700 urine Anatomy 0.000 description 1
- 235000013311 vegetables Nutrition 0.000 description 1
- 210000003462 vein Anatomy 0.000 description 1
- 239000002435 venom Substances 0.000 description 1
- 231100000611 venom Toxicity 0.000 description 1
- 210000001048 venom Anatomy 0.000 description 1
- 230000009385 viral infection Effects 0.000 description 1
- XLYOFNOQVPJJNP-UHFFFAOYSA-N water Substances O XLYOFNOQVPJJNP-UHFFFAOYSA-N 0.000 description 1
- DGVVWUTYPXICAM-UHFFFAOYSA-N β‐Mercaptoethanol Chemical compound OCCS DGVVWUTYPXICAM-UHFFFAOYSA-N 0.000 description 1
Classifications
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
- A61K39/29—Hepatitis virus
- A61K39/292—Serum hepatitis virus, hepatitis B virus, e.g. Australia antigen
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/12—Viral antigens
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/35—Allergens
- A61K39/36—Allergens from pollen
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K39/385—Haptens or antigens, bound to carriers
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61P—SPECIFIC THERAPEUTIC ACTIVITY OF CHEMICAL COMPOUNDS OR MEDICINAL PREPARATIONS
- A61P37/00—Drugs for immunological or allergic disorders
- A61P37/08—Antiallergic agents
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/005—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from viruses
- C07K14/01—DNA viruses
- C07K14/02—Hepadnaviridae, e.g. hepatitis B virus
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/415—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from plants
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/51—Medicinal preparations containing antigens or antibodies comprising whole cells, viruses or DNA/RNA
- A61K2039/53—DNA (RNA) vaccination
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/57—Medicinal preparations containing antigens or antibodies characterised by the type of response, e.g. Th1, Th2
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/60—Medicinal preparations containing antigens or antibodies characteristics by the carrier linked to the antigen
- A61K2039/6031—Proteins
- A61K2039/6075—Viral proteins
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/64—Medicinal preparations containing antigens or antibodies characterised by the architecture of the carrier-antigen complex, e.g. repetition of carrier-antigen units
-
- A—HUMAN NECESSITIES
- A61—MEDICAL OR VETERINARY SCIENCE; HYGIENE
- A61K—PREPARATIONS FOR MEDICAL, DENTAL OR TOILETRY PURPOSES
- A61K39/00—Medicinal preparations containing antigens or antibodies
- A61K2039/64—Medicinal preparations containing antigens or antibodies characterised by the architecture of the carrier-antigen complex, e.g. repetition of carrier-antigen units
- A61K2039/645—Dendrimers; Multiple antigen peptides
-
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K2319/00—Fusion polypeptide
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2730/00—Reverse transcribing DNA viruses
- C12N2730/00011—Details
- C12N2730/10011—Hepadnaviridae
- C12N2730/10111—Orthohepadnavirus, e.g. hepatitis B virus
- C12N2730/10122—New viral proteins or individual genes, new structural or functional aspects of known viral proteins or genes
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2730/00—Reverse transcribing DNA viruses
- C12N2730/00011—Details
- C12N2730/10011—Hepadnaviridae
- C12N2730/10111—Orthohepadnavirus, e.g. hepatitis B virus
- C12N2730/10134—Use of virus or viral component as vaccine, e.g. live-attenuated or inactivated virus, VLP, viral protein
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Chemical & Material Sciences (AREA)
- Medicinal Chemistry (AREA)
- General Health & Medical Sciences (AREA)
- Virology (AREA)
- Pharmacology & Pharmacy (AREA)
- Immunology (AREA)
- Animal Behavior & Ethology (AREA)
- Public Health (AREA)
- Veterinary Medicine (AREA)
- Organic Chemistry (AREA)
- Microbiology (AREA)
- Mycology (AREA)
- Epidemiology (AREA)
- Gastroenterology & Hepatology (AREA)
- Biochemistry (AREA)
- Biophysics (AREA)
- Genetics & Genomics (AREA)
- Molecular Biology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Communicable Diseases (AREA)
- Botany (AREA)
- Engineering & Computer Science (AREA)
- Pulmonology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Nuclear Medicine, Radiotherapy & Molecular Imaging (AREA)
- Peptides Or Proteins (AREA)
- Medicines Containing Antibodies Or Antigens For Use As Internal Diagnostic Agents (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Polipéptido capaz de generar respuestas de IgG contra al menos un alérgeno de tipo silvestre y capaz de inducir la producción de IL-10 e IFN-gamma, en donde la IgG generada de este modo se centra en los epítopos de IgE del alérgeno de tipo silvestre y puede bloquear la unión a dicho alérgeno de tipo silvestre, comprendiendo dicho polipéptido al menos 4 y hasta 6 fragmentos peptídicos hipoalergénicos que consisten en de 20 a 50 restos de aminoácido consecutivos de al menos un alérgeno de tipo silvestre, fusionados a los extremos N y C-terminales de una proteína PreS de hepatitis B (SEQ ID NO: 21) que actúa como transportador, en donde dichos fragmentos de péptido muestran una capacidad de unión a IgE nula o reducida en comparación con el alérgeno de tipo silvestre, son péptidos de unión a linfocitos B y muestran una reactividad con linfocitos T nula o sustancialmente nula, en donde la proteína PreS de hepatitis B comprende al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo N-terminal y al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo C-terminal.
Description
DESCRIPCIÓN
Proteínas de fusión transportadoras de péptido como vacunas para alergias
La presente invención se refiere a la fusión de fragmentos de proteína de alérgeno a una proteína vírica que son útiles para el tratamiento de alergias.
La alergia de tipo I es una enfermedad por hipersensibilidad mediada por IgE que afecta a prácticamente un 25 % de la población. Se basa en el reconocimiento de antígenos inofensivos transportados por el aire, de insectos, de veneno, alérgenos alimentarios y alérgenos de contacto procedentes de fuentes de antígeno de por sí inofensivas, tales como polen, insectos, moho y proteínas de animales mediante inmunoglobulinas E específicas. El sobrecruzamiento de los anticuerpos IgE unidos a células efectoras provoca una liberación de mediadores inflamatorios (por ejemplo, histamina, leucotrienos) y por lo tanto, a los síntomas inmediatos de la alergia (por ejemplo, rinoconjuntivitis, asma, dermatitis, anafilaxia). La activación de linfocitos T mediante mecanismos dependientes de IgE así como independientes de IgE contribuye a la inflamación alérgica.
Probablemente, la única forma de tratamiento de las causas de la alergia es la inmunoterapia con antígenos específicos, que se basa en la administración repetida de cantidades crecientes de extractos de alérgeno de la mayoría de las fuentes. Numerosos estudios clínicos han documentado la eficacia clínica de la inyección de inmunoterapia y hay pruebas de varios mecanismos inmunológicos que respaldan este tratamiento. Debido a la dificultad para preparar extractos de alérgeno de alta calidad para ciertas fuentes de alérgeno y al hecho de que la administración de alérgenos a los pacientes puede provocar graves efectos secundarios, solo puede recomendarse la inmunoterapia con alérgenos específicos para ciertos grupos de pacientes y manifestaciones de la enfermedad. Es especialmente complicado tratar a pacientes sensibles a varias fuentes de alérgenos diferentes y a pacientes que padecen manifestaciones graves de la enfermedad, tales como asma alérgica. El asma alergia es una de las manifestaciones más vigorosas de la alergia, debido a que afecta gravemente a la calidad de vida diaria, provoca una elevada tasa de ingresos hospitalarios y puede manifestarse en formas graves, potencialmente letales que requieren cuidados intensivos del paciente.
Los extractos de alérgenos preparados de fuentes de alérgenos naturales se encuentran en bruto y es imposible influenciar la calidad y las cantidades de los alérgenos individuales en dichas preparaciones mediante medios técnicos. También contienen numerosos componentes no alergénicos no definidos y varios estudios recientes han destacado la baja calidad de dichos extractos y documentan su gran heterogeneidad.
En la última década, se han hecho grandes progresos en el campo de la caracterización molecular de alérgenos usando tecnología de ADN recombinante. Un gran número de los alérgenos causantes de enfermedades más importantes se han caracterizado hasta el nivel molecular y se han producido alérgenos recombinantes que imitan la complejidad epitópica de los extractos de alérgeno naturales. Por otra parte, varios grupos de investigación han usado el conocimiento relativo a las estructuras de alérgeno para desarrollar nuevas vacunas contra la alergia definidas. Se ha empleado ingeniería genética, química de péptidos sintéticos y conjugación de alérgenos con secuencias de ADN inmunoestimuladoras para reducir la actividad alergénica de las nuevas vacunas y por lo tanto, la frecuencia de efectos secundarios inducidos por la terapia. Los primeros estudios clínicos prometedores se llevaron a cabo con dichos derivados de alérgenos. De forma interesante, se dio el caso de que aunque la reactividad con IgE de los alérgenos recombinantes modificados por ingeniería genética y los péptidos que contienen epítopos de linfocitos T sintéticos procedentes de alérgenos pueden reducirse significativamente o incluso suprimirse, estos derivados siguen pudiendo inducir efectos secundarios sistémicos que aparecen varias horas después de la inyección. Por ejemplo, se ha comunicado que los péptidos de epítopo de linfocitos T del alérgeno principal de gato, Fel d 1, indujeron asma e hiperreactividad bronquial varias horas después de su inyección intracutánea y hay sólidos indicios de que este efecto está mediado por linfocitos T y restringido al MHC.
Estos resultados indican que la eliminación de la reactividad de IgE reduce los efectos secundarios mediados por IgE ya que no se registraron reacciones inmediatas durante el transcurso de estos estudios de inmunoterapia. Sin embargo, los epítopos de linfocitos T específicos de alérgeno que se han preservado en los derivados de alérgeno recombinantes, así como en las mezclas de péptidos, son responsables de los efectos secundarios tardíos (por ejemplo, dermatitis muy problemática o atópica, manifestación alérgica en la piel crónica mediada por linfocitos T). Los efectos secundarios provocados en el caso de los derivados de alérgeno recombinantes fueron relativamente leves y en el caso de las vacunas de péptido de linfocitos T, se pueden superar mediante una dosificación adecuada. Por lo tanto, estas dos nuevas estrategias parecen muy prometedoras para la inmunoterapia de la rinoconjuntivitis alérgica, pero pueden tener limitaciones cuando se aborda el tratamiento de las formas graves del asma alérgica, donde puede ser problemática la inducción de efectos secundarios tardíos en el pulmón.
Para administrar y por consiguiente provocar una respuesta inmunitaria eficaz contra péptidos, polipéptidos y proteínas, pueden usarse regularmente adyuvantes y/o transportadores. por ejemplo, el adyuvante completo de Freund (CFA) es uno de los adyuvantes más potentes disponibles. Existe la necesidad de composiciones de vacuna capaces de inducir fuertes respuestas inmunitarias contra péptidos y polipéptidos procedentes de alérgenos y obviamente, de otros antígenos con o sin el uso de adyuvante completo de Freund. Además, aunque se ha usado con éxito BSA como portador en modelos animales, puede no ser adecuado para su uso en composiciones de vacuna
para seres humanos debido al riesgo de reacciones adversas, tal como el riesgo de transmisión de enfermedad por priones (variante de la enfermedad de Creutzfeldt-Jakob). Otro reto para el desarrollo de una vacuna eficaz contra alérgenos es la necesidad de una respuesta inmunitaria capaz de reducir los alérgenos rápidamente en un individuo 0 animal. Por lo tanto, se necesitan elevadas concentraciones de anticuerpos específicos para alérgenos, que sean principalmente del subtipo IgG. En las superficies mucosas, también son importantes los anticuerpos IgA.
La toxina colérica, una proteína transportadora conocida en la técnica, también se usa regularmente como adyuvante. Sin embargo, la toxina colérica aumenta los niveles totales y específicos de anticuerpo IgE y provoca reacciones inflamatorias asociadas con IgE.
Debido a los efectos secundarios provocados por la mayoría de proteínas transportadoras usadas para vacunación, existe la necesidad de sistemas transportadores que sean capaces de estimular las respuestas inmunitarias contra alérgenos u otros antígenos, sin usar adyuvantes tóxicos, sin usar proteínas transportadoras poco toleradas y, en ciertas situaciones, sin estimular respuestas inmunitarias potencialmente patológicas. Pueden usarse nuevos sistemas transportadores que cumplan estas especificaciones para la formación de nuevos conjugados y composiciones adecuadas para el tratamiento o la prevención de enfermedades, tales como enfermedades alérgicas.
En Bohle B. et al. (J. Immunol. 172 (11) (2004): 6642-6648) se describe una proteína de fusión recombinante que comprende un resto de proteína de capa S y un resto de Bet v 1. Esta molécula comprende el alérgeno natural Bet v 1 que incluye epítopos para linfocitos T específicos de Bet v 1.
El documento WO 2004/004761 se refiere a partículas víricas que se fusionan a un inmunógeno y que pueden usarse para inmunización.
En el documento WO 2004/003143 se divulga el uso de proteínas de fusión que comprenden una partícula de tipo vírico y una molécula alergénica como inmunógeno para vacunación.
En el documento WO 2007/140505 y en Edlmayr et al. (J. Immunol. 182 (10) (2009) 6298-6306) se describe el uso de proteínas de fusión que comprenden diversas moléculas de transportador fusionadas a péptidos procedentes de alérgeno para inducir anticuerpos IgG específicos para alérgenos, pero estas construcciones no muestran un efecto inmunomodulador que pueda considerarse ventajoso para pacientes alérgicos, tales como la inducción de IL-10 o de inmunidad de Th1. La Fig. 4 de Edlmayr et al muestra que los péptidos fusionados a KLH inducen la citocina de Th2, IL-5 y las proteínas de fusión de VP1 no inducen IL-10 o IFN-gamma.
En Niespodziana et al (J. Allergy Clin. Immunol. 127 (6) (2011) 1562-1570) se describe el uso de proteínas de fusión, comprendiendo cada una PreS procedente de hepatitis B y dos péptidos procedentes del alérgeno principal de gato, Fel d 1, para inducir anticuerpos IgG específicos de alérgeno. Sin embargo, no se ha descrito un régimen adecuado para la vacunación de seres humanos y los péptidos contenían epítopos de linfocitos T específicos de alérgenos. Un objeto de la presente invención es proporcionar medicamentos y portadores que superen los inconvenientes anteriormente mencionados y permitan una vacunación con alérgeno con efectos secundarios reducidos.
Por lo tanto, la presente invención se refiere a un polipéptido capaz de generar respuestas de IgG contra al menos un alérgeno de tipo silvestre, en donde la IgG generada de este modo puede bloquear la unión de IgG a dicho alérgeno de tipo silvestre y capaz de inducir la producción de IL-10 e IFN-gamma; comprendiendo dicho polipéptido al menos 4 y hasta 6 fragmentos peptídicos hipoalergénicos que consisten en de 20 a 50 restos de aminoácido consecutivos de al menos un alérgeno de tipo silvestre, fusionado al extremo N y C-terminal de una proteína transportadora, la proteína PreS de hepatitis B (SEQ ID NO: 21), en donde dicho fragmento es un polipéptido de superficie de un virus de la familia hepadnaviridae o al menos un fragmento de dicho polipéptido de superficie, en donde los al menos cuatro fragmentos de péptido muestran una capacidad de unión a IgE nula o reducida en comparación con el alérgeno de tipo silvestre, son péptidos de unión a linfocitos B y muestran una reactividad con linfocitos B nula o sustancialmente nula, en donde la proteína PreS de hepatitis B comprende al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo N-terminal y al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo C-terminal.
A fin de provocar una respuesta inmunitaria potenciada contra una molécula, en particular contra una molécula alergénica o hipoalergénica de acuerdo con la presente invención, se fusionan (por ingeniería genética) al menos cuatro fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre a un polipéptido PreS de hepatitis B. Se ha observado, sorprendentemente, que a diferencia de las proteínas transportadoras empleadas de manera convencional y regular, como KLH (hemocianina de lapa californiana), un polipéptido de la superficie de un virus de la familia hepadnaviridae, un polipéptido PreS de la hepatitis B, produce una formación potenciada de anticuerpos dirigidos a aquellos péptidos que están unidos a los mismos.
Se ha observado que los anticuerpos IgG específicos para alérgenos inducidos mediante la inmunización con más de tres fragmentos de péptido procedentes de alérgeno adecuadamente seleccionados, fusionados al polipéptido PreS de la hepatitis B, están mejor enfocados hacia los epítopos de IgE del alérgeno, mientras que la inmunización con el
alérgeno de tipo silvestre produce IgG que están dirigidas a todas las partes del alérgeno, incluyendo aquellas que no son reactivas con IgE. En un experimento normalizado para títulos de IgG, esto produce una mejor capacidad de bloqueo de la IgG inducida por PreS/péptido en comparación con la inducida por el alérgeno de tipo silvestre (Fig. 12). También sorprendentemente, se observó que en cultivos de PBMC humanas, las proteínas de fusión de fragmentos de péptido procedentes de alérgeno con el polipéptido PreS de la hepatitis B indujeron fuertemente las citocinas IL-10 e FN-gamma, que se han atribuido como indicadores positivos para una inmunoterapia exitosa contra alergias. Por el contrario, la inducción de IL-10 e IFN-gamma fue significativamente menor con alérgeno de tipo silvestre, fragmentos de péptido procedentes de alérgeno solos o solo PreS (Fig. 10).
"Fusionado al extremo N y C-terminal", como se usa en el presente documento, significa que están fusionados al menos dos péptidos al extremo N-terminal de una proteína PreS de hepatitis B y al menos dos péptidos están fusionados al extremo C-terminal de una proteína PreS de hepatitis B. Para ilustración, en la construcción más básica, una proteína PreS de hepatitis B puede comprender dos péptidos en el extremo N-terminal y dos péptidos en el extremo C-terminal.
El polipéptido de la presente invención comprende al menos cuatro y hasta seis fragmentos peptídicos, péptidos de unión a linfocitos B, procedentes de un alérgeno, siendo más preferidos cuatro péptidos.
De acuerdo con la presente invención, la proteína transportadora es el polipéptido PreS de hepatitis B con la siguiente secuencia de aminoácidos (SEQ ID NO: 21):

"Hipoalergénico", como se usa en el presente documento, se refiere a moléculas con un potencial alergénico reducido o nulo (es decir, la reactividad con IgE se determina con ensayos de unión a IgE conocidos en la técnica). Dichas moléculas tienen una capacidad reducida para provocar reacciones alérgicas en un individuo en comparación con la proteína de tipo silvestre de la que proceden estas moléculas.
Los al menos cuatro y hasta seis fragmentos peptídicos fusionados al extremo N y C-terminal de una proteína PreS de la hepatitis B consisten en 20 a 50 aminoácidos consecutivos, del al menos un alérgeno de tipo silvestre y muestran reactividad reducida con IgE en comparación con el alérgeno de tipo silvestre del que proceden los fragmentos de péptido. Estos fragmentos de péptido se diseñan preferentemente para excluir epítopos de linfocitos T específicos de alérgeno que pueden provocar efectos secundarios mediados por linfocitos T. Pueden determinarse e identificarse epítopos de linfocitos T y moléculas que muestran respuesta de linfocitos T reducida mediante métodos conocidos por los expertos en la materia (por ejemplo, Bercovici N. et al. Clin Diagn Lab Immunol. (2000) 7:859-864).
Los al menos cuatro y hasta seis fragmentos de péptido que consisten en 20 a 50 aminoácidos consecutivos de al menos un alérgeno pueden proceder de uno y del mismo alérgeno. En caso de que dos o más fragmentos procedan del mismo alérgeno, estos dos o más fragmentos no están ubicados de manera adyacente en el alérgeno de tipo silvestre y/o tienen un orden en el polipéptido de la presente invención que no corresponde al orden en el alérgeno de tipo silvestre.
La expresión "fragmento de péptido", como se usa en el presente documento, significa una parte/fragmento de un polipéptido o proteína de fusión hipoalergénicos de la invención que procede de la estructura primaria de un alérgeno de tipo silvestre y consiste en de 20 a 50 aminoácidos consecutivos de este alérgeno de tipo silvestre.
Las expresiones "procedente de un alérgeno" y "procedente de al menos un alérgeno de tipo silvestre", como se usan en el presente documento, significan que los fragmentos de péptido para los fines de la presente invención se obtienen directamente de un alérgeno mediante fragmentación o truncamiento. La secuencia de aminoácidos de estos fragmentos de péptido es preferentemente al menos un 80 % idéntica, más preferentemente, al menos un 90 % idéntica, lo más preferentemente, al menos un 95 % idéntica, en particular, un 100 % idéntica, a la serie de la secuencia de aminoácidos del alérgeno de tipo silvestre, de los que proceden los fragmentos de péptido. Sin embargo, los péptidos que no son un 100 % idénticos a los fragmentos de alérgeno de tipo silvestre han de ser capaces de unirse con al menos un 60 %, preferentemente, al menos un 70 %, más preferentemente, al menos un 80 %, lo más preferentemente, al menos un 90 %, de la fuerza a un anticuerpo o anticuerpos, preferentemente a anticuerpos IgG, que están dirigidos contra dichos fragmentos de alérgeno de tipo silvestre. "Al menos un alérgeno de tipo silvestre" significa que el polipéptido de la presente invención puede comprender péptidos de unión a linfocitos B o más de uno, preferentemente dos, más preferentemente tres, alérgenos de tipo silvestre diferentes (es decir, fuentes) (por ejemplo, un péptido procede de Bet v 1, uno de Amb a 1 y uno de PhI p 1 o dos péptidos proceden de Bet v 1 y uno de Amb a
i).
Puede determinarse el grado de identidad de una primera secuencia de aminoácidos respecto de una segunda secuencia de aminoácidos mediante una comparación directa entre ambas secuencias de aminoácido usando ciertos algoritmos. Dichos algoritmos se incorporan, por ejemplo, en varios programas informáticos (por ejemplo, "BLAST 2 SEQUENCES (blastp)" (Tatusova et al. (1999) FEMS Microbiol. Lett. 174:247-25; Corpet F, Nucl. Acids Res. (1988) 16:10881-10890).
Los polipéptidos de la presente invención pueden obtenerse mediante métodos recombinantes o síntesis química. Como alternativa, también es posible, obviamente, obtener las moléculas mediante escisión enzimática o química del alérgeno de tipo silvestre o de un polipéptido/proteína que porte la molécula de interés.
Sorprendentemente, se ha descubierto que las proteínas de fusión de péptido-portador con propiedades mejoradas pueden obtenerse empleando proteínas superficiales del virus de la hepatitis B de humano. Pueden fusionarse desde cuatro hasta seis fragmentos de péptido hipoalergénico al extremo C-terminal y al extremo N-terminal de una proteína PreS de la hepatitis B. Por lo tanto, la presente invención se refiere a proteínas de fusión formadas por al menos 4 y hasta 6 fragmentos de péptido hipoalergénico con una proteína transportadora procedente de los antígenos de superficie del virus de la hepatitis B de humano. Dichas proteínas de fusión usan como portador la proteína PreS. Una realización más preferida de la presente invención es proteínas de fusión en las que se fusionan 4 fragmentos de péptido hipoalergénico a la proteína portadora PreS. Los fragmentos de péptido (hipoalergénico) pueden ser iguales o diferentes y pueden proceder de una o varias proteínas alergénicas y el locus de los péptidos dentro de la proteína de fusión es el extremo C-terminal y el extremo N-terminal de la proteína portadora. Pueden fusionarse de uno hasta tres fragmentos de péptido (hipoalergénico) a cada uno del extremo C-terminal y el extremo N-terminal de tal forma que la suma de los fragmentos de péptido (hipoalergénico) será, por ejemplo, tres o cuatro a seis. Los términos "fusionado" o "proteína de fusión", se refieren a una realización preferida de la invención, dando a entender que la proteína portadora no alergénica y los fragmentos de péptido (hipoalergénico) en los extremos C y N-terminales del portador se expresan y preparan en forma de una sola cadena de polipéptido recombinante.
Una realización extremadamente preferida de la presente invención es proteínas de fusión de la proteína PreS del virus de la hepatitis B, que portan fragmentos de péptido (hipoalergénico) procedentes de un alérgeno específico, de tal forma que uno o dos, preferentemente dos, fragmentos de péptido se fusionan cada uno al extremo C-terminal y al extremo N-terminal del portador. A modo ilustrativo, los polipéptidos preferidos de la presente invención pueden tener la estructura molecular general representada por las siguientes estructuras genéticas:
Estructura 1 Principio de construcción general de realizaciones preferidas

Se entiende que los péptidos A, B, C y D pueden ser iguales o diferentes y pueden proceder del mismo alérgeno para cada proteína de fusión individual o procederán de diferentes alérgenos.
Los péptidos (hipoalergénicos) que se fusionan al extremo N y C-terminal del polipéptido de superficie de un virus de la familia hepadnaviridae o al menos un fragmento de dicho polipéptido de superficie, preferentemente la proteína PreS o un fragmento de la misma, se seleccionan preferentemente entre el grupo que consiste en alérgenos principales del polen de abedul, en particular, Bet v 1 y Bet v 4, alérgenos principales del polen de la hierba timotea, en particular, PhI p 1, Phl p 2, Phl p 5, Phl p 6 y Phl p 7, alérgenos principales de ácaros del polvo doméstico, en particular, Der p 1, Der p 2, Der p 5, Der p 7, Der p 21 y Der p 23, alérgeno principal de gato, Fel d 1, el alérgeno principal de la ambrosía, Amb a 1, los alérgenos principales del cedro japonés, Cry j 1 y Cry j 2, alérgenos principales de abeja, alérgenos principales de avispa, profilinas, especialmente Phl p 7, Phl p 12.
Otros alérgenos adecuados para su uso de acuerdo con la presente invención pueden obtenerse de la tabla 2 a continuación, sin restringirse a dicha tabla.
Tabla 2 Fuentes de péptidos hipoalergénicos










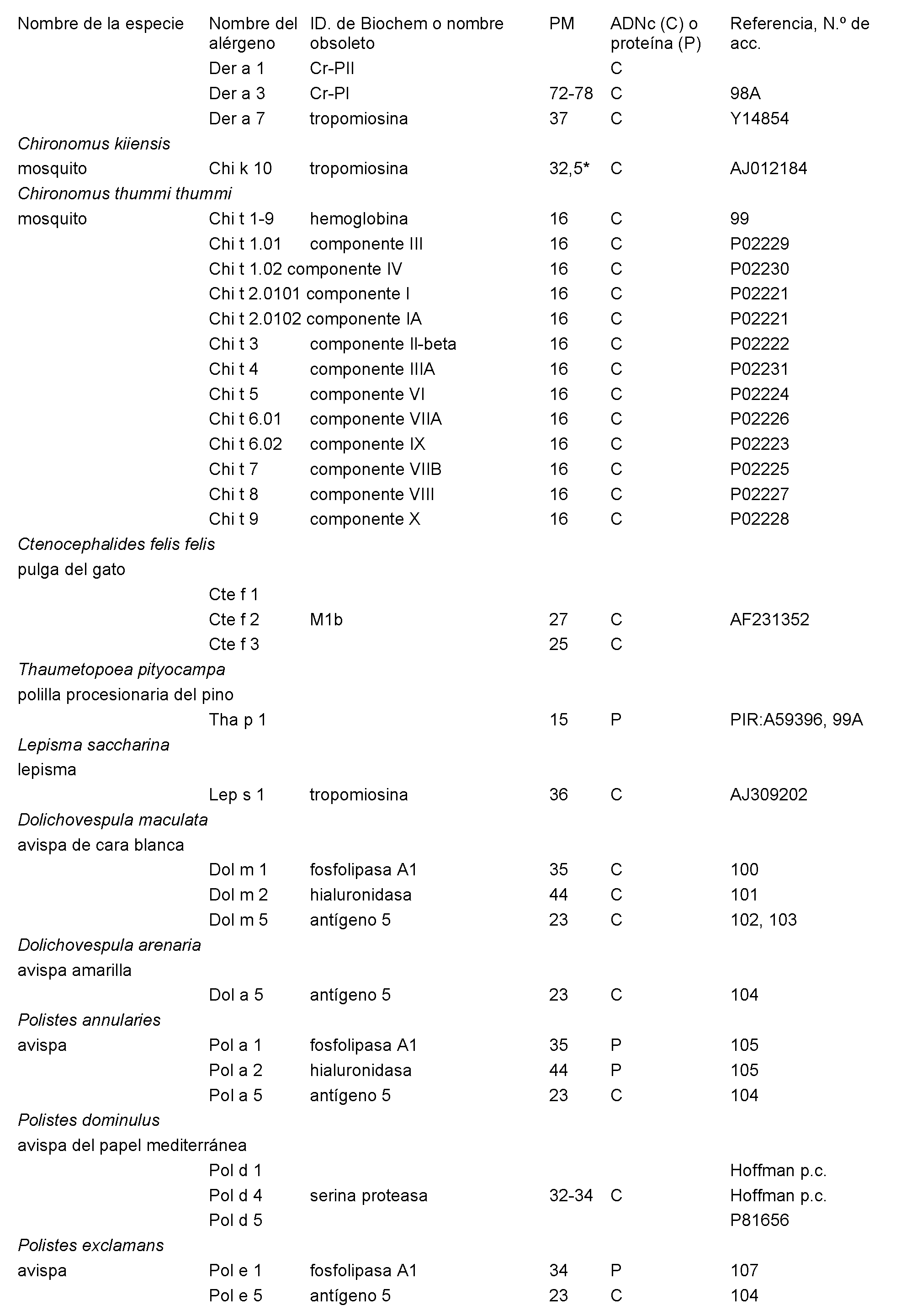

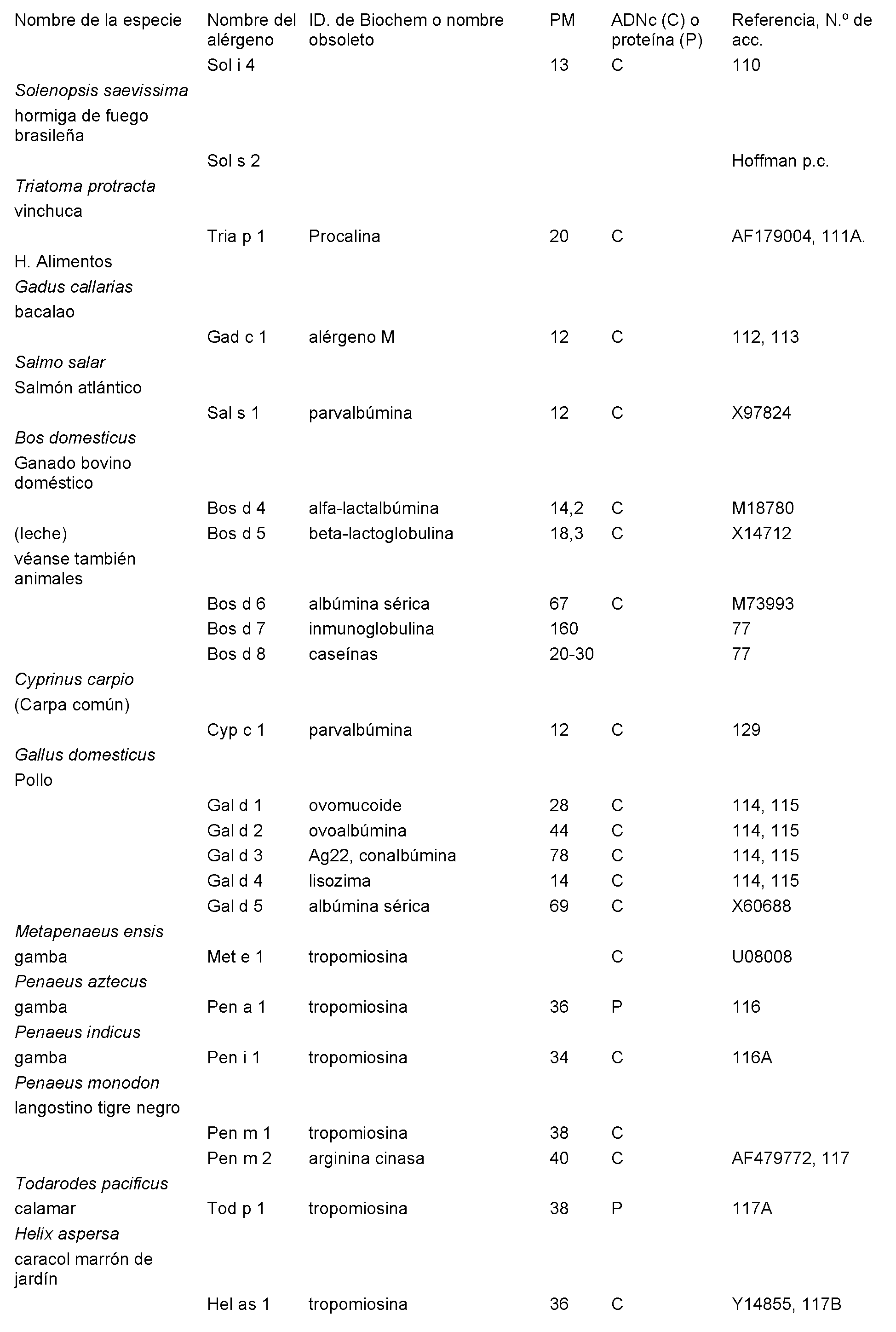

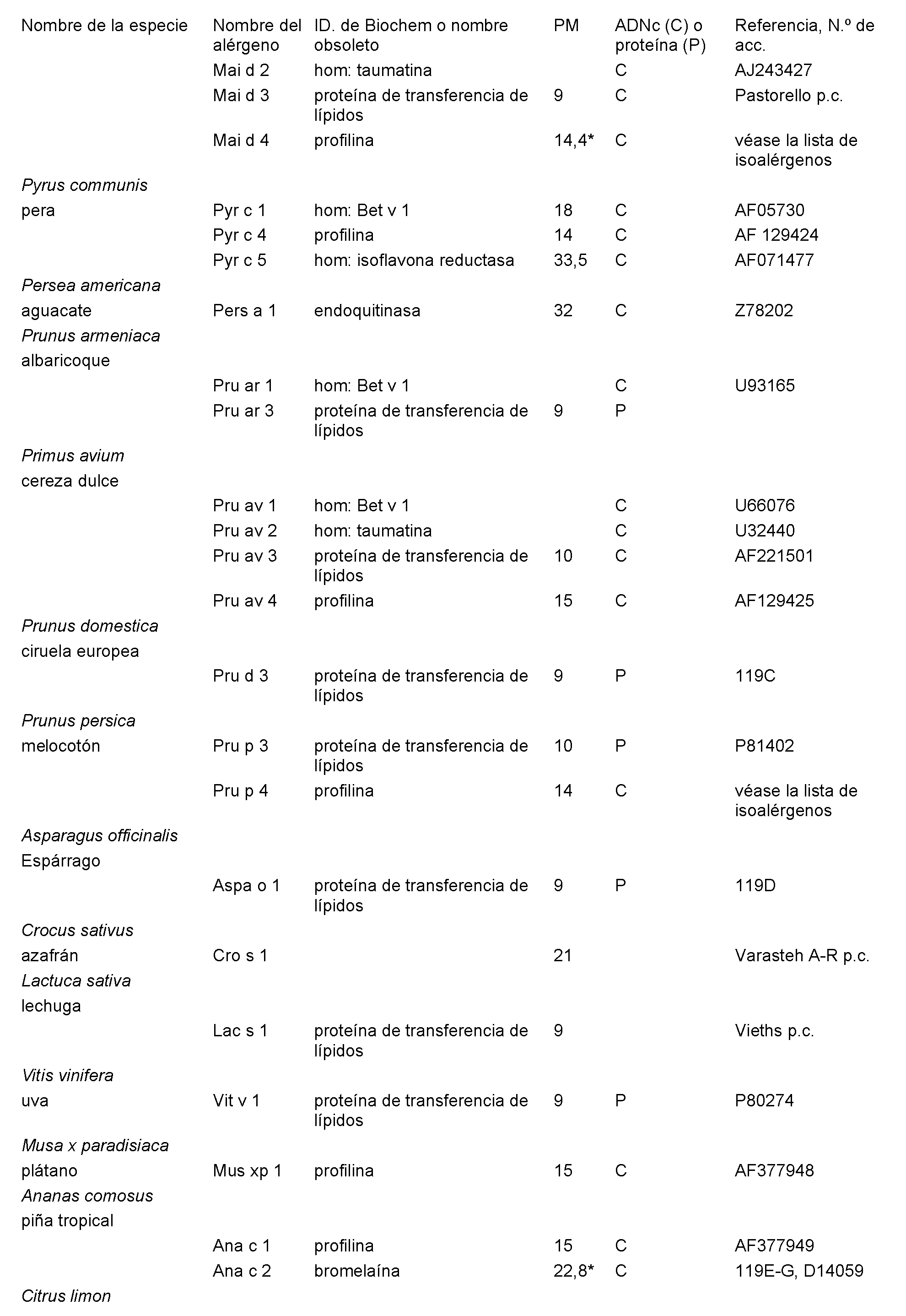



Referencias
1 Marsh, D.G., y L.R. Freidhoff. 1992. ALBE, an allergen database. IUIS, Baltimore, MD, Edición 1.0.
2 Marsh, D. G. et al. 1986. Allergen nomenclatura. Bull WHO 64:767-770.
3 King, T.P. et al. 1964. Biochemistry 3:458-468.
4 Lowenstein, H. 1980. Allergy 35:188-191.
5 Aukrust, L. 1980. Allergy 35:206-207.
6 Demerec, M. et al. 1966. Genetics 54:61-75.
7 Bodmer, J. G.et al. 1991. Immunogenetics 33:301-309.
8 Griffith, I.J. et al. 1991. Int. Arch. Allergy Appl. Immunol. 96:296-304.
9 Roebber, M. et al. 1985. J. Immunol. 134:3062-3069.
10 Metzler, W. J. et al. 1992. Biochemistry 31:5117-5127.
11 Metzler, W. J. et al. 1992. Biochemistry 31:8697-8705.
12 Goodfriend, L. et al. 1979. Fed. Proc. 38:1415.
13 Ekramoddoullah, A. K. M. et al. 1982. Mol. Immunol. 19:1527-1534.
14 Ansari, A. A. et al. 1987. J. Allergy Clin. Immunol. 80:229-235.
15 Morgenstern, J.P. et al. 1991. Proc. Natl. Acad. Sci. USA 88:9690-9694. 16 Griffith, I.J. et al. 1992. Gene 113:263-268.
17 Weber, A. et al. 1986. Biochem. Physiol. 83B:321-324.
18 Weber, A. et al. 1987. Allergy 42:464-470.
19 Stanworth, D. R. et al. 1990. Bulletin WHO 68:109-111.
20 Rafnar, T. et al. 1991. J. Biol. Chem. 266: 1229-1236.
21 Rogers, B.L. et al. 1991. J. Immunol. 147:2547-2552.
22 Klapper, D.G. et al. 1980. Biochemistry 19:5729-5734.
23 Ghosh, B. et al. 1993. J. Immunol. 150:5391-5399.
24 Roebber, M. et al. 1983. J. Immunol. 131:706-711.
25 Lubahn, B., y D.G. Klapper. 1993. J. Allergy Clin. Immunol. 91:338.
26 Roebber, M., y D.G. Marsh. 1991. J. Allergy Clin. Immunol. 87:324.
27 Goodfriend L. et al. Mol Immunol 22: 899-906, 1985.
28 Himly M. et al. FASEB J 17: 106-108, 2003.
28A Nilsen, B. M. et al. 1991. J. Biol. Chem. 266:2660-2668.
29 Wopfner N. et al. Biol Chem 383: 1779-1789, 2002.
29A Jimenez A. et al. 1994. Int Arch Allergy Immunol 105:297-307.
29B Barderas R. et al. Int Arch Allergy Immunol 127: 47-54, 2002.
29C Carnés J. et al. Allergy 56, Suplemento 68: 274, 2001.
29D Giuliani A. et al. Allergy 42: 434-440, 1987.
30 Smith,P.M. et al. 1996. J. Allergy Clin. Immunol. 98:331-343.
31 Suphioglu,C. et al. 1997. FEBS Lett. 402:167-172.
31a Asturias J.A. et al. 1997. Clin Exp Allergy 27:1307-1313.
32 Mecheri, S. et al. 1985. Allergy Appl. Immunol. 78:283-289.
33 Roberts, A.M. et al. 1993. Allergy 48:615-623.
33a Guerin-Marchand,C. et al. 1996. Mol. Immunol. 33:797-806.
34 Klysner, S. et al. 1992. Clin. Exp. Allergy 22: 491-497.
35 Perez, M. et al. 1990. J. Biol. Chem. 265:16210-16215.
36 Griffith, I. J. et al. 1991. FEBS Letters 279:210-215.
37 Ansari, A. A. et al. 1989. J. Biol. Chem. 264:11181-11185.
37a Sidoli,A. et al. 1993. J. Biol. Chem. 268:21819-21825.
38 Ansari, A. A. et al. 1989. Biochemistry 28:8665-8670.
39 Singh, M. B. et al. 1991. Proc. Natl. Acad. Sci. 88:1384-1388.
39a van Ree R. et al. 1995. J Allergy Clin Immunol 95:970-978.
40 Suphioglu,C. y Singh,M.B. 1995. Clin. Exp. Allergy 25:853-865.
41 Dolecek,C. et al. 1993. FEBS Lett. 335:299-304.
41A Fischer S. et al. 1996. J Allergy Clin Immunol 98:189-198.
42 Matthiesen, F., y H. Lowenstein. 1991. Clin. Exp. Allergy 21:297-307.
43 Petersen,A. et al. 1995. Int. Arch. Allergy Immunol. 108:55-59.
43A Marknell DeWitt A. et al. Clin Exp Allergy 32: 1329-1340, 2002.
44 Valenta, R. et al. 1994. Biochem. Biophys. Res. Commun. 199:106-118. 46 Esch, R. E., y D. G. Klapper. 1989. Mol. Immunol. 26:557-561.
47 Olsen, E. et al. 1991. J. Immunol. 147:205-211.
48 Avjioglu, A. et al. 1993. J. Allergy Clin. Immunol. 91:340.
52 Kos T. et al. 1993. Biochem Biophys Res Commun 196:1086-92.
53 Diaz-Perales A. et al. 2000. Clin Exp Allergy 30:1403-1410.
54 Ipsen, H., y O.C. Hansen. 1991. Mol. Immunol. 28: 1279-1288.
55 Taniai, M. et al. 1988. FEBS Lett. 239:329-332.
56 Griffith, I.J. et al. 1993. J. Allergy Clin. Immunol. 91:339.
57 Sakaguchi, M. et al. Allergy 45: 309-312, 1990.
57A Yokoyama M. et al. Biochem Biophys Res Commun 275: 195-202, 2000.
57B Midoro-Horiuti T. et al. J Immunol 164: 2188-2192, 2000.
57C Tinghino R. et al. J. Allergy Clin. Immunol. 101: 772-777, 1998.
58 Gross GN et al. Scand J Immunol 8: 437-441, 1978.
58A Obispo TM et al. Clin Exp Allergy 23: 311-316, 1993.
58B Midoro-Horiuti T. et al. Clin Exp Allergy 31: 771-778, 2001.
59 Lombardero M. et al. Clin. Exp. Allergy 24: 765-770, 1994.
60 Villalba, M. et al. Eur. J. Biochem. 216: 863-869, 1993.
60A Asturias JA et al. J Allergy Clin Immunol 100: 365-372, 1997.
60B Batanero E. et al. Eur J Biochem 241: 772-778, 1996.
60C Batanero E. et al. FEBS Lett. 410: 293-296, 1997.
60D Tejera ML et al. J Allergy Clin Immunol 104: 797-802, 1999.
60E Ledesma A. et al. FEBS Lett 466: 192-196, 2000.
60F Barral P. et al. J Immunol 172: 3644-3651,2004.
61 Yi FC et al. Clin Exp Allergy 32: 1203-1210, 2002.
61A Ramos JD et al. Int Arch Allergy Immunol 126: 286-293, 2001.
62 Chua, K. Y. et al. J. Exp. Med. 167: 175-182, 1988.
62A Chua, K. Y. et al. Int. Arch. Allergy Appl. Immunol. 91: 118-123, 1990.
62B Smith AM et al. Int Arch Allergy Immunol 124: 61-63, 2001.
62C Smith AM et al. J Allergy Clin Immunol 107: 977-984, 2001.
63 Smith WA, Thomas WR. Int Arch Allergy Immunol 109:133-140,1996.
64 Lake, F.R. et al. J. Allergy Clin. Immunol. 87: 1035-1042, 1991.
65 Tovey, E. R. et al. J. Exp. Med. 170: 1457-1462, 1989.
66 Yasueda, H., T. Shida, T. Ando, S. Sugiyama y H. Yamakawa. 1991. Allergenic and proteolytic properties of fourth allergens from Dermatophagoides mites. En: "Dust Mite Allergens and Asthma. Report of the 2nd international workshop" A. Todt, Ed., UCB Institute of Allergy, Bruselas, Bélgica, págs. 63-64.
67 Shen, H.-D. et al. Clin. Exp. Allergy 23: 934-940, 1993.
67A O'Neil GM et al. Biochim Biophys Acta,1219: 521-528, 1994.
67B King C. et al. J Allergy Clin Immunol 98: 739-747, 1996.
68 Lind P. et al. J. Immunol. 140: 4256-4262, 1988.
69 Dilworth, R. J. et al. Clin. Exp. Allergy 21: 25-32, 1991.
70 Nishiyama, C. et al. Int. Arch. Allergy Immunol. 101: 159-166, 1993.
70A Trudinger, M. et al. Clin. Exp. Allergy 21: 33-38, 1991.
71 Shen HD et al. Clin Exp Allergy 25: 1000-1006, 1995.
71A Tategaki A. et al. ACI International supl. 1: 74-76, 2000.
72 Aki T. et al. J Allergy Clin Immunol 96: 74-83, 1995.
72A Tsai L. et al. Clin Exp Allergy 29: 1606-1613, 1999.
72B Gafvelin G. et al. J Allergy Clin Immunol 107: 511-518, 2001.
73 van Hage-Hamsten. et al. J. Allergy Clin. Immunol. 91:353, 1993.
74 Varela J. et al. Eur J Biochem 225: 93-98, 1994.
74A Schmidt M. et al. FEBS Lett 370: 11-14, 1995.
75 Eriksson TLJ et al. Eur. J. Biochem. 268: 287-294, 2001.
75A Saame T. et al. Int Arch Allergy Immunol 130: 258-265, 2003.
75B Eriksson TL et al. Eur. J. Biochem. 251 (1-2), 443-447, 1998.
76 Rautiainen J, Rytkonen M, Pelkonen J, Pentikainen J, Perola O, Virtanen T, Zeiler T, Mantyjarvi R. BDA20, a major bovine dander allergen characterised at the sequence level is Bos d 2. Remitido.
77 Gjesing B, Lowenstein H. Ann Allergy 53:602, 1984.
78 de Groot, H. et al. J. Allergy Clin. Immunol. 87:1056-1065, 1991.
79 Konieczny, A. Personal communication; Immunologic Pharmaceutical Corp.
79A Bulone, V. Eur J Biochem 253: 202-211, 1998.
79B Swiss-Prot acc. P81216, P81217.
79C Dandeu J. P. et al. (1993). J. Chromatogr. 621:23-31.
79D Goubran Botros H. et al. 1998. J. Chromatogr. B 710:57-65.
79E Hilger C. et al. Allergy 52: 179-187; y Hilger C. et al. Gene 169:295-296, 1996.
79F Ichikawa K. et al. Clin Exp Allergy, In Press 2001.
80 Fahlbusch B. et al. Allergy 57: 417-422, 2002.
81 McDonald, B. et al. 1988. J. Allergy Clin. Immunol. 83:251.
81A Clarke, A. J. et al. 1984. EMBO J 3:1045-1052.
82 Longbottom, J. L. 1983. Characterisation of allergens from the urines of experimental animals. McMillan Press, Londres, págs. 525-529.
83 Laperche, Y. et al. 1983. Cell 32:453-460.
83A Bush RK et al. 1999. J Allergy Clin Immunol 104:665-671.
83B Aukrust L, Borch SM. 1979. Int Arch Allergy Appl Immunol 60:68-79.
83C Sward-Nordmo M. et al. 1988. Int Arch Allergy Appl Immunol 85:288-294.
84 Shen, et al. J. Allergy Clin. Immunol. 103:S157, 1999.
84A Crameri R. Epidemiology and molecular basis of the involvement of Aspergillus fumigatus in allergic diseases. Contrib. Microbiol. Vol. 2, Karger, Basilea (en imprenta).
84B Shen, et al. (manuscrito enviado), 1999
84C Shen HD et al. Vacuolar serine proteinase: A major allergen of Aspergillus fumigatus. 10th International Congress of Immunology, Abstract, 1998.
85 Kumar A. et al. 1993. J. Allergy Clin. Immunol. 91:1024-1030.
85A Saxena S. et al. 2003. Clin Exp Immunol 134:86-91.
85B Baur X. et al. Allergy 57: 943-945, 2002.
86A Shen HD et al. 1996. Clin Exp Allergy 26:444-451.
86B Shen, et al. Abstract; The XVIII Congress of the European Academy of Allergology and Clinical Immunology, Bruselas, Bélgica, 3-7 de julio de 1999.
87 Shen HD et al. Clin Exp Allergy 29: 642-651, 1999.
87A Shen HD et al. Clin Exp Allergy 25: 350-356, 1995.
87B Shen HD et al. J Lab Clin Med 137: 115-124,2001.
88 Woodfolk JA et al. 1998. J Biol Chem 273:29489-96.
88A Deuell, B. et al. 1991. J. Immunol. 147:96-101.
89 Shen, H.D. et al. 1991. Clin. Exp. Allergy 21:675-681.
89A Horner WE et al. 1995. Int Arch Allergy Immunol 107:298-300.
89B Chang CY et al. J Biomed Sci 9: 645-655, 2002.
90 Yasueda H. et al. Biochem Biophys Res Commun 248: 240-244, 1998. NB: cepa TIMM2782 (Teikyo University Institute for Medical Mycology) igual a la cepa CBS 1878 (Central Bureau von Schimmelkulturen).
90A Onishi Y. et al. Eur J Biochem 261: 148-154, 1999. nB: cepa TIMM2782 (Teikyo University Institute for Medical Mycology) equal to strain CBS 1878 (Central Bureau von Schimmelkulturen).
91 Schmidt M. et al. Eur J Biochem 246:181-185, 1997. NB: cepa ATCC no. 42132 (American Type Culture Collection).
91A Rasool O. et al. Eur J Biochem 267: 4355-4361,2000. NB: cepa ATCC no. 42132 (American Type Culture Collection).
91B NB: ; cepa 4625 (Indian Agricultural Research Institute, PUSA; Nueva Deli, India ).
92 Kuchler, K. et al. 1989. Eur. J. Biochem. 184:249-254.
93 Gmachl, M., y G. Kreil. 1993. Proc. Natl. Acad. Sci. USA 90:3569-3573.
93A Hoffman dR. 1977. J Allergy Clin. Immunol. 59:364-366.
94 Habermann, E. 1972. Science 177:314-322.
95 Hoffman DR, Jacobson RS. 1996. J. Allergy Clin. Immunol. 97:812-821.
95A Hoffman DR, El-Choufani AE, Smith MM, de Groot H. 2001. Occupational allergy to bumblebee venom: Allergens of Bombus terrestris. J Allergy Clin Immunol In press.
95B Helm R. et al. 1996. J Allerg Clin Immunol 98:172-180.
95C Pomes A. et al. 1998. J Biol Chem 273:30801-30807.
96 Arruda LK et al. J Biol Chem 270:19563-19568, 1995.
97 Arruda LK et al. J Biol Chem 270:31196-31201, 1995.
98 Arruda LK et al. Int Arch Allergy Immunol 107:295-297, 1995.
98A Wu CH et al. 1998. J Allergy Clin Immunol 101:832-840.
98B Melen E. et al. 1999. J Allergy Clin Immunol 103:859-64.
98C Wu CH et al. J Biol Chem 271:17937-17943, 1996.
98D Wu CH et al. Molecular Immunol 34:1-8, 1997.
98E Santos ABR et al. 1999. J Allergy Clin Immunol 104:329-337.
98F Asturias JA et al. 1999. J Immunol 162:4342-4348.
99 Mazur, G. et al. 1990. Monog. Allergy 28:121-137.
99A Moneo I. et al. Allergy 58: 34-37, 2003.
100 Soldatova, L. et al. 1993. FEBS Letters 320:145-149.
101 Lu, G. et al. 1994. J. Allergy Clin. Immunol. 93:224.
102 Fang, K. S. F. et al. 1988. Proc. Natl. Acad. Sci., USA 85:895-899.
103 King, T. P. et al. 1990. Prot. Seq. Data Anal. 3:263-266.
104 Lu, G. et al. 1993. J. Immunol. 150: 2823-2830.
105 King, T. P. y Lu, G. 1997. Datos no publicados.
105A King TP et al. 1996. J. Allergy Clin. Immunol. 98:588-600.
106 Hoffman, D.R. 1993. J. Allergy Clin. Immunol. 92:707-716.
107 Hoffman DR. 1992. Datos no publicados.
108 Hoffman DR. J. Allergy Clin. Immunol. 91:187, 1993.
109 Jacobson RS et al. J. Allergy Clin. Immunol. 89:292, 1992.
110 Hoffman DR. J. Allergy Clin. Immunol 91: 71-78, 1993.
111 Schmidt M. et al. FEBS Letters 319: 138-140, 1993.
111A Paddock CD et al. J Immunol 167: 2694-2699, 2001.
112 Elsayed S, Bennich H. Scand J Immunol 3: 683-686, 1974.
113 Elsayed S. et al. Immunochemistry 9: 647-661, 1972.
114 Hoffman, D. R. 1983. J. Allergy Clin. Immunol. 71: 481-486.
115 Langeland, T. 1983. Allergy 38:493-500.
116 Daul CB, Slattery M, Morgan JE, Lehrer SB. 1993. Common Crustacea allergens: identification of B cell epitopes with the shrimp specific monoclonal antibodies. En: "Molecular Biology and Immunology of Allergens" (D. Kraft and A. Sehon, eds.). CRC Press, Boca Raton, págs. 291-293.
116A Shanti KN et al. J. Immunol. 151: 5354-5363, 1993.
117 Yu CJ et al. J Immunol 170: 445-453, 2003.
117A Miyazawa M et al. J. Allergy Clin. Immunol. 98: 948-953, 1996.
117B Asturias JA et al. Int Arch Allergy Immunol 128: 90-96, 2002.
117C Lopata AL et al. J. Allergy Clin. Immunol. 100: 642-648, 1997.
117D Hoffmann-Sommergruber K. et al. Clin. Exp. Allergy 29: 840-847, 1999.
118 Monsalve RI et al. Biochem. J. 293: 625-632 1993.
118A. Monsalve RI et al. 1997. Clin Exp Allergy 27:833-841.
119 Mena, M. et al. Plant Molec. Biol. 20: 451-458, 1992.
119A Palosuo K. et al. J. Allergy Clin. Immunol. 108: 634-638, 2001.
119B Xu H. et al. Gene 164: 255-259, 1995.
119C Pastorello EA et al. J. Allergy Clin. Immunol. 94: 699-707, 1994.
119D Diaz-Perales A. et al. J Allergy Clin Immunol 110: 790-796, 2002.
119E Galleguillos F, Rodriguez JC. Clin Allergy 8: 21-24, 1978.
119F Baur X. Clin Allergy 9: 451-457, 1979.
119G Gailhofer G. et al. Clin Allergy 18: 445-450, 1988.
120 Menendez-Arias, L. et al. 1988. Eur. J. Biochem. 177:159-166.
120A Gonzalez R. et al. Lancet 346:48-49, 1995.
120B Kleine-Tebbe J. et al. J Allergy Clin Immunol 110: 797-804, 2002.
120C Sanchez-Monge R. et al. J. Allergy Clin. Immunol. 106: 955-961, 2000.
121 Gavrovic-Jankulovic M. et al. J Allergy Clin Immun ol 110: 805-810, 2002.
121A Pastorello EA et al. J. Chromatogr. B Biomed. Sci. Appl. 756: 85-93, 2001.
121B Moneo I. et al. J. Allergy Clin. Immunol. 106: 177-182, 2000.
121C Asturias JA et al. 2000. Allergy 55:898-890.
122 Christie, J. F. et al. 1990. Immunology 69:596-602.
122A Baur X. et al. Clin Allergy 12: 9-17, 1982.
122B Onisuka R. et al. Int Arch Allergy Immunol 125: 135-143, 2001.
123 Czuppon AB et al. J Allergy Clin Immunol 92:690-697, 1993.
124 Attanayaka DPSTG et al. 1991. Plant Mol Biol 16:1079-1081.
125 Chye ML, Cheung KY. 1995. Plant Mol Biol 26:397-402.
126 Alenius H. et al. 1993. Int Arch Allergy Immunol 102:61-66.
127 Yeang HY, Cheong KF, Sunderasan E, Hamzah S, Chew NP, Hamid S, Hamilton RG, Cardosa MJ. 1996. The 14.6 kD (REF, Hev b 1) and 24 kD (Hev b 3) rubber particle proteins are recognised by IgE from Spina Bifida patients with Latex allergy. J Allerg Clin Immunol in press.
128 Sunderasan E. et al. 1995. J nat Rubb Res 10:82-99.
129 Swoboda I. et al. 2002. J Immunol. 168:4576-84.
130 Vrtala et al., 2007. J Immunol. 179:1731-1739.
131 Valenta y Niederberger, 2007. J Allergy Clin Immunol. 119(4):826-830.
132 Niespodziana (13.3.2011); J. Allergy & Clin. Immunol. 127(6):1562-1570
De acuerdo con la presente invención, los al menos cuatro y hasta seis péptidos procedentes del al menos un alérgeno de tipo silvestre son péptidos de unión a linfocitos B.
Los "péptidos de unión a linfocitos B" para su uso en la vacunación contra la alergia de acuerdo con la invención proceden de o se encuentran próximos a los sitios de unión a IgE de los alérgenos, pero en sí muestran una reactividad nula o mínima con IgE en comparación con el alérgeno de tipo silvestre (Focke M et al. Clinical & Experimental Allergy 40(2010):385-397). Los requisitos para su producción y selección son conocer la secuencia primaria del alérgeno y tomar en consideración los sitios de unión a IgE. Tras la inmunización, los péptidos de unión a linfocitos B fusionados a un portador inmunogénico adecuado, son capaces de inducir la producción de IgG específicas para el alérgeno que pueden bloquear la unión de IgE al alérgeno. Puede determinarse si la IgG inducida con la proteína de fusión puede reconocer el alérgeno, por ejemplo, evaluando la reactividad de la IgG por el alérgeno completo. Los métodos adecuados incluyen ELISA, transferencia de puntos o ensayos de transferencia de Western. Se prefieren aquellos péptidos que inducen IgG que bloquean la unión de las IgE del paciente al alérgeno.
La presente invención muestra que el uso de péptidos de unión a linfocitos B adecuados, en particular cuando se fusionan cuatro o más a un portador adecuado de acuerdo con la presente invención, permite la inducción de respuestas de IgG que están más enfocadas hacia los epítopos de IgE que aquellas inducidas mediante inmunización, incluso con un alérgeno completo. Además, la invención demuestra que la combinación de los péptidos adecuados y su número con un portador adecuado puede dirigir la respuesta inmunitaria específica de alérgeno hacia una respuesta inmunitaria antialérgica favorable (caracterizada por la inducción preferencial de respuestas de IgG y no de IgE específicas de alérgeno y respuestas de citocinas tolerogénicas (IL-10) y de Th1 (interferón gamma)).
Por otra parte, se observó sorprendentemente que, a pesar del hecho de que carecen de epítopos de linfocitos T específicos de alérgeno, los polipéptidos de acuerdo con la invención que contienen 4 o más péptidos de unión a linfocitos B fusionados a un portador inmunogénico son capaces de reducir las reacciones de linfocitos T específicos de antígeno. Esto se demostró por el hecho de que la presencia de IgG específicas de alérgeno inducidas mediante la vacunación terapéutica con los polipéptidos hipoalergénicos de la presente invención reduce la activación de linfocitos T específica para el alérgeno provocada por la presentación de antígenos facilitada por IgE en las PBMC de individuos humanos alérgicos vacunados. (Fig. 16).
Dichos al menos cuatro y hasta seis péptidos muestran una capacidad de unión a IgE nula o reducida en comparación con el alérgeno de tipo silvestre y dichos al menos cuatro y hasta seis péptidos de unión a linfocitos B muestran una reactividad con linfocitos T nula o sustancialmente nula.
La presencia de epítopos de linfocitos T específicos de alérgeno puede dar lugar a efectos secundarios mediados por linfocitos T no deseados. Por lo tanto, los péptidos que muestran una reactividad con linfocitos T nula o
sustancialmente nula se usan para obtener los polipéptidos de la presente invención.
"Que muestran capacidad de unión a IgE reducida", como se usa en el presente documento, significa que las moléculas de acuerdo con la presente invención muestran una capacidad o actividad de unión a IgE significativamente reducida (una capacidad de unión al menos un 50 % menor, preferentemente al menos un 70 % menor, más preferentemente, al menos un 80 % menor, aún más preferentemente, al menos un 90 % menor, lo más preferentemente al menos un 95 % menor, en comparación con el alérgeno de tipo silvestre) o incluso una total ausencia de unión a IgE.
La actividad/capacidad de unión a IgE de las moléculas como péptidos y proteínas puede determinarse mediante, por ejemplo, un ensayo inmunoabsorbente ligado a enzimas (ELISA) usando, por ejemplo, suero obtenido de un sujeto, es decir, un sujeto alérgico, que se ha estado expuesto previamente al alérgeno de tipo silvestre. En resumen, un péptido que se va a evaluar se recubre sobre los pocillos de una placa de microtitulación. Tras lavar y bloquear los pocillos, se incubó en los pocillos una solución de anticuerpo que consiste en el plasma de un sujeto alérgico, que ha estado expuesto al péptido que se está ensayando o a la proteína de la que procede. Se añade a los pocillos un anticuerpo secundario marcado y se incuba. Posteriormente, se cuantifica la cantidad de unión a IgE y se compara con la cantidad de IgE unida a un alérgeno de tipo silvestre purificado.
Como alternativa, puede determinarse la actividad de unión de un péptido mediante análisis de transferencia de Western. Por ejemplo, se hace correr un péptido que se vaya a ensayar en un gel de poliacrilamida usando SDS-PAGE. Después, se transfiere el péptido a nitrocelulosa y se incuba posteriormente con suero de un sujeto alérgico. Tras la incubación con el anticuerpo secundario marcado, se determina y cuantifica la cantidad de IgE unida.
Otro ensayo que puede usarse para determinar la actividad de unión a IgE de un péptido es un ensayo ELISA de competición. En resumen, se genera una reserva de anticuerpo IgE combinando plasma de sujetos alérgicos que han mostrado mediante ELISA directo que tienen reactividad de IgE con el alérgeno de tipo silvestre. Esta reserva se usa en ensayos ELISA de competición para comparar la unión de IgE al alérgeno de tipo silvestre respecto del péptido ensayado. Después, se determina y cuantifica la unión de IgE al alérgeno de tipo silvestre y al péptido que se está ensayando.
Un "epítopo de linfocitos T" significa una proteína, péptido o polipéptido (por ejemplo, alérgeno) o un fragmento del mismo, para el que un linfocito T tiene un sitio de unión específico de antígeno, siendo el resultado de la unión a dicho sitio de unión la activación del linfocito T. La expresión "que muestra reactividad reducida con linfocitos T", como se usa en el presente documento, se refiere a moléculas que muestran una reactividad con linfocitos T que está significativamente reducida en comparación con la estimulación inducida por el alérgeno de tipo silvestre del que procede la molécula hipoalergénica usando cantidades equimolares en ensayos convencionales conocidos en la técnica (la reactividad con linfocitos T reducida significa al menos un 30 %, preferentemente, al menos un 50 %, más preferentemente, al menos un 70 %, lo más preferentemente, al menos un 90 %, menos de estimulación de moléculas hipoalergénicas, en comparación con el alérgeno de tipo silvestre en cantidades equimolares). En una realización particular preferida de la presente invención, las moléculas pueden "carecer" de epítopos para linfocitos T y por lo tanto, la molécula muestra una reactividad con linfocitos T reducida en los individuos que se van a tratar (es decir, quien va a recibir una molécula de plataforma de valencia presentadora de epítopos). Es probable que, por ejemplo, una molécula derivada de un alérgeno pueda carecer de epítopos para linfocitos T respecto de un individuo o un grupo de individuos, mientras que poseen epítopos para linfocitos T respecto de otros individuos. Los métodos para detectar la presencia de un epítopo para linfocitos T se conocen en la técnica e incluyen ensayos que detectan la proliferación de linfocitos T (tal como la incorporación de timidina). Generalmente, se considera que los inmunógenos que no logran inducir una incorporación estadísticamente significativa de timidina por encima del nivel de fondo (es decir, una p generalmente menor de 0,05 usando métodos estadísticos convencionales) carecen de epítopos para linfocitos T, aunque se apreciará que puede variar la cantidad cuantitativa de incorporación de timidina, dependiendo del inmunógeno que se esté evaluando (véase, por ejemplo, Zhen L. et al. (Infect Immun. (2003) 71:3920-3926)). En general, un índice de estimulación por debajo de aproximadamente 2-3, más preferiblemente menor de aproximadamente 1, indica la ausencia de reactividad con y de epítopos para linfocitos T. También puede determinarse la presencia de epítopos para linfocitos T midiendo la secreción de linfocinas procedentes de linfocitos T de acuerdo con métodos convencionales. Puede calcularse el índice de estimulación (IE) dividiendo la tasa de proliferación (captación de timidina) de las células estimuladas entre la tasa de proliferación de las células no estimuladas solo en medio. Un IE=1 significa falta de estimulación y un IE > 1 indica estimulación de las células. La ubicación y el contenido de epítopos para linfocitos T, en caso de estar presentes, pueden determinarse empíricamente.
Además de la estimulación de linfocitos T, puede determinarse la secreción de citocinas. Por ejemplo, El IFN-gamma y la IL-10, como biomarcadores para una actividad aumentad de linfocitos T reguladores, se han reconocido como citocinas que acompañan a una inmunoterapia contra la alergia exitosa.
Los fragmentos peptídicos de la presente invención están compuestos o consisten preferentemente en los aminoácidos 151 a 177, 87 a 117, 1 a 30, 43 a 70 o 212 a 241 de Phl p 1, los aminoácidos 1 a 33, 8 a 39, 34 a 65 o 66 a 96 de Phl p 2, los aminoácidos 93 a 128, 98 a 128, 26 a 53, 26 a 58, 132 a 162, 217 a 246, 252 a 283 o 176 a 212 de Phl p 5, los aminoácidos 23 a 54, 56 a 90, 73 a 114 o 95 a 127 de Phl p 6, los aminoácidos 1 a 34 o 35 a 70
de la cadena 1 de Fel d 1, los aminoácidos 1 a 34, 35 a 63 o 64 a 92 de la cadena 2 de Fel d 1, los aminoácidos 30 a 59, 50 a 79, 75 a 104, 30 a 74 o 60 a 104 de Bet v 1, los aminoácidos 1 a 30, 52 a 84 o 188 a 222 de Der p 1, los aminoácidos 1 a 33, 21 a 51, 42 a 73, 62 a 103 o 98 a 129 de Der p 2, los aminoácidos 1 a 30, 20 a 50, 50 a 80, 90 a 125, 125 a 155 o 165 a 198 de Der p 7, los aminoácidos 1-35, 36-70, 71-110, 111-145, 140-170, 175-205, 210-250 o 250-284 de Der p 10, los aminoácidos 1 a 35, 35 a 72, 70 a 100 o 90 a 122 de Der p 21, los aminoácidos 1 a 32, 15 a 48 o 32 a 70, 32 a 60, 52 a 84, 32 a 70 (Cys->Ser) de Der p 23, los aminoácidos 19 a 58, 59 a 95, 91 a 120 o 121 a 157 de Alt a 1, los aminoácidos 31 a 60, 45 a 80, 60 a 96 o 97 a 133 de Par j 2, los aminoácidos 1 a 40, 36 a 66, 63 a 99, 86 a 120 o 107 a 145 de Ole e 1, los aminoácidos 25 a 58, 99 a 133, 154 a 183, 277 a 307, 334 a 363, 373 a 402, 544 a 573, 579 a 608, 58 a 99, 125 a 165, 183 a 224, 224 a 261, 252 a 289, 303 a 340, 416 a 457, 460 a 500 o 501 a 542 de Fel d 2, los aminoácidos 19 a 58, 52 a 91, 82 a 119, 106 a 144 o 139 a 180 de Can f 2, los aminoácidos 19 a 56, 51 a 90, 78 a 118, 106 a 145 o 135-174 de Can f 1, los aminoácidos 27 a 70, 70 a 100 o 92 a 132 de Art v 1, los aminoácidos 31 a 70, 80 a 120, 125 a 155, 160 a 200, 225 a 263, 264 a 300, 305 a 350 o 356 a 396 de Amb a 1, los aminoácidos 1 a 34, 35 a 74, 74 a 115, 125 a 165, 174 a 213, 241 a 280, 294 a 333, 361 a 400 o 401 a 438 de Alt a 6, los aminoácidos 1 a 40, 41 a 80, 81 a 120, 121 a 160 de Alt a 2 o fragmentos o variaciones de secuencia de los mismos.
Las secuencias de aminoácido específicas de las moléculas procedentes de alérgeno anteriormente identificadas son (los péptidos en la tabla a continuación que tienen un resto de cisteína N y/o C-terminal (C) que se usa en el polipéptido de la presente invención pueden carecer de dicho resto de cisteína):
Péptido Posición Secuencia SEQ ID NO:
Pep Alt a 1.1 19-58 APLESRQDTASCPVTTEGDYVWKISEFYGRKPEGTYYN 23
SL
Pep Alt a 1.2 59-95 GFNIKATNGGTLDFTCSAQADKLEDHKWYSCGENSFM 24 Pep Alt a 1.3 91-120 ENSFMDFSFDSDRSGLLLKQKVSDDITYVA 25 Pep Alt a 1.4 121-157 TATLPNYCRAGGNGPKDFVCQGVADAYITLVTLPKSS 26 Pep Alt a 2.1 1-40 MHSSNNFFKDNIFRSLSKEDPDYSRNIEGQVIRLHWDW 27
AQ
Pep Alt a 2.2 41-80 LLMLSAKRMKVAFKLDIEKDQRVWDRCTADDLKGRN 28
GFKR
Pep Alt a 2.3 81-120 CLQFTLYRPRDLLSLLNEAFFSAFRENRETIINTDLEYAA 29 Pep Alt a 2.4 121-160 KSISMARLEDLWKEYQKIFPSIQVITSAFRSIEPELTVYT 30 Pep Alt a 2.5 161-190 CLKKIEASFELIEENGDPKITSEIQLLKAS 31 Pep Alt a 6.1 1-34 MTITKIHARSVYDSRGNPTVEVDIVTETGLHRAI 32 Pep Alt a 6.2 35-74 VTETGLHRAIVPSGASTGSHEACELRDGDKSKWGGKGV 33
TK
Pep Alt a 6.3 74-115 APALIKEKLDVKDQSAVDAFLNKLDGTTNKTNLGANAI 34
LGVS
Pep Alt a 6.4 125-165 EKGVPLYAHISDLAGT KKPYVLPVPF 35
QNVLNGGSHAGGRLA
Pep Alt a 6.5 174-213 CEAPTFSEAMRQGAEVYQKLKALAKKTYGQSAGNVGD 36
EGG
Pep Alt a 6.6 241-280 IKIAMDVASSEFYKADEKKYDLDFKNPDSDKSKWLTYE 37
QL
Pep Alt a 6.7 294-333 VSIEDPFAEDDWEAWSYFFKTYDGQIVGDDLTVTNPEFI 38
K
Péptido Posición Secuencia SEQ ID NO: Pep Alt a 6.8 361-400 AKDAF GAGWGVMV SHRSGETEDVTIADIWGLRSGQIK 39
TG
Pep Alt a 6.9 401-438 APARSERLAKLNQILRIEEELGDNAVYAGNNFRTAVNL 40 Pep Amb a 1.1 31-70 EILPVNETRRLTTSGAYNIIDGCWRGKADWAENRKALA 41
DC
Pep Amb a 1.2 80-120 GGKDGDIYTVTSELDDDVANPKEGTLRFGAAQNRPLWI 42
IFE
Pep Amb a 1.3 125-155 IRLDKEMW N SDKTIDGRGAKVEIINAGFTL 43 Pep Amb a 1.4 160-200 NVIIHNINMHDVKVNPGGLIKSNDGPAAPRAGSDGDAIS 44
IS
Pep Amb a 1.5 225-263 GTTRLTVSNSLFTQHQFVLLFGAGDENIEDRGMLATVA 45
F
Pep Amb a 1.6 264-300 NTFTDNVDQRMPRCRHGFFQWNNNYDKWGSYAIGGS 46 Pep Amb a 1.7 305-350 ILSQGNRFCAPDERSKKNVLGRHGEAAAESMKWNWRT 47
NKDVLENGA
Pep Amb a 1.8 356-396 GVDPVLTPEQSAGMIPAEPGESALSLTSSAGVLSCQPGA 48
PC
Pep Art v 1.1 27-70 SKLCEKTSKTYSGKCDNKKCDKKCIEWEKAQHGACHK 49
REAGKES
Pep Art v 1.2 70-100 SCFCYFDCSKSPPGATPAPPGAAPPPAAGGS 50 Pep Art v 1.3 92-132 APPPAAGGSPSPPADGGSPPPPADGGSPPVDGGSPPPPST 51
H
Can f 1 P e p 1 19-56 QDTPALGKDTVAVSGKWYLKAMTADQEVPEKPDSVTP 52
M
Can f 1 Pep 2 51-90 DSVTPMILKAQKGGNLEAKITMLTNGQCQNITWLHKT 53
SE
Can f 1 Pep 3 78-118 CQNITWLHKTSEPGKYTAYEGQRWFIQPSPVRDHYIL 54
YC
Can f 1 Pep 4 106-145 QPSPVRDHYILYCEGELHGRQIRMAKLLGRDPEQSQEA 55
LE
Can f 1 Pep 5 135-174 RDPEQSQEALEDFREFSRAKGLNQEILELAQSETCSPGG 56
Q
Can f 2 Pep 1 19-58 QEGNHEEPQGGLEELSGRWHSVALASNKSDLIKPWGHF 57
RV
Can f 2 Pep 2 52-91 PWGHFRVFIHSMSAKDGNLHGDILIPQDGQCEKVSLTAF 58
K
Can f 2 Pep 3 82-119 CEKVSLTAFKTATSNKFDLEYWGHNDLYLAEVDPKSYL 59
Péptido Posición Secuencia SEQ ID NO: Can f 2 Pep 4 106-144 NDLYLAEVDPKSYLILYMINQYNDDTSLVAHLMVRDLS 60
R
Can f 2 Pep 5 139-180 VRDLSRQQDFLPAFESVCEDIGLHKDQIWLSDDDRCQ 61
GSRD
Fel d 2 Pep 1 25-58 EAHQSEIAHRFNDLGEEHFRGLVLVAFSQYLQQC 62 Fel d 2 Pep 2 99-133 CTVASLRDKY GEMADCCEKKEPERNECFLQHKDDN 63 Fel d 2 Pep 3 154-183 NEQRFLGKYLYEIARRHPYFYAPELLYYAE 64 Fel d 2 Pep 4 277-307 CADDRADLAKYICENQDSISTKLKECCGKPV 65 Fel d 2 Pep 5 334-363 VEDKEVCKNYQEAKDVFLGTFLYEYSRRHP 66 Fel d 2 Pep 6 373-402 LAKEYEATLEKCCATDDPPACYAHVFDEFK 67 Fel d 2 Pep 7 544-573 EKQIKKQSALVELLKHKPKATEEQLKTVMG 68 Fel d 2 Pep 8 579-608 VDKCCAAEDKEACFAEEGPKLVAAAQAALA 69 Fel d 2 Pep 9 58-99 CPFEDHVKLVNEVTEFAKGCVADQSAANCEKSLHELLG 70
DKLC
Fel d 2 Pep 10 125-165 CFLQHKDDNPGFGQLVTPEADAMCTAFHENEQRFLGK 71
YLYE
Fel d 2 Pep 11 183-224 EEYKGVFTECCEAADKAACLTPKVDALREKVLASSAKE 72
RLKC
Fel d 2 Pep 12 224-261 CASLQKFGERAFKAWSVARLSQKFPKAEFAEISKLVTD 73 Fel d 2 Pep 13 252-289 FAEISKLVTDLAKIHKECCHGDLLECADDRADLAKYIC 74 Fel d 2 Pep 14 303-340 CGKPVLEKSHCISEVERDELPADLPPLAVDFVEDKEVC 75 Fel d 2 Pep 15 416-457 CELFEKLGEYGFQNALLVRYTKKVPQVSTPTLVEVSRSL 76
GKV
Fel d 2 Pep 16 460-500 CTHPEAERLSCAEDYLSWLNRLCVLHEKTPVSERVTK 77
C
Fel d 2 Pep 17 501-542 CTESLVNRRPCFSALQVDETYVPKEFSAETFTFHADLCT 78
LPE
Pep Ole e 1.1 1-40 EDIPQPPVSQFHIQGQVYCDTCRAGFITELSEFIPGASLR 79 Pep Ole e 1.2 36-66 GASLRLQCKDKENGDVTFTEVGYTRAEGLYS 80 Pep Ole e 1.3 63-99 GLYSMLVERDHKNEFCEITLISSGRKDCNEIPTEGWA 81 Pep Ole e 1.4 86-120 GRKDCNEIPTEGWAKPSLKFKLNTVNGTTRTVNPL 82 Pep Ole e 1.5 107-145 LNTVNGTTRTVNPLGFFKKEALPKCAQVYNKLGMYPP 83
NM
Pep Par j 2.1 31-60 GEEACGKWQDIMPCLHFVKGEEKEPSKEC 84 Pep Par j 2.2 45-80 CLHFVKGEEKEPSKECCSGTKKLSEEVKTTEQKREA 85 Pep Par j 2.3 60-96 CCSGTKKLSEEVKTTEQKREACKCIVRATKGISGIKN 86 Pep Par j 2.4 97-133 ELVAEVPKKCDIKTTLPPITADFDCSKIQSTIFRGYY 87
Péptido Posición Secuencia SEQ ID NO: Der p 1 Pep 1 1-30 TNACSINGNAPAEIDLRQMRTVTPIRMQGG 88 Der p 1 Pep 2 52-84 NQSLDLAEQELVDCASQHGCHGDTIPRGIEYIQ 89 Der p 1 Pep 3 85-115 HNGWQESYYRYVAREQSCRRPNAQRFGISN 90 Der p 1 Pep 4 99-135 REQSCRRPNAQRFGISNYCQIYPPNVNKIREALAQTH 91 Der p 1 Pep 5 145-175 KDLDAFRHYDGRTIIQRDNGYQPNYHAVNIV 92 Der p 1 Pep 6 155-187 GRTIIQRDNGYQPNYHAVNIVGYSNAQGVDYWI 93 Der p 1 Pep 7 175-208 VGYSNAQGVDYWIVRNSWDTNWGDNGYGYFAANI 94 Der p 1 Pep 8 188-222 VRNSWDTNWGDNGYGYFAANIDLMMIEEYPYWIL 95 Der p 1 Pep 1,2 1-41 TNACSINGNAPAEIDLRQMRTVTPIRMQGGCGSCWAFS 143
GVA
Der p 1 Pep 2,2 42-82 ATESAYLAYRNQSLDLAEQELVDCASQHGCHGDTIPRG 144
IEYIQ
Der p 1 Pep 9 27-57 MQGGCGSCWAFSGVAATESAYLAYRNQSLD 145 Der p 2 Pep 1 1-33 DQVDVKDCANHEIKKVLVPGCHGSEPCIIHRGK 96 Der p 2 Pep 2 21-51 CHGSEPCIIHRGKPFQLEAVFEANQNSKTAK 97 Der p 2 Pep 3 42-73 EANQNSKTAKIEIKASIEGLEVDVPGIDPNAC 98 Der p 2 Pep 4 62-103 EVDVPGIDPNACHYMKCPLVKGQQYDIKYTWIVPKIAP 99
KSEN
Der p 2 Pep 5 98-129 APKSENVWTVKVMGDNGVLACAIATHAKIRD 100 Der p 5 Pep 1 1-35 MEDKKHDYQNEFDFLLMERIHEQIKKGELALF YLQ 101 Der p 5 Pep 2 25-60 KKGELALFYLQEQINHFEEKPTKEMKDKIVAEMDTI 102 Der p 5 Pep 3 65-95 DGVRGVLDRLMQRKDLDIFEQYNLEMAKKSG 103 Der p 5 Pep 4 78-114 DLDIFEQYNLEMAKKSGDILERDLKKEEARVKKIEV 104 Der p 7 Pep 1 1-30 DPIHYDKITEEINKAVDEAVAAIEKSETFD 105 Der p 7 Pep 2 20-50 VAAIEKSETFDPMKVPDHSDKFERHIGIIDL 106 Der p 7 Pep 3 50-80 LKGELDMRNIQVRGLKQMKRVGDANVKSEDG 107 Der p 7 Pep 4 90-125 VHDDWSMEYDLAYKLGDLHPNTHVISDIQDFWEL 108 Der p 7 Pep 5 125-155 LSLEVSEEGNMTLTSFEVRQFANWNHIGGL 109 Der p 7 Pep 6 165-198 LSDVLTAIFQDTVRAEMTKVLAPAFKKELERNNQ 110 Der p 10 Pep 1 1-35 MEAIKKKMQAMKLEKDNAIDRAEIAEQKARDANLR 111 Der p 10 Pep 2 36-70 AEKSEEEVRALQKKIQQIENELDQVQEQLSAANTK 112 Der p 10 Pep 3 71-110 LEEKEKALQTAEGDVAALNRRIQLIEEDLERSEERLKIA 113
T
Der p 10 Pep 4 111-145 AKLEEASQSADESERMRKMLEHRSITDEERMEGLE 114 Der p 10 Pep 5 140-170 RMEGLENQLKEARMMAEDADRKYDEVARKLA 115 Der p 10 Pep 6 175-205 DLERAEERAETGESKIVELEEELRWGNNLK 116
Péptido Posición Secuencia SEQ ID NO: Der p 10 Pep 7 210-250 SEEKAQQREEAHEQQIRIMTTKLKEAEARAEFAERSVQ 117
KLQ
Der p 10 Pep 8 250-284 QKEVDRLEDELVHEKEKYKSISDELDQTFAELTGY 118 Der p 21 Pep 1 1-35 MFIVGDKKEDEWRMAFDRLMMEELETKIDQVEKGL 119 Der p 21 Pep 2 35-72 LHLSEQYKELEKTKSKELKEQILRELTIGENFMKGAL 120 Der p 21 Pep 3 70-100 GALKFFEMEAKRTDLNMFERYNYEFALESIK 121 Der p 21 Pep 4 90-122 YNYEFALESIKLLIKKLDELAKKVKAVNPDEYY 122 Der p 23 Pep 1 1-32 MANDNDDDPTTTVHPTTTEQPDDKFECPSRFG 123 Der p 23 Pep 2 15-48 PTTTEQPDDKFECPSRFGYFADPKDPHKFYICSN 124 Der p 23 Pep 3 32-70 GYFADPKDPHKFYICSNWEAVHKDCPGNTRWNEDEE 125
TCT
Der p 23 Pep 4 32-60 GYFADPKDPHKFYICSNWEAVHKDCPGNT 146 Der p 23 Pep 5 42-70 KFYICSNWEAVHKDCPGNTRWNEDEETCT 147 Der p 23 Pep 6 32-70* GYFADPKDPHKFYISSNWEAVHKDSPGNTRWNEDEETS 148
(Cys- T
>Ser)
Bet v 1 Pep 1 30-59 LFPKVAPQAIS SVENIEGNGGPGTIKKJSF 126 Bet v 1 Pep 2 50-79 GPGTIKKISFPEGFPFKYVKDRVDEVDHTN 127 Bet v 1 Pep 3 75-104 YDHTNFKYNY SYIEGGPIGDTLEKISNEIK 128 Bet v 1 Pep A 30-74 LFPKVAPQAISSVENIEGNGGPGTIKKISFPEGFPFKYVK 143
DRVDE
Bet v 1 Pep B 60-104 PEGFPFKYVKDRVDEVDHTNFKYNYSVIEGGPIGDTLEK 144
ISNEIKI
Fel d 1 cadena 1 Pep 1-34 EICPAVKRDVDLFLTGTPDEYVEQVAQYKALPWC 129 1
Fel d 1 cadena 1 Pep 35-70 LENARILKNCVDAKMTEEDKENALSLLDKIYTSPLC 130 2
Fel d 1 cadena 2 Pep 1-34 VKMAITCPIFYDVFFAVANGNELLLDLSLTKVNAC 131 1
Fel d 1 cadena 2 Pep 35-63 TEPERTAMKKIQDCYVENGLISRVLDGLVC 132 2
Fel d 1 cadena 2 Pep 64-92 CMTTISSSKDCMGEAVQNTVEDLKLNTLGR 133 3
Phl p 5 Pep 1 98-128 CGAASNKAFAEGLSGEPKGAAESSSKAALTSK 134 Phl p 5 Pep 2 26-58 ADLGY GP ATP AAP AAGYTP ATP AAP AEA AP AGKC 135 Phl p 5 Pep 3 132-162 AYKLAYKTAEGATPEAKYDAYVATLSEALRIC 136 Phl p 5 Pep 4 217-246 CEAAFNDAIKASTGGAYESYKFIPALEAAVK 137 Phl p 5 Pep 5 252-283 TVATAPEVKYTVFETALKKAITAMSEAQKAAKC 138 Phl p 5 Pep 6 176-212 CAEEVKVIPAGELQVIEKVDAAFKVAATAANAAPAND 139
K
Phl p 5 Pep 1a 93-128 CFVATFGAASNKAFAEGLSGEPKGAAESSSKAALTSK 141
Péptido Posición Secuencia SEQ ID NO:
Phl p 5 Pep 2b 26-53 ADLGYGPATPAAPAAGYTPATPAAPAEAC 142 Phl p 5 Pep 7 59-91 ATTEEQKLIEKINAGFKAALAAAAGVQPADKYR 22 Phl p 1 Pep 1 151-171 HVEKGSNPNYLALLVKYVNGDGDWAVC 1
Phl p 1 Pep 2 87-117 EPVWHITDDNEEPIAPYHFDLSGHAFGAMAC 2
Phl p1 Pep 3 1-30 IPKVPPGPNITATYGDKWLDAKSTWYGKPTGC 3
Phl p 1 Pep 4 43-70 GYKDVDKPPFSGMTGCGNTPIFKSGRGC 4
Phl p1 Pep 5 212-241 CVRYTTEGGTKTEAEDVIPEGWKADTSYESK 5
Phl p 2 Pep 1 1-33 VPKVTFTVEKGSNEKHLAVLVKYEGDTMAEVELC 6
Phl p 2 Pep 2 28-39 CVEKGSNEKHLAVLVKYEGDTMAEVELREHGSD 7
Phl p 2 Pep 3 34-65 REHGSDEWVAMTKGEGGVWTFDSEEPLQGPFNC 8
Phl p 2 Pep 4 66-96 CFRFLTEKGMKNYFDDWPEKYTIGATYAPEE 9
Phl p 6 Pep 1 23-54 GKATTEEQKLIEDVNASFRAAMATTANVPPAD 10 Phl p 6 Pep 2 56-90 YKTFEAAFTVSSKRNLADAVSKAPQLVPKLDEVYN 11 Phl p 6 Pep 3 95-127 AADHAAPEDKY EAFVLHFSEALRIIAGTPEVHA 12 Phl p 6 Pep 4 73-114 DAVSKAPQLVPKLDEVYNAAYNAADHAAPEDKY 13 i Cisteínas intercambiadas por serinas (marcadas en negrita)
Las expresiones "fragmentos de los mismos" y "variaciones de secuencia de los mismos" se refieren a péptidos que se deducen de las moléculas procedentes de alérgenos divulgadas en el presente documento y muestran propiedades bioquímicas (por ejemplo, la capacidad para prevenir la unión de IgE al alérgeno del que proceden estas moléculas) que son comparables o idénticas a dichas moléculas derivadas de alérgeno. Los fragmentos para los fines de la presente invención constan de 20 a 50 restos de aminoácido consecutivos de la molécula procedente de alérgeno. La expresión "variación de secuencia" incluye modificaciones de los péptidos, tal como fragmentación (véase lo anterior), sustituciones de aminoácido (en particular, pueden intercambiarse restos de cisteína o metionina por serina, alanina u otros aminoácidos o derivados de aminoácidos naturales o no naturales), eliminaciones o adiciones. "Variación de secuencia" también se refiere a dichas moléculas procedentes de alérgeno de la tabla anterior, en donde al menos 1, preferentemente al menos 2, más preferentemente al menos 3, aún más preferentemente al menos 4 (5, 6, 7, 8, 9, 10, 15, 20) restos de aminoácido se añaden al extremo C y/o N-terminal.
Cabe destacar que el alérgeno citado en el presente documento como "alérgeno de clon 30" es un alérgeno procedente del ácaro del polvo doméstico, Dermatophagoides pteronyssinus y consiste en la siguiente secuencia: MANDNDDDPTTTVHPTTTEQPDDKFECPSRFGYFADPKDPHKFYICSNWEAVHKDCP GNTRWNEDEETCT (SEQ ID NO: 140; véase también el documento WO 2007/124524). Mientras tanto, se ha asignado el nombre del alérgeno Der p 23 al alérgeno de clon 30. Esto significa que Der p 23 y alérgeno de clon 30 son sinónimos.
De acuerdo con la presente invención, la proteína PreS de hepatitis B comprende al menos dos fragmentos peptídicos de unión a linfocitos B procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo N-terminal y al menos dos fragmentos peptídicos de unión a linfocitos B procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo C-terminal.
Al menos dos de dichos tres péptidos de unión a linfocitos B pueden ser idénticos.
El polipéptido de la presente invención puede usarse como vacuna en el tratamiento o la prevención de una alergia en un ser humano o un animal.
El polipéptido se administra preferentemente a un individuo en una cantidad de 0,01 microgramos por kg de peso corporal a 5 mg/kg de peso corporal, preferentemente de 0,1 microgramos por kg de peso corporal a 10 microgramos por kg de peso corporal.
Los polipéptidos de la presente invención pueden administrarse a un individuo en una cantidad de al menos 10 pg, preferentemente, de al menos 20 pg por polipéptido. La cantidad máxima de polipéptidos a administrar puede variar, pero preferentemente es menor de 100 pg, más preferentemente, menor de 50 pg, aún más preferentemente, 40 pg o menos, por polipéptido.
La cantidad de polipéptidos que puede combinarse con los excipientes para producir una forma farmacéutica unitaria variará dependiendo del hospedador tratado y del modo de administración particular. La dosis de la vacuna puede variar dependiendo de factores tales como la patología, la edad, el sexo y el peso del individuo y de la capacidad del anticuerpo para provocar una respuesta deseada en el individuo. Las pautas posológicas se pueden ajustar para proporcionar la respuesta terapéutica óptima. Por ejemplo, pueden administrarse varias dosis divididas a diario o puede reducirse la dosis proporcionalmente según indiquen las exigencias de la situación terapéutica. También puede variarse la dosis de la vacuna para provocar una respuesta a la dosis preventiva óptima, dependiendo de las circunstancias. Por ejemplo, pueden administrarse los polipéptidos y la vacuna de la presente invención a un individuo en intervalos de varios días, una o dos semanas o incluso meses, siempre dependiendo del nivel de inducción de IgG específica para el alérgeno.
El polipéptido/vacuna puede aplicarse entre 2 y 10, preferentemente entre 2 y 7, aún más preferentemente, hasta 5 y lo más preferentemente hasta 3 veces. El intervalo de tiempo entre las vacunaciones posteriores puede seleccionarse para que sea entre 2 semanas y 5 años, preferentemente entre 1 mes y hasta 3 años, más preferentemente entre 2 meses y 1,5 años. La administración repetida del péptido/vacuna de la presente invención puede maximizar el efecto final de una vacunación terapéutica.
Pueden unirse tres o más péptidos de unión a linfocitos B seleccionados entre el grupo que consiste en SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 5, SEQ ID NO: 9, SEQ ID NO: 137, SEQ ID NO: 139, SEQ ID NO: 142 y SEQ ID NO: 10 en N y C-terminal a un polipéptido de superficie del virus de la familia hepadnaviridae, preferentemente, el polipéptido PreS de hepatitis o fragmentos del mismo.
Los polipéptidos de la presente invención que comprenden los al menos tres péptidos de unión a linfocitos B procedentes de al menos un alérgeno de tipo silvestre se seleccionan preferentemente entre el grupo que consiste en SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18 y SEQ ID NO: 19.
Otro aspecto de la presente invención se refiere a una molécula de ácido nucleico que codifica un polipéptido de acuerdo con la presente invención.
Otro aspecto de la presente invención se refiere a un vector que comprende una molécula de ácido nucleico de acuerdo con la presente invención.
Dicho vector es preferentemente un vector de expresión.
El vector que porta la molécula de ácido nucleico de la presente invención puede usarse con fines de clonación o para la producción de vectores de expresión. Dicho vector puede ser un plásmido, cósmido, virus, bacteriófago o cualquier otro vector comúnmente usado en ingeniería genética y puede incluir, además de la molécula de ácido nucleico de la invención, elementos de eucariota o procariota para el control de la expresión, tal como secuencias reguladoras para el inicio y la terminación de la transcripción y/o la traducción, potenciadores, promotores, secuencias de señal y similares.
El vector puede ser un vector bacteriano, fúngico, de insecto, vírico o de mamífero.
El vector de la presente invención puede emplearse preferentemente para fines de clonación y expresión en diversos hospedadores, tales como bacterias, levaduras, hongos filamentosos, células de mamífero, células de insecto, células vegetales o cualquier otra célula procariota o eucariota. Por lo tanto, dicho vector comprende, aparte de un ácido nucleico que codifica una molécula hipoalergénica o proteína de fusión de acuerdo con la presente invención, secuencias reguladoras específicas del hospedador.
Otro aspecto de la presente invención se refiere a un hospedador que comprende una molécula de ácido nucleico o un vector de acuerdo con la presente invención.
La molécula de ácido nucleico y el vector de acuerdo con la presente invención pueden introducirse en un hospedador adecuado. Dicha molécula puede incorporarse en el genoma del hospedador. El vector puede existir de manera extracromosómica en el citoplasma o incorporarse en el cromosoma del hospedador.
Otro aspecto de la presente invención se refiere a una formulación de vacuna que comprende al menos un polipéptido de acuerdo con la presente invención. Por ejemplo, puede contener al menos dos, tres y más preferentemente al menos cuatro polipéptidos de este tipo.
La vacuna puede comprender al menos uno, preferentemente, al menos dos, preferentemente, al menos tres, preferentemente, al menos cuatro, preferentemente al menos 5, polipéptidos que tienen una secuencia de aminoácidos seleccionada entre el grupo que consiste en SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151 y SEQ ID NO: 152. Dependiendo de la composición, dicha vacuna puede usarse en el tratamiento y/o la prevención de alergias al polen
de gramíneas, alergias al polen de abedul, alergias al ácaro del polvo doméstico o una combinación de dichas alergias en individuos que padecen dichas alergias o que se encuentran en riesgo de padecerlas.
El término "prevenir", como se usa en el presente documento, abarca medidas no solo para prevenir la aparición de la enfermedad, tal como la reducción de factores de riesgo, sino también a detener su progresión y reducir sus consecuencias una vez establecida. "Prevenir" también significa prevenir la sensibilización de un individuo que se encuentra en riesgo de adquirir una alergia.
Como se usa en el presente documento, el término "tratamiento" o los equivalentes gramaticales abarcan la mejora y/o reversión de los síntomas de enfermedad (por ejemplo, alergia). Un compuesto que provoca una mejora en cualquier parámetro asociado con la enfermedad, cuando se usa en los métodos de exploración de la presente invención, puede identificarse, por tanto, como un compuesto terapéutico. El término "tratamiento" se refiere tanto a tratamiento terapéutico como a medidas profilácticas o preventivas.
De acuerdo con una de las realizaciones más preferidas de la presente invención, la vacuna comprende polipéptidos que tienen la secuencia de aminoácidos de SEQ ID NO: 14, s Eq ID NO: 15, SEQ ID NO: 16 y SEQ ID NO: 17. De acuerdo con otra realización preferida de la presente invención, la vacuna comprende polipéptidos que tienen la secuencia de aminoácidos de SEQ ID NO: 18 y/o SEQ ID NO: 19.
De acuerdo con una realización particularmente preferida de la presente invención, la vacuna comprende polipéptidos de la presente invención que comprenden fragmentos de alérgeno procedentes de alérgenos del ácaros del polvo doméstico. Se prefieren particularmente los restos de aminoácido 1 a 33, 21 a 51, 42 a 73, 62 a 103 o 98 a 129 de Der p 2, los restos de aminoácido 1 a 30, 20 a 50, 50 a 80, 90 a 125, 125 a 155 o 165 a 198 de Der p 7, los restos de aminoácido 1 a 35, 35 a 72, 70 a 100 o 90 a 122 de Der p 21, los aminoácidos 1 a 32, 15 a 48 o 32 a 70, 32 a 60, 52 a 84, 32 a 70 (Cys->Ser) de Der p 23, los restos de aminoácido 1 a 30, 1 a 41, 27 a 57, 42 a 82, 52 a 84, 85 a 115, 99 a 135, 145 a 175, 155 a 187, 175 a 208 o 188 a 222 de Der p 1. Más preferentemente, la vacuna comprende al menos uno de los polipéptidos de SEQ ID NO: 149 a 152 (mostrados en las Fig. 18A-D).
El polipéptido/vacuna de la presente invención puede administrarse 4 veces por año de tratamiento a lo largo de un periodo de tratamiento total de 1 a 5 años, preferentemente durante 2 a 3 años. De dichas 4 administraciones anuales, 3 se aplican dentro de un periodo de 6 a 12, preferentemente, 8, semanas que tienen intervalos de 3 a 6 semanas, preferentemente 4 semanas, entre la administración 1 y 2 y otras 3 a 6 semanas, preferentemente 4 semanas, entre las administraciones 2 y 3. La cuarta aplicación se aplica de 3 a 7 meses después de la tercera administración. En caso de que el periodo de tratamiento total sea superior a 1 año, se aplica la misma pauta posológica en los siguientes años de tratamiento.
Para el tratamiento de las alergias estacionales (por ejemplo, alergias al polen, tales como alergia al polen de gramíneas o alergias al polen de abedul), las administraciones 1, 2 y 3 se programan preferentemente antes de la estación respectiva con exposición al alérgeno (estación de polen) y la cuarta administración se pauta después de la estación.
La formulación de vacuna de acuerdo con la presente invención puede formularse como se conoce en la técnica y ajustarse según sea necesario al modo de administración de dicha formulación de vacuna.
Las formas de administración preferidas de la formulación de vacuna (de la presente invención) incluyen todos los regímenes de administración convencionales descritos y sugeridos para la vacunación en general y específicamente para la inmunoterapia contra alergias (por vía oral, transdérmica, intravenosa, intranasal, por vía mucosa, rectal, etc.). Sin embargo, se prefiere particularmente administrar las moléculas y proteínas de acuerdo con la presente invención por vía subcutánea o intramuscular.
La formulación de vacuna de acuerdo con la presente invención puede comprender únicamente una proteína de la cápside vírica o fragmentos de la misma de un miembro del género hepadnaviridae. Dicha formulación comprende además preferentemente al menos un adyuvante, excipiente farmacéuticamente aceptable y/o conservante.
A fin de aumentar la inmunogenicidad de las moléculas hipoalergénicas de acuerdo con la presente invención, pueden usarse adyuvantes, por ejemplo, en un medicamento de acuerdo con la presente invención. Un adyuvante de acuerdo con la presente invención es un agente auxiliar que, cuando se administra junto con o en paralelo con un antígeno, aumenta su inmunogenicidad y/o influencia la calidad de la respuesta inmunitaria. Por lo tanto, el adyuvante puede, por ejemplo, tener una influencia considerable en el alcance de la respuesta inmunitaria humoral o celular. Los adyuvantes convencionales son, por ejemplo, compuestos de aluminio, compuestos que contienen lípidos o microbacterias inactivadas.
En general, los adyuvantes pueden ser de diferentes formas, a condición de que sean adecuados para su administración a seres humanos. Los ejemplos adicionales de dichos adyuvantes son emulsiones de aceite de origen mineral o vegetal, compuestos minerales, tales como fosfato o hidróxido de aluminio o fosfato de calcio, productos
bacterianos y derivados, tales como P40 (procedente de la pared celular de Corynebacterium granulosum), lípido A de monofosforilo (MPL, derivado del LPS) y derivados del péptido de muramilo y conjugados del mismo (derivados de componentes de microbacterium), alumbre, adyuvante incompleto de Freund, liposina, saponina, escualeno, etc. (véase, por ejemplo, Gupta R. K. et al. (Vaccine 11:293-306 (1993)) y Johnson A. G. (Clin. Microbiol. Rev. 7:277-289). El medicamento de la presente invención comprende como adyuvante, más preferentemente, alumbre.
Se prefiere particularmente una combinación de más de una proteína de fusión que contiene péptidos hipoalergénicos y la proteína PreS de hepatitis B. Estas combinaciones pueden derivarse de péptidos de un solo alérgeno o de diferentes alérgenos de la misma fuente de alérgeno o de varias fuentes de alérgeno diferentes.
Se prefiere particularmente una mezcla de cuatro proteínas de fusión que contienen péptidos hipoalergénicos de Phl p 1, Phl p 2, Phl p 5 y Phl p 6 y la proteína PreS del virus de la hepatitis B.
Se prefiere particularmente una proteína de fusión o una mezcla de 2 proteínas de fusión que contienen péptidos hipoalergénicos de Bet v 1 y la proteína PreS del virus de la hepatitis B.
Se prefiere una mezcla de al menos 2 proteínas de fusión que contienen péptidos hipoalergénicos de alérgenos de ácaros del polvo doméstico, más preferentemente seleccionados entre Der p 1, Der p 2, Der p 5, Der p 7, Der p 21 y Der p 23 y la proteína PreS del virus de la hepatitis B. Más preferentemente, la mezcla contiene 3 proteínas de fusión que contienen péptidos hipoalergénicos procedentes de Der p 1, Der p 2 y Der p 23. Se prefiere particularmente que la mezcla comprenda al menos uno, preferentemente, al menos dos, más preferentemente al menos tres, de los polipéptidos mostrados en las SEQ ID NO: 149 a 152 (véase también las Fig. 18A-D).
En general, pueden prepararse formulaciones de vacuna específicas de acuerdo con la presente invención para el tratamiento o la prevención de diferentes alergias mediante la combinación de polipéptidos hipoalergénicos de la invención que representan los alérgenos clínicamente relevantes de una fuente de alérgeno. Los métodos para determinar los alérgenos clínicamente relevantes de una fuente de alérgeno se conocen en la técnica y se han descrito con anterioridad (Valenta y Niederberger, 2007, J Allergy Clin. Immunol, 119 (4): 826-830). Los polipéptidos hipoalergénicos de dicha formulación de vacuna específica se adsorben preferentemente en un adyuvante que puede usarse en humanos (por ejemplo, hidróxido de aluminio) y la mezcla se administra 3-4 veces al año durante 1-3 años, aplicando más de 10 pg de cada polipéptido presente en la formulación de vacuna por dosis.
Dichas formulaciones pueden comprender de 10 ng a 1 g, preferentemente de 100 ng a 10 mg, especialmente de 0,5 pg a 200 pg de dicha molécula hipoalergénica o anticuerpo.
Puede usarse una proteína hipoalergénica de acuerdo con la presente invención para fabricar un medicamento para el tratamiento o la prevención de una infección vírica y/o una alergia en un ser humano o animal.
Dicho medicamento comprende además preferentemente al menos un adyuvante, excipiente farmacéuticamente aceptable y/o conservante.
El medicamento de acuerdo con la presente invención puede usarse para inmunización tanto activa (administración de la proteína hipoalergénica y/o moléculas de la invención) como pasiva (anticuerpos dirigidos a la proteína hipoalergénica y/o moléculas de la invención).
Dicho medicamento puede comprender de 10 ng a 1 g, preferentemente de 100 ng a 10 mg, especialmente de 0,5 pg a 200 pg de dicha molécula hipoalergénica, molécula de ácido nucleico, vector, hospedador o anticuerpo.
El medicamento se administra preferentemente a un individuo en una cantidad de 0,01 pg/kg de peso corporal a 5 mg/kg de peso corporal, preferentemente de 0,1 pg/kg de peso corporal a 10 pg/kg de peso corporal.
El medicamento puede administrarse en una dosis que contiene una cantidad absoluta de 5 - 200 pg, más preferentemente, de 10 - 80 pg, más preferentemente, de 20 - 40 pg de cada uno, incluyendo el polipéptido hipoalergénico.
La pauta posológica particular, es decir, dosis, tiempo y repetición, dependerá del individuo particular y del historial médico del individuo. Las consideraciones empíricas, tales como la semivida, contribuirán generalmente a la determinación de la dosis. Puede determinarse y ajustarse la frecuencia de la administración durante el transcurso de la terapia.
Más preferentemente, la pauta posológica para el medicamento consistirá en 4 inyecciones subcutáneas anuales de una y la misma dosis a lo largo de un periodo de tratamiento de 2 a 3 años. De dichas 4 inyecciones subcutáneas anuales, 3 se aplican dentro de un periodo de 6 a 12, preferentemente, 8, semanas que tienen intervalos de 3 a 6 semanas, preferentemente 4 semanas, entre la inyecciones 1 y 2 y otras 3 a 6 semanas, preferentemente 4 semanas, entre las inyecciones 2 y 3. La cuarta inyección se aplica de 4 a 6 meses después de la tercera administración. Se aplica la misma pauta posológica en los siguientes años de tratamiento.
Para el tratamiento de las alergias estacionales (por ejemplo, alergias al polen, tales como alergia al polen de gramíneas o alergias al polen de abedul), las administraciones 1, 2 y 3 se programan preferentemente antes de la estación respectiva con exposición al alérgeno (estación de polen) y la cuarta administración se pauta después de la estación.
El individuo a quien se administra el medicamento de acuerdo con la presente invención es preferentemente un individuo o animal que tiene o se encuentra en riesgo de tener una alergia.
Los sujetos que tienen o que se encuentran en riesgo de tener una alergia, afección alérgica, trastorno alérgico o enfermedad alérgica incluyen sujetos con una afección alérgica existente o una predisposición conocida o sospechada hacia el desarrollo de un síntoma asociado con o provocado por una afección alérgica. Por lo tanto, el sujeto puede tener una afección, trastorno o enfermedad alérgica crónica activa, un episodio alérgico agudo o una afección, trastorno o enfermedad alérgica latente. Ciertas afecciones alérgicas se asocian con factores ambientales estacionales o geográficos. Por lo tanto, los sujetos en riesgo incluyen aquellos que se encuentran en riesgo de padecer una afección basándose en un historial personal o familiar previo y la estación o la ubicación física, pero en los que la afección o un síntoma asociado con la afección pueden no manifestarse en ese momento en el sujeto.
La administración del medicamento de acuerdo con la presente invención, que comprende al menos una molécula hipoalergénica como se describe en el presente documento, a un individuo puede prevenir la sensibilización de dicho individuo o puede inducir una respuesta inmunitaria adecuada a los alérgenos. En caso de que el medicamento de la presente invención se use para prevenir la sensibilización, debe administrarse a un individuo antes del primer contacto con dicho alérgeno. Por lo tanto, se prefiere administrar el medicamento de acuerdo con la presente invención a neonatos y niños. También se ha dado el caso de que la administración del medicamento de acuerdo con la presente invención a gestantes inducirá la formación de anticuerpos dirigidos contra alérgenos en el feto. Es especialmente beneficioso usar moléculas hipoalergénicas de acuerdo con la presente invención para dichas terapias, debido a la ausencia de epítopos de linfocitos T específicos de alérgeno, pueden reducirse significativamente los efectos secundarios durante el transcurso de la inmunoterapia con alérgeno o incluso evitarse completamente.
Puede usarse una proteína de la cápside vírica de un virus de la familia hepadnaviridae como transportador en medicamentos o vacunas.
Una de las ventajas de dicho transportador es que no solo puede exponerse el antígeno fusionado o conjugado al sistema inmunitario, sino que también se induce una respuesta inmunitaria contra la proteína de la cápside de un hepadnavirus. Por consiguiente, dicha vacunación puede proporcionar prevención y/o tratamiento de enfermedades provocadas por hepadnavirus. El virus es preferentemente de la especie del virus de la hepatitis B humana.
Otro aspecto de la presente invención se refiere a una molécula hipoalergénica derivada de Phl p 5 (n.° de GenBank X7435) que tiene un truncamiento C y/o N-terminal y carece sustancialmente de capacidad de unión a IgE.
El polen de gramíneas es una de las fuentes estacionales externas más potentes de alérgenos transportados por el aire responsable de la fiebre del heno y el asma alérgica.
Más de un 40 % de los individuos alérgicos presentan reactividad de IgE con alérgenos de polen de gramíneas, que se dividen en más de 11 grupos. Más de un 80 % de los pacientes alérgicos al polen de gramíneas reaccionan con alérgenos del grupo 5.
Los alérgenos del grupo 5 son proteínas altamente homólogas no glucosiladas con una masa molecular en el intervalo de 25-33kD. Se han clonado y/o caracterizado inmunológicamente varios alérgenos del grupo 5.
El ensayo para reducir la actividad alérgica introduciendo mutaciones puntuales, mutaciones de varios aminoácidos seguidos o eliminaciones no mostró efecto (Schramm G, et al. J Immunol 1999; 162: 2406-1435). Las regiones de unión a IgE de Phl p 5 (Flicker S, et al. J Immunol 2000; 165: 3849-3859) ya se han descrito y se ha resuelto la estructura tridimensional (Maglio O, et al. 2002. Protein Eng. 15:635-642).
Se ha observado que en particular, los péptidos Phl p 5 de acuerdo con la presente invención, que están truncados en C y/o N-terminal y que carecen de capacidad de unión a IgE, pueden emplearse para la vacunación activa de individuos.
Esta molécula truncada carece sustancialmente de epítopos para linfocitos T y, por lo tanto, carece de reactividad de linfocitos T específica de Phl p 5.
Como se ha indicado anteriormente, pueden reducirse significativamente los efectos secundarios de la inmunoterapia con alérgeno o incluso evitarse en caso de que las moléculas hipoalergénicas carezcan sustancialmente de epítopos de linfocitos T específicos de alérgeno.
Las moléculas de Phl p 5 que carecen de epítopos para linfocitos T están formadas por los aminoácidos 93 a 128, 98
a 128, 26 a 53, 26 a 58 o 252 a 283 de Phl p 5 o fragmentos o variaciones de secuencia del mismo.
En particular, estas moléculas truncadas muestran una reactividad con linfocitos T específicos de antígeno sustancialmente baja o nula y son, no obstante, capaces de provocar una respuesta inmunitaria adecuada dirigida contra el alérgeno de tipo silvestre.
El Phl p 5 truncado hipoalergénico puede estar compuesto por los aminoácidos 132 a 162, 217 a 246 o 176 a 212 de Phl p 5 o variaciones de secuencia del mismo.
Estas moléculas hipoalergénicas comprenden uno o más epítopos para linfocitos T pero carecen de capacidad de unirse a IgE.
Se prefieren moléculas de Phl p 1 truncadas que carecen de epítopos para linfocitos T, que están compuestas por los aminoácidos 1 a 30, 43 a 70, 87 a 117, 151 a 171 o 214 a 241 de Phl p 1 o variaciones de secuencia de las mismas fusionadas a una proteína transportadora vírica, preferentemente la proteína PreS de hepatitis B.
Se prefieren moléculas de Phl p 2 truncadas que carecen de epítopos para linfocitos T, que están compuestas por los aminoácidos 1 a 33, 8 a 39, 34 a 65 o 66 a 96 de Phl p 2 o variaciones de secuencia de las mismas fusionadas a una proteína transportadora vírica, preferentemente la proteína PreS de hepatitis B.
Se prefieren moléculas de Phl p 6 truncadas que carecen de epítopos para linfocitos T, que están compuestas por los aminoácidos 23 a 54, 56 a 90, 73 a 114 o 95 a 127 de Phl p 6 o variantes de secuencia de las mismas fusionadas a una proteína transportadora vírica, preferentemente la proteína PreS de hepatitis B.
Se prefieren moléculas de Bet v 1 truncadas que carecen de epítopos para linfocitos T, que están compuestas por los aminoácidos 30 a 59, 50 a 79, 75 a 104, 30 a 74 o 60 a 104 de Bet v 1.
Se prefieren combinaciones o mezclas de moléculas truncadas de Phleum pratense que carecen de epítopos para linfocitos T, fusionadas a una proteína transportadora vírica, preferentemente la proteína PreS de hepatitis B, como se ha descrito anteriormente.
Se prefieren combinaciones o mezclas de moléculas de Phleum pratense que carecen de epítopos para linfocitos T, que están compuestas por una o más de dichas proteínas de fusión de Phl p 1, Phl p 2, Phl p 5 y Phl p 6 truncadas, como se ha descrito anteriormente.
Una molécula hipoalergénica procedente de Fel d 1 (n.° de GenBank X62477 y X62478) puede tener un truncamiento C y/o N-terminal y carecer de capacidad de unión a IgE.
Las alergias a animales afectan a hasta un 40 % de los pacientes alérgicos. En el ambiente doméstico, las alergias a las mascotas más populares, gatos y perros, son particularmente prevalentes y están conectadas con síntomas perennes. Los alérgenos de los animales están presentes en la caspa, epitelio, saliva, suero u orina. La exposición a los alérgenos puede producirse tanto mediante contacto directo con la piel como por la inhalación de partículas que portan los alérgenos. Se ha demostrado que los alérgenos principales de gatos y perros están ampliamente presentes y pueden detectarse incluso en hogares sin mascotas y en lugares públicos, por ejemplo, escuelas. Esto puede atribuirse al elevado y creciente número de hogares que tienen mascotas en los países industrializados (aproximadamente un 50 %) y a la elevada estabilidad de los alérgenos, que son transportados y distribuidos.
Se ha identificado Fel d 1 como el principal alérgeno de gato, que es reconocido por más de un 90 % de los pacientes alérgicos a los gatos. Fel d 1 representa una glucoproteína ácida de 38 kDa con una función biológica desconocida. Consta de dos heterodímeros idénticos unidos de manera no covalente, que, de nuevo, están formados por dos cadenas de polipéptido unidas de manera antiparalela por tres enlaces disulfuro. La cadena 1 y la cadena 2 están codificadas por genes distintos, consistiendo cada uno en 3 exones. Fel d 1 recombinante (rFel d 1), expresada como una proteína de fusión de cadena 2 a cadena 1, se ha generado en E. coli. Esta Fel d 1 recombinante puede imitar completamente las propiedades inmunológicas del alérgeno de tipo silvestre.
Son adecuados los péptidos procedentes del alérgeno principal de gato, Fel d 1 y que carezcan de capacidad de unión a IgE, por ejemplo, para inmunoterapia y vacunación profiláctica contra la alergia. Los péptidos sintéticos derivados de Fel d 1, como Phl p 5 y los péptidos derivados de alérgeno divulgados en el presente documento, son capaces de inducir una respuesta de IgG, es decir, la producción de los denominados "anticuerpos de bloqueo" o "anticuerpos protectores". Estos anticuerpos impiden la unión de IgE al alérgeno Fel d 1. De este modo, puede lograrse una reducción significativa en los síntomas de alergia.
La molécula truncada divulgada en el presente documento puede mostrar reactividad reducida con linfocitos T. A fin de evitar o de reducir significativamente los efectos secundarios tardíos, la molécula hipoalergénica derivada de Fel d 1 muestra una reactividad con linfocitos T reducida como se define en la presente invención.
El Fel d 1 truncado está compuesto preferentemente por los aminoácidos 1 a 34 o 35 a 70 de la cadena 1 de Fel d 1, los aminoácidos 1 a 34, 35 a 63 o 64 a 92 de la cadena 2 de Fel d 1 o variaciones de secuencia del mismo.
En el presente documento se divulgan moléculas hipoalergénicas que están compuestas por o que comprenden los aminoácidos 1 a 33, 21 a 51, 42 a 73, 62 a 103 o 98 a 129 de Der p 2, los aminoácidos 1 a 30, 20 a 50, 50 a 80, 90 a 125, 125 a 155 o 165 a 198 de Der p 7, los aminoácidos 1 a 35, 35 a 72, 70 a 100 o 90 a 122 de Der p 21, los aminoácidos 1 a 32, 15 a 48 o 32 a 70, 32 a 60, 52 a 84, 32 a 70 (Cys->Ser) de Der p 23, los aminoácidos 19 a 58, 59 a 95, 91 a 120 o 121 a 157 de Alt a 1, los aminoácidos 31 a 60, 45 a 80, 60 a 96 o 97 a 133 de Par j 2, los aminoácidos 1 a 40, 36 a 66, 63 a 99, 86 a 120 o 107 a 145 de Ole e 1, los aminoácidos 25 a 58, 99 a 133, 154 a 183, 277 a 307, 334 a 363, 373 a 402, 544 a 573, 579 a 608, 58 a 99, 125 a 165, 183 a 224, 224 a 261, 252 a 289, 303 a 340, 416 a 457, 460 a 500 o 501 a 542 de Fel d 2, los aminoácidos 19 a 58, 52 a 91, 82 a 119, 106 a 144 o 139 a 180 de Can f 2, los aminoácidos 19 a 56, 51 a 90, 78 a 118, 106 a 145 o 135-174 de Can f 1, los aminoácidos 27 a 70, 70 a 100 o 92 a 132 de Art v 1, los aminoácidos 31 a 70, 80 a 120, 125 a 155, 160 a 200, 225 a 263, 264 a 300, 305 a 350 o 356 a 396 de Amb a 1, los aminoácidos 1 a 34, 35 a 74, 74 a 115, 125 a 165, 174 a 213, 241 a 280, 294 a 333, 361 a 400 o 401 a 438 de Alt a 6, los aminoácidos 1 a 40, 41 a 80, 81 a 120, 121 a 160 de Alt a 2 o fragmentos o variaciones de secuencia de los mismos.
Los métodos para la producción de proteínas de fusión se conocen bien en la técnica y pueden encontrarse en las referencias convencionales de biología molecular, tales como Sambrook et al. (Molecular Cloning, 2.a ed., Cold Spring Harbor Laboratory Press, 1989) y Ausubel et al. (Short Protocols in Molecular Biology, 3.a ed; Wiley and Sons, 1995). En general, una proteína de fusión se produce construyendo en primer lugar un gen de fusión que se inserta en un vector de expresión adecuado, que, a su vez, se usa para transfectar un una célula hospedadora adecuada. En general, las construcciones de fusión recombinantes se producen mediante una serie de digestiones con enzimas de restricción y de reacciones de ligamiento que dan como resultado la incorporación de las secuencias deseadas en un plásmido. En caso de que no estén disponibles los sitios de restricción adecuados, pueden usarse adaptadores o enlazadores oligonucleotídicos sintéticos, como es conocido por los expertos en la materia y como se describe en las referencias citadas anteriormente. Las secuencias de polinucleótido que codifican alérgenos y proteínas nativas pueden ensamblarse antes de su inserción en un vector adecuado o puede insertarse la secuencia que codifica el alérgeno adyacente a una secuencia que codifica una secuencia nativa ya presente en un vector. La inserción de la secuencia dentro del vector ha de ser en fase, de tal forma que la secuencia puede transcribirse en una proteína. Resultará evidente para los expertos habituales en la materia que las enzimas de restricción exactas, los enlazadores y/o adaptadores necesarios, así como las condiciones de reacción exactas, variarán dependiendo de las secuencias y los vectores de clonación empleados. El ensamblaje de construcciones de ADN, sin embargo, es rutinario en la técnica y puede lograrse fácilmente por una persona experta en la materia.
Una ventaja específica e inesperada es que las proteínas de fusión procedentes de moléculas de alérgeno hipoalergénicas truncadas y la proteína PreS de la hepatitis B humana pueden expresarse de manera reproducible en sistemas de expresión convencionales y pueden producirse fácilmente en procesos de alto rendimiento y de manera reproducible en sistemas de expresión convencionales conocidos por los expertos en la materia, más particularmente, usando como sistema de expresión Escherichia coli. Dicho proceso de fabricación comprende normalmente la expresión de las moléculas de acuerdo con la invención mediante el cultivo de células en un biorreactor (por ejemplo, en un fermentador, matraz de agitación), seguido de recogida de las células (por ejemplo, por filtración, centrifugación, etc.) y rotura de las células (por ejemplo, mediante homogeneización a alta presión, sonicación, ciclos de congelación/descongelación, lisis enzimática o química, etc.), purificación de las moléculas (por ejemplo, por cromatografía, filtración, precipitación, ultra/diafiltración, etc.) y formulación del producto final. A fin de obtener un alto rendimiento de las moléculas de acuerdo con la invención, se emplean preferentemente procesos de cultivo de alta densidad celular, mediante la aplicación de fermentación discontinua alimentada.
En el presente documento también se divulga una molécula de ácido nucleico que codifica una proteína de fusión como se describe en el presente documento.
La molécula de ácido nucleico de la presente invención puede emplearse, por ejemplo, para producir dichas moléculas de manera recombinante.
Dicha molécula de ácido nucleico, en otro aspecto de la presente invención, puede estar comprendida en un vector. Este vector es preferentemente un vector de expresión.
La presente invención se ilustra adicionalmente mediante las siguientes figuras y ejemplos, sin embargo, sin limitarse a los mismos.
La Fig. 1 A muestra una representación esquemática del vector HBV_Phlp1_4xP5
La Fig. 1 B muestra una representación esquemática del vector HBV_Phlp2_4xP3
La Fig. 1 C muestra una representación esquemática del vector HBV_Phlp5_V2
La Fig. 1 D muestra una representación esquemática del vector HBV_Phlp6_4xP1
La Fig.2 A muestra la secuencia primaria de la proteína de fusión HBV_PhlP1_4xP5 (BM321, SEQ ID NO: 14) La Fig. 2 B muestra la secuencia primaria de la proteína de fusión HBV_Phlp2_4xP3 (BM322, SEQ ID NO: 15) La Fig. 2 C muestra la secuencia primaria de la proteína de fusión HBV_Phlp5_V2 (BM325, SEQ ID NO: 16) La Fig. 2 D muestra la secuencia primaria de la proteína de fusión HBV_Phlp6_4xP1 (B326, SEQ ID NO: 17) La Fig. 2 E muestra la secuencia primaria de la proteína de fusión HBV_Betv1_4PA (BM31a, SEQ ID NO: 18) La Fig.2 F muestra la secuencia primaria de la proteína de fusión HBV_Betv1_2PA2PB (BM31, SEQ ID NO: 19) La Fig. 2 G muestra la secuencia primaria de la proteína de fusión HBV_Phlp5_V1 (SEQ ID NO: 20)
La Fig. 3 A muestra un gel de SDS Page al 12 % teñido con azul de Coomassie que contiene la proteína de fusión HBV_Phlp1_4xP5 purificada (BM 321, carril 1 y 10: 5 ug de marcador molecular, carril 2, 3, 11 y 125ug de BM321, carril 4 y 132 ug de BM321, carril 5 y 141 ug de BM321, carril 6 y 150,5 ug de BM321, carril 7 y 160,25 ug de BM321, carril 8 y 170,1 ug de BM 321, carril 9 y 180,05 ug de Bm321). Los carriles 1 a 9 son en condiciones reductoras y los carriles 10-18 en no reductoras.
La Fig. 3 B muestra un gel de SDS Page al 12 % teñido con azul de Coomassie que contiene la proteína de fusión HBV_Phlp2_4xP3 purificada (BM 322, carril 1 y 10: 5 ug de marcador molecular, carril 2, 3, 11 y 125ug de BM322, carril 4 y 132 ug de BM322, carril 5 y 141 ug de BM322, carril 6 y 150,5 ug de BM322, carril 7 y 160,25 ug de BM322, carril 8 y 170,1 ug de BM 322, carril 9 y 180,05 ug de Bm322). Los carriles 1 a 9 son en condiciones reductoras y los carriles 10-18 en no reductoras.
La Fig. 3 C muestra un gel de SDS Page al 12 % teñido con azul de Coomassie que contiene la proteína de fusión HBV_Phlp5_V2 purificada (BM 325, carril 1 y 10: 5 ug de marcador molecular, carril 2, 3, 11 y 125ug de BM325, carril 4 y 132 ug de BM325, carril 5 y 141 ug de BM325, carril 6 y 150,5 ug de BM325, carril 7 y 160,25 ug de BM325, carril 8 y 170,1 ug de BM 325, carril 9 y 180,05 ug de BM325). Los carriles 1 a 9 son en condiciones reductoras y los carriles 10-18 en no reductoras.
La Fig. 3 D muestra un gel de SDS Page al 12 % teñido con azul de Coomassie que contiene la proteína de fusión HBV_Phlp6_4xP1 purificada (BM 326, carril 1 y 10: 5 ug de marcador molecular, carril 2, 3, 11 y 125ug de BM326, carril 4 y 132 ug de BM326, carril 5 y 141 ug de BM326, carril 6 y 150,5 ug de BM326, carril 7 y 160,25 ug de BM326, carril 8 y 170,1 ug de BM 326, carril 9 y 180,05 ug de Bm326). Los carriles 1 a 9 son en condiciones reductoras y los carriles 10-18 en no reductoras.
La Fig. 4 demuestra la ausencia de reactividad con IgE de los péptidos de fusión procedentes de alérgenos del polen de gramíneas. Se evaluó la unión a IgE de las proteínas de fusión en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros del número indicado de pacientes alérgicos al polen de gramíneas con proteínas añadidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión de IgE para ninguna de las cuatro proteínas de fusión de péptido-transportador, a) muestra los resultados del ensayo de transferencia por puntos usando HBV_Phlp1_4XP5 (BM321); b) muestra los resultados del ensayo de transferencia por puntos usando HBV_Phlp2_4xP3 (BM322); c) muestra los resultados del ensayo de transferencia por puntos usando HBV_Phlp5_V2 (BM325); d) muestra los resultados del ensayo de transferencia por puntos usando HBV_Phlp6_4xP1 (BM326).
La Fig. 5 muestra la baja actividad alergénica de la proteína de fusión derivada de alérgeno del polen de gramíneas HBV_Phlp1_4xP5 (BM321) determinada mediante la expresión de CD203c en basófilos de pacientes alérgicos. Se incubaron PBMC de pacientes alérgicos al polen de gramíneas con diluciones seriadas de Phl p 1 (barras de color gris claro) o BM321 (barras de color gris oscuro). Se midió la inducción de CD203c como la intensidad media de fluorescencia y los índices de estimulación calculados se muestran en el eje y.
La Fig.6 muestra la baja actividad alergénica de la proteína de fusión derivada de alérgeno del polen de gramíneas HBV_Phlp6_4xP1 (BM326) determinada mediante la expresión de CD203c en basófilos de pacientes alérgicos. Se incubaron PBMC de pacientes alérgicos al polen de gramíneas con diluciones seriadas de Phl p 6 (barras de color gris claro) o BM326 (barras de color gris oscuro). Se midió la inducción de CD203c como la intensidad media de fluorescencia y los índices de estimulación calculados se muestran en el eje y.
La Fig. 7 muestra las respuestas de IgG1 específicas para el alérgeno del polen de hierba timotea en ratones. Se inmunizó a grupos de 4 ratones con 20 ug de proteínas de fusión (proteínas de fusión individuales y combinación de 4 proteínas de fusión) y 10 jg de cada uno (Phl p 1 y 5) o 5 |jg de cada uno (Phl p 2 y 6) del alérgeno de tipo silvestre en las semanas 0 y 3 del estudio, seguido de una inmunización de refuerzo en la semana 17 del estudio. Los antígenos se administraron por vía subcutánea en la región dorsal de los animales. Se recogió sangre en las semanas 0, 3, 6, 9, 12, 17, 20 y 22 del estudio de la vena caudal de los ratones. Se recogió sangre en las semanas del estudio con inmunizaciones un día antes de la inmunización. Se investigaron los sueros inmunes de los ratones respecto de la presencia de IgG1 específica de alérgeno mediante ELISA. Los sueros preinmunes antes de la primera inmunización fueron negativos en todos los animales. Se compararon las proteínas de fusión individuales con la aplicación de una mezcla de proteínas de fusión.
a) Respuesta inmunitaria contra el antígeno rPhl p 1 para HBV_Phlp1_4xP5 (BM321 como componente único), BM321 en una mezcla con BM322, BM325 y BM326 y ratones inmunizados con rPhl p 1.
b) Respuesta inmunitaria contra el antígeno rPhl p 2 para HBV_Phlp2_4xP3 (BM321 como componente único), BM322 en una mezcla con BM321, BM325 y BM326 y ratones inmunizados con rPhl p 2.
c) Respuesta inmunitaria contra el antígeno rPhl p 5 para HBV_Phlp5_V2 (BM325 como componente único), BM325 en una mezcla con BM321, BM322 y BM326 y ratones inmunizados con rPhl p 5.
d) Respuesta inmunitaria contra el antígeno rPhl p 6 para HBV_Phlp6_4xP1 (BM326 como componente único), BM326 en una mezcla con BM321, BM322 y BM325 y ratones inmunizados con rPhl p 6.
La figura 8 muestra la caracterización molecular e inmunológica de las proteínas de fusión recombinantes. A. SDS-PAGE teñido con Coomassie que muestra cuatro proteínas de fusión de PreS con péptidos derivados de Bet v1 (carril 1: 2xPA-PreS, carril 2: 2xPB-PreS, carril 3: 4xPA-PreS, carril 4: 2xPA2xPB-PreS) y el transportador PreS (carril 5). B. Se sondaron proteínas de fusión recombinantes y PreS añadidas por puntos en nitrocelulosa con suero anti-PreS de conejo (carril 1), suero preinmune de conejo (carril 3) y tampón de control para anticuerpos de conejo (carril 3) y anticuerpos monoclonales dirigidos contra péptido P2' (mAb2) (carril 4) y P4' (mAb12) (carril 5) procedentes de Bet v 1 y tampón de control para anticuerpos monoclonales de ratón (carril 6).
La figura 9 A muestra la reactividad con IgE de rBet v 1 y de proteínas de fusión recombinantes de PreS con péptidos derivados de Bet v 1. Se evaluaron sueros de pacientes alérgicos al polen de abedul, de controles no alérgicos y de solo tampón respecto de su reactividad con rBet v 1, las cuatro proteínas de fusión recombinantes (2PA-PreS, 2PB-PreS, 4PA-PreS, 2PA2PB-PreS) y PreS sola transferidas por puntos. Se detectó IgE humana unida con anticuerpos anti-IgE humana marcados con 125I. Las cuentas por minuto (cpm) correspondientes a IgE unida se miden con un contador y y se indican en el eje Y. Los diagramas de cajas muestran los resultados de 50 pacientes alérgicos al polen de abedul.
La figura 9B muestra la activación de basófilos por rBet v1 y las cuatro proteínas de fusión de PreS medida por la regulación positiva de CD 203c. Se expusieron las muestras de sangre de pacientes alérgicos al polen de abedul a concentraciones crecientes (0,001-1 pg/ml) de antígenos, anti-IgE o control de tampón (Co). Se muestran los resultados de un paciente representativo. Se detectó la expresión de CD 203c mediante análisis FACS y se muestra como índice de estimulación (IE (eje y)). Se muestran las medias de mediciones por triplicado y se indican las desviaciones estándar.
La figura 10 muestra las respuestas linfoproliferativas y la producción de citocinas de PBMC de pacientes alérgicos al polen de abedul. Las PBMC de pacientes alérgicos al polen de abedul se han estimulado con cantidades equimolares de rBet v 1, los péptidos derivados de Bet v 1 PA y PB, solo PreS y las proteínas de fusión PreS (es decir, 2PA-PreS, 2PB-PreS, 4PA-PreS, 2PAPB-PreS). Se muestran los índices de estimulación (IE) (ejes y).
(A) Los IE para la concentración más elevada (5 pg/pocillo de Bet v 1 y cantidades equimolares de los péptidos, PreS y proteínas de fusión de PreS) de 6 pacientes alérgicos al polen de abedul se muestran como diagramas de cajas, donde un 50 % de los valores se encuentran dentro de las cajas y los valores no anómalos se encuentran entre las barras.
Las líneas dentro de las cajas indican los valores de la mediana.
(B) Los IE para cuatro concentraciones (1=5 pg/pocillo, 2=2,5 pg/pocillo, 3=1,25 pg/ml, 4=0,63 pg/pocillo de rBet v1 y cantidades equimolares de los péptidos, PreS y proteínas de fusión PreS) se muestran para un paciente representativo.
(C) Se ha medido la producción de citocinas en los sobrenadantes de PBMC de 6 pacientes alérgicos al polen de abedul, estimuladas con 2,5 pg/ml de rBet v1 y cantidades equimolares de los péptidos PA y PB, PreS y cuatro proteínas de fusión. Las concentraciones observadas (pg/ml) (ejes y) después de la estimulación con antígenos se muestran en diagramas de cajas, donde un 50 % de los valores se encuentran dentro de las cajas y los valores no anómalos se encuentran entre las barras. Las líneas dentro de las cajas indican los valores de la mediana.
La figura 11 muestra la inducción de anticuerpos IgG específicos para rBet v 1 y alérgenos homólogos de Bet v 1 tras la inmunización subcutánea mediante proteínas de fusión de PreS en conejos.
(A) Los conejos se han inmunizado con proteínas de fusión (2PA-PreS, 2PB-PreS, 4PA-PreS, 2PAPB-PreS) y rBet v 1 adsorbidas en hidróxido de aluminio (alumbre) (parte superior) o adsorbidas en adyuvante completo de Freund (CFA) (parte inferior). Se midieron las IgG de conejo específicas para rBet v 1 y se presentan los valores medios de la densidad óptica (DO) para mediciones duplicadas (ejes y) para diferentes diluciones de antisuero de conejo (ejes x).
(B1) Alineamiento de múltiples secuencias de Bet v 1 y alérgenos homólogos a Bet v 1 en aliso (Aln g 1), avellano (Cor a 1) y manzano (Mal d 1). Los aminoácidos iguales se indican como puntos, los huecos se indican como guiones. En el lado derecho se muestra el porcentaje de identidad de los alérgenos homólogos a Bet v 1 respecto de Bet v 1. El péptido A (PA, línea discontinua) y el péptido B (PB, línea continua) procedentes de Bet v 1 se encuentran recuadrados.
(B2) Los anticuerpos IgG de antisueros de conejo (rab a-2PA-PreS, rab a -2PB-PreS, rab a -4PA-PreS, rab a -2PAPB-PreS) dirigidos contra rBet v 1, rAln g 1, rCor a 1 y rMal d 1 (eje x) se han medido mediante ELISA. Se muestran las medias de mediciones duplicadas. La densidad óptica (DO) correspondiente a IgG específicas de alérgeno en suero de conejo (post) se presenta en comparación con los sueros preinmunes correspondientes (pre) (ejes y).
(C) Los anticuerpos IgG de conejos inmunizados con rBet v 1 y proteínas de fusión recombinantes (2PA-PreS, 2PB-PreS, 4PA-PreS, 2PAPB-PreS) dirigidas contra seis péptidos derivados de Bet v 1 (P1'-P6') (eje x) se han medido mediante ELISA. Se muestran las medias de los valores de densidad óptica (DO) para mediciones duplicadas (eje y).
La figura 12 muestra la inhibición del suero de conejo anti-2xPA2xPB-PreS contra IgE de pacientes alérgicos en comparación con suero de conejo contra rBet v 1 completo. Se determinó el porcentaje de inhibición de la unión de IgE a rBet v 1 (ejes y) obtenidos con sueros de conejo anti-2xPA2xPB-PreS y anti-rBet v 1 mediante ELISA de
inhibición y se presentan como diagramas de cajas, donde un 50 % de los valores se encuentran dentro de las cajas y los valores no anómalos se encuentran entre las barras. Las líneas dentro de las cajas indican los valores de la mediana. Se muestran los resultados de 21 pacientes alérgicos al polen de abedul.
La figura 13 muestra una titulación de IgG de conejo generadas tras la inmunización con proteínas de fusión PreS que contenían 2 o 4 copias de un péptido derivado de Phl p 6. Para las pruebas de inmunogenicidad, se inmunizaron conejos (conejos New Zealand White) con las diferentes proteínas de fusión usando hidróxido de aluminio como adyuvante. La inducción de anticuerpos específicos se monitorizó en ensayos ELISA. Los resultados muestran que las proteínas de fusión que contienen 4 péptidos son más inmunogénicos que las proteínas de fusión que contienen 2 péptidos.
La figura 14 muestra la inducción de una robusta respuesta de IgG dirigida contra los alérgenos del polen de gramíneas Phl p 1 (A), Phl p2 (B), Phl p 5 (C) y Phl p 6 (D) en pacientes humanos alérgicos al polen de gramíneas después de la inmunización subcutánea con una formulación de vacuna (BM32) que comprende una mezcla de las 4 proteínas de fusión hipoalergénicas de SEQ ID NO: 14, SEQ ID NO: 15, SeQ ID NO: 16 y SEQ ID NO: 17. La determinación de IgG se llevó a cabo mediante ELISA. Se compararon los niveles de IgG antes del tratamiento (pre) con los niveles de IgG después del tratamiento (post).
La figura 15 muestra los resultados de ensayos de proliferación de linfocitos T llevados a cabo en linfocitos T de individuos alérgicos al polen de gramíneas tras la inmunización con una formulación de vacuna que consiste en una mezcla de las 4 proteínas de fusión hipoalergénicas de SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 y SEQ ID NO: 17. La reactividad de linfocitos T se reduce fuertemente o se encuentra ausente si se compara con el polen de gramíneas. El eje y de la gráfica refleja el índice de estimulación.
La figura 16 muestra que las IgG inducidas mediante la terapia con una formulación de vacuna (BM32) que comprende una mezcla de las 4 proteínas de fusión hipoalergénicas de SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 y SEQ ID NO: 17 reduce las respuestas linfoproliferativas contra los alérgenos del polen de gramíneas en PBMC humanas. (a) Configuración experimental. (b) Resultados de los ensayos de proliferación de linfocitos T llevados a cabo en ausencia (+ suero antes) y presencia (+ suero después) de IgG inducidas por el tratamiento. El eje y de la gráfica refleja el índice de estimulación. P1-P5 indican los resultados de diferentes participantes en el estudio.
La figura 17 muestra la configuración de un estudio clínico llevado a cabo en 69 pacientes alérgicos al polen de gramíneas usando la formulación de vacuna BM32 que comprende una mezcla de las 4 proteínas de fusión hipoalergénicas de SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 y SEQ ID NO: 17.
La Fig. 18 A muestra la secuencia primaria de la proteína de fusión HBV Der p2-2xP2-2xP4 (SEQ ID NO: 149) La Fig. 18 B muestra la secuencia primaria de la proteína de fusión HBV Der p2-3xP2-3xP4 (SEQ ID NO: 150) La Fig. 18 C muestra la secuencia primaria de la proteína de fusión HBV Der p23-2xP4-2xP5 (SEQ ID NO: 151) La Fig. 18 D muestra la secuencia primaria de la proteína de fusión HBV Der p23-4xP6 (SEQ ID NO: 152) La figura 19A muestra el cambio en los síntomas nasales inducidos por el tratamiento con 3 inyecciones subcutáneas de la formulación de vacuna BM32 que comprende una mezcla de las 4 proteínas de fusión hipoalergénica de SEQ ID NO: 14, SEQ ID NO: 15, SeQ ID NO: 16 y SEQ ID NO: 17. Barras negras: antes del tratamiento, barras grises: después del tratamiento.
La figura 19B muestra el cambio en el área media de la roncha entre la prueba del pinchazo en la piel titulada y después del tratamiento con la formulación de vacuna BM32. Se llevó a cabo la prueba del pinchazo en la piel titulada usando 8 diluciones seriadas de extracto de polen de gramíneas (sin diluir a 1:128).
La figura 20 muestra la unión a IgE de los péptidos derivados de Der p 2 en comparación con el alérgeno completo probado mediante un ensayo de transferencia por puntos de IgE. Se incubaron sueros de 26 pacientes alérgicos al ácaro del polvo doméstico con péptidos conjugados a KLH punteados y se detectó la IgE unida con anti-IgE humana marcada con 125I. No se detectó unión de IgE para ninguno de los 5 péptidos como en el ejemplo 26.
Ejemplos:
Ejemplo 1: Construcción del plásmido de expresión para HBV_Phlp1_4xP5 (BM321)
Se ensambló el gen de BM321 sintético a partir de oligonucleótidos sintéticos y/o productos de la PCR y se clonaron en un vector convencional adecuado (pMK-RQkanR). El plásmido se purificó a partir de una cepa de E. coli K12 transformada (DH10B-T1R) y la concentración se determinó mediante espectroscopía UV. La secuencia de ADN de BM321 sintética final y con codones optimizados se clonó adicionalmente en el vector de expresión pET28b(+) usando sitios de restricción adecuados (sitio de NcoI en el extremo 5' y de EcoRI en el extremo 3'). El aDn de plásmido se purificó a partir de E. coli K12 DH10B (dam+ dcm+) transformadas y se determinó la concentración mediante espectroscopía UV. La construcción final se verificó mediante secuenciación del inserto. A continuación se muestra un resumen de los datos del plásmido y un mapa del plásmido del vector de expresión final "pBM-321".
R m n l n i BM 21 l n n l r x r i n fin l ET-2

Ejemplo 2: Transformación del plásmido de expresión en el hospedador de expresión para HBV_Phlp1_4xP5 (BM321)
Se transformaron células de E. coli BL21(DE3) químicamente competentes con el plásmido de expresión mediante el método de choque térmico. Las células transformadas se sembraron en placas de LB-agar que consistía en cloruro de sodio al 0,5 %, peptona de soja al 1 %, extracto de levadura al 0,5 %, agar al 1,5 % y 50 pg/ml de kanamicina para la selección. Las células en las placas de LB se cultivaron mediante cultivo durante una noche a 37 °C. Se aislaron las colonias individuales de células de E. coli (BL21 (DE3), se cultivaron en medio LB y se exploraron respecto del crecimiento y la expresión de BM321. Se seleccionó el clon con mejor rendimiento para establecer posteriormente el banco celular maestro.
Ejemplo 3: Preparación de un banco celular maestro para HBV_Phlp1_4xP5 (BM321)
Se usó una alícuota del clon seleccionado para la inoculación de 150 ml de medio de cultivo (composición: cloruro de sodio al 0,5 %, peptona de soja al 1 %, extracto de levadura al 0,5 %, 50 pg/ml de kanamicina). El cultivo del banco celular maestro (MCB) se incubó a 37 °C con agitación constante a 200 rpm hasta que el cultivo alcanzó una densidad óptica DO600 = 1 - 2. Se añadió glicerol para obtener una concentración final de glicerol del 15 % v/v y se separó el MCB en alícuotas en viales de 1 ml y se almacenaron en un congelador ultra profundo a -75 ± 10 °C.
Ejemplo 4: Fermentación discontinua alimentada de alta densidad celular de HBV_PhlP1_4xP5 (BM321) Se inoculó medio de cultivo sintético (100 ml, pH = 6,8, sales y oligoelementos, 10 g/l de glucosa como fuente de carbono) con 1 ml de banco celular maestro (E. coli BL21(DE3) / pBM321) y se cultivó en un matraz de agitación (37 °C, 200 rpm) hasta que se alcanzó un valor diana de densidad óptica DO = 1. Se usó un fermentador de acero inoxidable de 22 l para llevar a cabo la fermentación discontinua alimentada. Para un control automático y reproducible de la alimentación, se programó una receta que permitía una velocidad de crecimiento, una velocidad de alimentación, una duración de la fase discontinua y una duración de la fase de alimentación exponencial específicas predefinidas. A fin de aumentar la velocidad de transferencia de oxígeno al fermentador, se controló la presión retrógrada y se ajustó a 1 bar. El fermentador se esterilizó in situ con el medio de cultivo sintético como se ha mencionado anteriormente y se comenzó la fermentación mediante incubación con precultivo. Tras agotar la glucosa, se comenzó la fase de alimentación exponencial a fin de mantener una velocidad de crecimiento específica de p = 0,25 h-1. Al alcanzarse una DO = 45, se indujo la expresión de BM321 recombinante mediante la adición embolada de IPTG (concentración final de 0,8 mM). El cultivo se recogió a una DO600 = 73. El título de producto de BM321 obtenido de la fermentación discontinua alimentada fue de 1,2 g por litro de caldo de cultivo. A continuación, se enfrió el caldo de cultivo hasta < 20 °C y se centrifugó a 7.000 rpm (5.500 g) a 4 °C durante 15 min. Se separó en alícuotas la biomasa celular húmeda y se almacenó a -75 °C.
Ejemplo 5: Disrupción celular y aclarado
Para la disrupción celular, se descongelaron 748 gramos de biomasa del ejemplo 6 y se subdividieron en alícuotas de 125 gramos y se resuspendieron en un tampón de homogeneización (Tris 20 mM, EDTA 1 mM, Triton X-100 al 0,1 %, pH 11,0) con agitación mecánica a temperatura ambiente durante 30 min. Para la disrupción celular, se aplicó un procedimientos de congelación-descongelación congelando a -75 °C y descongelando posteriormente, seguido de homogeneización mecánica. Se ajustó el pH del homogeneizado a pH = 10,0. Se sometió al homogeneizado celular en bruto a una etapa de centrifugación a 7.000 rpm (5.500 g) a 4 °C durante 30 min. Se sometieron los sobrenadantes a precipitación con PEI (polietilenimina) con agitación mecánica. La materia no soluble se separó mediante una etapa de centrifugación posterior. Los sobrenadantes aclarados se sometieron a la etapa de cromatografía posterior. Ejemplo 6: Purificación por cromatografía de HBV_Phlp1_4xP5 (BM321)
Se cargaron un total de 1840 ml del sobrenadante de precipitación con PEI de la etapa de clarificación como se describe en el ejemplo 7 en una columna Q-Sepharose f F de 5 x 30 cm y se equilibró con tampón A (Tris-HCl, EDTA). El material no unido se retiró mediante lavado con tampón A, seguido de un lavado con tampón C (fosfato de sodio 1, EDTA, pH 7,0). Se logró la elución de la fracción de producto mediante una elución de gradiente lineal con tampón E para bM32 al 0-100 % (fosfato de sodio, EDTA, NaCl, pH 7,0) en tampón C de BM32. La selección de fracciones que contienen producto para su agrupamiento se llevó a cabo mediante análisis de SDS-PAGE, mediante la evaluación densitométrica de la pureza de la fracción y mediante la intensidad de la banda de producto.
Las fracciones agrupadas de la etapa de captura se ajustaron a una conductividad de 115 mS/cm mediante la adición de cloruro de sodio 2,5 M y esta alimentación se cargó en una columna de Fenil Sepharose HP equilibrada con tampón D (fosfato de sodio, EDTA, NaCl, pH 7,0). Se retiró el material no unido mediante lavado con tampón D. La elución de la fracción de producto se logró mediante una elución de gradiente de tampón C al 40-100 % (fosfato de sodio, EDTA, pH 7,0) en tampón D. La selección de fracciones que contienen producto para su agrupamiento se llevó a cabo mediante análisis de SDS-PAGE, mediante la evaluación densitométrica de la pureza de la fracción y mediante la intensidad de la banda de producto.
Las fracciones agrupadas de la etapa intermedia se ajustaron hasta una conductividad de 80 mS/cm mediante la adición de cloruro de sodio 2,5 M y se cargó esta alimentación en una columna Toyopearl Butilo 650-S equilibrada con
una mezcla de tampón F (fosfato de sodio, EDTA, NaCI, pH 7,0). El material no unido se retiró mediante un lavado de gradiente con tampón F de BM32 al 80-0 % en tampón C (fosfato de sodio, EDTA, pH 7,0). La elución de la fracción se logró mediante una elución de gradiente de tampón G al 0-1 (fosfato de sodio, EDTA, isopropanol, pH 7,0) en tampón C. La selección de fracciones que contienen producto para su agolpamiento se llevó a cabo mediante análisis de SDS-PAGE, mediante la evaluación densitométrica de la pureza de la fracción y mediante la intensidad de la banda de producto.
Ejemplo 7: Producción de HBV_Phlp2_4xP3 (BM322), HBV_Phlp5_V2 (BM325) y HBV_Phlp6_4xP1 (BM326):
Para la expresión y producción de las moléculas recombinantes de acuerdo con la invención, a saber, HBV_Phlp2_4xP3 (BM322), HBV_Phlp5_V2 (BM325) y HBV_Phlp6_4xP1 (BM326), se aplicaron los mismos procedimientos a los descritos en el ejemplo 1, el ejemplo 2, el ejemplo 3, el ejemplo 4, el ejemplo 5 y el ejemplo 6 u otros comparables.
Ejemplo 8: Preparación de una formulación inyectable que consiste en una mezcla de HBV_PhlP1_4xP5 (BM321), HBV_PhlP2_4xP3 (BM322), HBV_PhlP5_V2 (BM325), y HBV_PhlP6_4xP1 (BM326)
Se disolvió cada una de las proteínas recombinantes purificadas en un tampón isotónico que contenía cloruro de sodio al 0,9 % y fosfato de sodio 2 mM y se añadió a cada solución de proteína una cantidad adecuada de hidróxido de aluminio. Se preparó una mezcla que contenía partes iguales de las cuatro suspensiones resultantes y se separó en alícuotas en condiciones estériles en viales sellados. La formulación inyectable obtenida mediante este procedimiento contenía 0,4 mg/ml de cada uno de HBV_PhlP1_4xP5, HBV_PhlP2_4xP3, HBV_PhlP5_V2 y HBV_PhlP6_4xP1. Ejemplo 9: Preparación de HBV_Betv1_4xPA marcado con his
Se sintetizó el gen que codifica las proteínas de fusión que consisten en PreS fusionada con péptido PA procedente de Bet v 1 dos veces en los extremos N y C-terminales (es decir, 4PA-PreS) por ATG:biosynthetics, Merzhausen, Alemania y se insertó en los sitios de Ndel/Xhol del vector pET-17b (Novagen, Alemania). Se confirmaron las secuencias de ADN mediante secuenciación automatizada de ambas hebras de ADN (Microsynth, Balgach, Suiza). La proteína de fusión se expresó en la cepa BL21 de E. coli (DE3; Stratagene, La Jolla, CA). Las células se cultivaron en medio Luria-Bertani que contenía 50 pg/ml de kanamicina hasta una DO de 0,6. La expresión de proteína se indujo añadiendo isopropil-p-D-tiogalactopiranósido hasta una concentración final de 1 mmol/l durante una noche a 37 °C. Las células se recogieron por centrifugación a 3500 rpm durante 10 minutos. El producto de proteína se detectó principalmente en la fracción de cuerpos de inclusión. Se solubilizó en GuHCl 6M, NaH2PO4 100 mM, Tris 10 mM, pH 8,0 durante una noche. El homogeneizado se centrifugó a 14.000 g durante 18 minutos. Los sobrenadantes se incubaron con 2 ml de una resina de Ni-NTA previamente equilibrada durante 4 horas (Qiagen, Hilden, Alemania) y posteriormente, se cargaron las suspensiones en una columna, se lavaron con 2 volúmenes de columna de tampón de lavado (8 mol/l de urea, 100 mmol/l de NaH2PO4 y 10 mmol/l de Tris-HCl [pH = 6,1]) y se eluyeron con el mismo tampón (pH 3,5). La proteína purificada se dializó contra agua.
La pureza de las proteínas recombinantes se analizó mediante SDS-PAGE teñida con Coomassie (12,5%) en condiciones reductoras.
Se confirmó la identidad de la proteína de fusión mediante transferencia por puntos usando anticuerpos monoclonales, específicos para los péptidos P2' (mAb2) y P4' (mAb12) derivados de Bet v 1 y anticuerpos de conejo específicos para PreS, así como las IgG preinmunes de conejo correspondientes. Se inmovilizaron sobre nitrocelulosa un pg de las proteínas de fusión de PreS, PreS y HSA (control) y se incubaron con sueros monoclonales, así como de conejo diluidos a 1:1000 a 4 °C. Los anticuerpos unidos se detectaron con anti-IgG de ratón procedente de conejo marcada con yodo 125 (mAb2, mAb12) o anti-IgG de conejo procedente de cabra marcada con 125I (anti-PreS procedente de conejo, preinmune de conejo) (Perkin-Elmer, Waltham, Massachusetts) diluido a 1:500 durante 2 horas y se visualizó mediante autorradiografía. Además, se recubrieron placas de ELISA (Maxisorp, Nunc, Dinamarca) con 2 pg de proteína de fusión PreS y PreS, diluidas en tampón carbonato 0,1 mol/l, pH 9,6, se lavaron con PBS que contenía un 0,05 % vol/vol de Tween 20 (PBST) 3 veces y se bloquearon durante 2 horas con BSA-PBST al 1 %. Posteriormente, se incubaron las placas con mAb2, mAb12, suero de conejo anti-PreS y anticuerpos de conejo anti-Bet v 1 en una dilución de 1:5000 (tampón de dilución: BSA al PBST al 0,5 % p/v) durante una noche a 4 °C. Tras lavar 5 veces, se detectaron los anticuerpos unidos con un anticuerpo de oveja anti-ratón marcado con HRP (para mAb2, mAb12) o anticuerpo de burro anti-conejo marcado con HRP (suero de conejo) (ambos de GE Healthcare, Uppsala, Suecia) y se reveló la reacción de color.
Ejemplo 10: Preparación de HBV_Betv1_2xPA2xPB (BM31) marcado con his
Se sintetizaron los genes que codifican la proteína de fusión que consiste en PreS fusionada dos veces con péptidos derivados de Bet v 1 en los extremos N y C-terminales (2xPA2xPB-PreS) por GenScript, Piscataway, NJ, Ee. UU., 2PAPB-Pres) y se insertaron en los sitios de Ndel/Xhol del vector pET-17b (Novagen, Alemania). Se confirmaron las secuencias de ADN mediante secuenciación automatizada de ambas hebras de ADN (Microsynth, Suiza).
Las proteínas de fusión de PreS recombinantes se expresaron en la cepa BL21 de E. coli (DE3; Stratagene, CA). Las células se cultivaron en medio Luria-Bertani que contenía 50 |jg/ml de kanamicina hasta una DO de 0,6. La expresión de proteína se indujo añadiendo isopropil-p-D-tiogalactopiranósido hasta una concentración final de 1 mmol/l durante una noche a 37 °C. Las células se recogieron por centrifugación a 3500 rpm durante 10 minutos. Las proteínas se detectaron principalmente en la fracción de cuerpos de inclusión. La proteína resultante se solubilizó en GuHCl 6M, NaH2PO4 100 mM, Tris 10 mM, pH 8,0 durante una noche. El homogeneizado se centrifugó a 14.000 g durante 18 minutos. Los sobrenadantes se incubaron con 2 ml de una resina de Ni-NTA previamente equilibrada durante 4 horas (Qiagen, Hilden, Alemania) y posteriormente, se cargaron las suspensiones en una columna, se lavaron con 2 volúmenes de columna de tampón de lavado (8 mol/l de urea, 100 mmol/l de NaH2PO4 y 10 mmol/l de Tris-HCl [pH = 6,1]) y se eluyeron con el mismo tampón (pH 3,5). La proteína se dializó contra NaH2PO4 10 mM.
La pureza de las proteínas recombinantes se analizó mediante SDS-PAGE teñida con Coomassie (12,5%) en condiciones reductoras. Se confirmó la identidad de las proteínas de fusión mediante transferencia por puntos usando anticuerpos monoclonales, específicos para los péptidos P2' (mAb2) y P4' (mAb12) derivados de Bet v 1 y anticuerpos de conejo específicos para PreS, así como las IgG preinmunes de conejo correspondientes. Se inmovilizaron sobre nitrocelulosa un jg de las proteínas de fusión de PreS, PreS y HSA (control) y se incubaron con sueros monoclonales, así como de conejo diluidos a 1:1000 a 4 °C. Los anticuerpos unidos se detectaron con anti-IgG de ratón procedente de conejo marcada con yodo 125 (mAb2, mAb12) o anti-IgG de conejo procedente de cabra marcada con 125I (anti-PreS procedente de conejo, preinmune de conejo) (Perkin-Elmer, Waltham, Massachusetts) diluido a 1:500 durante 2 horas y se visualizó mediante autorradiografía. Además, se recubrieron placas de ELISA (Maxisorp, Nunc, Rosklide, Dinamarca) con 2 jg de proteína de fusión PreS y PreS, diluidas en tampón carbonato 0,1 mol/l, pH 9,6, se lavaron con PBS que contenía un 0,05 % vol/vol de Tween 20 (PBST) 3 veces y se bloquearon durante 2 horas con BSA-PBST al 1 %. Posteriormente, se incubaron las placas con mAb2, mAb12, suero de conejo anti-PreS y anticuerpos de conejo anti-Bet v 1 en una dilución de 1:5000 (tampón de dilución: BSA al PBST al 0,5 % p/v) durante una noche a 4 °C. Tras lavar 5 veces, se detectaron los anticuerpos unidos con un anticuerpo de oveja anti-ratón marcado con HRP (para mAb2, mAb12) o anticuerpo de burro anti-conejo marcado con HRP (suero de conejo) (ambos de GE Healthcare, Uppsala, Suecia) y se reveló la reacción de color.
Ejemplo 11: Detección de reactividad con IgE de la proteína de fusión HBV_Phlpl_4xP5 (BM321)
Se evaluó la unión a IgE en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros de pacientes alérgicos al polen de gramíneas con proteínas transferidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión a IgE para HBV_Phlp1_4xP5 (BM321), como se muestra en la Fig. 4A.
Ejemplo 12: Detección de reactividad con IgE de la proteína de fusión HBV_Phlp2_4xP3 (BM322)
Se evaluó la unión a IgE en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros de pacientes alérgicos al polen de gramíneas con proteínas transferidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión a IgE para HBV_Phlp2_4xP3 (BM321), como se muestra en la Fig. 4B.
Ejemplo 13: Detección de reactividad con IgE de la proteína de fusión HBV_Phlp5_V2 (BM325)
Se evaluó la unión a IgE en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros de pacientes alérgicos al polen de gramíneas con proteínas transferidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión a IgE para HBV_Phlp5_V2 (BM325), como se muestra en la Fig. 4C.
Ejemplo 14: Detección de reactividad con IgE de la proteína de fusión HBV_Phlp6_4xP1 (BM326)
Se evaluó la unión a IgE en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros de pacientes alérgicos al polen de gramíneas con proteínas transferidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión a IgE para HBV_Phlp1_4xP1 (BM326), como se muestra en la Fig. 4D.
Ejemplo 15: Detección de reactividad con IgE de las proteínas de fusión HBV_etV1_4xPA y HBV_Betv1_2xPA2xPB (BM31)
Se evaluó la unión a IgE en comparación con el alérgeno completo mediante ensayo de transferencia de puntos de IgE. Se incubaron sueros de pacientes alérgicos al polen de gramíneas con proteínas transferidas por puntos y se detectó la IgE unida mediante anticuerpo anti-IgE humana marcado con 125I. No se detectó unión de IgE para ambas proteínas de fusión, como se muestra en la Fig. 5.
Ejemplo 16: Los anticuerpos de conejo anti-r89P5 bloquean la unión de las IgE del paciente a rPhl p 1 Para determinar la capacidad de la Ig de conejo inducida por péptido para inhibir la unión de los anticuerpos IgE de pacientes alérgicos a rPhl p 1, Se recubrieron placas de ELISA con 1 pg/ml de rPhl p 1, se lavaron y se bloquearon. Las placas se preincubaron con anticuerpo de conejo anti-péptido (HBV_Phlp1_4xP5, KLHP5) diluido a 1:100, un anticuerpo de conejo anti-rPhl p 1 y, para fines de control, con los sueros preinmunes correspondientes. Después del lavado, se incubaron las placas con suero humano procedentes de pacientes alérgicos a Phl p 1 (diluido a 1:3) y la IgE unida se detectó con anticuerpo de ratón anti-IgE humana (Pharmigen, 1:1000) y después con anticuerpo de oveja anti-IgG de ratón acoplado a POX (Amersham Bioscience) a 1:2000. Se calculó del siguiente modo el porcentaje de inhibición de la unión de IgE mediante preincubación con el antisuero anti-péptido del siguiente modo: 100-DOi/DOpx 100.
DOi y DOp representan las extinciones tras la preincubación con el suero inmune y preinmune de conejo, respectivamente. La tabla 1 muestra la capacidad de los anticuerpos anti-péptido Phl p 1 para inhibir la unión de las IgE de 13 pacientes alérgicos a rPhl p 1 completo. Los sueros anti-proteína de fusión bloquearon la unión de las IgE en la misma medida que el suero contra rPhl p 1 y KLHP5. La tabla 2 muestra la inhibición (en %) de los 13 pacientes.
Tabla 1: % de inhibición de la unión de las IgE de 13 pacientes a rPhl p 1 tras la incubación de anticuerpo de conejo
- - -

Ejemplo 17: Los anticuerpos de conejo anti-HBV_Phlp2_4xP3 bloquean la unión de las IgE del paciente a rPhl p 2
Para determinar la capacidad de la Ig de conejo inducida por péptido para inhibir la unión de los anticuerpos IgE de pacientes alérgicos a rPhl p 2, Se recubrieron placas de ELISA con 1 pg/ml de rPhl p 2, se lavaron y se bloquearon. Las placas se preincubaron con anticuerpo de conejo anti-péptido (HBV_Phlp2_4xP3, KLHP3) diluido a 1:100, un anticuerpo de conejo anti-rPhl p 2 y, para fines de control, con los sueros preinmunes correspondientes. Después del lavado, se incubaron las placas con suero humano procedentes de pacientes alérgicos a Phl p 2 (diluido a 1:3) y la IgE unida se detectó con anticuerpo de ratón anti-IgE humana (Pharmigen, 1:1000) y después con anticuerpo de oveja anti-IgG de ratón acoplado a POX (Amersham Bioscience) a 1:2000. Se calculó del siguiente modo el porcentaje de inhibición de la unión de IgE mediante preincubación con el antisuero anti-péptido del siguiente modo: 100-DOí/DOpx 100.
DOi y DOP representan las extinciones tras la preincubación con el suero inmune y preinmune de conejo, respectivamente. La tabla 2 muestra la capacidad de los anticuerpos anti-péptido Phl p 2 para inhibir la unión de las IgE de 19 pacientes alérgicos a rPhl p 2 completo. Los sueros anti-proteína de fusión bloquearon la unión de las IgE en la misma medida que el suero contra rPhl p 2 y KLHP3. La tabla 2 muestra la inhibición (en %) de los 19 pacientes.
Tabla 2: % de inhibición de la unión de las IgE de 19 pacientes a rPhl p 2 tras la incubación de anticuerpo de conejo
- - -


Ejemplo 18: Los anticuerpos de conejo anti-HBV_Phlp5_V2 bloquean la unión de las IgE del paciente a rPhl p 5
Para determinar la capacidad de la Ig de conejo inducida por péptido para inhibir la unión de los anticuerpos IgE de pacientes alérgicos a rPhl p 5, Se recubrieron placas de ELISA con 1 |jg/ml de rPhl p 5, se lavaron y se bloquearon. Las placas se preincubaron con anticuerpo de conejo anti-péptido (HBV_Phlp2_V2) diluido a 1:100, un anticuerpo de conejo anti-rPhl p 5 y, para fines de control, con los sueros preinmunes correspondientes. Después del lavado, se incubaron las placas con suero humano procedentes de pacientes alérgicos a Phl p 5 (diluido a 1:3) y la IgE unida se detectó con anticuerpo de ratón anti-IgE humana (Pharmigen, 1:1000) y después con anticuerpo de oveja anti-IgG de ratón acoplado a POX (Amersham Bioscience) a 1:2000. Se calculó del siguiente modo el porcentaje de inhibición de la unión de IgE mediante preincubación con el antisuero anti-péptido del siguiente modo: 100-DOí/DOp x 100.
DOi y DOp representan las extinciones tras la preincubación con el suero inmune y preinmune de conejo, respectivamente. La tabla 3 muestra la capacidad de los anticuerpos anti-péptido Phl p 5 para inhibir la unión de las IgE de 16 pacientes alérgicos a rPhl p 5 completo. Los sueros anti-proteína de fusión bloquearon la unión de las IgE en la misma medida que el suero contra rPhl p 5 y mejor que la mezcla de péptido KLH. La tabla 3 muestra la inhibición (en %) de los 16 pacientes.
Tabla 3: % de inhibición de la unión de las IgE de 13 pacientes a rPhl p 5 tras la incubación de anticuerpo de conejo - - -


Ejemplo 19: Los anticuerpos de conejo anti-HBV_Phlp6_4xP1 bloquean la unión de las IgE del paciente a rPhl p 6
Para determinar la capacidad de la Ig de conejo inducida por péptido para inhibir la unión de los anticuerpos IgE de pacientes alérgicos a rPhl p 6, Se recubrieron placas de ELISA con 1 |jg/ml de rPhl p 6, se lavaron y se bloquearon. Las placas se preincubaron con anticuerpo de conejo anti-péptido (HBV_Phlp6_4xP1, KLHP1) diluido, un anticuerpo de conejo anti-rPhl p 6 y, para fines de control, con los sueros preinmunes correspondientes. Después del lavado, se incubaron las placas con suero humano procedentes de pacientes alérgicos a Phl p 6 (diluido a 1:3) y la IgE unida se detectó con anticuerpo de ratón anti-IgE humana (Pharmigen, 1:1000) y después con anticuerpo de oveja anti-IgG de ratón acoplado a POX (Amersham Bioscience) a 1:2000. Se calculó del siguiente modo el porcentaje de inhibición de la unión de IgE mediante preincubación con el antisuero anti-péptido del siguiente modo: 100-DOi/DOp x 100. DOi y DOp representan las extinciones tras la preincubación con el suero inmune y preinmune de conejo, respectivamente. La tabla 4 muestra la capacidad de los anticuerpos anti-péptido Phl p 6 para inhibir la unión de las IgE de 21 pacientes alérgicos a rPhl p 6 completo. Los sueros anti-proteína de fusión bloquearon la unión de las IgE en la misma medida que el suero contra rPhl p 6 y KLHP1. La tabla 4 muestra la inhibición (en %) de los 21 pacientes.
Tabla 4: % de inhibición de la unión de las IgE de 21 pacientes a rPhl p 6 tras la incubación de anticuerpo de conejo ni-rPhl ni-HB Phl 4xP1 n i r ni-KLHP1

Ejemplo 20: Reactividad con IgE de las proteínas de fusión de PreS determinada mediante transferencia por puntos y ELISA
Se probó la reactividad con IgE de las proteínas de fusión recombinantes de rBet v 1, 4xPA-PreS y 2xPA2xPB-PreS mediante ensayos de transferencias por puntos no desnaturalizantes basados en RAST. Se transfirieron por puntos dos |jg de las proteínas purificadas y con fines de control, HSA sobre tiras de membrana de nitrocelulosa (Schleicher & Schuell, Dassel, Alemania).
Las tiras de nitrocelulosa se bloquearon en tampón A (Vrtala, J Clin Invest, 1997) y se incubaron con sueros de pacientes alérgicos al polen de abedul (n=50), sueros de personas no alérgicas (n=3) diluidos a 1:10, control de tampón y control positivo (antisuero de conejo anti-rBet v 1 diluido a 1:1000). Los anticuerpos IgE unidos se detectaron con anticuerpos anti-IgE humana marcados con 125I (BSM Diagnostica, Viena, Austria), anticuerpos de conejo unidos con un antisuero de cabra anti-conejo marcado con 125I (Perkin-Elmer) y se visualizaron mediante autorradiografía (Valenta et al., 1992). Adicionalmente, las placas de ELISA se recubrieron con rBet v 1 y las proteínas de fusión de PreS purificadas (5 jg/ml). Tras lavar y bloquear como se ha descrito anteriormente, las placas se incubaron con sueros de pacientes alérgicos al polen de abedul (n=21) y tres sueros de control no alérgico diluidos a 1:5. La IgE unida se detectó mediante anticuerpo de ratón anti-IgE humana purificado (BD Pharmigen) diluido a 1:1000 durante una noche y se visualizó con anticuerpo de oveja anti-IgG de ratón marcado con HRP (GE Healthcare) diluido a 1:2000. Después del lavado, se determinó la reacción de color como se ha descrito anteriormente.
Ejemplo 21: Regulación positiva inducida por alérgeno de CD203c de basófilos de pacientes alérgicos Se obtuvieron muestras de sangre heparinizadas de pacientes alérgicos al abedul tras otorgar consentimiento informado y se incubaron con concentraciones crecientes de rBet v 1,4PA-PreS, 2PAPB-PreS en el intervalo de 0,001 a 1 mg/ml, un anticuerpo monoclonal anti-IgE (Immunotech, Marsella, Francia) como control positivo o PBS (control negativo) durante 15 min (37 °C). La expresión de CD 203c se determinó como se ha descrito anteriormente.
Ejemplo 22: Respuestas linfoproliferativas e inducción de citocinas en PBMC de pacientes alérgicos al polen de abedul
Se aislaron PBMC de pacientes alérgicos al polen de abedul (n=6) mediante centrifugación de gradiente de densidad de Ficoll (Amersham Biosciences, Uppsala, Suecia). Posteriormente, se resuspendieron las PBMC en medio AIM V (Life Technologies, Grand Island, NY) hasta una concentración final de 2 x 105 células/pocillo y se estimularon con dosis decrecientes de antígeno (cantidades equimolares de 5 jg/pocillo de rBet v 1, PA, PB, PreS, 2PA-PreS, 2PB-PreS, 4PA-PreS, 2PAPB-PreS), con medio solo (control negativo) o con IL-2 (4 IE/pocillo) (control positivo). Después de 6 días, se midieron las respuestas proliferativas mediante incorporación de [3H] timidina y se expresaron como índices de estimulación (IE).
Además, la producción de 17 citocinas diferentes (es decir, IL-1p, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12, IL-13, IL-17, IFN-y, TNF-a, G-CSF, GM-CSF, MIP-1p, MCP-1) se midió tras 6 días de estimulación con el panel de 17 canales de citocinas humanas Bio-plex Pro (Bio-Rad Laboratories) de acuerdo con las instrucciones del fabricante. En resumen, se mezclaron los sobrenadantes no diluidos con anticuerpos monoclonales de ratón anticitocinas/quimiocinas acoplados a diferentes perlas como anticuerpos de captura (Bio-Rad). Se usó una curva patrón de 8 puntos para lograr una baja sensibilidad final. Después del lavado, se añadió anticuerpo de detección biotinilado anti-citocinas. La reacción se visualizó añadiendo ficoeritrina (PE) marcada con estreptavidina y tampón de ensayo. Las muestras se analizaron en un instrumento Luminex 100 (Biosource, Nivelles, Bélgica) y se recogieron los datos usando el programa informático Bio-Plex Manager 6.0. Todas las muestras se analizaron en un ciclo. Los resultados se muestran en la Fig. 10.
Ejemplo 23: Análisis de sueros de conejo inmunizados con rBet v 1 y proteínas de fusión de PreS respecto de su reconocimiento de alérgenos homólogos de rBet v E Bet v 1 y péptidos derivados de Bet v 1 mediante ELISA
Se recubrieron placas de ELISA (Maxisorp, Nunc) con 1 jg/ml de rBet v 1 o de alérgenos homólogos de aliso (rAln g 1), avellano (rCor a 1), manzano (rMal d 1) y adicionalmente con varios péptidos derivados de Bet v 1 a una concentración de 1 jg/ml durante una noche a 4 °C. Tras lavar y bloquear como se ha descrito anteriormente, los sueros de conejos inmunizados con rBet v 1 y las proteínas de fusión de PreS conjugadas a alumbre o CFA, se incubaron en diluciones seriadas de 1:2 en el intervalo de 1:500 a 1: 1280000 y en una concentración de 1:1000. La IgG de conejo unida se detectó con anticuerpos de burro anti-conejo marcados con HRP (GE Healthcare) y la reacción de color como se ha descrito anteriormente.
Ejemplo 24: Inhibición de la unión de IgE de pacientes alérgicos a rBet v 1
Se usó un ELISA de inhibición para estudiar la inhibición de la unión de IgE de pacientes alérgicos al polen de abedul a rBet v 1. Se recubrieron placas de ELISA con rBet v 1 a una concentración de 1 jg/ml a 4 °C durante una noche. Tras lavar y bloquear las placas, se preincubaron con sueros de conejo dirigidos contra la proteína de fusión de PreS,
2PAPB-PreS y suero de conejo anti-Bet v 1 en una dilución de 1:80 y 1:160 en comparación con suero preinmune de conejo durante una noche a 4 °C. Tras una etapa de lavado adicional, se añadieron sueros de pacientes alérgicos al polen de abedul diluidos a 1:5 durante una noche a 4 °C y se detectaron las IgE humanas unidas con un anticuerpo monoclonal de ratón anti-IgE humana conjugado a fosfatasa alcalina diluido a 1:1000 (BD Pharmigen). El porcentaje de inhibición de la unión de IgE a rBet v 1 tras la preincubación con antisueros de conejo contra 2PAPB-PreS y anticuerpos de conejo contra Bet v 1 se calculó del siguiente modo: porcentaje de inhibición = 100 - (DOi x 100/ DOp). DOp y DOi representan las extinciones tras la preincubación con IgG específica de conejo (DOi) o sueros preinmunes (DOp), respectivamente. (Fig. 12)
Ejemplo 25: Uso de una formulación de vacuna que comprende una mezcla de 4 proteínas de fusión hipoalergénicas para el tratamiento de la alergia al polen de gramíneas en individuos humanos alérgicos al polen de gramíneas
Se preparó una formulación inyectable de proteínas de fusión hipoalergénicas de SEQ ID NO: 14, SEQ ID NO: 15, SEQ iD NO: 16 y SEQ ID NO: 17 con hidróxido de aluminio, como se ha descrito en el ejemplo 8. Durante el transcurso de un estudio clínico, se administró la vacuna 3 veces por vía subcutánea a 69 sujetos humanos alérgicos al polen de gramíneas. (Fig. 17)
La vacunación con la formulación de vacuna ocasionó una robusta respuesta inmunitaria de IgG. La inducción de IgG específicas para el alérgeno tras la inyección s.c. de los 3 niveles de dosis diferentes de la vacuna y placebo se determinó mediante ELISA en los sueros recogidos de los participantes en el estudio antes y después del tratamiento con 3 inyecciones s.c. de la formulación de vacuna. (Fig. 14).
Para este fin, las placas de ELISA (Nunc Maxisorp, Roskilde, Dinamarca) se recubrieron con 5 pg/ml de los antígenos Phl p 1, Phl p 2, Phl p 5 y Phl p 6 o albúmina sérica humana (HSA) como control durante una noche a 4 °C. Tras lavar con PBS que contenía Tween 20 al 0,5 % (PT) y bloquear con BSA en PT al 2 % p/v, se incubaron las placas con sueros de pacientes diluidos de 1:10 a 1:100, suero de un individuo no atópico o solo tampón en triplicados durante una noche a 4 °C. Los anticuerpos IgE unidos se detectaron con anticuerpos anti-IgE humana acoplados a HRP diluidos en PT, BSA al 0,5 %. El revelado del color se llevó a cabo mediante la adición de soluciones de tinción ABTS (ácido 2,2'-Azino-bis(3-etilbenztiazolin-6-sulfónico) sal de diamonio; Sigma-Aldrich, St.Louis, Missouri, EE. UU.) (100 pl/pocillo). La densidad óptica se midió usando un lector de ELISA a 405 nm. Los resultados de las determinaciones de IgG se muestran en la figura 14.
La vacuna no provocó ninguna reactividad de linfocitos T relevante contra las proteínas de fusión hipoalergénicas presentes en la formulación de vacuna como se determinó mediante un ensayo de proliferación de linfocitos T in vitro (Fig. 15), demostrando de este modo la ausencia de reactividad con linfocitos T de las proteínas de fusión hipoalergénicas.
Se llevaron a cabo ensayos de proliferación de linfocitos T usando el siguiente procedimiento: Se aislaron células mononucleares de sangre periférica (PBMC) de muestras de sangre heparinizadas de los pacientes alérgicos al polen de gramíneas mediante centrifugación de gradiente de densidad de Ficoll (Amersham Pharmacia Biotech, Little Chalfont, R. U.). Después, se cultivaron las PBMC (2x105) por triplicado en placas de 96 pocillos (Nunclone; Nalge Nunc International, Roskilde, Dinamarca) en 200 pl de medio de cultivo Ultra asérico (BioWhittaker, Rockland, ME) complementado con L-glutamina 2mM (SIGMA, St. Louis, MO), b-mercaptoetanol 50 pM (SIGMA) y 0,1 mg de gentamicina por ml (SIGMA) a 37 °C y CO2 al 5 % en una atmósfera humidificada. Las células se estimularon con una mezcla que contenía 0,25 pg de cada componente polipeptídico de la vacuna y por comparación, una concentración equimolar de extracto de polen de gramíneas o para fines de control, con 4 U de interleucina-2 por pocillo (Boeringer Mannheim, Alemania) o solo medio. Tras 6 días de cultivo, se añadieron 0,5 pCi de [3H]timidina (Amersham Pharmacia Biotech) por pocillo y 16 h después se midió la radiactividad incorporada mediante conteo de centelleo líquido usando un contador de centelleo Microbeta (Wallac ADL, Friburgo, Alemania). Se calculó la media de cpm a partir de los triplicados y se calcularon los índices de estimulación (EI) como el cociente de las cpm obtenidas mediante la estimulación con antígeno o interleucina-2 y el control no estimulado. Los resultados de los ensayos de proliferación se muestran en la Fig. 15.
El tratamiento con la vacuna indujo anticuerpos IgG con la capacidad de modular la respuesta de linfocitos T específica de antígeno como se demostró por una respuesta proliferativa reducida tras la estimulación con alérgenos del polen de gramíneas en presencia de IgG inducidas por el tratamiento. (Fig. 16).
Para este fin, se llevaron a cabo ensayos de proliferación de linfocitos T con PBMC aisladas de participantes en el estudio tras el tratamiento, como se ha descrito anteriormente, salvo por que la estimulación se llevó a cabo con una mezcla de los 4 alérgenos del polen de gramíneas, Phl p 1, Phl p 2, Phl p5 y Phl p 6 (0,25 pg por alérgeno) junto con suero recogido del mismo participante antes y después del tratamiento. La configuración experimental y los resultados se muestran en la figura 1 6.
Se observó reducción de los síntomas nasales de alergia inducidos mediante exposición en una cámara de polen y reducción de la reactividad cutánea, determinada mediante una prueba de pinchazo en la piel titulada, en pacientes
que habían recibido 3 inyecciones que contenían o 20 jg o 40 |jg de cada uno de los 4 polipéptidos, mientras que no hubo reducción en estos parámetros tras el tratamiento con dosis de 10 jg de cada polipéptido, (véase la Fig. 19). Ejemplo 26: Selección de péptidos procedentes del alérgeno del ácaro del polvo doméstico Der p 2 y diseño de proteínas de fusión de PreS usando estos péptidos
Los 5 péptidos derivados de Der p 2 sin unión a IgE, Der p 2 Pep1 (SEQ ID NO: 96), Der p 2 Pep2 (SEQ ID NO: 97), Der p 2 Pep3 (SEQ ID NO: 98), Der p 2 Pep4 (SEQ ID NO: 99) y Der p 2 Pep5 (SEQ ID NO: 100), se exploraron respecto de
• sus propiedades de unión a IgE (ensayo de transferencia por puntos)
• su potencial para inducir reacciones de linfocitos T específicas para Der p 2 (ensayo de proliferación de linfocitos T) y
• su capacidad para inducir anticuerpos específicos para Der p 2 con la capacidad de bloquear las IgE del paciente humano para Der p 2. (ELISA de inhibición usando IgG de conejo anti-péptido)
Para este fin, se acopló químicamente cada uno de los péptidos a KLH. Se usó KLH y acoplamiento químico de los péptidos en este experimento de exploración debido a que es un sistema modelo fácil de usar y directo que permite la comparación inicial de los diferentes péptidos.
Se evaluó la unión a IgE de los péptidos derivados de Der p 2 en comparación con el alérgeno completo mediante el ensayo de transferencia por puntos de IgE. Se incubaron sueros de 26 pacientes alérgicos al ácaro del polvo doméstico con péptidos conjugados a KLH punteados y se detectó la IgE unida con anti-IgE humana marcada con 125I. No se detectó unión de IgE para ninguno de los 5 péptidos, como se muestran a continuación.
Para identificar péptidos que inducen una baja respuesta linfoproliferativa en PBMC de pacientes alérgicos al ácaro del polvo doméstico, se estimularon PBMC aisladas de 10 pacientes solamente con los 5 péptidos derivados de Der p 2, los péptidos conjugados a KLH y Der p 2 de tipo silvestre por comparación.
Se estimularon las PBMC de los 10 pacientes con Der p 2 de tipo silvestre y hubo una proliferación baja o nula tras la estimulación con Der p 2 Pep1, Der p 2 Pep2 y Der p 2 Pep4. Sin embargo, la estimulación con Der p 2 Pep3 y Der p 2 Pep5, dio como resultado una proliferación significativa de las PBMC en 4 de 10 y 3 de 10 casos, respectivamente, lo que indica que los péptidos 3 y 5 contienen epítopos para linfocitos T importantes.
Para identificar la capacidad de los péptidos para inducir IgG de bloqueo, se inmunizó a conejos con los 5 conjugados de KLH-péptido individuales. Posteriormente, Se investigó la capacidad de la Ig de conejo inducida por péptido para inhibir la unión de los anticuerpos IgE de pacientes alérgicos a rDer p 2 mediante ELISA. Se recubrieron placas de ELISA con 1 jg/ml de rDer p 2, se lavaron y se bloquearon. Las placas se preincubaron con anticuerpo de conejo antipéptido (KLH-P1, KLH-P2, KLH-P3, KLH-P4 y KLH-P5) diluido a 1:100, un anticuerpo de conejo anti-rDer p 2 y, para fines de control, con los sueros preinmunes correspondientes. Después del lavado, las placas se incubaron con sueros humano procedentes de pacientes alérgicos al ácaro del polvo doméstico sensibilizados a Der p 2 (diluido a 1:3) y la IgE unida se detectó con anticuerpo de ratón anti-IgE humana (Pharmigen, 1:1000) y después con anticuerpo de oveja anti-IgG de ratón acoplado a POX (Amersham Bioscience) a 1:2000. Se calculó del siguiente modo el porcentaje de inhibición de la unión de IgE mediante preincubación con el antisuero anti-péptido del siguiente modo: 100-DOí/DOpx 100.
Tabla 5: Capacidad de los anticuerpos anti-péptido Der p 2 para inhibir la unión de las IgE de 20 pacientes alérgicos a rDer p 2 completo. Los sueros anti-péptido KLH inducidos por los péptidos 2, 3 y 4 bloquearon la unión de IgE en la misma medida que los sueros contra Der p 2 de tipo silvestre. La tabla 5 muestra la inhibición (en %) de los 20 pacientes.

Claims (12)
1. Polipéptido capaz de generar respuestas de IgG contra al menos un alérgeno de tipo silvestre y capaz de inducir la producción de IL-10 e IFN-gamma, en donde la IgG generada de este modo se centra en los epítopos de IgE del alérgeno de tipo silvestre y puede bloquear la unión a dicho alérgeno de tipo silvestre, comprendiendo dicho polipéptido al menos 4 y hasta 6 fragmentos peptídicos hipoalergénicos que consisten en de 20 a 50 restos de aminoácido consecutivos de al menos un alérgeno de tipo silvestre, fusionados a los extremos N y C-terminales de una proteína PreS de hepatitis B (SEQ ID NO: 21) que actúa como transportador, en donde dichos fragmentos de péptido muestran una capacidad de unión a IgE nula o reducida en comparación con el alérgeno de tipo silvestre, son péptidos de unión a linfocitos B y muestran una reactividad con linfocitos T nula o sustancialmente nula, en donde la proteína PreS de hepatitis B comprende al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo N-terminal y al menos dos fragmentos peptídicos procedentes de al menos un alérgeno de tipo silvestre fusionado a su extremo C-terminal.
2. Polipéptido de acuerdo con la reivindicación 1, caracterizado por que el alérgeno de tipo silvestre se selecciona entre el grupo que consiste en los alérgenos principales del polen de abedul, en particular, Bet v 1, alérgenos principales del polen de la hierba timotea, preferentemente Phl p 1, Phl p 2, Phl p 5, Phl p 6 y Phl p 7, alérgenos principales de ácaros del polvo doméstico, preferentemente, Der p 1, Der p 2 y Der p 23, alérgeno principal de gato, Fel d 1 y Fel d 2, alérgenos principales de abeja, alérgenos principales de avispa, profilinas, especialmente Phl p 12, alérgenos del olivo, preferentemente Ole e 1, alérgenos de Parietaria judaica, preferentemente Par j 2, alérgenos de ambrosía, preferentemente Amb a 1, alérgenos del polen de artemisa, preferentemente Art v 1 y alérgeno del polen de cedro japonés, preferentemente Cry j 1 o Cry j 2.
3. Polipéptido de acuerdo con la reivindicación 1 o 2, caracterizado por que el fragmento de péptido se selecciona entre el grupo que consiste en los aminoácidos 151 a 177, 87 a 117, 1 a 30, 43 a 70 o 212 a 241 de Phl p 1, los aminoácidos 1 a 33, 8 a 39, 34 a 65 o 66 a 96 de Phl p 2, los aminoácidos 93 a 128, 98 a 128, 26 a 53, 26 a 58, 132 a 162, 217 a 246, 252 a 283 o 176 a 212 de Phl p 5, los aminoácidos 23 a 54, 56 a 90, 73 a 114 o 95 a 127 de Phl p 6 , los aminoácidos 1 a 34 o 35 a 70 de la cadena 1 de Fel d 1, los aminoácidos 1 a 34, 35 a 63 o 64 a 92 de la cadena 2 de Fel d 1, los aminoácidos 30 a 59, 50 a 79, 75 a 104, 30 a 74 o 60 a 104 de Bet v 1, los aminoácidos 1 a 30, 52 a 84 o 188 a 222 de Der p 1, los aminoácidos 1 a 33, 21 a 51,42 a 73, 62 a 103 o 98 a 129 de Der p 2, los aminoácidos 1 a 30, 20 a 50, 50 a 80, 90 a 125, 125 a 155 o 165 a 198 de Der p 7, los aminoácidos 1-35, 36-70, 71-110, 111-145, 140-170, 175-205, 210-250 o 250-284 de Der p 10, los aminoácidos 1 a 35, 35 a 72, 70 a 100 o 90 a 122 de Der p 21, los aminoácidos 1 a 32, 15 a 48 o 32 a 70, 32 a 60, 52 a 84, 32 a 70 (Cys->Ser) de Der p 23, los aminoácidos 19 a 58, 59 a 95, 91 a 120 o 121 a 157 de Alt a 1, los aminoácidos 31 a 6 0 , 45 a 80, 60 a 96 o 97 a 133 de Par j 2, los aminoácidos 1 a 40, 36 a 6 6 , 63 a 99, 86 a 120 o 107 a 145 de Ole e 1, los aminoácidos 25 a 58, 99 a 133, 154 a 183, 277 a 307, 334 a 363, 373 a 402, 544 a 573, 579 a 608, 58 a 99, 125 a 165, 183 a 224, 224 a 261, 252 a 289, 303 a 340, 416 a 457, 460 a 500 o 501 a 542 de Fel d 2, los aminoácidos 19 a 58, 52 a 91, 82 a 119, 106 a 144 o 139 a 180 de Can f 2, los aminoácidos 19 a 56, 51 a 90, 78 a 118, 106 a 145 o 135-174 de Can f 1, los aminoácidos 27 a 70, 70 a 100 o 92 a 132 de Art v 1, los aminoácidos 31 a 70, 80 a 120, 125 a 155, 160 a 200, 225 a 263, 264 a 300, 305 a 350 o 356 a 396 de Amb a 1, los aminoácidos 1 a 34, 35 a 74, 74 a 115, 125 a 165, 174 a 213, 241 a 280, 294 a 333, 361 a 400 o 401 a 438 de Alt a 6 , los aminoácidos 1 a 40, 41 a 80, 81 a 120, 121 a 160 de Alt a 2 o fragmentos o variaciones de secuencia de los mismos.
4. Polipéptido de acuerdo con una cualquiera de las reivindicaciones 1 a 3, caracterizado por que el polipéptido tiene una secuencia de aminoácidos seleccionada entre el grupo que consiste en SEQ ID NO: 14, s Eq ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17, SEQ ID NO: 18, SEQ ID NO: 19, SEQ ID NO: 20, SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151 y SEQ ID NO: 152.
5. Polipéptido de acuerdo con una cualquiera de las reivindicaciones 1 a 4 para su uso en el tratamiento o la prevención de una alergia en un ser humano o un animal.
6. Moléculas de ácido nucleico que codifican un polipéptido de acuerdo con una cualquiera de las reivindicaciones 1 a 4.
7. Vector que comprende una molécula de ácido nucleico de acuerdo con la reivindicación 6.
8. Hospedador que comprende una molécula de ácido nucleico de acuerdo con la reivindicación 6 o un vector de acuerdo con la reivindicación 7.
9. Formulación de vacuna que comprende al menos un polipéptido de acuerdo con una cualquiera de las reivindicaciones 1 a 4.
10. Formulación de vacuna de acuerdo con la reivindicación 9 para su uso en el tratamiento o la prevención de la alergia al polen de gramíneas que contiene una mezcla de polipéptidos hipoalergénicos procedentes de alérgenos del polen de gramíneas, caracterizado por que al menos uno de los polipéptidos se selecciona entre SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16 y SEQ ID NO: 17.
11. Formulación de vacuna de acuerdo con la reivindicación 9, para su uso en el tratamiento o la prevención de la alergia al polen de abedul que contiene al menos un polipéptido hipoalergénico seleccionado entre la SEQ ID NO: 18 y la SEQ ID NO: 19.
12. Formulación de vacuna de acuerdo con la reivindicación 9 para su uso en el tratamiento o la prevención de la alergia al ácaro del polvo doméstico que contiene al menos un polipéptido derivado de alérgenos de ácaros del polvo doméstico, caracterizado por que al menos uno de los polipéptidos se selecciona entre SEQ ID NO: 149, SEQ ID NO: 150, SEQ ID NO: 151 y SEQ ID NO: 152.
Applications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| EP11169365 | 2011-06-09 | ||
| PCT/EP2012/061040 WO2012168487A1 (en) | 2011-06-09 | 2012-06-11 | Peptide carrier fusion proteins as allergy vaccines |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| ES2720140T3 true ES2720140T3 (es) | 2019-07-18 |
Family
ID=46354215
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| ES12729450T Active ES2720140T3 (es) | 2011-06-09 | 2012-06-11 | Proteínas de fusión transportadoras de péptido como vacunas para alergias |
Country Status (17)
| Country | Link |
|---|---|
| US (2) | US9308251B2 (es) |
| EP (1) | EP2717910B1 (es) |
| JP (1) | JP6170912B2 (es) |
| CN (1) | CN103687617B (es) |
| AU (1) | AU2012266246B2 (es) |
| BR (1) | BR112013031505B1 (es) |
| CA (1) | CA2838217C (es) |
| DK (1) | DK2717910T3 (es) |
| ES (1) | ES2720140T3 (es) |
| HU (1) | HUE042761T2 (es) |
| MX (1) | MX343386B (es) |
| PL (1) | PL2717910T3 (es) |
| PT (1) | PT2717910T (es) |
| RU (1) | RU2630652C2 (es) |
| TR (1) | TR201905619T4 (es) |
| WO (1) | WO2012168487A1 (es) |
| ZA (1) | ZA201309399B (es) |
Families Citing this family (20)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| AT503530A1 (de) * | 2006-04-28 | 2007-11-15 | Biomay Ag | Polypeptid mit allergenen eigenschaften |
| RU2630652C2 (ru) * | 2011-06-09 | 2017-09-11 | Биомей Аг | Гибридные белки-носители пептидов в качестве вакцин от аллергии |
| CA2839832C (en) * | 2011-06-27 | 2018-08-28 | Anergis S.A. | Contiguous overlapping peptides for treatment of ragweed pollen allergy |
| BR112016011930A2 (pt) | 2013-12-23 | 2017-09-26 | Alk Abello As | combinações de peptídeos e seus usos em tratamento de alergia de ácaro de poeira |
| WO2015131053A1 (en) | 2014-02-28 | 2015-09-03 | Alk-Abelló A/S | Polypeptides derived from phl p and methods and uses thereof for immune response modulation |
| EP3153520A4 (en) * | 2014-06-04 | 2018-05-16 | Abbcn, S.L. | Peptide tags for marking proteins by fusion, and antibodies for the detection thereof |
| WO2017004561A1 (en) | 2015-07-01 | 2017-01-05 | Alk-Abelló A/S | Peptide combinations and uses thereof for treating grass allergy |
| EP3138579A1 (en) * | 2015-09-05 | 2017-03-08 | Biomay Ag | Fusion protein for use in the treatment of a hepatitis b virus infection |
| WO2017179025A1 (en) | 2016-04-15 | 2017-10-19 | Alk-Abelló A/S | Epitope polypeptides of ragweed pollen allergens |
| EP3295956A1 (en) | 2016-09-20 | 2018-03-21 | Biomay Ag | Polypeptide construct comprising fragments of allergens |
| EP3532084A4 (en) | 2016-10-31 | 2020-05-27 | Georgia State University Research Foundation, Inc. | VIRUS-LIKE HEPATITIS-B-VIRUS-PRÄ-S-PROTEIN PARTICLE |
| EP3569612A1 (en) * | 2018-05-18 | 2019-11-20 | Biomay Ag | Treatment and prevention of house dust mite allergies |
| RU2761431C9 (ru) | 2020-10-26 | 2022-04-18 | Федеральное государственное бюджетное учреждение "Государственный научный центр "Институт иммунологии" Федерального медико-биологического агентства России (ФГБУ "ГНЦ Институт иммунологии" ФМБА России) | Рекомбинантный полипептид на основе аллергена пыльцы березы и аллергена яблока в качестве вакцины от аллергии |
| EP4134674A1 (en) | 2021-08-13 | 2023-02-15 | Euroimmun Medizinische Labordiagnostika AG | Methods and reagents for detecting an antibody to a ses ipr-10 allergen |
| US20240382584A1 (en) | 2021-09-23 | 2024-11-21 | Viravaxx AG | Hbv vaccine inducing pres-specific neutralizing antibodies |
| EP4539877A1 (en) | 2022-06-17 | 2025-04-23 | Danmarks Tekniske Universitet | Allergy vaccines based on consensus allergens |
| CN116063442A (zh) * | 2022-09-28 | 2023-05-05 | 深圳市龙岗区耳鼻咽喉医院 | Der p2的3个B细胞表位肽及其应用 |
| JP2025537236A (ja) * | 2022-11-07 | 2025-11-14 | ウォルグ ファーマシューティカルズ(チョーチアン)カンパニー,リミテッド | アレルギー治療用ワクチン |
| CN116726198B (zh) * | 2023-03-13 | 2024-02-13 | 陕西师范大学 | 一种抗肿瘤药物及其在脑胶质瘤治疗中的应用 |
| WO2025194374A1 (zh) * | 2024-03-20 | 2025-09-25 | 深圳赫兹生命科学技术有限公司 | 猫过敏原多肽片段及应用 |
Family Cites Families (9)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6541011B2 (en) * | 1998-02-11 | 2003-04-01 | Maxygen, Inc. | Antigen library immunization |
| RU2285042C2 (ru) * | 2000-11-16 | 2006-10-10 | Альк-Абелло А/С | Новые мутантные аллергены |
| CA2490866A1 (en) | 2002-06-26 | 2004-01-08 | Advanced Bionutrition Corporation | Viruses and virus-like particles for multiple antigen and target display |
| ES2431963T3 (es) | 2002-07-05 | 2013-11-28 | Folia Biotech Inc. | Partícula viral adyuvante |
| AT503530A1 (de) | 2006-04-28 | 2007-11-15 | Biomay Ag | Polypeptid mit allergenen eigenschaften |
| AT503690A1 (de) * | 2006-06-09 | 2007-12-15 | Biomay Ag | Hypoallergene moleküle |
| US20090214593A1 (en) * | 2007-08-16 | 2009-08-27 | Tripep Ab | Immunogen platform |
| EP2295076A1 (en) | 2009-09-10 | 2011-03-16 | Biomay Ag | Hypoallergenic hybrid polypeptides for the treatment of allergy |
| RU2630652C2 (ru) * | 2011-06-09 | 2017-09-11 | Биомей Аг | Гибридные белки-носители пептидов в качестве вакцин от аллергии |
-
2012
- 2012-06-11 RU RU2013157115A patent/RU2630652C2/ru active
- 2012-06-11 TR TR2019/05619T patent/TR201905619T4/tr unknown
- 2012-06-11 JP JP2014514111A patent/JP6170912B2/ja active Active
- 2012-06-11 US US14/124,925 patent/US9308251B2/en active Active
- 2012-06-11 MX MX2013014439A patent/MX343386B/es active IP Right Grant
- 2012-06-11 PL PL12729450T patent/PL2717910T3/pl unknown
- 2012-06-11 WO PCT/EP2012/061040 patent/WO2012168487A1/en not_active Ceased
- 2012-06-11 CA CA2838217A patent/CA2838217C/en active Active
- 2012-06-11 PT PT12729450T patent/PT2717910T/pt unknown
- 2012-06-11 AU AU2012266246A patent/AU2012266246B2/en active Active
- 2012-06-11 ES ES12729450T patent/ES2720140T3/es active Active
- 2012-06-11 CN CN201280028169.7A patent/CN103687617B/zh active Active
- 2012-06-11 DK DK12729450.2T patent/DK2717910T3/en active
- 2012-06-11 EP EP12729450.2A patent/EP2717910B1/en active Active
- 2012-06-11 BR BR112013031505-9A patent/BR112013031505B1/pt active IP Right Grant
- 2012-06-11 HU HUE12729450A patent/HUE042761T2/hu unknown
-
2013
- 2013-12-12 ZA ZA2013/09399A patent/ZA201309399B/en unknown
-
2015
- 2015-05-01 US US14/701,848 patent/US9844591B2/en active Active
Also Published As
| Publication number | Publication date |
|---|---|
| DK2717910T3 (en) | 2019-04-15 |
| JP6170912B2 (ja) | 2017-07-26 |
| US20150335735A1 (en) | 2015-11-26 |
| TR201905619T4 (tr) | 2019-05-21 |
| CA2838217C (en) | 2022-11-01 |
| CN103687617A (zh) | 2014-03-26 |
| PL2717910T3 (pl) | 2019-07-31 |
| MX2013014439A (es) | 2014-07-24 |
| BR112013031505A2 (pt) | 2018-06-26 |
| CN103687617B (zh) | 2017-09-15 |
| RU2630652C2 (ru) | 2017-09-11 |
| MX343386B (es) | 2016-11-03 |
| AU2012266246A1 (en) | 2014-01-09 |
| HUE042761T2 (hu) | 2019-07-29 |
| JP2014518635A (ja) | 2014-08-07 |
| EP2717910B1 (en) | 2019-01-23 |
| PT2717910T (pt) | 2019-05-22 |
| US20140193450A1 (en) | 2014-07-10 |
| WO2012168487A1 (en) | 2012-12-13 |
| CA2838217A1 (en) | 2012-12-13 |
| AU2012266246B2 (en) | 2017-05-11 |
| ZA201309399B (en) | 2016-03-30 |
| US9308251B2 (en) | 2016-04-12 |
| US9844591B2 (en) | 2017-12-19 |
| EP2717910A1 (en) | 2014-04-16 |
| BR112013031505B1 (pt) | 2023-01-10 |
| RU2013157115A (ru) | 2015-07-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| ES2720140T3 (es) | Proteínas de fusión transportadoras de péptido como vacunas para alergias | |
| DK2035457T3 (en) | Vaccine carrier | |
| JP2015514126A (ja) | 非特異的アレルギー免疫療法において使用するための植物プロフィリンポリペプチド | |
| AU2013222053B2 (en) | Vaccine carrier | |
| AU2014352986A1 (en) | Pan pollen immunogens and methods and uses thereof for immune response modulation | |
| BRPI0712421B1 (pt) | Proteína de fusão hipoalergênica derivada de phl p 5 |