JP2009095237A - L−アミノ酸の製造法 - Google Patents
L−アミノ酸の製造法 Download PDFInfo
- Publication number
- JP2009095237A JP2009095237A JP2006025620A JP2006025620A JP2009095237A JP 2009095237 A JP2009095237 A JP 2009095237A JP 2006025620 A JP2006025620 A JP 2006025620A JP 2006025620 A JP2006025620 A JP 2006025620A JP 2009095237 A JP2009095237 A JP 2009095237A
- Authority
- JP
- Japan
- Prior art keywords
- gene
- amino acid
- promoter
- enzyme
- culture
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
Images


Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/70—Vectors or expression systems specially adapted for E. coli
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/38—Chemical stimulation of growth or activity by addition of chemical compounds which are not essential growth factors; Stimulation of growth by removal of a chemical compound
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/08—Lysine; Diaminopimelic acid; Threonine; Valine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/14—Glutamic acid; Glutamine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/22—Tryptophan; Tyrosine; Phenylalanine; 3,4-Dihydroxyphenylalanine
- C12P13/222—Phenylalanine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/22—Tryptophan; Tyrosine; Phenylalanine; 3,4-Dihydroxyphenylalanine
- C12P13/227—Tryptophan
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Life Sciences & Earth Sciences (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- General Chemical & Material Sciences (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Biomedical Technology (AREA)
- Virology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Medicinal Chemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Molecular Biology (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
【課題】従来よりもさらに改良された発酵法によるL−アミノ酸の製造法を提供する。
【解決手段】phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物を培地で培養して、L−アミノ酸を該培地に生成蓄積させ、該培地よりL−アミノ酸を採取する、L−アミノ酸の製造方法であって、前記培地中のリン濃度が、前記プロモーターによる発現が誘導される濃度である方法。
【選択図】なし
【解決手段】phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物を培地で培養して、L−アミノ酸を該培地に生成蓄積させ、該培地よりL−アミノ酸を採取する、L−アミノ酸の製造方法であって、前記培地中のリン濃度が、前記プロモーターによる発現が誘導される濃度である方法。
【選択図】なし
Description
本発明は微生物を用いたL−アミノ酸の製造法、特にL−リジン、L−スレオニン、L−トリプトファン等のL−アミノ酸の製造法に関する。L−リジン、L−スレオニン、L−トリプトファンは、動物飼料用の添加物、健康食品の成分、アミノ酸輸液等として、産業上有用なL−アミノ酸である。
L−アミノ酸は、ブレビバクテリウム属、コリネバクテリウム属、エシェリヒア属等に属する微生物を用いた発酵法により工業生産されている。これらの製造法においては、自然界から分離された菌株または該菌株の人工変異株、さらには、組換えDNA技術により塩基性L−アミノ酸生合成酵素の活性が増大するように改変された微生物などが用いられている。(特許文献1〜9)
また、一般的に細菌を用い、生産培養を行う際には、微生物の過剰な生育は基質の目的生産物への分配を減少させることになるために、培地中の栄養分の添加量を必要に応じて制限することが必要となる場合がある。制限する栄養分としては、要求アミノ酸やリンが挙げられる。培養液中のリン濃度を一定範囲内に制限し、細菌の過剰な生育を抑制するような培養方法としては、特許文献1に記載の方法が挙げられる。
EP0643135B
EP0733712B
EP1477565A
EP0796912A
EP0837134A
WO01/53459
EP1170376A
WO2005/010175
WO96/17930
US5,763,230
本発明は、従来よりもさらに改良された発酵法によるL−アミノ酸の製造法を提供することを課題とする。
本発明者らは、上記課題を解決するために鋭意検討を重ねた結果、腸内細菌科に属する微生物において、細胞内、あるいは培養液中のリンの濃度がある濃度以下に低下したときに、phoレギュロン(regulon)プロモーター配列の下流にL-アミノ酸生合成系の酵素をコードする遺伝子が接続された発現構築物を導入することにより、その細菌によるL−アミノ酸の生産が改善されることを見いだし、この知見に基づいて本発明を完成した。
本発明は下記のものを提供する。
1. phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物を培地で培養して、L−アミノ酸を該培地に生成蓄積させ、該培地よりL−アミノ酸を採取す
る、L−アミノ酸の製造方法であって、前記培地中のリン濃度が、前記プロモーターによる発現が誘導される濃度である方法。
2. 前記phoレギュロンプロモーターが、phoA, phoB, phoE, phoH, asr, argP, ugpB, pstS, psiE及びphnCから選択される遺伝子のプロモーターである1に記載の製造法。
3. 前記phoレギュロンプロモーターが、pho boxを有するプロモーターである、1又は2に記載の製造法。
4. 前記培地中のリン濃度が、100μM/L以下に制御される1〜3のいずれか一項に記載の製造法。
5. 前記L−アミノ酸生合成系酵素が、改変前の微生物においては、リンが枯渇した条件下で発現レベルが低下するものである、1〜4のいずれか一項に記載の製造法。
6. 前記DNA断片がマルチコピー型ベクターに搭載されている、又は、微生物の染色体DNA上に多コピー導入されている、1〜5のいずれか一項に記載の方法。
7. 前記腸内細菌科に属する微生物が、エシェリヒア属細菌、エンテロバクター属細菌、パントエア属細菌、クレブシエラ細菌、セラチア属細菌からなる群より選ばれる微生物である、1〜6のいずれか一項に記載の方法。
8. 前記L−アミノ酸がL−リジン、L−スレオニン、L−トリプトファン、L−グルタミン酸からなる群より選択される1種または2種以上のL−アミノ酸である1〜7のいずれか一項に記載の方法。
9. 前記L−アミノ酸がL−リジンであり、L−アミノ酸生合成系酵素がジヒドロジピコリン酸レダクターゼ、ジアミノピメリン酸脱炭酸酵素、ジアミノピメリン酸デヒドロゲナーゼ、ホスホエノールピルビン酸カルボキシラーゼ、アスパラギン酸アミノトランスフェラーゼ、ジアミノピメリン酸エピメラーゼ、アスパラギン酸セミアルデヒド脱水素酵素、テトラヒドロジピコリン酸スクシニラーゼ、及び、スクシニルジアミノピメリン酸デアシラーゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
10. 前記L−アミノ酸がL−スレオニンであり、L−アミノ酸生合成系酵素がアスパルトキナーゼIII、アスパラギン酸セミアルデヒドデヒドロゲナーゼ、thrオペロンにコードされるアスパルトキナーゼI、ホモセリンキナーゼ、及び、スレオニンシンターゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
11. 前記L−アミノ酸がL−グルタミン酸であり、L−アミノ酸生合成系酵素が、グルタミン酸デヒドロゲナーゼ、グルタミンシンテターゼ、グルタミン酸シンターゼ、イソクエン酸デヒドロゲナーゼ、アコニット酸ヒドラターゼ、クエン酸シンターゼ、ホスホエノールピルビン酸カルボキシラーゼ、ピルビン酸カルボキシラーゼ、ピルビン酸デヒドロゲナーゼ、ピルビン酸キナーゼ、ホスホエノールピルビン酸シンターゼ、6−ホスホグルコン酸デヒドラターゼ、及び、2−ケト−3−デオキシ−6−ホスホグルコン酸アルドラーゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
1. phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物を培地で培養して、L−アミノ酸を該培地に生成蓄積させ、該培地よりL−アミノ酸を採取す
る、L−アミノ酸の製造方法であって、前記培地中のリン濃度が、前記プロモーターによる発現が誘導される濃度である方法。
2. 前記phoレギュロンプロモーターが、phoA, phoB, phoE, phoH, asr, argP, ugpB, pstS, psiE及びphnCから選択される遺伝子のプロモーターである1に記載の製造法。
3. 前記phoレギュロンプロモーターが、pho boxを有するプロモーターである、1又は2に記載の製造法。
4. 前記培地中のリン濃度が、100μM/L以下に制御される1〜3のいずれか一項に記載の製造法。
5. 前記L−アミノ酸生合成系酵素が、改変前の微生物においては、リンが枯渇した条件下で発現レベルが低下するものである、1〜4のいずれか一項に記載の製造法。
6. 前記DNA断片がマルチコピー型ベクターに搭載されている、又は、微生物の染色体DNA上に多コピー導入されている、1〜5のいずれか一項に記載の方法。
7. 前記腸内細菌科に属する微生物が、エシェリヒア属細菌、エンテロバクター属細菌、パントエア属細菌、クレブシエラ細菌、セラチア属細菌からなる群より選ばれる微生物である、1〜6のいずれか一項に記載の方法。
8. 前記L−アミノ酸がL−リジン、L−スレオニン、L−トリプトファン、L−グルタミン酸からなる群より選択される1種または2種以上のL−アミノ酸である1〜7のいずれか一項に記載の方法。
9. 前記L−アミノ酸がL−リジンであり、L−アミノ酸生合成系酵素がジヒドロジピコリン酸レダクターゼ、ジアミノピメリン酸脱炭酸酵素、ジアミノピメリン酸デヒドロゲナーゼ、ホスホエノールピルビン酸カルボキシラーゼ、アスパラギン酸アミノトランスフェラーゼ、ジアミノピメリン酸エピメラーゼ、アスパラギン酸セミアルデヒド脱水素酵素、テトラヒドロジピコリン酸スクシニラーゼ、及び、スクシニルジアミノピメリン酸デアシラーゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
10. 前記L−アミノ酸がL−スレオニンであり、L−アミノ酸生合成系酵素がアスパルトキナーゼIII、アスパラギン酸セミアルデヒドデヒドロゲナーゼ、thrオペロンにコードされるアスパルトキナーゼI、ホモセリンキナーゼ、及び、スレオニンシンターゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
11. 前記L−アミノ酸がL−グルタミン酸であり、L−アミノ酸生合成系酵素が、グルタミン酸デヒドロゲナーゼ、グルタミンシンテターゼ、グルタミン酸シンターゼ、イソクエン酸デヒドロゲナーゼ、アコニット酸ヒドラターゼ、クエン酸シンターゼ、ホスホエノールピルビン酸カルボキシラーゼ、ピルビン酸カルボキシラーゼ、ピルビン酸デヒドロゲナーゼ、ピルビン酸キナーゼ、ホスホエノールピルビン酸シンターゼ、6−ホスホグルコン酸デヒドラターゼ、及び、2−ケト−3−デオキシ−6−ホスホグルコン酸アルドラーゼからなる群より選択される1種または2種以上の酵素である8に記載の方法。
本発明により、L−リジンなどのL−アミノ酸の生産性の高いエシェリヒア属細菌などの腸内細菌科に属する微生物が提供され、この微生物を用いることによりL−リジンなどのL−アミノ酸を高収率で得ることができる。
<1>本発明の微生物
本発明の微生物は、phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系の酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物である。ここで、L−アミノ酸生産能とは、本発明の微生物を培地中で培養したときに、培地中または菌体内にL−アミノ酸を生成し、蓄積する能力をいう。なお、本発明の微生物は複数のL−アミノ酸の生産能を有するものであってもよい。L−アミノ酸
の生産能を有する微生物としては、本来的にL−アミノ酸の生産能を有するものであってもよいが、下記<1−2>で説明するような微生物を、変異法や組換えDNA技術を利用して、L−アミノ酸の生産能を有するように改変したものであってもよい。
本発明の微生物は、phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系の酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物である。ここで、L−アミノ酸生産能とは、本発明の微生物を培地中で培養したときに、培地中または菌体内にL−アミノ酸を生成し、蓄積する能力をいう。なお、本発明の微生物は複数のL−アミノ酸の生産能を有するものであってもよい。L−アミノ酸
の生産能を有する微生物としては、本来的にL−アミノ酸の生産能を有するものであってもよいが、下記<1−2>で説明するような微生物を、変異法や組換えDNA技術を利用して、L−アミノ酸の生産能を有するように改変したものであってもよい。
L-アミノ酸の種類は特に制限されないが、L-リジン、L-オルニチン、L-アルギニン、L-ヒスチジン、L−シトルリン等の塩基性アミノ酸、L-イソロイシン、L-アラニン、L-バリン、L-ロイシン、L-グリシン等の脂肪族アミノ酸、L-スレオニン、L-セリン等のヒドロキシモノアミノカルボン酸であるアミノ酸、L-プロリン等の環式アミノ酸、L-フェニルアラニン、L-チロシン、L-トリプトファン等の芳香族アミノ酸、L-システイン、L-シスチン、L-メチオニン等の含硫アミノ酸、L-グルタミン酸、L-アスパラギン酸、L-グルタミン、L-アスパラギン等の酸性アミノ酸又はその酸アミドが挙げられるが、中でもL−リジン、L−トリプトファン、L-スレオニン、L−グルタミン酸が好ましい。本発明の微生物は2種類以上のアミノ酸の生産能を有するものであってもよい。
<1−1>本発明のDNA断片
本発明に用いるDNA断片は、phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系の酵素をコードする構造遺伝子が接続されたDNA断片である。
本発明に用いるDNA断片は、phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系の酵素をコードする構造遺伝子が接続されたDNA断片である。
phoレギュロンとは、細胞内のリン濃度が低下した際に発現が誘導される遺伝子群を意味し、通常には、phoB-phoRのヒスチジン−アスパラギン酸リン酸リレー制御により転写因子を活性化する経路を経て制御を受ける遺伝子群を意味する。
センサーキナーゼのPhoRが細胞内のリン濃度を感知し、ヒスチジン残基を自己リン酸化し、レスポンスレギュレーターであり転写因子であるPhoBタンパク質の特定のアスパラギン酸残基にリン酸を転移する。レスポンスレギュレーターであるPhoBタンパク質はこのリン酸化によって活性化され、多くの遺伝子の転写を制御する。
例えばphoレギュロンに属する遺伝子としては、pstSCAB, ugpBAEC, ugpQ, bap, phnSTUV, phnCDE, phoE, phoA等があげられる。
例えばphoレギュロンに属する遺伝子としては、pstSCAB, ugpBAEC, ugpQ, bap, phnSTUV, phnCDE, phoE, phoA等があげられる。
また、phoレギュロンプロモーターとは、phoレギュロンに属する遺伝子のプロモーターである。通常には、細胞内のリンの濃度が低下した際にphoB-phoRの二成分制御系によって転写調節を受ける遺伝子の上流に存在するプロモーターであり、PhoBが結合する領域を有する。具体的には、phoA, phoB, phoE, phoH, asr, argP, ugpB, pstS, psiE及びphnCから選択される遺伝子のプロモーターが挙げられる。これらの遺伝子の情報を以下に示す。
また、phoレギュロンプロモーター配列としては、pho BOXを有するプロモーターが好ましい。Pho BOXとは、phoBが結合する領域で微生物で高度に保存されている配列である。具体的には、開始コドンの約100bp〜10bp上流に保存されている配列で、好ましくはCTGTCATA(A/T)A(T/A)CTGT(C/A)A(C/T) (配列番号21)の配列を有し、-35領域の一部であるCTGTCATが最も高度に保存されている配列である。(Neidhardt, F. C. et al., Escherich
ia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D. C.,Chapter 87,Figure 6 )
ia coli and Salmonella Typhimurium, American Society for Microbiology, Washington D. C.,Chapter 87,Figure 6 )
プロモーターの配列には、細胞内のリン濃度が低下した際に発現を誘導する活性に影響しない変異が存在してもよい。例えば、phoA, phoB, phoE, phoH, asr, argP, ugpB, pstS, psiE及びphnCから選択される遺伝子のプロモーターの配列(配列番号1〜20)に対して、通常90%以上、好ましくは95%以上、より好ましくは97%の相同性を有し、細胞内のリン濃度が低下した際に発現を誘導する活性を有するものであり得る。
塩基配列の相同性(同一性)は、例えばKarlin and AltschulによるアルゴリズムBLAST(Pro. Natl. Acad. Sci. USA, 90, 5873(1993))やFASTA(Methods Enzymol., 183, 63 (1990))を用いて決定することができる。このアルゴリズムBLASTに基づいて、BLASTNやBLASTXとよばれるプログラムが開発されている (hwww.ncbi.nlm.nih.gov参照)。相同性は、通常には、このようなプログラムにより、その規定値を用いて算出される。
本発明において、「L−アミノ酸生合成系酵素」とは、代謝上L-アミノ酸生合成に関与する酵素であればいずれでもよいが、培養後半のリン濃度が低下している条件で、発現量が低下している酵素が好ましい。本発明において培養後半とは、主にアミノ酸生産期を意味し、菌体増殖期と区別される。本発明における「菌体増殖期」とは、培養開始から3時間、好ましくは6時間、特に好ましくは、10時間以内の、リン及び炭素源が主に菌体生育に使用されている時期、すなわち微生物が対数的に増殖している時期を意味し、本発明における「培養後半」とは、培養終了まで6時間、好ましくは10時間、特に好ましくは20時間前で、炭素源が主にL−アミノ酸生産に用いられている時期でを意味する。
培養後半のリン濃度が低下している条件で発現量が低下している酵素は、培養後半と菌体の増殖期(対数増殖期)である培養前半の酵素活性を比較することにより確認できる。また、DNAマクロアレイや、RT-PCR等によって培養後半と培養前半のmRNA量を比較することによっても確認できる。
L-アミノ酸生合成系酵素をコードする遺伝子は、L-アミノ酸生産に効果がある遺伝子である限り、1遺伝子のみを用いてもよいし、複数の遺伝子の組み合わせて用いてもよい。また、目的遺伝子は、エシェリヒア・コリの染色体上に存在する内因性の遺伝子であってもよいし、他微生物由来の外因性の遺伝子であってもよい。
以下、L-アミノ酸生合成系酵素をコードする遺伝子に関して詳細に説明する。
L−リジン生合成系酵素をコードする遺伝子としては、ジヒドロジピコリン酸合成酵素遺伝子(dapA)、アスパルトキナーゼ遺伝子(lysC)、ジヒドロジピコリン酸レダクターゼ遺伝子(dapB)、ジアミノピメリン酸脱炭酸酵素遺伝子(lysA(配列番号28))、ジアミノピメリン酸デヒドロゲナーゼ遺伝子(ddh)(以上、国際公開第96/40934号パンフレット)、ホスホエノールピルビン酸カルボキシラーゼ遺伝子(ppc) (特開昭60-87788号公報)、アスパラギン酸アミノトランスフェラーゼ遺伝子(aspC)(特公平6-102028号公報)、ジアミノピメリン酸エピメラーゼ遺伝子(dapF)(特開2003-135066号公報)、アスパラギン酸セミアルデヒド脱水素酵素遺伝子(asd)(国際公開第00/61723号パンフレット)等のジアミノピメリン酸経路の酵素の遺伝子、あるいはホモアコニット酸ヒドラターゼ遺伝子(特開2000-157276号公報)等のアミノアジピン酸経路の酵素等の遺伝子が挙げられる。また、テトラヒドロジピコリン酸スクシニラーゼ遺伝子(dapD)、スクシニルジアミノピメリン酸デアシラーゼ遺伝子(dapE)が挙げられる。これらのうち、dapB、lysA、ddh、pepC、aspC、dapF、asd、dapD、及び、dapEが好ましい。エシェリヒア・コリの全塩基配列は既に明らかにされており(Science, 277, 1453-1474 (1997))、この文献に報告されている目
的遺伝子、あるいはGenBankに登録されている遺伝子を元に遺伝子配列を取得できる。
的遺伝子、あるいはGenBankに登録されている遺伝子を元に遺伝子配列を取得できる。
L−グルタミン酸生合成系酵素をコードする遺伝子としては、L−グルタミン酸デヒドロゲナーゼ遺伝子(gdh)、グルタミンシンテターゼ遺伝子(glnA)、グルタミン酸シンターゼ遺伝子(gltBD)、イソクエン酸デヒドロゲナーゼ遺伝子(icd)、アコニット酸ヒドラターゼ遺伝子(acn)、クエン酸シンターゼ遺伝子(gltA)、ピルビン酸デヒドロゲナーゼ遺伝子(pdh)などが挙げられる(米国特許6,197,559号、6,331,419号明細書、欧州特許0999282号明細書)。また、ホスホエノールピルビン酸カルボキシラーゼ遺伝子(pepC)、ピルビン酸カルボキシラーゼ遺伝子(pc)、ピルビン酸キナーゼ遺伝子(pk)、ホスホエノールピルビン酸シンターゼ遺伝子(pps)、6−ホスホグルコン酸デヒドラターゼ遺伝子(edd)、及び、2−ケト−3−デオキシ−6−ホスホグルコン酸アルドラーゼ遺伝子(eda) (欧州特許1352966号)などが挙げられる。
L−スレオニン生合成系酵素をコードする遺伝子としては、アスパルトキナ−ゼIII遺伝子(lysC)、アスパラギン酸セミアルデヒドデヒドロゲナーゼ遺伝子(asd)、thrオペロンにコードされるアスパルトキナ−ゼI遺伝子(thrA)、ホモセリンキナーゼ遺伝子(thrB)、スレオニンシンターゼ遺伝子(thrC)が挙げられる。また、L−スレオニンはL−リジンと生合成系が共通しており、L-リジン生合成系酵素をコードする遺伝子を増幅してもよい。
L−スレオニン生合成系酵素は、最終産物のL−スレオニンによって酵素活性が抑制される。L−スレオニンによるフィードバック阻害を受けないようにL−スレオニン生合成系酵素をコードする遺伝子を改変した遺伝子を構造遺伝子として用いることが望ましい(国際公開第02/26993号パンフレット、Biotechnology Letters vol24,No.21,November 2002、国際公開第2005/049808号パンフレット参照)。
L−スレオニン生合成系酵素は、最終産物のL−スレオニンによって酵素活性が抑制される。L−スレオニンによるフィードバック阻害を受けないようにL−スレオニン生合成系酵素をコードする遺伝子を改変した遺伝子を構造遺伝子として用いることが望ましい(国際公開第02/26993号パンフレット、Biotechnology Letters vol24,No.21,November 2002、国際公開第2005/049808号パンフレット参照)。
L−トリプトファン、L−フェニルアラニン、L−チロシンは共に芳香族アミノ酸で生合成系が共通しており、芳香族アミノ酸の生合成系酵素をコードする遺伝子としては、デオキシアラビノ−ヘプツロン酸リン酸シンターゼ遺伝子(aroG)、3−デヒドロキネートシンターゼ遺伝子(aroB)、シキミ酸デヒドラターゼ遺伝子、シキミ酸キナーゼ遺伝子(aroL)、5−エノール酸ピルビンシキミ酸3−リン酸シンターゼ遺伝子(aroA)、コリスミ酸シンターゼ遺伝子(aroC)が挙げられる(欧州出願公開763127号明細書)。
L−トリプトファン生合成系酵素をコードする遺伝子としては、アントラニル酸合成酵素遺伝子(trpE)、ホスホグリセリン酸デヒドロゲナーゼ遺伝子(serA)もしくはトリプトファンシンターゼ遺伝子(trpAB)が挙げられ、ホスホグリセレートデヒドロゲナーゼ遺伝子(serA)を、フィードバック阻害を受けないように変異させ、得られた変異型遺伝子を用いるとより有効である(国際公開WO93/12235号パンフレット)。また、トリプトファンオペロンを含む組換えDNAを構造遺伝子として用いてもよい。具体的には、脱感作型アントラニル酸合成酵素をコードする遺伝子を含むトリプトファンオペロンが挙げられる(特開昭57-71397号公報、特開昭62-244382号公報、米国特許第4,371,614明細書)。また、トリプトファンオペロンのうち、トリプトファンシンターゼをコードする遺伝子(trpBA)の発現を強化することによっても、L−トリプトファン生産能を向上又は付与することができる。トリプトファンシンターゼは、α及びβサブユニットからなり、それぞれtrpA、trpBによってコードされている(米国特許第4,371,614明細書)。
L−フェニルアラニン、L−チロシン生合成系酵素をコードする遺伝子としては、プレフェン酸デヒドラターゼ遺伝子(tyrA;米国特許 4371614号明細書)、チロシンアミノトランスフェラーゼ遺伝子(tyrB;米国特許第5,091,314号明細書)があげられる。
L−アルギニン生合成系酵素をコードする遺伝子としては、N−アセチルグルタミン酸
シンターゼ遺伝子(argA)、N−アセチルグルタミルリン酸レダクターゼ遺伝子(argC)、オルニチンアセチルトランスフェラーゼ遺伝子(argJ)、N-アセチルグルタミン酸キナーゼ遺伝子(argB)、アセチルオルニチントランスアミナーゼ遺伝子(argD)、アセチルオルニチンデアセチラーゼ遺伝子(argE)、オルニチンカルバモイルトランスフェラーゼ遺伝子(argF)、アルギニノコハク酸シンターゼ遺伝子(argG)、アルギニノコハク酸リアーゼ遺伝子(argH)、及び、カルバモイルリン酸シンターゼ(carAB)から選ばれる1種又は2種以上が挙げられる(特開昭63-79597号)。N-アセチルグルタミン酸シンターゼ遺伝子(argA)は、野生型の15位〜19位に相当するアミノ酸配列が置換されたL−アルギニンによるフィードバック阻害が解除された変異型の遺伝子を用いるとより好適である(欧州出願公開1170361号明細書)。
シンターゼ遺伝子(argA)、N−アセチルグルタミルリン酸レダクターゼ遺伝子(argC)、オルニチンアセチルトランスフェラーゼ遺伝子(argJ)、N-アセチルグルタミン酸キナーゼ遺伝子(argB)、アセチルオルニチントランスアミナーゼ遺伝子(argD)、アセチルオルニチンデアセチラーゼ遺伝子(argE)、オルニチンカルバモイルトランスフェラーゼ遺伝子(argF)、アルギニノコハク酸シンターゼ遺伝子(argG)、アルギニノコハク酸リアーゼ遺伝子(argH)、及び、カルバモイルリン酸シンターゼ(carAB)から選ばれる1種又は2種以上が挙げられる(特開昭63-79597号)。N-アセチルグルタミン酸シンターゼ遺伝子(argA)は、野生型の15位〜19位に相当するアミノ酸配列が置換されたL−アルギニンによるフィードバック阻害が解除された変異型の遺伝子を用いるとより好適である(欧州出願公開1170361号明細書)。
L−ロイシン、L-バリン、L-イソロイシンは分岐鎖アミノ酸で生合成系が共通しており、分岐鎖アミノ酸生合成系の共通の酵素をコードする遺伝子としては、ピルビン酸デヒドロゲナーゼ遺伝子(aceE)が挙げられる(国際公開第03076635号パンフレット)。
L-バリン、L-イソロイシン生合成系酵素をコードする遺伝子としては、アセトヒドロキシ酸シンターゼ遺伝子(ilvGM)、分岐鎖アミノ酸アミノトランスフェラーゼ遺伝子(ilvE)、ジヒドロキシ酸デヒドラターゼ遺伝子(ilvD)、スレオニンデヒドラターゼ遺伝子(ilvA)が挙げられる。中でもilvGMEDAはオペロンを構成しており、オペロンとして用いてもよいし、個々の遺伝子を単独で使用してもよい。尚、ilvGMEDAオペロンは、L−バリン及び/又はL−イソロイシン及び/又はL−ロイシンによるオペロンの発現調節(アテニュエーション)を受けるので、生成するL−バリンによる発現抑制を解除するために、アテニュエーションに必要な領域が除去又は変異されていることが好ましい(米国特許5,998,178号明細書)。
また、L−イソロイシンは、L−スレオニンを前駆体として生成される。従って、L−イソロイシンの生産能を高めるには、L−スレオニンの供給を増加すること、すなわちL−スレオニンの生合成系を強化することが好ましい。従って、L−イソロイシン生合成系酵素をコードする遺伝子として、L−イソロイシン固有の生合成系酵素をコードする遺伝子とともに、前記のL−スレオニン生合成系を強化してもよい。
L−ロイシン生合成系酵素をコードする遺伝子としては、2−イソプロピルリンゴ酸シンターゼ遺伝子(leuA)、2−イソプロピルリンゴ酸イソメラーゼ遺伝子(leuD)、2−イソプロピルリンゴ酸デヒドロゲナーゼ遺伝子(leuB)、分岐鎖アミノ酸アミノトランスフェラーゼ遺伝子(ilvE;カナダ特許1341352号明細書)が挙げられる。2−イソプロピルリンゴ酸シンターゼは、L-ロイシンによってフィードバック阻害を受けるので、L−ロイシンによるイソプロピルリンゴ酸シンターゼのフィードバック阻害が脱感作されたleuAを用いることが好ましい(米国特許第6,403,342号明細書)。
L−ヒスチジン生合成系酵素をコードする遺伝子としては、ATP ホスホリボシルトランスフェラーゼ遺伝子(hisG)、ホスホリボシルAMP サイクロヒドロラーゼ遺伝子(hisI)、ホスホリボシル-ATP ピロホスホヒドラーゼ遺伝子(hisIE)、ホスホリボシルフォルミミノ−5−アミノイミダゾールカルボキシアミドリボタイドイソメラーゼ遺伝子(hisA)、アミドトランスフェラーゼ遺伝子(hisH)、ヒスチジノールホスフェートアミノトランスフェラーゼ遺伝子(hisC)、ヒスチジノールフォスファターゼ遺伝子(hisB)、ヒスチジノールデヒドロゲナーゼ遺伝子(hisD)等が挙げられる(米国特許4,388,405号明細書)。
L-システイン生合成系酵素をコードする遺伝子としては、ホスホグリセリン酸デヒドロゲナーゼ遺伝子(serA)、セリンアセチルトランスフェラーゼ遺伝子(cysE;国際公開第20
05/007841号パンフレット)、システインシンターゼ遺伝子(cysK)が挙げられる(国際公開第03/06666号パンフレット)。
05/007841号パンフレット)、システインシンターゼ遺伝子(cysK)が挙げられる(国際公開第03/06666号パンフレット)。
本発明のDNA断片は、以下のような方法で取得できる。
まず、phoレギュロンプロモーター配列と、L-アミノ酸生合成系遺伝子は別々にPCR等によってクローニングされる。PCRに用いるオリゴヌクレオチドは、公知のデータベースを参考にして設計される。またPCRのオリゴヌクレオチドのN末端側に制限酵素サイトを連結しておくと、簡単に2種のDNAを連結できる。
まず、phoレギュロンプロモーター配列と、L-アミノ酸生合成系遺伝子は別々にPCR等によってクローニングされる。PCRに用いるオリゴヌクレオチドは、公知のデータベースを参考にして設計される。またPCRのオリゴヌクレオチドのN末端側に制限酵素サイトを連結しておくと、簡単に2種のDNAを連結できる。
遺伝子のクローニングに使用されるプラスミドとしては、腸内細菌科において自律複製可能なものであればよく、具体的には、pBR322、pTWV228(宝バイオ社)、pMW119(ニッポンジーン社)、pUC19、pSTV29(宝バイオ社製)、RSF1010 (Gene vol.75 (2), p271-288, 1989)等が挙げられる。他にもファージDNAのベクターも利用できる。
次に本発現DNA断片を腸内細菌科に属する微生物に導入する方法について説明する。
例えば、以下のようにして、DNA断片を宿主に導入することができる。すなわち、DNA断片を、宿主微生物で機能するベクター、好ましくはマルチコピー型ベクターと連結して組み換えDNAを作製し、これで宿主を形質転換することにより導入できる。
目的遺伝子(phoレギュロンプロモーターの制御下に発現する遺伝子)を上記ベクターに連結して組み換えDNAを調製するには、目的遺伝子を含むDNA断片の末端に合うような制限酵素でベクターを切断する。連結は、T4 DNAリガーゼ等のリガーゼを用いて行うのが普通である。目的遺伝子は、それぞれ別個のベクターに搭載してもよく、同一のベクターに搭載してもよい。DNAの切断、連結、その他、染色体DNAの調製、PCR、プラスミドDNAの調製、形質転換、プライマーとして用いるオリゴヌクレオチドの設定等の方法は、当業者によく知られている通常の方法を採用することができる。これらの方法は、Sambrook, J., Fritsch, E. F., and Maniatis, T., "Molecular Cloning A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, (1989)等に記載されている。上記のようにして調製した組換えDNAを微生物に導入するには、十分な形質転換効率が得られる方法ならば、いかなる方法を用いてもよいが、例えば、エレクトロポレーション法(Canadian Journal of Microbiology, 43. 197(1997))が挙げられる。
目的遺伝子の発現は、通常の発現増大手段により増大させることが好ましい。例えば、細胞内でのコピー数を2以上、好ましくは3以上、さらに好ましくは4コピー以上に増大させることが挙げられる。
コピー数の増大は、目的遺伝子をマルチコピー型ベクターに搭載することにより達成できる。
また、目的遺伝子を微生物の染色体DNA上に多コピー導入することによっても達成できる。微生物の染色体DNA上に目的遺伝子を多コピーで導入するには、染色体DNA上に多コピー存在する配列を標的に利用して相同組換えにより行う。このような相同組換えを利用した遺伝子置換による部位特異的変異導入は既に確立しており、直鎖上DNAを用いる方法や温度感受性複製起点を含むプラスミドを用いる方法などがある(米国特許第6303383号、又は特開平05-007491号公報)。染色体DNA上に多コピー存在する配列としては、レペティティブDNA、転移因子の端部に存在するインバーティッド・リピートが利用できる。あるいは、特開平2-109985号公報に開示されているように、目的遺伝子をトランスポゾンに搭載してこれを転移させて染色体DNA上に多コピー導入することも可能で
ある。いずれの方法によっても形質転換株内の目的遺伝子のコピー数が上昇する結果、L−リジン生合成系の酵素活性が増大する。
ある。いずれの方法によっても形質転換株内の目的遺伝子のコピー数が上昇する結果、L−リジン生合成系の酵素活性が増大する。
目的遺伝子の発現増強は、上記の遺伝子増幅による以外に、phoレギュロンプロモーターの上流あるいは下流のL-アミノ酸生合成系遺伝子のプロモーター配列をより強力なものに置換することによっても達成される(特開平1-215280号公報参照)。たとえば、lacプロモーター、trpプロモーター、trcプロモーター、tacプロモーター、ラムダファージのPRプロモーター、PLプロモーター、tetプロモーター等が強力なプロモーターとして知られている。これらのプロモーターへの置換により、目的遺伝子の発現が強化されることによって酵素活性が増幅される。プロモーターの強度の評価法及び強力なプロモーターの例は、Goldsteinらの論文(Prokaryotic promoters in biotechnology. Biotechnol. Annu. Rev., 1995, 1, 105-128)等に記載されている。これらのプロモーターは、pho レギュロンプロモーターの上流あるいは下流に連結し、phoレギュロンプロモーターと強力なプロモーター共に発現が制御されていることが望ましい。
また、L-アミノ酸生合成系遺伝子の固有のプロモーターに国際公開WO00/18935に開示されているように、目的遺伝子のプロモーター領域に数塩基の塩基置換を導入し、より強力なものに改変することも可能である。さらに、リボソーム結合部位(RBS)と開始コドンとの間のスペーサ、特に開始コドンのすぐ上流の配列における数個のヌクレオチドの置換がmRNAの翻訳効率に非常に影響を及ぼすことが知られており、これらを改変することも可能である。目的遺伝子の発現調節領域は、プロモーター検索ベクターやGENETYX等の遺伝子解析ソフトを用いて決定することが出来る。発現調節配列の置換は、例えば、上述の温度感受性プラスミドを用いた遺伝子置換と同様にして行うことができる。
また、本発明のL-アミノ酸生合成系酵素をコードする遺伝子は、エシェリヒア・コリの種や菌株によって目的遺伝子の塩基配列に差異が存在することがあるため、本発明に用いる遺伝子は目的遺伝子がコードする活性を有する限り、1若しくは複数の位置での1若しくは数個のアミノ酸の置換、欠失、挿入又は付加等を含む配列を有するタンパク質をコードする変異体又は人為的な改変体であってもよい。ここで、「数個」とは、アミノ酸残基のタンパク質の立体構造における位置や種類によっても異なるが、具体的には2から20個、好ましくは、2から10個、より好ましくは2から5個である。上記の1若しくは数個のアミノ酸の置換、欠失、挿入、または付加は、酵素活性が維持される保存的変異である。保存的変異の代表的なものは、保存的置換であり、保存的置換とみなされる置換としては、AlaからSer又はThrへの置換、ArgからGln、His又はLysへの置換、AsnからGlu、Gln、Lys、His又はAspへの置換、AspからAsn、Glu又はGlnへの置換、CysからSer又はAlaへの置換、GlnからAsn、Glu、Lys、His、Asp又はArgへの置換、GluからAsn、Gln、Lys又はAspへの置換、GlyからProへの置換、HisからAsn、Lys、Gln、Arg又はTyrへの置換、IleからLeu、Met、Val又はPheへの置換、LeuからIle、Met、Val又はPheへの置換、LysからAsn、Glu、Gln、His又はArgへの置換、MetからIle、Leu、Val又はPheへの置換、PheからTrp、Tyr、Met、Ile又はLeuへの置換、SerからThr又はAlaへの置換、ThrからSer又はAlaへの置換、TrpからPhe又はTyrへの置換、TyrからHis、Phe又はTrpへの置換、及び、ValからMet、Ile又はLeuへの置換が挙げられる。
また、L-アミノ酸生合成系酵素をコードする遺伝子は、コードされる酵素の活性が維持される限り、その塩基配列または同塩基配列から調製され得るプローブとストリンジェントな条件下でハイブリダイズするDNAであってもよい。「ストリンジェントな条件」とは、いわゆる特異的なハイブリッドが形成され、非特異的なハイブリッドが形成されない条件をいう。一例を示せば、相同性が高いDNA同士、例えば70%以上の相同性を有するDNA同士がハイブリダイズし、それより相同性が低いDNA同士がハイブリダイズし
ない条件、あるいは通常のサザンハイブリダイゼーションの洗いの条件である60℃、1×SSC,0.1%SDS、好ましくは、0.1×SSC、0.1%SDSさらに好ましくは、68℃、0.1×SSC、0.1%SDSに相当する塩濃度、温度で、1回より好ましくは2〜3回洗浄する条件が挙げられる。プローブの長さは、ハイブリダイゼーションの条件により適宜選択されるが、通常には、100bp〜1Kbpである。
ない条件、あるいは通常のサザンハイブリダイゼーションの洗いの条件である60℃、1×SSC,0.1%SDS、好ましくは、0.1×SSC、0.1%SDSさらに好ましくは、68℃、0.1×SSC、0.1%SDSに相当する塩濃度、温度で、1回より好ましくは2〜3回洗浄する条件が挙げられる。プローブの長さは、ハイブリダイゼーションの条件により適宜選択されるが、通常には、100bp〜1Kbpである。
さらに、L-アミノ酸生合成系酵素をコードする遺伝子は、野生型の配列と比較して80%以上、好ましくは90%以上、より好ましくは95%以上、特に好ましくは97%以上の相同性を有するタンパク質をコードし、かつ、L-アミノ酸生合成系酵素の活性を有するタンパク質をコードする配列を用いることが出来る。相同性の算出方法は、プロモーター配列に関して説明したのと同様である。また、それぞれ導入する宿主により、遺伝子の縮重性が異なるので、それぞれ導入される宿主で使用しやすいコドンに置換したものでもよい。同様に目的遺伝子は、L-アミノ酸生合成系酵素の活性を有するタンパク質をコードする限り、N末端側、C末端側が延長したものあるいは削られているものでもよい。例えば延長・削除する長さは、アミノ酸残基で50以下、好ましくは20以下、より好ましくは10以下、特に好ましくは5以下である。より具体的には、アミノ酸配列のN末端側50アミノ酸から5アミノ酸、C末端側50アミノ酸から5アミノ酸延長・削除したものでもよい。
<1−2>親株
本発明の微生物の親株としては、エシェリヒア属細菌、パントエア属細菌を代表とする腸内細菌科に属する微生物を用いることができる。その他の腸内細菌科に属する微生物としては、エンテロバクター(Enterobacter)属、クレブシエラ(Klebsiella)属、セラチア(Serratia)属、エルビニア(Erwinia)属、サルモネラ(Salmonella)属、モルガネラ(Morganella)属などのγ−プロテオバクテリアに属する腸内細菌科に属する微生物が挙げられる。
本発明の微生物の親株としては、エシェリヒア属細菌、パントエア属細菌を代表とする腸内細菌科に属する微生物を用いることができる。その他の腸内細菌科に属する微生物としては、エンテロバクター(Enterobacter)属、クレブシエラ(Klebsiella)属、セラチア(Serratia)属、エルビニア(Erwinia)属、サルモネラ(Salmonella)属、モルガネラ(Morganella)属などのγ−プロテオバクテリアに属する腸内細菌科に属する微生物が挙げられる。
エシェリヒア属細菌としては、ナイトハルトらの著書((Backmann, B. J. 1996. Derivations and Genotypes of some mutant derivatives of Escherichia coli K-12, p. 2460-2488. Table 1. In F. D. Neidhardt (ed.), Escherichia coli and Salmonella Cellular and Molecular Biology/Second Edition, American Society for Microbiology Press, Washington, D.C.)に挙げられるもの、例えばエシェリヒア・コリ等が利用できる。エシェリヒア・コリの野生株としては、例えばK12株又はその誘導体、エシェリヒア・コリ MG1655株(ATCC No.47076)、及びW3110株(ATCC No.27325)等が挙げられる。これらを入手するには、例えばアメリカン・タイプ・カルチャー・コレクション(ATCC)より分譲を受けることができる(住所 P.O. Box 1549, Manassas, VA 20108, United States of America )。
また、エンテロバクター属細菌としては、エンテロバクター・アグロメランス(Enterobacter agglomerans)、エンテロバクター・アエロゲネス(Enterobacter aerogenes)等、パントエア属細菌としてはパントエア・アナナティス(Pantoea ananatis)が挙げられる。尚、近年、エンテロバクター・アグロメランスは、16S rRNAの塩基配列解析などにより、パントエア・アグロメランス(Pantoea agglomerans)又はパントエア・アナナティス(Pantoea ananatis)、パントエア・スチューアルティ(Pantoea stewartii)アグロメランス等に再分類されているものがある。本発明においては、腸内細菌科に分類されるものであれば、エンテロバクター属又はパントエア属のいずれに属するものであってもよい。パントエア・アナナティスを遺伝子工学的手法を用いて育種する場合には、パントエア・アナナティスAJ13355株(FERM BP−6614)、AJ13356株(FERM BP−6615)、AJ13601株(FERM BP−7207)及びそれらの誘導体を用いることができる。これらの株は、分離された当時はエンテロバクター・アグロメランスと同定され、エンテロバクター・アグロメランスとして寄託された
が、上記のとおり、16S rRNAの塩基配列解析などにより、パントエア・アナナティスに再分類されている。
が、上記のとおり、16S rRNAの塩基配列解析などにより、パントエア・アナナティスに再分類されている。
L−アミノ酸生産能を付与するには、栄養要求性変異株、アナログ耐性株又は代謝制御変異株の取得や、L−アミノ酸の生合成系酵素の発現が増大した組換え株の創製等、従来、エシェリヒア属細菌等の育種に採用されてきた方法を適用することができる(アミノ酸発酵、(株)学会出版センター、1986年5月30日初版発行、第77〜100頁参照)。ここで、L−アミノ酸生産菌の育種において、付与される栄養要求性、アナログ耐性、代謝制御変異等の性質は、単独でもよく、2種又は3種以上であってもよい。また、発現が増強されるL−アミノ酸生合成系酵素も、単独であっても、2種又は3種以上であってもよい。さらに、栄養要求性、アナログ耐性、代謝制御変異等の性質の付与と、生合成系酵素の活性の増大が組み合わされてもよい。
L−アミノ酸生産能を有する栄養要求性変異株、L−アミノ酸のアナログ耐性株、又は代謝制御変異株は、親株又は野生株を通常の変異処理、すなわちX線や紫外線の照射、またはN−メチル−N'−ニトロ−N−ニトロソグアニジン(NTG)、エチルメタンスルフォネート(EMS)等の変異剤処理などによって処理し、得られた変異株の中から、栄養要求性、アナログ耐性、又は代謝制御変異を示し、かつL−アミノ酸生産能を有するものを選択することによって得ることができる。
L−スレオニン生産菌
L−スレオニン生産菌の親株の例としては、E. coli TDH-6/pVIC40 (VKPM B-3996) (米国特許第5,175,107号、米国特許第5,705,371号)、E. coli 472T23/pYN7 (ATCC 98081) (米国特許第5,631,157号)、E. coli NRRL-21593 (米国特許第5,939,307号)、E. coli FERM
BP-3756 (米国特許第5,474,918号)、E. coli FERM BP-3519及びFERM BP-3520 (米国特許第5,376,538号)、E. coli MG442 (Gusyatiner et al., Genetika (in Russian), 14, 947-956 (1978))、E. coli VL643及びVL2055 (EP 1149911 A)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−スレオニン生産菌の親株の例としては、E. coli TDH-6/pVIC40 (VKPM B-3996) (米国特許第5,175,107号、米国特許第5,705,371号)、E. coli 472T23/pYN7 (ATCC 98081) (米国特許第5,631,157号)、E. coli NRRL-21593 (米国特許第5,939,307号)、E. coli FERM
BP-3756 (米国特許第5,474,918号)、E. coli FERM BP-3519及びFERM BP-3520 (米国特許第5,376,538号)、E. coli MG442 (Gusyatiner et al., Genetika (in Russian), 14, 947-956 (1978))、E. coli VL643及びVL2055 (EP 1149911 A)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
TDH-6株はthrC遺伝子を欠損し、スクロース資化性であり、また、そのilvA遺伝子がリーキー(leaky)変異を有する。この株はまた、rhtA遺伝子に、高濃度のスレオニンまたはホモセリンに対する耐性を付与する変異を有する。B-3996株は、RSF1010由来ベクターに、変異thrA遺伝子を含むthrA*BCオペロンを挿入したプラスミドpVIC40を保持する。この変異thrA遺伝子は、スレオニンによるフィードバック阻害が実質的に解除されたアスパルトキナーゼホモセリンデヒドロゲナーゼIをコードする。B-3996株は、1987年11月19日、オールユニオン・サイエンティフィック・センター・オブ・アンチビオティクス(Nagatinskaya Street 3-A, 117105 Moscow, Russia)に、受託番号RIA 1867で寄託されている。この株は、また、1987年4月7日、ルシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオルガニズムズ(VKPM) (1 Dorozhny proezd., 1 Moscow 117545, Russia) に、受託番号B-3996で寄託されている。
E. coli VKPM B-5318 (EP 0593792B)は、L-スレオニン生産菌の親株としても使用できる。B-5318株は、イソロイシンに関し原栄養性であり、プラスミドpVIC40中のスレオニンオペロンの制御領域が、温度感受性ラムダファージC1リプレッサー及びPRプロモーターにより置換されている。VKPM B-5318は、1990年5月3日、ルシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオルガニズムズ(VKPM)に、受託番号VKPM B-5318で寄託されている。
好ましくは、本発明の細菌は、さらに、下記の遺伝子の1種以上の発現が増大するように改変される。
スレオニンによるフィードバック阻害に耐性のアスパルトキナーゼホモセリンデヒドロゲナーゼIをコードする変異thrA遺伝子
ホモセリンキナーゼをコードするthrB遺伝子
スレオニンシンターゼをコードするthrC遺伝子
推定トランスメンブランタンパク質をコードするrhtA遺伝子
アスパラギン酸−β−セミアルデヒドデヒドロゲナーゼをコードするasd遺伝子
アスパラギン酸アミノトランスフェラーゼ(アスパラギン酸トランスアミナーゼ)をコードするaspC遺伝子
スレオニンによるフィードバック阻害に耐性のアスパルトキナーゼホモセリンデヒドロゲナーゼIをコードする変異thrA遺伝子
ホモセリンキナーゼをコードするthrB遺伝子
スレオニンシンターゼをコードするthrC遺伝子
推定トランスメンブランタンパク質をコードするrhtA遺伝子
アスパラギン酸−β−セミアルデヒドデヒドロゲナーゼをコードするasd遺伝子
アスパラギン酸アミノトランスフェラーゼ(アスパラギン酸トランスアミナーゼ)をコードするaspC遺伝子
Escherichia coliのアスパルトキナーゼホモセリンデヒドロゲナーゼIをコードするthrA遺伝子が知られている(ヌクレオチド番号337〜2799, GenBank accession NC_000913.2,
gi: 49175990)。thrA遺伝子は、E. coli K-12の染色体において、thrL遺伝子とthrB遺伝子との間に位置する。Escherichia coliのホモセリンキナーゼをコードするthrB遺伝子が知られている(ヌクレオチド番号2801〜3733, GenBank accession NC_000913.2, gi: 49175990)。thrB遺伝子は、E. coli K-12の染色体において、thrA遺伝子とthrC遺伝子との間に位置する。Escherichia coliのスレオニンシンターゼをコードするthrC遺伝子が知られている(ヌクレオチド番号3734〜5020, GenBank accession NC_000913.2, gi: 49175990)。thrC遺伝子は、E. coli K-12の染色体において、thrB遺伝子とyaaXオープンリーディングフレームとの間に位置する。三つの遺伝子は、全て、単一のスレオニンオペロンとして機能する。スレオニンオペロンの発現を増大させるには、転写に影響するアテニュエーター領域を、好ましくは、オペロンから除去する(WO2005/049808, WO2003/097839)。
gi: 49175990)。thrA遺伝子は、E. coli K-12の染色体において、thrL遺伝子とthrB遺伝子との間に位置する。Escherichia coliのホモセリンキナーゼをコードするthrB遺伝子が知られている(ヌクレオチド番号2801〜3733, GenBank accession NC_000913.2, gi: 49175990)。thrB遺伝子は、E. coli K-12の染色体において、thrA遺伝子とthrC遺伝子との間に位置する。Escherichia coliのスレオニンシンターゼをコードするthrC遺伝子が知られている(ヌクレオチド番号3734〜5020, GenBank accession NC_000913.2, gi: 49175990)。thrC遺伝子は、E. coli K-12の染色体において、thrB遺伝子とyaaXオープンリーディングフレームとの間に位置する。三つの遺伝子は、全て、単一のスレオニンオペロンとして機能する。スレオニンオペロンの発現を増大させるには、転写に影響するアテニュエーター領域を、好ましくは、オペロンから除去する(WO2005/049808, WO2003/097839)。
スレオニンによるフィードバック阻害に耐性のアスパルトキナーゼホモセリンデヒドロゲナーゼIをコードする変異thrA遺伝子、ならびに、thrB遺伝子及びthrC遺伝子は、スレオニン生産E. coli VKPM B-3996株に存在する周知のプラスミドpVIC40から一つのオペロンとして取得できる。プラスミドpVIC40の詳細は、米国特許第5,705,371号に記載されている。
rhtA遺伝子は、グルタミン輸送系の要素をコードするglnHPQ オペロンに近いE. coli染色体の18分に存在する。rhtA遺伝子は、ORF1 (ybiF遺伝子, ヌクレオチド番号764〜1651,
GenBank accession number AAA218541, gi:440181)と同一であり、pexB遺伝子とompX遺伝子との間に位置する。ORF1によりコードされるタンパク質を発現するユニットは、rhtA遺伝子と呼ばれている(rht: ホモセリン及びスレオニンに耐性)。また、rhtA23変異が、ATG開始コドンに対して-1位のG→A置換であることが判明している(ABSTRACTS of the 17th
International Congress of Biochemistry and Molecular Biology in conjugation with Annual Meeting of the American Society for Biochemistry and Molecular Biology,
San Francisco, California August 24-29, 1997, abstract No. 457, EP 1013765 A)。
GenBank accession number AAA218541, gi:440181)と同一であり、pexB遺伝子とompX遺伝子との間に位置する。ORF1によりコードされるタンパク質を発現するユニットは、rhtA遺伝子と呼ばれている(rht: ホモセリン及びスレオニンに耐性)。また、rhtA23変異が、ATG開始コドンに対して-1位のG→A置換であることが判明している(ABSTRACTS of the 17th
International Congress of Biochemistry and Molecular Biology in conjugation with Annual Meeting of the American Society for Biochemistry and Molecular Biology,
San Francisco, California August 24-29, 1997, abstract No. 457, EP 1013765 A)。
E. coliのasd遺伝子は既に知られており(ヌクレオチド番号3572511〜3571408, GenBank
accession NC_000913.1, gi:16131307)、その遺伝子の塩基配列に基づいて作製されたプライマーを用いるPCRにより得ることができる(White, T.J. et al., Trends Genet., 5, 185 (1989)参照)。他の微生物のasd遺伝子も同様に得ることができる。
accession NC_000913.1, gi:16131307)、その遺伝子の塩基配列に基づいて作製されたプライマーを用いるPCRにより得ることができる(White, T.J. et al., Trends Genet., 5, 185 (1989)参照)。他の微生物のasd遺伝子も同様に得ることができる。
また、E. coliのaspC遺伝子も既に知られており(ヌクレオチド番号983742〜984932, GenBank accession NC_000913.1, gi:16128895)、PCRにより得ることができる。他の微生物のaspC遺伝子も同様に得ることができる。
L−リジン生産菌
エシェリヒア属に属するL−リジン生産菌の例としては、L−リジンアナログに耐性を有する変異株が挙げられる。L−リジンアナログはエシェリヒア属に属する細菌の生育を
阻害するが、この阻害は、L−リジンが培地に共存するときには完全にまたは部分的に解除される。L−リジンアナログの例としては、オキサリジン、リジンヒドロキサメート、S−(2−アミノエチル)−L−システイン(AEC)、γ−メチルリジン、α−クロロカプロラクタムなどが挙げられるが、これらに限定されない。これらのリジンアナログに対して耐性を有する変異株は、エシェリヒア属に属する細菌を通常の人工変異処理に付すことによって得ることができる。L−リジンの生産に有用な細菌株の具体例としては、Escherichia coli AJ11442 (FERM BP-1543, NRRL B-12185; 米国特許第4,346,170号参照)及びEscherichia coli VL611が挙げられる。これらの微生物では、アスパルトキナーゼのL−リジンによるフィードバック阻害が解除されている。
エシェリヒア属に属するL−リジン生産菌の例としては、L−リジンアナログに耐性を有する変異株が挙げられる。L−リジンアナログはエシェリヒア属に属する細菌の生育を
阻害するが、この阻害は、L−リジンが培地に共存するときには完全にまたは部分的に解除される。L−リジンアナログの例としては、オキサリジン、リジンヒドロキサメート、S−(2−アミノエチル)−L−システイン(AEC)、γ−メチルリジン、α−クロロカプロラクタムなどが挙げられるが、これらに限定されない。これらのリジンアナログに対して耐性を有する変異株は、エシェリヒア属に属する細菌を通常の人工変異処理に付すことによって得ることができる。L−リジンの生産に有用な細菌株の具体例としては、Escherichia coli AJ11442 (FERM BP-1543, NRRL B-12185; 米国特許第4,346,170号参照)及びEscherichia coli VL611が挙げられる。これらの微生物では、アスパルトキナーゼのL−リジンによるフィードバック阻害が解除されている。
WC196株が、Escherichia coliのL−リジン生産菌として使用できる。この菌株は、Escherichia coli K-12に由来するW3110株にAEC耐性を付与することにより育種された。同株は、Escherichia coli AJ13069株と命名され、1994年12月6日、工業技術院生命工学工業技術研究所(現 独立行政法人 産業技術総合研究所 特許生物寄託センター、〒305-8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に受託番号FERM P-14690として寄託され、1995年9月29日にブダペスト条約に基づく国際寄託に移管され、受託番号FERM BP-5252が付与されている(米国特許第5,827,698号)。
L-リジン産生菌の親株の例としては、L−リジン生合成系酵素をコードする遺伝子の1種以上の発現が増大している株も挙げられる。L−リジン生合成系酵素をコードする遺伝子の例としては、ジヒドロジピコリン酸シンターゼ遺伝子(dapA)、アスパルトキナーゼ遺伝子(lysC)、ジヒドロジピコリン酸レダクターゼ遺伝子(dapB)、ジアミノピメリン酸デカルボキシラーゼ遺伝子(lysA)、ジアミノピメリン酸デヒドロゲナーゼ遺伝子(ddh) (米国特許第6,040,160号)、ホスホエノールピルビン酸カルボキシラーゼ遺伝子(ppc)、アスパラギン酸セミアルデヒドデヒドロゲナーゼ遺伝子(asd)及びアスルターゼ遺伝子(aspA) (EP 1253195 A)が挙げられるが、これらに限定されない。また、親株は、エネルギー効率に関与する遺伝子(cyo) (EP 1170376 A)、ニコチンアミドヌクレオチドトランスヒドロゲナーゼをコードする遺伝子(pntAB) (米国特許第5,830,716号)、ybjE遺伝子(WO2005/073390)、または、これらの組み合わせの発現量が増大していてもよい。
L−リジン産生菌の親株の例としては、L−リジンの生合成経路から分岐してL−リジン以外の化合物を生成する反応を触媒する酵素の活性が低下または欠損している株も挙げられる。L−リジンの生合成経路から分岐してL−リジン以外の化合物を生成する反応を触媒する酵素の例としては、ホモセリンデヒドロゲナーゼ、リジンデカルボキシラーゼ(米国特許第5,827,698号)、及び、リンゴ酸酵素(WO2005/010175)が挙げられる。
L−システイン生産菌
L−システイン生産菌の親株の例としては、フィードバック阻害耐性のセリンアセチルトランスフェラーゼをコードする異なるcysEアレルで形質転換されたE. coli JM15(米国特許第6,218,168号、ロシア特許出願第2003121601号)、細胞に毒性の物質を排出するのに適したタンパク質をコードする過剰発現遺伝子を有するE. coli W3110 (米国特許第5,972,663号)、システインデスルフォヒドラーゼ活性が低下したE. coli株 (JP11155571A2)、cysB遺伝子によりコードされる正のシステインレギュレーターの活性が上昇したE. coli W3110 (WO0127307A1)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−システイン生産菌の親株の例としては、フィードバック阻害耐性のセリンアセチルトランスフェラーゼをコードする異なるcysEアレルで形質転換されたE. coli JM15(米国特許第6,218,168号、ロシア特許出願第2003121601号)、細胞に毒性の物質を排出するのに適したタンパク質をコードする過剰発現遺伝子を有するE. coli W3110 (米国特許第5,972,663号)、システインデスルフォヒドラーゼ活性が低下したE. coli株 (JP11155571A2)、cysB遺伝子によりコードされる正のシステインレギュレーターの活性が上昇したE. coli W3110 (WO0127307A1)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−ロイシン生産菌
L−ロイシン生産菌の親株の例としては、ロイシン耐性のE. coil株 (例えば、57株 (VKPM B-7386, 米国特許第6,124,121号)またはβ−2−チエニルアラニン、3−ヒドロキシロイシン、4−アザロイシン、5,5,5-トリフルオロロイシンなどのロイシンアナログ耐性
のE.coli株(JP 62-34397 B及びJP 8-70879 A)、WO96/06926に記載された遺伝子工学的方法で得られたE. coli株、E. coli H-9068 (JP 8-70879 A)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−ロイシン生産菌の親株の例としては、ロイシン耐性のE. coil株 (例えば、57株 (VKPM B-7386, 米国特許第6,124,121号)またはβ−2−チエニルアラニン、3−ヒドロキシロイシン、4−アザロイシン、5,5,5-トリフルオロロイシンなどのロイシンアナログ耐性
のE.coli株(JP 62-34397 B及びJP 8-70879 A)、WO96/06926に記載された遺伝子工学的方法で得られたE. coli株、E. coli H-9068 (JP 8-70879 A)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
本発明の細菌は、L-ロイシン生合成に関与する遺伝子の1種以上の発現が増大されることにより改良されていてもよい。このような遺伝子の例としては、L-ロイシンによるフィードバック阻害を受けないイソプロピルリンゴ酸シンターゼをコードする変異leuA遺伝子(米国特許第6,403,342号)に代表されるleuABCDオペロンの遺伝子が挙げられる。さらに、本発明の細菌は、細菌の細胞からL-アミノ酸を排出するタンパク質をコードする遺伝子の1種以上の発現が増大されることにより改良されていてもよい。このような遺伝子の例としては、b2682遺伝子及びb2683遺伝子(ygaZH遺伝子) (EP 1239041 A2)が挙げられる。
L−ヒスチジン生産菌
L−ヒスチジン生産菌の親株の例としては、E. coli 24株 (VKPM B-5945, RU2003677)、E. coli 80株 (VKPM B-7270, RU2119536)、E. coli NRRL B-12116 - B12121 (米国特許第4,388,405号)、E. coli H-9342 (FERM BP-6675)及びH-9343 (FERM BP-6676) (米国特許第6,344,347号)、E. coli H-9341 (FERM BP-6674) (EP1085087)、E. coli AI80/pFM201 (米国特許第6,258,554号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−ヒスチジン生産菌の親株の例としては、E. coli 24株 (VKPM B-5945, RU2003677)、E. coli 80株 (VKPM B-7270, RU2119536)、E. coli NRRL B-12116 - B12121 (米国特許第4,388,405号)、E. coli H-9342 (FERM BP-6675)及びH-9343 (FERM BP-6676) (米国特許第6,344,347号)、E. coli H-9341 (FERM BP-6674) (EP1085087)、E. coli AI80/pFM201 (米国特許第6,258,554号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L-ヒスチジン生産菌の親株の例としては、L-ヒスチジン生合成系酵素をコードする遺伝子の1種以上の発現が増大した株も挙げられる。L-ヒスチジン生合成系酵素をコードする遺伝子の例としては、ATPホスホリボシルトランスフェラーゼ遺伝子(hisG)、ホスホリボシルAMPサイクロヒドロラーゼ遺伝子(hisI)、ホスホリボシル-ATPピロホスホヒドロラーゼ遺伝子(hisIE)、ホスホリボシルフォルミミノ-5-アミノイミダゾールカルボキサミドリボタイドイソメラーゼ遺伝子(hisA)、アミドトランスフェラーゼ遺伝子(hisH)、ヒスチジノールホスフェートアミノトランスフェラーゼ遺伝子(hisC)、ヒスチジノールフォスファターゼ遺伝子(hisB)、ヒスチジノールデヒドロゲナーゼ遺伝子(hisD)などが挙げられる。
hisG及びhisBHAFIにコードされるL-ヒスチジン生合成系酵素はL-ヒスチジンにより阻害されることが知られており、従って、L-ヒスチジン生産能は、ATPホスホリボシルトランスフェラーゼ遺伝子(hisG)にフィードバック阻害への耐性を付与する変異を導入することにより効率的に増大させることができる(ロシア特許第2003677号及び第2119536号)。
L-ヒスチジン産生能を有する株の具体例としては、L-ヒスチジン生合成系酵素をコードするDNAを保持するベクターを導入したE. coli FERM-P 5038及び5048 (JP 56-005099 A)、アミノ酸輸送の遺伝子であるrhtを導入したE.coli株(EP1016710A)、スルファグアニディン、DL-1,2,4-トリアゾール-3-アラニン及びストレプトマイシンに対する耐性を付与したE. coli 80株(VKPM B-7270, ロシア特許第2119536号)などが挙げられる。
L−グルタミン酸生産菌
L−グルタミン酸生産菌の親株の例としては、E. coli VL334thrC+ (EP 1172433)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。E. coli VL334 (VKPM B-1641)は、thrC遺伝子及びilvA遺伝子に変異を有するL-イソロイシン及びL-スレオニン要求性株である(米国特許第4,278,765号)。thrC遺伝子の野生型アレルは、野生型E. coli K12株 (VKPM B-7)の細胞で増殖したバクテリオファージP1を用いる一般的形質導入により導入された。この結果、L-イソロイシン要求性株L-グルタミン酸生産菌 VL334thrC+ (VKPM B-8961) が得られた。
L−グルタミン酸生産菌の親株の例としては、E. coli VL334thrC+ (EP 1172433)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。E. coli VL334 (VKPM B-1641)は、thrC遺伝子及びilvA遺伝子に変異を有するL-イソロイシン及びL-スレオニン要求性株である(米国特許第4,278,765号)。thrC遺伝子の野生型アレルは、野生型E. coli K12株 (VKPM B-7)の細胞で増殖したバクテリオファージP1を用いる一般的形質導入により導入された。この結果、L-イソロイシン要求性株L-グルタミン酸生産菌 VL334thrC+ (VKPM B-8961) が得られた。
L-グルタミン酸生産菌の親株の例としては、L-グルタミン酸生合成系酵素をコードする
遺伝子の1種以上の発現が増大した株が挙げられるがこれらに限定されない。L-グルタミン酸生合成系酵素をコードする遺伝子の例としては、グルタミン酸デヒドロゲナーゼ遺伝子(gdh)、グルタミンシンテターゼ遺伝子(glnA)、グルタミン酸シンテターゼ遺伝子(gltAB)、イソクエン酸デヒドロゲナーゼ遺伝子(icd)、アコニット酸ヒドラターゼ遺伝子(acn)、クエン酸シンターゼ遺伝子(glta)、ホスホエノールピルビン酸カルボキシラーゼ遺伝子(pepC)、ピルビン酸カルボキシラーゼ遺伝子(pyc)、ピルビン酸デヒドロゲナーゼ遺伝子(pdh)、ピルビン酸キナーゼ遺伝子(pykA,pykF)、ホスホエノールピルビン酸シンターゼ遺伝子(pps)、エノラーゼ遺伝子(eno)、ホスホグリセロムターゼ遺伝子(pgm)、ホスホグリセレートキナーゼ遺伝子(pgk)、グリセルアルデヒド-3-ホスフェートデヒドロゲナーゼ遺伝子(gap)、トリオースホスフェートイソメラーゼ遺伝子(tpi)、フルクトースビスホスフェートアルドラーゼ遺伝子(fbp)、ホスホフルクトキナーゼ遺伝子(pfk)、グルコースホスフェートイソメラーゼ遺伝子(pgi)などが挙げられる。
遺伝子の1種以上の発現が増大した株が挙げられるがこれらに限定されない。L-グルタミン酸生合成系酵素をコードする遺伝子の例としては、グルタミン酸デヒドロゲナーゼ遺伝子(gdh)、グルタミンシンテターゼ遺伝子(glnA)、グルタミン酸シンテターゼ遺伝子(gltAB)、イソクエン酸デヒドロゲナーゼ遺伝子(icd)、アコニット酸ヒドラターゼ遺伝子(acn)、クエン酸シンターゼ遺伝子(glta)、ホスホエノールピルビン酸カルボキシラーゼ遺伝子(pepC)、ピルビン酸カルボキシラーゼ遺伝子(pyc)、ピルビン酸デヒドロゲナーゼ遺伝子(pdh)、ピルビン酸キナーゼ遺伝子(pykA,pykF)、ホスホエノールピルビン酸シンターゼ遺伝子(pps)、エノラーゼ遺伝子(eno)、ホスホグリセロムターゼ遺伝子(pgm)、ホスホグリセレートキナーゼ遺伝子(pgk)、グリセルアルデヒド-3-ホスフェートデヒドロゲナーゼ遺伝子(gap)、トリオースホスフェートイソメラーゼ遺伝子(tpi)、フルクトースビスホスフェートアルドラーゼ遺伝子(fbp)、ホスホフルクトキナーゼ遺伝子(pfk)、グルコースホスフェートイソメラーゼ遺伝子(pgi)などが挙げられる。
クエン酸シンテターゼ遺伝子、ホスホエノールピルビン酸カルボキシラーゼ遺伝子、及び/またはグルタミン酸デヒドロゲナーゼ遺伝子の発現が増大するように改変された株の例としては、EP1078989A、EP955368A及びEP952221Aに開示されたものが挙げられる。
L-グルタミン酸産生細菌の親株の例としては、L−グルタミン酸の生合成経路から分岐してL−グルタミン酸以外の化合物を生成する反応を触媒する酵素の活性が低下または欠損している株も挙げられる。このような酵素の例としては、イソクエン酸リアーゼ(aceA)、α-ケトグルタレートデヒドロゲナーゼ(sucA)、ホスホトランスアデチラーゼ(pta)、アセテートキナーゼ(ack)、アデトヒドロキシ酸シンターゼ(ilvG)、アセトラクテートシンターゼ(ilvI)、フォルメートアセチルトランスフェラーゼ(pfl)、ラクテートデヒドロゲナーゼ(ldh)、グルタミン酸デカルボキシラーゼ(gadAB)などが挙げられる(括弧内は、その酵素をコードする遺伝子の名である)。α-ケトグルタレートデヒドロゲナーゼ活性が欠損した、または、α-ケトグルタレートデヒドロゲナーゼ活性が低下したエシェリヒア属に属する細菌、及び、それらの取得方法は米国特許第5,378,616 号及び第5,573,945号に記載されている。
具体例としては下記のものが挙げられる。
E. coli W3110sucA::Kmr
E. coli AJ12624 (FERM BP-3853)
E. coli AJ12628 (FERM BP-3854)
E. coli AJ12949 (FERM BP-4881)
E. coli W3110sucA::Kmr
E. coli AJ12624 (FERM BP-3853)
E. coli AJ12628 (FERM BP-3854)
E. coli AJ12949 (FERM BP-4881)
E. coli W3110sucA::Kmr は、E. coli W3110のα-ケトグルタレートデヒドロゲナーゼ遺伝子(以下、「sucA遺伝子」ともいう)を破壊することにより得られた株である。この株は、α-ケトグルタレートデヒドロゲナーゼを完全に欠損している。
L-グルタミン酸産生菌の他の例としては、エシェリヒア属に属し、アスパラギン酸代謝拮抗物質に耐性を有するものが挙げられる。これらの株は、α-ケトグルタレートデヒドロゲナーゼを欠損していてもよく、例えば、E. coli AJ13199 (FERM BP-5807) (米国特許第5.908,768号)、さらにL-グルタミン酸分解能が低下したFFRM P-12379(米国特許第5,393,671号); AJ13138 (FERM BP-5565) (米国特許第6,110,714号)などが挙げられる。
L-グルタミン酸産生菌の例としては、α-ケトグルタレートデヒドロゲナーゼ活性が欠損した、または、α-ケトグルタレートデヒドロゲナーゼ活性が低下したパントエア属に属する細菌が挙げられ、上記のようにして得ることができる。このような株の例としては、Pantoea ananatis AJ13356(米国特許第6,331,419号)がある。Pantoea ananatis AJ13356は、1998年2月19日、工業技術院生命工学工業技術研究所(現 独立行政法人 産業技術総
合研究所 特許生物寄託センター、〒305-8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に受託番号FERM P-16645として寄託され、1999年1月11日にブダペスト条約に基づく国際寄託に移管され、受託番号FERM BP-6616が付与されている。Pantoea ananatis
AJ13356は、αKGDH-E1サブユニット遺伝子(sucA)の破壊によりα-ケトグルタレートデヒドロゲナーゼ活性が欠損している。この株は、単離された時には、Enterobacter agglomeransと同定され、Enterobacter agglomerans AJ13356として寄託された。しかし、16S rRNAの塩基配列などに基づき、Pantoea ananatisに再分類されている。AJ13356は、上記寄託機関にEnterobacter agglomeransとして寄託されているが、本明細書では、Pantoea ananatisとして記載する。
合研究所 特許生物寄託センター、〒305-8566 日本国茨城県つくば市東1丁目1番地1 中央第6)に受託番号FERM P-16645として寄託され、1999年1月11日にブダペスト条約に基づく国際寄託に移管され、受託番号FERM BP-6616が付与されている。Pantoea ananatis
AJ13356は、αKGDH-E1サブユニット遺伝子(sucA)の破壊によりα-ケトグルタレートデヒドロゲナーゼ活性が欠損している。この株は、単離された時には、Enterobacter agglomeransと同定され、Enterobacter agglomerans AJ13356として寄託された。しかし、16S rRNAの塩基配列などに基づき、Pantoea ananatisに再分類されている。AJ13356は、上記寄託機関にEnterobacter agglomeransとして寄託されているが、本明細書では、Pantoea ananatisとして記載する。
L−フェニルアラニン生産菌
L−フェニルアラニン生産菌の親株の例としては、E.coli AJ12739 (tyrA::Tn10, tyrR) (VKPM B-8197)、変異型pheA34遺伝子を保持するE.coli HW1089 (ATCC 55371) (米国特許第 5,354,672号)、E.coli MWEC101-b (KR8903681)、E.coli NRRL B-12141, NRRL B-12145, NRRL B-12146及びNRRL B-12147 (米国特許第4,407,952号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。また、親株として、E. coli K-12 [W3110 (tyrA)/pPHAB] (FERM BP-3566)、E. coli K-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659)、E. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662)及びAJ 12604と命名されたE.
coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB] (FERM BP-3579)も使用できる(EP 488424
B1)。さらに、yedA遺伝子またはyddG遺伝子にコードされるタンパク質の活性が増大したエシェリヒア属に属するL-フェニルアラニン産生菌も使用できる(米国出願公開2003/0148473 A1及び2003/0157667 A1)。
L−フェニルアラニン生産菌の親株の例としては、E.coli AJ12739 (tyrA::Tn10, tyrR) (VKPM B-8197)、変異型pheA34遺伝子を保持するE.coli HW1089 (ATCC 55371) (米国特許第 5,354,672号)、E.coli MWEC101-b (KR8903681)、E.coli NRRL B-12141, NRRL B-12145, NRRL B-12146及びNRRL B-12147 (米国特許第4,407,952号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。また、親株として、E. coli K-12 [W3110 (tyrA)/pPHAB] (FERM BP-3566)、E. coli K-12 [W3110 (tyrA)/pPHAD] (FERM BP-12659)、E. coli K-12 [W3110 (tyrA)/pPHATerm] (FERM BP-12662)及びAJ 12604と命名されたE.
coli K-12 [W3110 (tyrA)/pBR-aroG4, pACMAB] (FERM BP-3579)も使用できる(EP 488424
B1)。さらに、yedA遺伝子またはyddG遺伝子にコードされるタンパク質の活性が増大したエシェリヒア属に属するL-フェニルアラニン産生菌も使用できる(米国出願公開2003/0148473 A1及び2003/0157667 A1)。
L−トリプトファン生産菌
L−トリプトファン生産菌の親株の例としては、変異trpS遺伝子によりコードされるトリプトファニル-tRNAシンテターゼが欠損したE. coli JP4735/pMU3028 (DSM10122)及びJP6015/pMU91 (DSM10123) (米国特許第5,756,345号)、セリンによるフィードバック阻害を受けないホスホグリセリン酸デヒドロゲナーゼをコードするserAアレル及びトリプトファンによるフィードバック阻害を受けないアントラニル酸シンターゼをコードするtrpEアレルを有するE. coli SV164 (pGH5) (米国特許第6,180,373号)、トリプトファナーゼが欠損したE. coli AGX17 (pGX44) (NRRL B-12263)及びAGX6(pGX50)aroP (NRRL B-12264) (米国特許第4,371,614号)、ホスホエノールピルビン酸産生能が増大したE. coli AGX17/pGX50,pACKG4-pps (WO9708333, 米国特許第6,319,696号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−トリプトファン生産菌の親株の例としては、変異trpS遺伝子によりコードされるトリプトファニル-tRNAシンテターゼが欠損したE. coli JP4735/pMU3028 (DSM10122)及びJP6015/pMU91 (DSM10123) (米国特許第5,756,345号)、セリンによるフィードバック阻害を受けないホスホグリセリン酸デヒドロゲナーゼをコードするserAアレル及びトリプトファンによるフィードバック阻害を受けないアントラニル酸シンターゼをコードするtrpEアレルを有するE. coli SV164 (pGH5) (米国特許第6,180,373号)、トリプトファナーゼが欠損したE. coli AGX17 (pGX44) (NRRL B-12263)及びAGX6(pGX50)aroP (NRRL B-12264) (米国特許第4,371,614号)、ホスホエノールピルビン酸産生能が増大したE. coli AGX17/pGX50,pACKG4-pps (WO9708333, 米国特許第6,319,696号)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
さらに、yedA遺伝子またはyddG遺伝子にコードされるタンパク質の活性が増大したエシェリヒア属に属するL-トリプトファン産生菌も使用できる(米国出願公開2003/0148473 A1及び2003/0157667 A1)。
L-トリプトファン産生菌の親株の例としては、アンスラニレートシンターゼ、ホスホグリセレートデヒドロゲナーゼ(serA)、及び、トリプトファンシンターゼ(trpAB)から選ばれる酵素の活性の一種以上が増大した株も挙げられる。アンスラニレートシンターゼ及びホスホグリセレートデヒドロゲナーゼは共にL-トリプトファン及びL-セリンによるフィードバック阻害を受けるので、フィードバック阻害を解除する変異をこれらの酵素に導入してもよい。このような変異を有する株の具体例としては、脱感作型アンスラニレートシンターゼを保持するE. coli SV164、及び、E. coli SV164に、フィードバック阻害が解除されたホスホグリセレートデヒドロゲナーゼをコードする変異serA遺伝子を含むプラスミドpGH5 (WO 94/08031)を導入することにより得られた形質転換株が挙げられる。
L-トリプトファン生産菌の親株の例としては、脱感作型アンスラニレートシンターゼを
コードする遺伝子を含むトリプトファンオペロンが導入された株(JP 57-71397 A, JP 62-244382 A, 米国特許第4,371,614号)も挙げられる。さらに、L-トリプトファン生産能を、トリプトファンオペロン(trpBA)中のトリプトファンシンターゼをコードする遺伝子の発現を増大させることにより付与してもよい。トリプトファンシンターゼは、それぞれtrpA及びtrpBによりコードされるα及びβサブユニットからなる。さらに、L-トリプトファン生産能を、イソクエン酸リアーゼ-リンゴ酸シンターゼオペロンの発現を増大させることにより改良してもよい(WO2005/103275)。
コードする遺伝子を含むトリプトファンオペロンが導入された株(JP 57-71397 A, JP 62-244382 A, 米国特許第4,371,614号)も挙げられる。さらに、L-トリプトファン生産能を、トリプトファンオペロン(trpBA)中のトリプトファンシンターゼをコードする遺伝子の発現を増大させることにより付与してもよい。トリプトファンシンターゼは、それぞれtrpA及びtrpBによりコードされるα及びβサブユニットからなる。さらに、L-トリプトファン生産能を、イソクエン酸リアーゼ-リンゴ酸シンターゼオペロンの発現を増大させることにより改良してもよい(WO2005/103275)。
L−プロリン生産菌
L−プロリン生産菌の親株の例としては、ilvA遺伝子が欠損し、L-プロリンを産生できるE. coli 702ilvA (VKPM B-8012) (EP 1172433)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L−プロリン生産菌の親株の例としては、ilvA遺伝子が欠損し、L-プロリンを産生できるE. coli 702ilvA (VKPM B-8012) (EP 1172433)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
本発明の細菌は、L-プロリン生合成に関与する遺伝子の一種以上の発現を増大することにより改良されていてもよい。L-プロリン産生菌に好ましい遺伝子の例としては、L-プロリンによるフィードバック阻害が解除されたグルタミン酸キナーゼをコードするproB遺伝子(ドイツ特許第3127361号)が挙げられる。さらに、本発明の細菌は、細菌の細胞からL-アミノ酸を排出するタンパク質をコードする遺伝子の一種以上の発現が増大することにより改良されていてもよい。このような遺伝子としては、b2682 遺伝子及びb2683遺伝子(ygaZH遺伝子) (EP1239041 A2)が挙げられる。
L-プロリン生産能を有するエシェリヒア属に属する細菌の例としては、NRRL B-12403及びNRRL B-12404 (英国特許第2075056号)、VKPM B-8012 (ロシア特許出願2000124295)、ドイツ特許第3127361号に記載のプラスミド変異体、Bloom F.R. et al (The 15th Miami winter symposium, 1983, p.34)に記載のプラスミド変異体などのE. coli 株が挙げられる。
L−アルギニン生産菌
L−アルギニン生産菌の親株の例としては、E. coli 237株 (VKPM B-7925) (米国出願公開2002/058315 A1)、及び、変異N-アセチルグルタミン酸シンターゼを保持するその誘導体株(ロシア特許出願第2001112869号)、E. coli 382株 (VKPM B-7926) (EP1170358A1)、N-アセチルグルタミン酸シンテターゼをコードするargA遺伝子が導入されたアルギニン産生株(EP1170361A1)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L-アルギニン産生菌の親株の例としては、L-アルギニン生合成系酵素をコードする遺伝子の1種以上の発現が増大した株も挙げられる。L-アルギニン生合成系酵素をコードする遺伝子の例としては、N-アセチルグルタミルホスフェートレダクターゼ遺伝子(argC)、オルニチンアセチルトランスフェラーゼ遺伝子(argJ)、N-アセチルグルタミン酸キナーゼ遺伝子(argB)、アセチルオルニチントランスアミナーゼ遺伝子(argD)、オルニチンカルバモイルトランスフェラーゼ遺伝子(argF)、アルギノコハク酸シンテターゼ遺伝子(argG)、アルギノコハク酸リアーゼ遺伝子(argH)、カルバモイルリン酸シンターゼ遺伝子(carAB)が挙げられる。
L−アルギニン生産菌の親株の例としては、E. coli 237株 (VKPM B-7925) (米国出願公開2002/058315 A1)、及び、変異N-アセチルグルタミン酸シンターゼを保持するその誘導体株(ロシア特許出願第2001112869号)、E. coli 382株 (VKPM B-7926) (EP1170358A1)、N-アセチルグルタミン酸シンテターゼをコードするargA遺伝子が導入されたアルギニン産生株(EP1170361A1)などのエシェリヒア属に属する株が挙げられるがこれらに限定されない。
L-アルギニン産生菌の親株の例としては、L-アルギニン生合成系酵素をコードする遺伝子の1種以上の発現が増大した株も挙げられる。L-アルギニン生合成系酵素をコードする遺伝子の例としては、N-アセチルグルタミルホスフェートレダクターゼ遺伝子(argC)、オルニチンアセチルトランスフェラーゼ遺伝子(argJ)、N-アセチルグルタミン酸キナーゼ遺伝子(argB)、アセチルオルニチントランスアミナーゼ遺伝子(argD)、オルニチンカルバモイルトランスフェラーゼ遺伝子(argF)、アルギノコハク酸シンテターゼ遺伝子(argG)、アルギノコハク酸リアーゼ遺伝子(argH)、カルバモイルリン酸シンターゼ遺伝子(carAB)が挙げられる。
L−バリン生産菌
L−バリン生産菌の親株の例としては、ilvGMEDAオペロンを過剰発現するように改変された株(米国特許第5,998,178号)が挙げられるがこれらに限定されない。アテニュエーションに必要なilvGMEDAオペロンの領域を除去し、生産されるL-バリンによりオペロンの発現が減衰しないようにすることが好ましい。さらに、オペロンのilvA遺伝子は破壊され、スレオニンデアミナーゼ活性が減少することが好ましい。
L−バリン生産菌の親株の例としては、ilvGMEDAオペロンを過剰発現するように改変された株(米国特許第5,998,178号)が挙げられるがこれらに限定されない。アテニュエーションに必要なilvGMEDAオペロンの領域を除去し、生産されるL-バリンによりオペロンの発現が減衰しないようにすることが好ましい。さらに、オペロンのilvA遺伝子は破壊され、スレオニンデアミナーゼ活性が減少することが好ましい。
L-バリン産生菌の親株の例としては、アミノアシルt-RNAシンテターゼの変異を有する変異株(米国特許第5,658,766号)も挙げられる。例えば、イソロイシンtRNAシンテターゼをコードするileS 遺伝子に変異を有するE. coli VL1970が使用できる。E. coli VL1970は、1988年6月24日、ルシアン・ナショナル・コレクション・オブ・インダストリアル・マイクロオルガニズムズ(VKPM) (1 Dorozhny proezd., 1 Moscow 117545, Russia)に、受託番号VKPM B-4411で寄託されている。
さらに、生育にリポ酸を要求する、及び/または、H+-ATPaseを欠失している変異株(WO96/06926)を親株として用いることができる。
L−イソロイシン生産菌
L−イソロイシン生産菌の親株の例としては、6−ジメチルアミノプリンに耐性を有する変異株(JP 5-304969 A)、チアイソロイシン、イソロイシンヒドロキシメートなどのイソロイシンアナログに耐性を有する変異株、さらにDL-エチオニン及び/またはアルギニンヒドロキシメートに耐性を有する変異株(JP 5-130882 A).が挙げられるがこれらに限定されない。さらに、スレオニンデアミナーゼ、アセトヒドロキシ酸シンターゼなどのL-イソロイシン生合成に関与するタンパク質をコードする遺伝子で形質転換された組換え株もまた親株として使用できる(JP 2-458 A, FR 0356739, 及び米国特許第5,998,178号)。
L−イソロイシン生産菌の親株の例としては、6−ジメチルアミノプリンに耐性を有する変異株(JP 5-304969 A)、チアイソロイシン、イソロイシンヒドロキシメートなどのイソロイシンアナログに耐性を有する変異株、さらにDL-エチオニン及び/またはアルギニンヒドロキシメートに耐性を有する変異株(JP 5-130882 A).が挙げられるがこれらに限定されない。さらに、スレオニンデアミナーゼ、アセトヒドロキシ酸シンターゼなどのL-イソロイシン生合成に関与するタンパク質をコードする遺伝子で形質転換された組換え株もまた親株として使用できる(JP 2-458 A, FR 0356739, 及び米国特許第5,998,178号)。
<2>本発明の製造法
上記のようにして得られる本発明微生物を、培地中のリン濃度がphoレギュロンプロモーターによる発現が誘導される濃度である培地中で培養し、該培養物中にL−アミノ酸を生産蓄積させ、該培養物からL−アミノ酸を採取することにより、L−アミノ酸を効率よく製造することができる。
上記のようにして得られる本発明微生物を、培地中のリン濃度がphoレギュロンプロモーターによる発現が誘導される濃度である培地中で培養し、該培養物中にL−アミノ酸を生産蓄積させ、該培養物からL−アミノ酸を採取することにより、L−アミノ酸を効率よく製造することができる。
微生物を培養した場合、微生物自身では生合成できないような栄養源、あるいは栄養源の生合成を行うために必要な元素が枯渇した場合、微生物の生育は停止する。このような物質は多数あるが、この中でもっとも最初に枯渇した物質の量により、微生物の生育は制限される。従って、本発明においては、必要な栄養源のうち、リンが第1の生育の律速因子となっているような培地を、リン濃度が制限された培地と定義する。また、phoレギュロンプロモーターによる発現が誘導される濃度を「リン制限濃度」ともいう。
本発明で用いられる培地は、栄養源として炭素源、窒素源を含んでいればいずれでもよいが、発酵培地中のリンの濃度がリン制限濃度になるように調整された培地である。ここで、リンとは、リン分子を含んでいるものであればいずれでもよいが、中でもリン酸が好ましく、添加形態としては、リン酸塩が好ましい。塩としては特に制限されず、アンモニウム塩、カルシウム塩、ナトリウム塩でもよく、リン酸2水素カリウム、リン酸水素2カリウム、ピロリン酸などのリン酸ポリマー等が用いられる。また培地中にこれらの物質を1種のみ含んでいてもよいし、2種以上含んでいてもよい。リン制限濃度は、リンを大量に含む(すなわちリンが生育の律速因子とならない濃度でリンを含む)培地と比べてPho レギュロンプロモーターが活性化される濃度であれば、いずれでもよいが、具体的には、発酵培地に含まれるリン濃度で通常には100μM、好ましくは10μM以下、より好ましくは4μM以下である。培養後半であれば、培地中のリン濃度は0であってもよい。
尚、発酵培地には微生物の生育に最低限必要なリン量を含んでいればよく、一時的に不足している状態になってもよい。一時的とは、例えば発酵全体の時間のうち約20%、40%、最大で約60%の時間でリンが不足している状態でもよい。
本発明の培養法は、回分培養(batch culture)、流加培養(Fed−batch culture)、連続培養法(continuous culture)のいずれも用いることができる。また、本発明におい
ては、L−アミノ酸蓄積を一定以上に保つために、種培養と本培養と分けて行ってもよく、種培養をフラスコ等のしんとう培養、あるいは回分培養で行い、本培養を流加培養、連続培養で行ってもよく、種培養、本培養ともに回分培養で行ってもよい。
ては、L−アミノ酸蓄積を一定以上に保つために、種培養と本培養と分けて行ってもよく、種培養をフラスコ等のしんとう培養、あるいは回分培養で行い、本培養を流加培養、連続培養で行ってもよく、種培養、本培養ともに回分培養で行ってもよい。
ここで本発明において、「初発培地」とは、流加させる前の回分培養(batch)培養に用いる培地のことを意味し、「流加培地」とは、流加培養を行う際に発酵槽に供給する培地のことを意味する。また、本発明において「発酵培地」とは、発酵槽中の培地を意味し、この発酵培地からL−アミノ酸が回収される。また、本発明において、「発酵槽」とは、L−アミノ酸発酵を行う器を意味し、発酵タンクを用いてもジャーファーメンターを用いてもよい。また、その容量は、L−アミノ酸を生成・回収できる容量であればいずれでもよい。
リンは初発培地でリン制限濃度になるように調整されていてもよいし、流加培地でリン制限濃度になるように制限してもよいし、これらを組み合わせてもよい。例えば流加培養によりリンの濃度を制限する場合には、発酵培地中のリン濃度を好ましくは100μM以下、より好ましく10μM以下、より一層好ましくは4μM以下に制御する。
また、初発培地、流加培地とも、これらのリンを混合してもよいし、流加培地のリンを初発培地と変更してもよい。
尚、リンは、L-アミノ酸生産期である培養後半においてリン制限濃度に制限されていることが好ましい。例えば、本発明の方法が、L−アミノ酸生産能を持つ微生物を増殖させる段階(増殖期)と、L−アミノ酸を産生させる段階(L−アミノ酸生産期)を含む場合、L−アミノ酸生産期においてリンをリン制限濃度に制限すればよく、L−アミノ酸蓄積微生物を増殖させる増殖期においては、リンをリン制限濃度を超えて培地に含有させてもよく、リン制限濃度に制限してもよい。また、L−アミノ酸を産生させる段階においても、その段階の全期間でリンの含有量が前記の範囲である必要はなく、同段階の初期に含有量が前記範囲以上になるようにリンを存在させ、培養時間に応じて減少してもよい。
本発明において培養後半とは、主にアミノ酸生産期を意味し、菌体増殖期と区別される。本発明における「増殖期」とは、培養開始から3時間、好ましくは6時間、特に好ましくは、10時間以内の、リンが主に菌体生育に使用されている時期、すなわち微生物が対数的に増殖している時期を意味し、本発明における「培養後半」とは、培養終了まで6時間、好ましくは10時間、特に好ましくは20時間前の期間で、炭素源が主にL−アミノ酸生産に用いられている時期を意味する。
本発明に用いられる培地に含まれる炭素源としては、炭素源としては、グルコース、グリセロール、フラクトース、スクロース、マルトース、マンノース、ガラクトース、澱粉加水分解物、糖蜜等の糖類が使用でき、特にグルコース、スクロースが好ましい。その他、酢酸、クエン酸等の有機酸、エタノール等のアルコール類も単独あるいは他の炭素源と併用して用いることができる。また、炭素源となる原料としては、ケインモラセス、ビートモラセス、ハイテストモラセス、シトラスモラセスを用いてもよいし、セルロース、デンプン、コーン、シリアル、タピオカ等の天然原料の加水分解物を用いてもよい。また培養液中に溶存した二酸化炭素も炭素源として使用出来る。これらの炭素源が初発培地にも流加培地にも用いることができる。また、初発培地、流加培地とも、これらの炭素源を混合してもよいし、流加培地の炭素源を初発培地と変更してもよい。例えば、初発培地をグルコースで培養し、流加培地をスクロースで培養する場合である。
本発明の培地中に含まれる窒素源としては、アンモニア、硫酸アンモニウム、炭酸アンモニウム、塩化アンモニウム、リン酸アンモニウム、酢酸アンモニウム、ウレア等のアン
モニウム塩または硝酸塩等が使用することができ、pH調整に用いられるアンモニアガス、アンモニア水も窒素源として利用できる。また、ペプトン、酵母エキス、肉エキス、麦芽エキス、コーンスティープリカー、大豆加水分解物等も利用出来る。これらの窒素源は、初発培地にも流加培地にも用いることができる。また、初発培地、流加培地とも、これらの窒素源を混合してもよいし、流加培地の窒素源を初発培地と変更してもよい。
モニウム塩または硝酸塩等が使用することができ、pH調整に用いられるアンモニアガス、アンモニア水も窒素源として利用できる。また、ペプトン、酵母エキス、肉エキス、麦芽エキス、コーンスティープリカー、大豆加水分解物等も利用出来る。これらの窒素源は、初発培地にも流加培地にも用いることができる。また、初発培地、流加培地とも、これらの窒素源を混合してもよいし、流加培地の窒素源を初発培地と変更してもよい。
また本発明の培地には、炭素源、窒素源、リン源の他に硫黄源を含んでいることが好ましい。硫黄源としては、硫黄分子を含んでいるものであればいずれでもよいが、硫酸塩、チオ硫酸塩、亜硫酸塩等の硫酸塩、システイン、シスチン、グルタチオン等の含硫アミノ酸が望ましく、なかでも硫酸アンモニウムが望ましい。
また本発明の培地には、炭素源、窒素源、リン源の他に、増殖促進因子を含んでいてもよい。増殖促進因子とは、微量金属類、アミノ酸、ビタミン、脂肪酸、核酸、更にこれらのものを含有するペプトン、カザミノ酸、酵母エキス、大豆たん白分解物等が使用できる。
微量金属類としては、鉄、マンガン、マグネシウム、カルシウム等が挙げられ、ビタミンとしては、ビタミンB1、ビタミンB2、ビタミンB6、ニコチン酸、ニコチン酸アミド、ビタミンB12等が挙げられる。これらの増殖促進因子は初発培地に含まれていてもよいし、流加培地に含まれていてもよい。
また本発明の培地には、生育にアミノ酸などを要求する栄養要求性変異株を使用する場合には要求される栄養素を補添することが好ましい。
培養は、通常には発酵温度20〜45℃、特に好ましくは33〜42℃で通気培養を行う。ここで酸素濃度は、通常には5〜50%に、好ましくは10%程度に調節して行う。また、通常、pHを5〜9に制御し、通気培養を行う。培養中にpHが下がる場合には、例えば、炭酸カルシウムを加えるか、アンモニアガス、アンモニア水等のアルカリで中和する。このような条件下で、好ましくは10時間〜120時間程度培養することにより、培養液中に著量のL−アミノ酸が蓄積される。蓄積されるL−アミノ酸の濃度は野生株より高く、培地中から採取・回収できる濃度であればいずれでもよいが、50g/L以上、好ましくは75g/L以上、さらに好ましくは100g/L以上である。
培養終了後の培養液からL−アミノ酸を採取する方法は、公知の回収方法に従って行えばよい。例えば、培養液から遠心分離等によって菌体を除去した後に、濃縮晶析することによって採取される。
以下に、実施例を挙げて本発明を更に具体的に説明する
(1)大腸菌リジン生産菌のリン制限条件下での培養
大腸菌リジン生産菌WC196(FERM BP-5252)を用いて、リジンの生産能を評価した。グルコースを40g/L、MgSO47水和物を1g/L、KH2PO4を0または0.125、0.5、1g/L、(NH4)2SO4を16g/L、FeSO47水和物を10mg/L、MnSO44〜5水和物を10mg/L、酵母エキスを2g/L、CaCO3を50g/L含む培地にて500mL容坂口フラスコを用いて培養した。培養開始時の培養液量は20mLとし、回転速度120rpmで往復振とうし、37℃で培養を行った。
大腸菌リジン生産菌WC196(FERM BP-5252)を用いて、リジンの生産能を評価した。グルコースを40g/L、MgSO47水和物を1g/L、KH2PO4を0または0.125、0.5、1g/L、(NH4)2SO4を16g/L、FeSO47水和物を10mg/L、MnSO44〜5水和物を10mg/L、酵母エキスを2g/L、CaCO3を50g/L含む培地にて500mL容坂口フラスコを用いて培養した。培養開始時の培養液量は20mLとし、回転速度120rpmで往復振とうし、37℃で培養を行った。
培地、容器等は全てオートクレーブ滅菌を行った後に供した。このとき、培養液中の菌体濃度、グルコース濃度、及びL-リジン蓄積を測定した。菌体濃度(OD600)は、適当倍率に0.1規定塩酸で希釈した培養液を用い、600nmの濁度を分光光度計(ベックマン社)で測定
して求めた。残存グルコース濃度、及びL-リジン濃度は遠心分離により除菌した培養液上清を適当倍率に水で希釈した後にバイオテックアナライザー(サクラ精器)により測定した。培養は24〜65時間行い、培養液中のグルコースが全て消費されるまで培養を行った。
して求めた。残存グルコース濃度、及びL-リジン濃度は遠心分離により除菌した培養液上清を適当倍率に水で希釈した後にバイオテックアナライザー(サクラ精器)により測定した。培養は24〜65時間行い、培養液中のグルコースが全て消費されるまで培養を行った。
下表2に示すように、KH2PO4を0.5または1.0g/Lとして培養を行ったときには、OD600に示すように最終の到達菌体量がほぼ同一であるのに対して、KH2PO4を0または0.125g/Lとして培養を行ったときには、最終の到達菌体量が低下した。これは、リンが培養途中で欠乏している、すなわちリンが制限されているためと考えられる。更に、KH2PO4を0または0.125g/Lとして培養を行ったときには、リジン蓄積も対照(1.0g/L)に比較して減少が認められた。なお、培養液中にはKH2PO4由来のリンのほかに、酵母エキス由来のリンがKH2PO4換算で約0.25g/L含まれている。
すなわち、リンが制限されている条件においては、リジン生産菌は生育が低下し、リジンの生産能も低下することが示された。
(2)大腸菌野生株の遺伝子発現解析
大腸菌野生株MG1655を、22.2 mMグルコース、50 mM NaCl、0.523 mM NH4Cl、1 mM (NH4)2SO4、0.01 mM FeSO4・7H2O、0.005 mM CaCl2、0.01 mM MnSO4・4-5H2O、1 mMチアミン-HCl、40 mM MOPS-KOH(pH7.2)、及び、種々の濃度のKH2PO4を含む培地にて、500mL容坂口フラスコを用いて培養した。KH2PO4の濃度はリン酸充足条件においては1mM、リン酸不足条件では50μM として培養を行った。培養開始時の培養液量は50mLとし、回転速度120rpmで往復振とうし、37℃で培養を行った。培地、容器等は全てオートクレーブ滅菌を行った後に供した。
大腸菌野生株MG1655を、22.2 mMグルコース、50 mM NaCl、0.523 mM NH4Cl、1 mM (NH4)2SO4、0.01 mM FeSO4・7H2O、0.005 mM CaCl2、0.01 mM MnSO4・4-5H2O、1 mMチアミン-HCl、40 mM MOPS-KOH(pH7.2)、及び、種々の濃度のKH2PO4を含む培地にて、500mL容坂口フラスコを用いて培養した。KH2PO4の濃度はリン酸充足条件においては1mM、リン酸不足条件では50μM として培養を行った。培養開始時の培養液量は50mLとし、回転速度120rpmで往復振とうし、37℃で培養を行った。培地、容器等は全てオートクレーブ滅菌を行った後に供した。
リン酸充足条件の対数増殖期、及びリン酸不足条件でリン酸が枯渇して増殖が停止した2時間後にフラスコより約10mLの培養液をサンプリングし、直ちに氷上にて冷却した後に冷却遠心機にて10000×g、2分の条件で遠心分離を行い培養液上清を除去した。回収した菌体から、QIAGEN社のRNeasy kitを用い、添付のプロトコールによって全RNAを回収した。得られたRNAは、アガロースゲル電気泳動にて分解されることなく回収されていることを確認し、紫外線領域の吸光度を測定することにより濃度を定量した。得られたRNAは、-80℃にて密閉保存し、次のDNAマクロアレイを用いた遺伝子発現解析に供した。
得られたRNA20μgを鋳型に、dATP、dGTP、dTTPを各1mM、及び33PでラベルされたdCTPを比放射能で1500MBqの量で基質として用い、Promega社のReverse Transcription Kitを用いて逆転写反応を行い、各フェーズのラベルされたcDNAを得た。
得られたcDNAをプローブとして、Genosys社の大腸菌マクロアレイメンブレンに対して、プロトコールに従い、ハイブリダイゼーションを行い、ハイブリダイゼーション終了後にメンブレンの洗浄を行った。洗浄後のメンブレンを密閉し、イメージングプレートに48時間、暗黒下で密着させ感光させた。感光させたイメージングプレートを、富士フィルム社製フルオロイメージングアナライザーFLA-3000Gにより読取り、得られた読取り画像をDNAアレイ画像解析システムAISにとりこみ、各スポットの濃度を定量することにより、各フェーズでの遺伝子発現プロファイルのデータを得た。
得られたDNAアレイデータより、対数増殖期には発現レベルが高いが、リンが枯渇して増殖が停止した時期には発現レベルが低下するような遺伝子をスチューデントのt検定により、糖代謝系、及びリジン生合成系の遺伝子の中から探索した。その結果、lysA遺伝子の遺伝子発現が有意に低下していることを見出した(表3)。また、得られたDNAアレイデータより、対数増殖期には発現レベルが低いが、リンが枯渇して増殖が停止した時期には発現レベルが増加するような遺伝子をスチューデントのt検定により、大腸菌の全遺伝子
の中から探索した。その結果、phoA遺伝子の遺伝子発現が有意に増加していることを見出した(表3)。
の中から探索した。その結果、phoA遺伝子の遺伝子発現が有意に増加していることを見出した(表3)。
培養液中のリン酸濃度は、培養液を10000rpm、2分間の条件で遠心分離した後に培養液を適当な濃度に希釈し、ピーテストワコー(和光純薬工業)を用いて分析を行った。
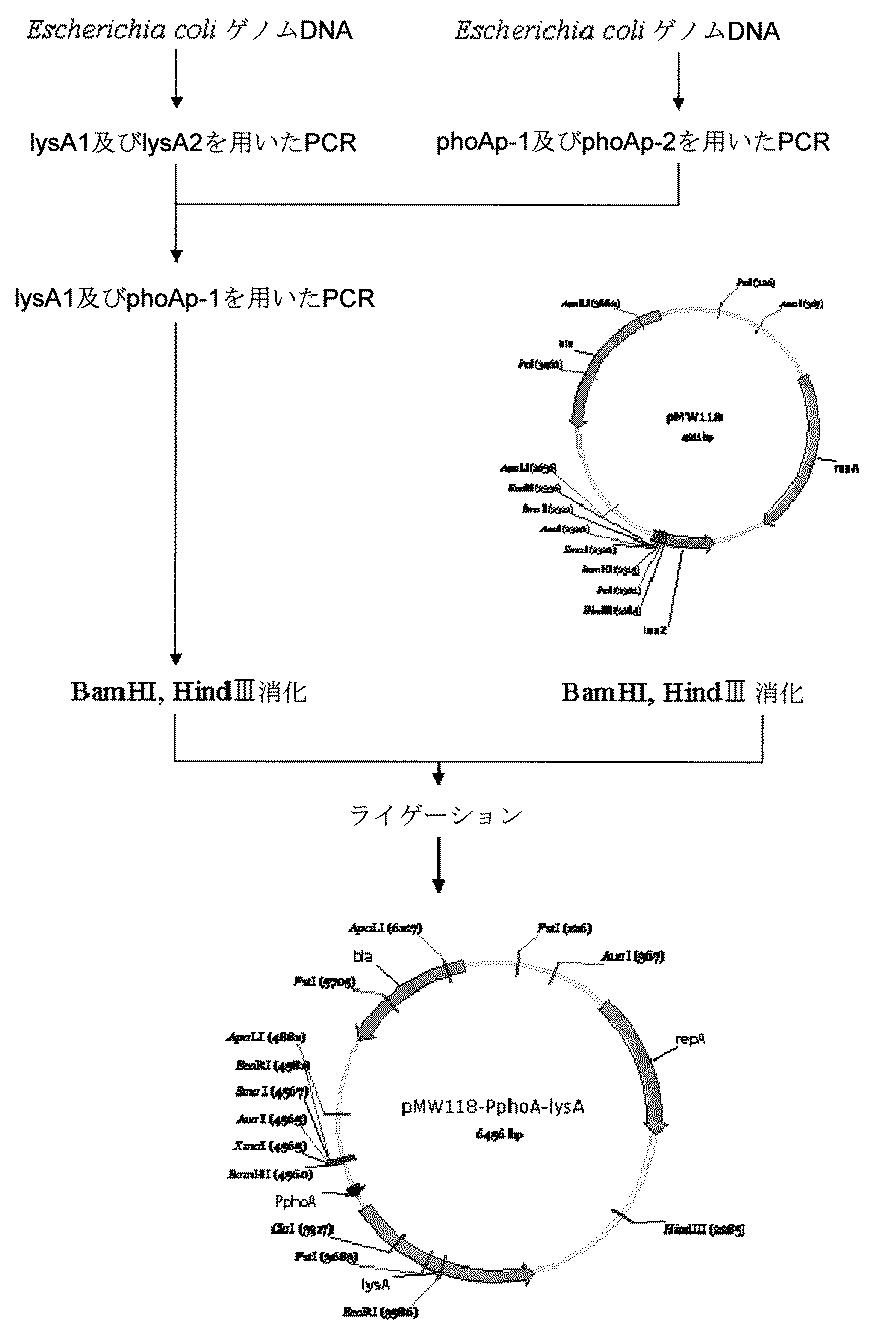
(3)プロモーター領域をphoA遺伝子のプロモーターに置換したlysA遺伝子を増幅した菌株の構築
プロモーターをphoA遺伝子のプロモーターに置換したlysA遺伝子(以下、PphoA-lysAと略す)は、クロスオーバーPCR法(Link AJ, Phillips D, Church GM、J.Bacteriol.Vol.179.p6228-6237,1997記載)により構築した。下表4に記載したプライマーLysA-1,LysA-2(配列番号22及び23)とphoAp-1,phoAp-2(配列番号26及び27)でE.coli K-12株より取得したゲノムDNAを鋳型にそれぞれPCRを行った。この際、プライマーLysA-1とLysA-2、及びphoAp-1とphoAp-2のモル比は10:1にて行った。得られた一回目のPCR産物を鋳型に、プライマーLysA-1とphoAp-1を用いて二回目のPCRを行った。得られたPCR産物はBamHIとHindIIIで切断し、pMW118(ニッポンジーン社)をBamHI-HindIIIで切断したものと合わせてDNA ligation kit ver.2(宝酒造)を用いてライゲーション反応を行った。この結合反応液にてDH5αコンピテント細胞(宝酒造)を形質転換し、アンピシリン(Ap)(ナカライテスク)を50 μg/mL含むLB寒天プレート(LB+Apプレート)にまき、37℃でコロニーを選択した。コロニーを50μg/mLのApを含むLB培地で37℃にて試験管培養し、Wizard Plus Miniprep(Promega社)を用いてプラスミドを抽出した。抽出したプラスミドは、BamHIとHindIIIで切断し、目的の長さの挿入配列を有するものを選択して、目的とするプラスミドpMW-PphoA-lysAを構築した。
プロモーターをphoA遺伝子のプロモーターに置換したlysA遺伝子(以下、PphoA-lysAと略す)は、クロスオーバーPCR法(Link AJ, Phillips D, Church GM、J.Bacteriol.Vol.179.p6228-6237,1997記載)により構築した。下表4に記載したプライマーLysA-1,LysA-2(配列番号22及び23)とphoAp-1,phoAp-2(配列番号26及び27)でE.coli K-12株より取得したゲノムDNAを鋳型にそれぞれPCRを行った。この際、プライマーLysA-1とLysA-2、及びphoAp-1とphoAp-2のモル比は10:1にて行った。得られた一回目のPCR産物を鋳型に、プライマーLysA-1とphoAp-1を用いて二回目のPCRを行った。得られたPCR産物はBamHIとHindIIIで切断し、pMW118(ニッポンジーン社)をBamHI-HindIIIで切断したものと合わせてDNA ligation kit ver.2(宝酒造)を用いてライゲーション反応を行った。この結合反応液にてDH5αコンピテント細胞(宝酒造)を形質転換し、アンピシリン(Ap)(ナカライテスク)を50 μg/mL含むLB寒天プレート(LB+Apプレート)にまき、37℃でコロニーを選択した。コロニーを50μg/mLのApを含むLB培地で37℃にて試験管培養し、Wizard Plus Miniprep(Promega社)を用いてプラスミドを抽出した。抽出したプラスミドは、BamHIとHindIIIで切断し、目的の長さの挿入配列を有するものを選択して、目的とするプラスミドpMW-PphoA-lysAを構築した。
このプラスミドpMW-PphoA-lysAにてE.coli WC196の形質転換を行い、目的とする菌株WC196/pMW-PphoA-lysAを得た。
(4)lysA遺伝子、及びlysR遺伝子を増幅した菌株の構築
下表4に記載したプライマーLysA-3,LysA-4(配列番号24及び25)を用い、E.coli K-12株より取得したゲノムDNAを鋳型にPCRを行った。得られたPCR産物はBamHIとHindIIIで切断し、pMW118(ニッポンジーン社)をBamHI-HindIIIで切断したものと合わせてDNA ligation kit ver.2(宝酒造)を用いてライゲーション反応を行った。この結合反応液にてDH5αコンピテント細胞(宝酒造)を形質転換し、アンピシリン(Ap)(ナカライテスク)を50 μg/mL含むLB寒天プレート(LB+Apプレート)にまき、37℃でコロニーを選択した。コロニーを50 μg/mL のApを含むLB培地で37℃にて試験管培養し、Wizard Plus Miniprep(Promega社)を用いてプラスミドを抽出した。抽出したプラスミドは、BamHIとHindIIIで切断し、目的の長さの挿入配列を有するものを選択して、目的とするプラスミドpMW-lysARを構築した。
下表4に記載したプライマーLysA-3,LysA-4(配列番号24及び25)を用い、E.coli K-12株より取得したゲノムDNAを鋳型にPCRを行った。得られたPCR産物はBamHIとHindIIIで切断し、pMW118(ニッポンジーン社)をBamHI-HindIIIで切断したものと合わせてDNA ligation kit ver.2(宝酒造)を用いてライゲーション反応を行った。この結合反応液にてDH5αコンピテント細胞(宝酒造)を形質転換し、アンピシリン(Ap)(ナカライテスク)を50 μg/mL含むLB寒天プレート(LB+Apプレート)にまき、37℃でコロニーを選択した。コロニーを50 μg/mL のApを含むLB培地で37℃にて試験管培養し、Wizard Plus Miniprep(Promega社)を用いてプラスミドを抽出した。抽出したプラスミドは、BamHIとHindIIIで切断し、目的の長さの挿入配列を有するものを選択して、目的とするプラスミドpMW-lysARを構築した。
このプラスミドpMW-lysARにてE.coli WC196の形質転換を行い、目的とする菌株WC196/pMW-lysARを得た。
(5)2種のlysA遺伝子増幅株によるリジン生産
上記(3)及び(4)にて作成した株、及びベクターのみのコントロールとしてWC196/pMW118を用いて、L-リジンの生産能を評価した。グルコースを40g/L、MgSO47水和物を1g/L、KH2PO4を0、0.125、または1g/L、(NH4)2SO4を16g/L、FeSO47水和物を10mg/L、MnSO44〜5水和物を10mg/L、酵母エキスを2g/L、CaCO3を50g/L、アンピシリンを100μg/mL含む培地にて500mL容坂口フラスコを用いて培養した。上記(1)で示したように、KH2PO4を0または0.125g/Lで培養した条件では、リンが制限されており、KH2PO4を1g/Lで培養した条件ではリンが充足している。培養開始時の培養液量は20mLとし、回転速度120rpmで往復振と
うし、37℃で培養を行った。培地、容器等は全てオートクレーブ滅菌を行った後に供した。このとき、培養液中の菌体濃度、グルコース濃度、及びリジン蓄積を測定した。菌体濃度(OD600)は、適当倍率に0.1規定塩酸で希釈した培養液を用い、600nmの濁度を分光光度計(ベックマン社)で測定して求めた。残存グルコース濃度、及びリジン濃度は遠心分離により除菌した培養液上清を適当倍率に水で希釈した後にバイオテックアナライザー(サクラ精器)により測定した。培養は24〜65時間行い、培養液中のグルコースが全て消費されるまで培養を行った。
上記(3)及び(4)にて作成した株、及びベクターのみのコントロールとしてWC196/pMW118を用いて、L-リジンの生産能を評価した。グルコースを40g/L、MgSO47水和物を1g/L、KH2PO4を0、0.125、または1g/L、(NH4)2SO4を16g/L、FeSO47水和物を10mg/L、MnSO44〜5水和物を10mg/L、酵母エキスを2g/L、CaCO3を50g/L、アンピシリンを100μg/mL含む培地にて500mL容坂口フラスコを用いて培養した。上記(1)で示したように、KH2PO4を0または0.125g/Lで培養した条件では、リンが制限されており、KH2PO4を1g/Lで培養した条件ではリンが充足している。培養開始時の培養液量は20mLとし、回転速度120rpmで往復振と
うし、37℃で培養を行った。培地、容器等は全てオートクレーブ滅菌を行った後に供した。このとき、培養液中の菌体濃度、グルコース濃度、及びリジン蓄積を測定した。菌体濃度(OD600)は、適当倍率に0.1規定塩酸で希釈した培養液を用い、600nmの濁度を分光光度計(ベックマン社)で測定して求めた。残存グルコース濃度、及びリジン濃度は遠心分離により除菌した培養液上清を適当倍率に水で希釈した後にバイオテックアナライザー(サクラ精器)により測定した。培養は24〜65時間行い、培養液中のグルコースが全て消費されるまで培養を行った。
結果を下表5に示す。リンが培養中常に充足しており、生育制限物質となっていない条件下(KH2PO4が1g/L含まれている培地)においては、いずれの株でも生育、リジン蓄積ともほぼ同等であった。一方、リンが培養途中に枯渇して生育制限物質となっている条件(KH2PO4が0.125または0g/L含まれている条件)においては、コントロール株、及びWC196/pMW-lysARにおいてはリジン蓄積が低下するのに対して、WC196/pMW-PphoA-lysAにおいてはこれらの株に比較してリジン蓄積が顕著に増加することが確認された。すなわち、phoAプロモーターにより発現するようなlysA遺伝子を導入することで、リンを制限した培養条件においてリジンの生産能が向上することが確認された。
Claims (11)
- phoレギュロンプロモーターの下流に、そのプロモーターにより発現するように、L−アミノ酸生合成系酵素をコードする構造遺伝子が接続されたDNA断片を導入することにより、前記プロモーターによる発現の誘導によってL−アミノ酸生合成系酵素の活性が増大するように改変された、腸内細菌科に属するL-アミノ酸生産能を有する微生物を培地で培養して、L−アミノ酸を該培地に生成蓄積させ、該培地よりL−アミノ酸を採取する、L−アミノ酸の製造方法であって、前記培地中のリン濃度が、前記プロモーターによる発現が誘導される濃度である方法。
- 前記phoレギュロンプロモーターが、phoA, phoB, phoE, phoH, asr, argP, ugpB, pstS, psiE及びphnCから選択される遺伝子のプロモーターである請求項1に記載の製造法。
- 前記phoレギュロンプロモーターが、pho boxを有するプロモーターである、請求項1又は2に記載の製造法。
- 前記培地中のリン濃度が、100μM/L以下に制御される請求項1〜3のいずれか一項に記載の製造法。
- 前記L−アミノ酸生合成系酵素が、改変前の微生物においては、リンが枯渇した条件下で発現レベルが低下するものである、請求項1〜4のいずれか一項に記載の製造法。
- 前記DNA断片がマルチコピー型ベクターに搭載されている、又は、微生物の染色体DNA上に多コピー導入されている、請求項1〜5のいずれか一項に記載の方法。
- 前記腸内細菌科に属する微生物が、エシェリヒア属細菌、エンテロバクター属細菌、パントエア属細菌、クレブシエラ細菌、セラチア属細菌からなる群より選ばれる微生物である、請求項1〜6のいずれか一項に記載の方法。
- 前記L−アミノ酸がL−リジン、L−スレオニン、L−トリプトファン、L−グルタミン酸からなる群より選択される1種または2種以上のL−アミノ酸である請求項1〜7のいずれか一項に記載の方法。
- 前記L−アミノ酸がL−リジンであり、L−アミノ酸生合成系酵素がジヒドロジピコリン酸レダクターゼ、ジアミノピメリン酸脱炭酸酵素、ジアミノピメリン酸デヒドロゲナーゼ、ホスホエノールピルビン酸カルボキシラーゼ、アスパラギン酸アミノトランスフェラーゼ、ジアミノピメリン酸エピメラーゼ、アスパラギン酸セミアルデヒド脱水素酵素、テトラヒドロジピコリン酸スクシニラーゼ、及び、スクシニルジアミノピメリン酸デアシラーゼからなる群より選択される1種または2種以上の酵素である請求項8に記載の方法。
- 前記L−アミノ酸がL−スレオニンであり、L−アミノ酸生合成系酵素がアスパルトキナーゼIII、アスパラギン酸セミアルデヒドデヒドロゲナーゼ、thrオペロンにコードされるアスパルトキナーゼI、ホモセリンキナーゼ、及び、スレオニンシンターゼからなる群より選択される1種または2種以上の酵素である請求項8に記載の方法。
- 前記L−アミノ酸がL−グルタミン酸であり、L−アミノ酸生合成系酵素が、グルタミン酸デヒドロゲナーゼ、グルタミンシンテターゼ、グルタミン酸シンターゼ、イソクエン酸デヒドロゲナーゼ、アコニット酸ヒドラターゼ、クエン酸シンターゼ、ホスホエノールピルビン酸カルボキシラーゼ、ピルビン酸カルボキシラーゼ、ピルビン酸デヒドロゲナーゼ、ピルビン酸キナーゼ、ホスホエノールピルビン酸シンターゼ、6−ホスホグルコン酸デヒドラターゼ、及び、2−ケト−3−デオキシ−6−ホスホグルコン酸アルドラーゼからなる群より選択される1種または2種以上の酵素である請求項8に記載の方法。
Priority Applications (5)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2006025620A JP2009095237A (ja) | 2006-02-02 | 2006-02-02 | L−アミノ酸の製造法 |
| PCT/JP2007/051815 WO2007088977A1 (ja) | 2006-02-02 | 2007-02-02 | L-アミノ酸の製造法 |
| EP07707964.8A EP1990416B1 (en) | 2006-02-02 | 2007-02-02 | Method for production of l-amino acid |
| US12/184,637 US8394612B2 (en) | 2006-02-02 | 2008-08-01 | Method for production of an L-amino acid |
| RU2008133840/10A RU2405040C2 (ru) | 2006-02-02 | 2008-08-19 | Способ получения l-аминокислоты |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JP2006025620A JP2009095237A (ja) | 2006-02-02 | 2006-02-02 | L−アミノ酸の製造法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| JP2009095237A true JP2009095237A (ja) | 2009-05-07 |
Family
ID=38327545
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2006025620A Pending JP2009095237A (ja) | 2006-02-02 | 2006-02-02 | L−アミノ酸の製造法 |
Country Status (5)
| Country | Link |
|---|---|
| US (1) | US8394612B2 (ja) |
| EP (1) | EP1990416B1 (ja) |
| JP (1) | JP2009095237A (ja) |
| RU (1) | RU2405040C2 (ja) |
| WO (1) | WO2007088977A1 (ja) |
Families Citing this family (12)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7915018B2 (en) * | 2004-10-22 | 2011-03-29 | Ajinomoto Co., Inc. | Method for producing L-amino acids using bacteria of the Enterobacteriaceae family |
| RU2396336C2 (ru) | 2007-09-27 | 2010-08-10 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) | СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia |
| US8692070B2 (en) | 2009-11-04 | 2014-04-08 | Iowa Corn Promotion Board | Plants with improved nitrogen utilization and stress tolerance |
| CN102181503A (zh) * | 2011-04-15 | 2011-09-14 | 江苏汉光生物工程有限公司 | 一种发酵生产l-苯丙氨酸的方法 |
| WO2014159850A2 (en) * | 2013-03-14 | 2014-10-02 | Codexis, Inc. | Low-phosphate repressible promoter |
| RU2014105547A (ru) | 2014-02-14 | 2015-08-20 | Адзиномото Ко., Инк. | СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА ENTEROBACTERIACEAE, ИМЕЮЩЕЙ СВЕРХЭКСПРЕССИРУЕМЫЙ ГЕН yajL |
| WO2017127567A1 (en) * | 2016-01-19 | 2017-07-27 | Genebiologics, Llc | Production of arginine-rich proteins and use as a fertilizer and germination enhancer |
| WO2019030808A1 (ja) * | 2017-08-07 | 2019-02-14 | 地方独立行政法人大阪産業技術研究所 | 芳香族化合物を産生する微生物 |
| KR102016050B1 (ko) * | 2018-03-20 | 2019-08-29 | 씨제이제일제당 주식회사 | 신규한 프로모터 및 이의 용도 |
| US12577572B2 (en) | 2019-10-23 | 2026-03-17 | Duke University | Compositions, systems, and methods for high level expression of recombinant protein |
| CN116555134A (zh) * | 2022-01-29 | 2023-08-08 | 廊坊梅花生物技术开发有限公司 | 产苏氨酸菌株的构建方法 |
| CN121022709B (zh) * | 2025-10-29 | 2026-02-03 | 天津科技大学 | 一种l-异亮氨酸生产菌株及其构建方法与应用 |
Family Cites Families (28)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| DE69124939T2 (de) | 1990-11-30 | 1997-10-02 | Ajinomoto Kk | Rekombinante DNS-Sequenzen kodierend für Enzyme frei von Feedback Inhibition, Plasmide diese Sequenzen enthaltend, transformierte Mikroorganismen nützlich für Produktion von aromatischen Aminosäuren, und deren Verfahren zur Herstellung durch Fermentation |
| JP3185261B2 (ja) * | 1991-02-05 | 2001-07-09 | 味の素株式会社 | 発酵法による芳香族アミノ酸の製造法 |
| CZ285453B6 (cs) | 1992-11-10 | 1999-08-11 | Ajinomoto Co.,Inc. | DNA, která obsahuje gen kódující aspartokinézu III, mikroorganizmus Escherichia coli a způsob výroby L-threoninu fermentací |
| SK283058B6 (sk) | 1993-10-28 | 2003-02-04 | Ajinomoto Co., Inc. | Spôsob produkcie cieľovej látky použitím mikroorganizmov |
| JPH07155184A (ja) | 1993-12-08 | 1995-06-20 | Ajinomoto Co Inc | 発酵法によるl−リジンの製造法 |
| KR100420743B1 (ko) | 1994-12-09 | 2004-05-24 | 아지노모토 가부시키가이샤 | 신규한리신데카복실라제유전자및l-리신의제조방법 |
| EP0796916A1 (en) | 1996-03-22 | 1997-09-24 | Triple-A B.V. | Improvement of amino acid fermentation processes |
| US5763230A (en) * | 1996-03-22 | 1998-06-09 | Triple-A B.V. P/A Produkschap Voor Veevoedor | Amino acid fermentation processes |
| JP4035855B2 (ja) * | 1996-06-05 | 2008-01-23 | 味の素株式会社 | L−リジンの製造法 |
| US5939307A (en) * | 1996-07-30 | 1999-08-17 | The Archer-Daniels-Midland Company | Strains of Escherichia coli, methods of preparing the same and use thereof in fermentation processes for l-threonine production |
| JP4088982B2 (ja) * | 1996-10-15 | 2008-05-21 | 味の素株式会社 | 発酵法によるl−アミノ酸の製造法 |
| DE19931317A1 (de) * | 1999-07-07 | 2001-01-11 | Degussa | L-Lysin produzierende coryneforme Bakterien und Verfahren zur Herstellung von L-Lysin |
| ES2313878T3 (es) | 2000-01-21 | 2009-03-16 | Ajinomoto Co., Inc. | Procedimiento para la produccion de l-lisina. |
| RU2212447C2 (ru) * | 2000-04-26 | 2003-09-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Штамм escherichia coli - продуцент аминокислоты (варианты) и способ получения аминокислот (варианты) |
| JP4380029B2 (ja) * | 2000-07-05 | 2009-12-09 | 味の素株式会社 | 微生物を利用した物質の製造法 |
| JP2002153285A (ja) | 2000-11-20 | 2002-05-28 | Tosoh Corp | 新規耐熱性アミノトランスフェラーゼ |
| EP1217076B1 (en) * | 2000-12-22 | 2006-06-14 | Ajinomoto Co., Inc. | Method of producing a target substance by fermentation |
| WO2003054140A2 (en) * | 2001-07-30 | 2003-07-03 | Genencor International, Inc. | Phosphate limited inducible promoter and a bacillus expression system |
| RU2229513C2 (ru) * | 2001-11-23 | 2004-05-27 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" | Способ получения l-аминокислот, штамм escherichia coli - продуцент l-аминокислоты (варианты) |
| AU2003205041A1 (en) * | 2002-07-12 | 2004-01-29 | Ajinomoto Co., Inc. | Method for producing target substance by fermentation |
| BR0304860A (pt) * | 2002-11-11 | 2004-08-31 | Ajinomoto Kk | Método para produzir uma substância alvo pela utilização de uma-bactéria pertencente ao gênero escherichia |
| BRPI0407966A (pt) * | 2003-03-12 | 2006-03-07 | Ajinomoto Kk | método para obter uma bactéria, bactéria produzindo l-aminoácido, e, método para produzir um l-aminoácido |
| JP2005021154A (ja) * | 2003-06-11 | 2005-01-27 | Ajinomoto Co Inc | L−アミノ酸の製造法 |
| WO2005010175A1 (en) * | 2003-07-29 | 2005-02-03 | Ajinomoto Co., Inc. | Method for producing l-lysine or l-threonine using escherichia bacteria having attnuated malic enzyme activity |
| BRPI0506369B1 (pt) * | 2004-01-30 | 2020-11-03 | Ajinomoto Co., Inc | microorganismo transgênico, e, método para produzir um l-aminoácido |
| US20050191684A1 (en) * | 2004-02-25 | 2005-09-01 | Zimenkov Danila V. | Method for producing L-amino acids |
| BRPI0507233B8 (pt) | 2004-03-11 | 2021-05-25 | Genentech Inc | processo para a produção de um polipeptídeo heterólogo em e. coli |
| US7915018B2 (en) * | 2004-10-22 | 2011-03-29 | Ajinomoto Co., Inc. | Method for producing L-amino acids using bacteria of the Enterobacteriaceae family |
-
2006
- 2006-02-02 JP JP2006025620A patent/JP2009095237A/ja active Pending
-
2007
- 2007-02-02 WO PCT/JP2007/051815 patent/WO2007088977A1/ja not_active Ceased
- 2007-02-02 EP EP07707964.8A patent/EP1990416B1/en active Active
-
2008
- 2008-08-01 US US12/184,637 patent/US8394612B2/en active Active
- 2008-08-19 RU RU2008133840/10A patent/RU2405040C2/ru active
Also Published As
| Publication number | Publication date |
|---|---|
| WO2007088977A1 (ja) | 2007-08-09 |
| US8394612B2 (en) | 2013-03-12 |
| US20100184162A1 (en) | 2010-07-22 |
| RU2405040C2 (ru) | 2010-11-27 |
| EP1990416A4 (en) | 2009-11-04 |
| EP1990416A1 (en) | 2008-11-12 |
| RU2008133840A (ru) | 2010-02-27 |
| EP1990416B1 (en) | 2014-09-10 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8460903B2 (en) | Method for producing an L-amino acid using a bacterium of the Enterobacteriaceae family | |
| US8293505B2 (en) | L-amino acid-producing microorganism and a method for producing an L-amino acid | |
| US8394612B2 (en) | Method for production of an L-amino acid | |
| US20090209011A1 (en) | Method for Producing an L-Amino Acid Using a Bacterium of the Enterobacteriaceae Family With Enhanced Expression of the fucPIKUR Operon | |
| EP2460873A1 (en) | A method for producing an L-amino acid using a bacterium of the Enterobacteriaceae family with attenuated expression of any of the cynT, cynS, cynX or cynR genes or a combination thereof | |
| US20140147909A1 (en) | Method for Producing an L-Amino Acid Using a Bacterium of the Enterobacteriaceae Family | |
| US20090155861A1 (en) | Method for producing an l-amino acid using a bacterium of the enterobacteriaceae family | |
| US8691537B2 (en) | Method for producing an L-amino acid using a bacterium of the Enterobacteriaceae family with attenuated expression of the rcsA gene | |
| US8703446B2 (en) | Method for producing an L-amino acid using a bacterium of the Enterobacteriaceae family | |
| JP2009060791A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| JP2010263789A (ja) | L−アミノ酸生産菌及びl−アミノ酸の製造法 | |
| WO2012002486A1 (ja) | L-アミノ酸の製造法 | |
| CA2595501A1 (en) | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family having a pathway of glycogen biosynthesis disrupted | |
| WO2008032757A1 (en) | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family with enhanced expression of the alsabc operon | |
| EP1856243B1 (en) | Process for producing an l-amino acid employing a bacterium of the enterobacteriaceae family with attenuated leuo expression | |
| WO2007119576A1 (en) | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family | |
| WO2010101053A1 (ja) | L-アミノ酸の製造法 | |
| WO2008004682A1 (en) | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family with attenuated expression of the yrah-r cluster | |
| WO2007083788A1 (en) | A method for producing an l-amino acid using a bacterium of enterobacteriaceae family with attenuated expression of the lrha gene | |
| WO2007083789A1 (en) | A method for producing an l-amino acid using a bacterium of enterobacteriaceae family with attenuated expression of the yfeh gene | |
| WO2007139220A1 (en) | A method for producing an l-amino acid using a bacterium of the enterobacteriaceae family with attenuated expression of the yehabcde cluster | |
| HUE025593T2 (en) | Method for L-Amino Acid Production by Enterobacteriaceae Bacteria by Reducing FimZ GENE Production |