JP2010507367A - モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 - Google Patents
モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 Download PDFInfo
- Publication number
- JP2010507367A JP2010507367A JP2009533566A JP2009533566A JP2010507367A JP 2010507367 A JP2010507367 A JP 2010507367A JP 2009533566 A JP2009533566 A JP 2009533566A JP 2009533566 A JP2009533566 A JP 2009533566A JP 2010507367 A JP2010507367 A JP 2010507367A
- Authority
- JP
- Japan
- Prior art keywords
- amino acid
- racemase
- seq
- monatin
- aminotransferase
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Pending
Links
- RMLYXMMBIZLGAQ-UHFFFAOYSA-N NC(CC(Cc1c[nH]c2ccccc12)(C(O)=O)O)C(O)=O Chemical compound NC(CC(Cc1c[nH]c2ccccc12)(C(O)=O)O)C(O)=O RMLYXMMBIZLGAQ-UHFFFAOYSA-N 0.000 description 1
Images











Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/1096—Transferases (2.) transferring nitrogenous groups (2.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P17/00—Preparation of heterocyclic carbon compounds with only O, N, S, Se or Te as ring hetero atoms
- C12P17/10—Nitrogen as only ring hetero atom
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Zoology (AREA)
- Engineering & Computer Science (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Microbiology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Biomedical Technology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Enzymes And Modification Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Indole Compounds (AREA)
- Seasonings (AREA)
Abstract
【化1】
Description
本出願は、2005年4月26日に出願された米国特許仮出願第60/674,932号の利益を主張する2006年4月26日に出願された米国特許出願第11/411,229号の一部継続出願である2006年10月20日に出願された米国特許出願第11/584,016号の一部継続出願である。

用語および方法の以下の説明は、本開示をより十分に記載し、かつ本開示の実施において当業者を誘導するために提供される。本明細書に用いた「含む(including)」は、「含む(comprising)」を意味する。用語「〜を含む」が用いるところはいかなるところでも、「〜を含むが、限定されない」と意味されることが、「限定されない」が明示的に述べられているまたはいないのいずれにせよ理解される。さらに、単数形「1つの(a)」または「1つの(an)」または「その(the)」は、文脈が明らかに指定しない限り、複数の言及を含む。例えば、「タンパク質を含む」についての言及は、1つまたは複数の上記のタンパク質を含み、「細胞を含む」についての言及は、1つまたは複数の細胞および当業者に知られている等価物についての言及を含む等とする。用語「約」は、いかなる測定においても起こる実験誤差の範囲を包含する。特に述べられていない限り、測定数はすべて、「約」という語が明確に用いられていなくとも、それらの前に「約」という語を有するとみなされる。
式中、Ra、Rb、Rc、Rd、およびReは、それぞれ独立して、水素原子、水酸基、C1〜C3アルキル基、C1〜C3アルコキシ基、アミノ基、またはヨウ素原子、臭素原子、塩素原子、もしくはフッ素原子のごときハロゲン原子から選択される任意の置換基を表す。しかしながら、Ra、Rb、Rc、Rd、およびReは、同時にすべて水素とすることはできない。あるいは、RbおよびRcならびに/またはRdおよびReは、それぞれ、C1〜C4アルキレン基を共に形成してもよい。
とりわけ国際公開第03/091396A2号(例えば、図1〜3および11〜13参照)に記載のように、モナチンは、生物学的変換(すなわち、基質から生成物への反応をポリペプチドで促進すること)を含む複数工程の経路によってトリプトファンから生成できる。記載した経路は、トリプトファンをインドール−3−ピルビン酸に生物学的に変換し、インドール−3−ピルビン酸を2−ヒドロキシ2−(インドール−3−イルメチル)−4−ケトグルタル酸(「MP」)に生物学的に変換し、MPをモナチンに生物学的に変換することを含む。モナチンを生成するために用いる本発明の生合成の経路は、1つまたは複数の下記に記載される工程、メカニズム、および/または経路を含んでいてもよいまたは本質的にそれからなってもよい。下記に記載される工程、メカニズム、および/または経路は、単に例示的であることが意図される。
・tnaT遺伝子を発現または過剰発現するように微生物を遺伝子操作する等、微生物によるトリプトファンの取り込みを増加させる、
・(i)L−アミノ酸の最小量のみを含有する培地中で微生物を成長させること、(ii)フェニルアラニンパーミアーゼタンパク質を改変すること、および/または(iii)一般的な芳香族アミノ酸パーミアーゼタンパク質の活性を付与する、配列の少なくともその部分およびフェニルアラニンパーミアーゼタンパク質の活性を付与する、配列の少なくともその部分を有するキメラタンパク質を作り出すこと等によって、微生物によるD−トリプトファンの取り込みを増加させる(Cosgriff AJ、Brasier G、Pi J、Dogovski C、Sarsero JP、Pittard AJ.「A study of AroP−PheP chimeric proteins and identification of a residue involved in tryptophan transport」、J Bacteriol.、182(8):2207〜17頁(2000年))、
・微生物によるインドール−3−ピルビン酸の分泌を阻止または低減する、
・微生物によるインドール−3−ピルビン酸の取り込みを増加させる、
・利用可能なインドール−3−ピルビン酸が微生物内で分解されるのを阻止または低減する、
・(i)デアシラーゼが欠乏していない微生物でtRNAtrpを過剰生成すること、(ii)適切なD−アミノ酸デアシラーゼを発現または過剰発現するように微生物を遺伝子操作すること、(iii)tRNAtrpを発現または過剰発現するようにおよび適切なD−アミノ酸デアシラーゼを発現または過剰発現するように微生物を遺伝子操作すること、(iv)遺伝子操作された微生物のバイオマスを生成し、次いでモナチンオペロンを誘導すること、(v)栄養素要求株を補完するためにフェニルアラニン、チロシン、およびイソロイシンに富むペプチドもしくはタンパク質を用いること、(vi)一般的な芳香族アミノ酸パーミアーゼを発現または過剰発現するように微生物を遺伝子操作すること、(vii)D−トリプトファン以外のD−アミノ酸を用いないもしくはその使用を最小化すること、(viii)外部からの基質としてL−トリプトファンを提供し、L−トリプトファンをD−トリプトファンに変換するラセマーゼを発現または過剰発現するように微生物を遺伝子操作すること、および/または(ix)D−トリプトファンの取り込みの阻害が起こる閾値未満のレベルでD−トリプトファンを提供すること等によって、D−トリプトファンの毒性を緩和する
ように改変された微生物とすることができる。
実施例
本実施例は、モナチン、モナチン前駆体(「MP」)、トリプトファン、アスパラギン酸、アラニン、およびグルタミン酸の存在を検出するために用いる方法を記載する。本実施例は、さらに、モナチンの4種の立体異性体の分離および検出のための方法を記載する。
in vitroまたはin vivo生化学反応から誘導されたモナチンおよびトリプトファンについての、混合物の分析は、クロマトグラフおよびMicromass社製Quattro Ultima三連四重極質量分析計の間に直列で設置されたWaters社製996フォトダイオードアレイ(PDA)吸光度モニターと共にWaters社製2795液体クロマトグラフを含むWaters/Micromass社製液体クロマトグラフィータンデム質量分析(LC/MS/MS)機器を用いて行った。LC分離は、40℃でXterra MS C8逆相クロマトグラフィーカラム、2.1mm×250mmを用いて行った。LC移動相は、A)(i)0.05%(容量/容量)トリフルオロ酢酸または(ii)0.3%ギ酸および10mMギ酸アンモニウムのいずれかを含有する水ならびにB)(i)0.05%(容量/容量)トリフルオロ酢酸または(ii)0.3%ギ酸および10mMギ酸アンモニウムのいずれかを含有するメタノールからなるものとした。
高分解能MS分析は、Applied Biosystems−Perkin Elmer社製 Q−Starハイブリッド四重極/飛行時間型質量分析計を用いて行った。プロトン化モナチンの測定質量には、トリプトファンを内部質量検量標準物質として用いた。元素組成C14H17N2O5に基づいた、プロトン化モナチンの算出質量は、293.1137である。実施例2および3に記載される生体触媒プロセスを用いて生成されるモナチンは、293.1144の測定質量を示した。これは、100万分の2(「ppm」)未満の質量測定誤差であり、酵素的に生成されたモナチンの元素組成の決定的証拠を提供する。
in vitroおよびin vivo反応におけるモナチンの立体異性体分布の決定は、1−フルオロ−2−4−ジニトロフェニル−5−L−アラニンアミド(「FDAA」)を用いた誘導体化に続けて、逆相LC/MS/MS MRM測定によって達成した。
50μLの試料または標準物質および10μLの内部標準物質に、アセトン中FDAAの1%溶液を100μLまたは200μL追加した。それぞれ20μLまたは40μLの1.0M炭酸水素ナトリウムを追加し、混合物は、時々混合しながら40℃で1時間インキュベートした。試料は、取り出して、冷却し、20μLの2.0M HClを用いて中和した(緩衝生物学的混合物の中和をもたらすために、より多くのHClを必要としてもよい)。脱気完了後、試料は、LC/MS/MSによる分析の準備が整った。
分析は、上記に記載されるLC/MS/MS計測装置を用いて行った。モナチン(特にFDAA−モナチン)の4種の立体異性体すべてを分離できるLC分離は、40℃で、Phenomenex社製Luna 2.0×250mm(3μm)C18(2)逆相クロマトグラフィーカラムで行った。LC移動相は、A)0.05%(質量/容量)酢酸アンモニウムを含有する水およびB)アセトニトリルからなるものとした。溶出は、0〜2分間は13%Bで均一溶媒とし、2〜15分間は13%Bから30%Bまで直線的であり、15〜16分間は30%Bから80%Bまで直線的であり、16〜21分間は80%Bで均一溶媒とし、21〜22分間は80%Bから13%Bまで直線的であり、実行の間の再平衡化期間は8分間とした。流速は、0.23mL/分とし、PDA吸光度は、200nmから400nmまでモニターした。ESI−MSのパラメータはすべて、FDAA−モナチンの脱プロトン化分子イオン([M−H]−)の発生および特徴的なフラグメントイオンの生成に基づいて最適化し、かつ選択した。
in vitroおよびin vivo反応におけるグルタミン酸およびアラニンの決定のためのポストカラム蛍光検出を有する液体クロマトグラフィー(LC/OPA)は、Waters社製474走査蛍光検出器およびWaters社製ポストカラム反応モジュールと組み合わせたWaters社製2690LCシステムまたはその等価物で行った。モナチンおよびトリプトファンの半定量分析もまたこの方法を用いて行った。LC分離は、60℃で、相互作用ナトリウム装填イオン交換カラムで行った。移動相Aは、Pickering社製Na 328緩衝液(Pickering Laboratories社;マウンテンビュー、CA)とした。移動相Bは、Pickering社製Na 740緩衝液とした。勾配溶出は、試料マトリックスに依存して、0〜20分間は0%Bから100%Bまでとし、20〜36分間は100%Bで均一溶媒とし、36〜37分間は100%Bから0%Bまで直線的であり、実行の間の再平衡化期間は少なくとも5分間とした。移動相の流速は、0.5mL/分とした。OPAポストカラム誘導体化溶液の流速は、0.5mL/分とした。蛍光検出器設定は、EX338〜340nmおよびEm420〜425nmとした。ノルロイシンを分析のための内部標準物質として使用した。アミノ酸の同定は、精製標準物質についてのクロマトグラフィーの保持時間データに基づくものとした。
生化学反応実験からのリシン、アラニン、メチオニン、チロシン、ロイシン、フェニルアラニン、トリプトファン、グルタミン酸、およびアスパラギン酸のごときL−アミノ酸およびD−アミノ酸の混合物を含有する試料は、タンパク質を変性させるためにギ酸を用いて最初に処理した。次いで、試料は、LC/MS/MS分析前に遠心分離し、0.45μmナイロンシリンジフィルターで濾過した。L−アミノ酸およびD−アミノ酸の同定は保持時間および質量選択的検出に基づくものとした。LC分離は、Waters社製2690液体クロマトグラフィーシステムおよびASTEC社製2.1mm×250mm Chirobiotic TAGクロマトグラフィーカラムをカラム温度を45℃に設定して用いることによって達成した。LC移動相AおよびBは、それぞれ、0.25%酢酸およびメタノール中0.25%酢酸とした。均一溶媒溶出は、L異性体およびD異性体を分離するためのすべての方法に用いた。リシンは、80%移動相Aおよび20%Bを用いて溶出した。グルタミン酸、アラニン、およびメチオニンは、60%移動相Aおよび40%Bならびに0.25mL/分の流速で溶出して分離した。アスパラギン酸、トリプトファン、チロシン、ロイシン、およびフェニルアラニンは、すべてについて、30%移動相Aおよび70%Bで異性体を分離し、フェニルアラニン以外は0.3mL/分の流速、フェニルアラニンは0.25mL/分の流速で実行した。
モナチンの生成
R,RモナチンおよびS,Sモナチンのラセミ混合物は、米国特許第5128482号に記載されるように合成して生成した。
R−MPは、アミノ受容体としてピルビン酸ナトリウムを用いて、0.1Mリン酸カリウム緩衝液中のAT−103広範囲D−アミノトランスフェラーゼ(BioCatalytics社)を用い、R,Rモナチンのアミノ基転移によって生成した。S−MPは、アミノ受容体としてピルビン酸ナトリウムを用いて、0.1Mリン酸カリウム緩衝液中のAT−102L−アミノトランスフェラーゼ(BioCatalytics社)を用い、S,Sモナチンのアミノ基転移によって生成した。両反応は、約20時間、約8.0〜8.3のpHで30℃で行った。両化合物は、Rohm and Haas社製(フィラデルフィア、PA)疎水性樹脂(XAD(商標)1600)を用いた調整用スケールのHPLCを用いて、精製し、水中に溶出させた。90%を超える純度のモナチン前駆体を含有する試料を収集し、凍結乾燥した。
AT−103トランスアミナーゼは、BioCatalytics社(パサデナ、CA)から購入したトランスアミナーゼライブラリーの一部であり、またこの酵素は、C.テストステローニからのProAアルドラーゼを用いる共役反応におけるモナチンの生成について試験した。アルドラーゼは、国際公開第03/091396A2号に記載のように調製した。AT−103は、アミノ酸供与体としてD−アミノ酸(D−グルタミン酸、D−アスパラギン酸、またはD−アラニンのごとき)を必要とするバチルス種からの広域特異性D−トランスアミナーゼ(EC2.6.1.21)である。酵素および付加的な成分/基質を、100mMリン酸カリウム緩衝液pH7.5、100mMアミノ供与体、および0.1mMピリドキサール−5’−リン酸(「PLP」)を含有したキット中に提供された反応緩衝液に直接的に追加した。1mLの反応緩衝液に、以下のものを追加した:4mgインドール−3−ピルビン酸、20mgピルビン酸、細胞抽出物中に提供される約50μg ProA、1μL 2M MgCl2、および2mgアミノトランスフェラーゼ酵素(AT−103)。反応は二通り行った。反応は、緩やかに振盪させながら(100rpm)、30℃で一晩インキュベートした。試料は、実施例1に記載のように濾過し、逆相LC/MS/MS分析にかけた。結果は、約370μg/mLモナチンがAT−103酵素を用いて生成されたことを示した。結果は、クロマトグラフ分離の間に分離する2種の立体異性体プールのピーク面積に基づき、R,R/S,Sモナチンに対するS,R/R,Sモナチンの比を決定するためにさらに分析した。AT−103によって生成された全モナチンのうち、69%は、混合異性体と比較してR,R/S,Sモナチンであった。この酵素(AT−103)は、D−アミノ酸に対して広域特異性を有することで知られている、国際公開第03/091396A2号に記載されるバチルス・スブチリスDAT酵素に相同である。キラル分析を、実施例1に記載されるFDAA方法論を用いて行い、D−アミノトランスフェラーゼが、予想どおり、R,RモナチンおよびいくらかのS,Rモナチンを圧倒的に作製することを検証した。基質としてS,SモナチンまたはR,Rモナチンおよびα−ケトグルタル酸を用いたさらなるアミノ基転移実験は、予想どおり、BioCatalytics社製酵素が、炭素4のD−立体配置に高度に選択的であることを検証した。これらの実験では、グルタミン酸は、基質としてS,Sモナチンおよびケトグルタル酸を用いた反応において検出されなかった。
以下のものを反応混合物1mL当たりに追加した:約60μg C.テストステローニProAアルドラーゼ(国際公開第03/091396A2号に記載のように細胞抽出物において供給)、4mM MgCl2、50mM D−トリプトファン、0.5mg BioCatalytics社製D−アミノトランスフェラーゼ(AT−103)、100mMピルビン酸ナトリウム、100mMリン酸カリウム緩衝液pH7.5または100mM酢酸ナトリウム緩衝液pH8、0.05mM PLP、3mMリン酸カリウム(アセテート反応にのみ)、および10mM α−ケトグルタル酸。実験は、アルドラーゼを追加しないネガティブコントロールと共に二通り実行した。試料は、緩やかに振盪させながら、30℃で、一晩(20時間)インキュベートした。酢酸ナトリウム試料の実際のpHは約5とし、リン酸緩衝試料の最終pHは約7とした。アルドラーゼのどれも、pH5で著しい活性を有するようではなく、ProAアルドラーゼを含有する試料は、ネガティブコントロールをわずかに超えたが、おそらく実験誤差を超えなかった。リン酸カリウム中では、ProAアルドラーゼは、1.7:1のR,R:S,Rの比で73.4ppmのモナチンを生成した(D−トリプトファンから約63%R,R)。
AT−103トランスアミナーゼ(広域特異性D−アミノトランスフェラーゼ)をBioCatalytics社(パサデナ、CA)から購入し、この酵素または実施例18で生成したB.スファエリクス組換え酵素のいずれかを、HMGアルドラーゼとの共役反応に用いて、米国特許出願公開第2005282260号に記載のように、D−トリプトファンおよびピルビン酸からモナチンを生成した。
特異的活性を示す。
配列番号21のアルドラーゼ遺伝子は、Diversa社、サンディエゴ、カリフォルニアから得た。配列番号21は、アルドラーゼ遺伝子についてDiversa社によってスクリーニングされた環境ライブラリーの一部であった。しかしながら、配列番号21のアルドラーゼ遺伝子は、当業者に知られているいずれかの方法によって再構築されてもよい。例えば、配列番号21のアルドラーゼ遺伝子は、実施例10、18、および19に記載のように、アセンブリーPCR法を利用して再構築されてもよい。
確認したアルドラーゼクローンは、BL21(DE3)またはBL21(DE3)pLysSコンピテント細胞(Novagen社、マディソン、WI)のいずれかに形質転換した。適切な抗生物質を用いて成長させた一晩培養物を、新鮮な培地中に希釈し(通常1:100)、37℃で通気しながらOD600〜0.6まで成長させた。次いで、培養物は、1mM イソプロピルチオガラクトシド(「IPTG」)を用いて誘発し、30℃(通気有り)に移し、インキュベーションを一晩継続した。細胞は、遠心分離によって採取した。細胞ペレットは、細胞溶解を助けるために、通常、1回の凍結解凍サイクルにかけた。細胞ペレットは、BugBusterおよびBenzonase(Novagen社、マディソン、WI)中で溶解させた(メーカーのプロトコールに従って)。細胞片は遠心分離によって除去した。粗タンパク質抽出物は、メーカーのプロトコールに従って調製したHisBindカラム(Novagen社、マディソン、WI)にかけた。カラムは洗浄し、タンパク質はメーカーのプロトコールに従って溶出した。精製タンパク質は、PD−10カラム(GE Healthcare社、ピスカタウェイ、NJ)を用いて脱塩した。交換に用いた緩衝液は、50mMリン酸カリウムpH7.5、100mM NaCl、4mM MgCl2とした。精製タンパク質は、Amicon社製遠心濃縮器(Millipore社、ビレリカ、MA)を用いて濃縮した。
配列番号104のアルドラーゼをコードする遺伝子(遺伝子のDNA配列は配列番号103として示される)を精製を考慮するためにN末端Hisタグを有するpET28b発現ベクター(EMD Biosciences/Novagen、Madison、WI)の中にサブクローニングした。
5'-ATAAGACATATGCCTATCGTTGTTACGAAG-3'(Nde I制限部位)(配列番号105)および
5'-ATAAGAGGATCCTTATTCCTCGGGCAGCCGCTC-3'(BamH I制限部位)(配列番号106)。
確認したアルドラーゼクローンを、BL21(DE3)pLysSコンピテント細胞(Novagen Madison、WI)中に形質転換した。配列番号22のアルドラーゼもまた活性の比較のために同時に調製した。誘発は、50μg/mLカナマイシンを用いて30℃でTerrific Broth(TartoffおよびHobbs、Bethesda Research Laboratories Focus 9:12(1987年)中で一晩行った。適切な抗生物質を用いて成長させた一晩培養物を、新鮮な培地中に希釈し(通常1:100)、37℃で通気しながらOD600〜0.6まで成長させた。次いで、培養は、1mM IPTGを用いて誘発し、30℃(通気有り)に移し、インキュベーションを一晩継続した。細胞は、遠心分離によって採取した。細胞ペレットは、細胞溶解を助けるために、通常、1回の凍結解凍サイクルにかけた。細胞ペレットは、BugBusterおよびBenzonase Nuclease(EMD Biosciences/Novagen、Madison、WI)中で溶解させた(メーカーのプロトコールに従って)。細胞片は遠心分離によって除去した。粗タンパク質抽出物は、メーカーのプロトコールに従って調製した10mg容量HIS−Bindカラム(EMD Biosciences/Novagen、Madison、WI)にかけた。カラムは洗浄し、タンパク質はメーカーのプロトコールに従って溶出した。精製タンパク質は、PD−10カラム(GE Healthcare、Piscataway、NJ)を用いて脱塩し、4mM MgCl2および200mM NaClを含有する50mMリン酸カリウム緩衝液、pH7.5中に溶出した。精製タンパク質は、Amicon遠心濃縮器(5000 MWカットオフ)(Millipore、Billerica、MA)を用いて濃縮させた。濃縮の後、配列番号104のアルドラーゼのうちのいくつかが沈殿したことに注目した。しかしながら、その沈殿は、酵素の活性のレベルに影響するようには考えられなかった。アッセイを行うまで、タンパク質は−80℃で保存した。タンパク質アッセイは、Pierce BCAキット(Rockford、IL)およびタンパク質標準物質としてウシ血清アルブミン(「BSA」)を用いるマイクロタイタープレートプロトコールを用いて行った。Experion Pro260電気泳動システム(Bio−Rad、Hercules、CA)は、精製試料中のアルドラーゼの百分率を算出するためにならびに可溶性細胞抽出物におけるおよび全タンパク質における発現レベルを評価するために用いた。
精製アルドラーゼは、D−トリプトファンからR,Rモナチンを生成するそれらの能力について試験した。アッセイは、アッセイ当たり同じ濃度の酵素(50μg/mL)を用いて、精製タンパク質を用いて、微量遠心チューブ中で二通り行った。2mg/mLのBiocatalytics AT−103をD−アミノトランスフェラーゼとして用いた。以下のものを反応混合物1mL当たりに追加した:アルドラーゼ、4mM MgCl2、50mM D−トリプトファン、D−アミノトランスフェラーゼ、200mMピルビン酸ナトリウム、100mMリン酸カリウム緩衝液pH7.5、および0.05mM PLP。試料は、振盪させながら30℃でインキュベートした。30分間、1時間、3時間、および一晩(19時間)の試料を測った。表9は、逆相ピーク面積によって決定されるように、各時点で生成された全モナチンの平均した結果および生成されたR,Rモナチン%を示す。実施例1に記載されるさらなるFDAA誘導体化LC/MS/MS分析を反応のうちのいくつかについて行った、また括弧中に示す。
atgcctatcg ttgttacgaa gatcgaccga cccagcgcgg cggacgtcga
aaggatcgcc gcctatggtg tcgcgacctt gcatgaagcg caaggacgaa
ccgggttgat ggcgtccaat atgcgcccaa tctatcgccc tgcgcacatt
gccgggcccg cggtgacctg ccttgtggcg cctggcgaca attggatgat
ccatgtcgcc gtcgaacagt gccagccggg agatgtcctg gtcgtggtac
cgaccagccc ctgcgaagac ggctatttcg gcgatctgct ggcgacctcg
ctgcggtcgc gcggggtcaa aggtctgatc atcgaggccg gcgtacgcga
tatcgcgaca ttgaccgaga tgaaattccc ggtctggtcc aaggcggtgt
tcgcgcaagg aacggtcaag gagaccatcg ccagcgtcaa tgtgcccctc
gtctgcgcgg gcgcccgcat cgtgccgggc gatctgatcg ttgccgacga
cgacggggtc gtcgtgattc caagacgttc cgttccggcg gtcctttcca
gcgccgaggc ccgcgaagag aaggaagccc gcaaccgcgc ccgcttcgaa
gctggcgagc tgggcctcga cgtctacaac atgcgccagc gcctggccga
caagggcttg cgctatgtcg agcggctgcc cgaggaatag(配列番号103)。
Met Pro Ile Val Val Thr Lys Ile Asp Arg Pro Ser Ala Ala Asp Val Glu Arg Ile Ala Ala Tyr Gly Val Ala Thr Leu His Glu Ala Gln Gly Arg Thr Gly Leu Met Ala Ser Asn Met Arg Pro Ile Tyr Arg Pro Ala His Ile Ala Gly Pro Ala Val Thr Cys Leu Val Ala Pro Gly Asp Asn Trp Met Ile His Val Ala Val Glu Gln Cys Gln Pro Gly Asp Val Leu Val Val Val Pro Thr Ser Pro Cys Glu Asp Gly Tyr Phe Gly Asp Leu Leu Ala Thr Ser Leu Arg Ser Arg Gly Val Lys Gly Leu Ile Ile Glu Ala Gly Val Arg Asp Ile Ala Thr Leu Thr Glu Met Lys Phe Pro Val Trp Ser Lys Ala Val Phe Ala Gln Gly Thr Val Lys Glu Thr Ile Ala Ser Val Asn Val Pro Leu Val Cys Ala Gly Ala Arg Ile Val Pro Gly Asp Leu Ile Val Ala Asp Asp Asp Gly Val Val Val Ile Pro Arg Arg Ser Val Pro Ala Val Leu Ser Ser Ala Glu Ala Arg Glu Glu Lys Glu Ala Arg Asn Arg Ala Arg Phe Glu Ala Gly Glu Leu Gly Leu Asp Val Tyr Asn Met Arg Gln Arg Leu Ala Asp Lys Gly Leu Arg Tyr Val Glu Arg Leu Pro Glu Glu(配列番号104)。
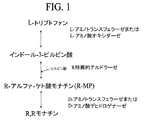
R,R−モナチンは、D−トリプトファンを出発物質として用いた場合に、D−アミノトランスフェラーゼおよびアルドラーゼを用いて生成した(実施例3)。それにもかかわらず、L−トリプトファンはいくつかの理由で好ましい出発物質となる可能性がある。例えば、L−トリプトファンは、D−トリプトファンよりも高価ではなく、容易に入手可能である可能性がある。本開示は、活性トリプトファンラセマーゼを得るための数種の方法を記載する。R,Rモナチンの収量は、R特異的アルドラーゼ、すなわち、R−MPを優先的にまたは選択的に生成するアルドラーゼを用いることより改善される。図1および2は、トリプトファンラセマーゼ、D−アミノトランスフェラーゼ、およびR特異的アルドラーゼを用いて、L−トリプトファンから立体異性的に豊富なR,Rモナチンを生成するための方法を示す。
ライブラリーの構築:
バークホルデリア・ピロシナ(Burkholderia pyrrocina)(ATCC15958)およびシュードモナス・クロロラフィス(ATCC15926)をAmerican Type Culture Collection、P.O.Box1549、マナサス、VA、20108、USAから得た。それらは、ATCCによって推奨されるように成長させ、ゲノムDNAは、Mekalanos,J.J.,「Duplication and amplification of toxin genes in Vibrio cholerae」,Cell 35:253−263,(1983)に記載される方法に従って調製した。ゲノムDNAは、Sau3AI制限酵素を用いて部分的に消化した。1〜3Kbp断片を、Qiagen社製QIAquick(登録商標)Gel Extraction Kit(Qiagen社、バレンシア、CA)を用いてゲル精製した。精製DNAは、上記のようにBamHIを用いて消化し、かつ精製したpTrc99a(Amersham社、ピスカタウェイ、NJ)にライゲーションした。ライゲーションは、ベクターに対する挿入断片の3:1のモル比を用いて、一晩のインキュベーションにより室温で行った。ライゲーションライブラリーは、TOP10F’化学的コンピテント細胞(Invitrogen社、カールスバード、CA)中に形質転換し、100μg/mlアンピシリンを用いてLB培地上で平板培養した。形質転換平板培養物の一晩のインキュベーションの後、コロニーを平板からこすり取り、液体LB培地で洗浄した。適切なサイズの細胞ペレットを、Qiagen社製QIAquick(登録商標)ミニプレップキット(Qiagen社、バレンシア、CA)を用いてミニプレップした。約30,000コロニーをプールし、ミニプレップした。
トリプトファンラセマーゼを可能性として有するとして同定されたクローンを、BL21のごとき組換えタンパク質の発現に一般的に用いられるイー・コリの株に形質転換する。細胞は、0.4〜0.6の600nmの光学濃度までLBブロス中で成長させる。ラセマーゼの発現を駆動するプロモーターは、IPTG(イソプロピル−β−D−チオガラクトピラノシド)(0.1mM最終濃度)を用いて誘発する。誘発の後、細胞に、37℃(通気有り)で1〜3時間タンパク質を発現させる。細胞は、採取し、フレンチプレス、超音波処理によって、または化学的手段(BugBuster(Novagen社、マディソン、WI)のごとき)によって溶解させる。溶解させた細胞は、細胞片を除去するために遠心分離する。清澄抽出物をアッセイに直接的に用いる。
トリプトファンラセマーゼは、硫安分画を含めた従来のタンパク質精製技術および従来のカラムクロマトグラフイーによって、植物源または微生物源のいずれかから精製できる。スポットが2次元ゲル上で単離できるようにタンパク質が精製されると、ペプチドマイクロシークエンシング技術または従来のEdman型アミノ酸配列決定が利用される(この種の作業に通常用いられるプロトコールおよび設備の記載について「golgi.harvard.edu/microchem/」をインターネットで参照されたい)。しかしながら、いくつかの場合では、生物のゲノム配列は、タンパク質精製のためのタンパク質源として使用できない、というのは、上記配列がまだ決定されていないからである。そういった状況では、一次縮重プライマーを、最も近い、知られている類縁のタンパク質源からの入手可能な配列に基づき設計してもよい。次いで、縮重PCRおよびゲノムウォーキングを、トリプトファンラセマーゼコード配列を単離するために確立されたプロトコールに従って行う。
ゲオバチルス・ステアロテルモフィルスからのアラニンラセマーゼ(配列番号41)をクローニングした。G.ステアロテルモフィルス(ATCC12980D)からのゲノムDNAはATCC(マナッサス、VA)から購入した。以下のプライマーは、G.ステアロテルモフィルスからのアラニンラセマーゼ遺伝子を増幅するために用いた:5'-atggacgagtttcaccgcga-3'(配列番号25)および5'-ttatgcatcgcttcatccgc-3'(配列番号26)。PCR生成物は、Zero Blunt TOPO PCRクローニングキット(Invitrogen社、カールスバード、CA)を用いてpCR−Blunt−TOPOにライゲーションした。適正なクローンは配列決定によって確認した(Agencourt社、ビバリー、MA)。適正なクローンは、引き続いてのPCR反応における鋳型として用いた。
5'-gccggacgacacgcacattnnkgcggtcgtgaaggcgaacgcc-3'(配列番号29);
5'-gtgaaggcgaacgcctatggannkggggatgtgcaggtggcaagg-3'(配列番号30);
5'-cctcccgcctggcggttgccnnkttggatgaggcgctcgctttaa-3'(配列番号31);
5'-caaccaggcgaaaaggtgagcnnkggtgcgacgtacactgcgcag-3'(配列番号32);
5'-gatcgggacgattccgatcggcnnkgcggacggctggctccgccg-3'(配列番号33);および
5'-gccatttggaaacgatcaacnnkgaagtgccttgcacgatcag-3'(配列番号34)
(n=任意のヌクレオチドおよびk=gまたはt)。
pET30Trpラセマーゼ構築物を有する一晩培養物は、適切な抗生物質(50μg/mlカナマイシンおよび20μg/mlクロラムフェニコール)を有する新鮮なLB培地中に継代培養させ、OD600を約0.6まで成長させた(通気有りで37℃)。発現を100μM IPTGを用いて誘発し、インキュベーションは、2時間通気有りで37℃で継続させた。細胞は、遠心分離によって採取し、使用まで−80℃で保存した。細胞ペレットは、氷上で解凍し、細胞は、BugBuster Primary Amine Free Cell Lysis ReagentおよびBenzonase Nuclease(Novagen社、マディソン、WI)を用いて溶解した。細胞片は、遠心分離によって除去し、上清は、粗タンパク質抽出物として用いた。粗タンパク質抽出物は、0.45μmシリンジフィルターを用いて濾過し、メーカーの指示に従って、あらかじめ平衡化したHisBindカラム(Novagen社、マディソン、WI)にかけた。カラムは洗浄し、タンパク質はメーカーのプロトコールに指示されるように溶出した。精製タンパク質は、溶出液として50mMリン酸カリウムpH8.0、10μMピリドキサール−5’−リン酸(「PLP」))を用いて、PD−10カラム(GE Healthcare社、ピスカタウェイ、NJ)を用いて脱塩した。脱塩したタンパク質は、Amicon社製遠心濃縮器(Millipore社、ビレリカ、MA)を用いて濃縮した。野生型のアラニンラセマーゼは、上記に記載のように精製した。
精製ラセマーゼは、数種のアッセイにおいて試験した。あるアッセイにおいて、D−アミノ酸オキシダーゼによる過酸化水素の生成を検出システムとして用いた。オキシダーゼについてのD−トリプトファン基質は、本実施例において記載のように単離されたラセマーゼ酵素を介してL−トリプトファンから生成された。アッセイは、アッセイ当たりに0、1、10、25、50、100、200μgの酵素、50mMリン酸カリウムpH8.0、10μM PLP、50mM L−トリプトファンを含んだ。アッセイは、37℃で1時間インキュベートした。インキュベーションの後、100mg/ml D−アミノ酸オキシダーゼ(AOD−101 BioCatalytics社、パサデナ、CA)および0.5mM FADを反応ミックスに追加した。過酸化水素の発生は、Amplex Red試薬キット(Molecular Probes社、ユージーン、OR)およびPerkin Elmer社製HTS 7000 Plus BioAssay Reader Fluorometer(ウェルズリー、MA)を用いて測定した。アッセイデータを下記の表12および13に要約する。
以下のものを反応混合物1mL当たりに追加した:配列番号22の約50μgアルドラーゼ、16mg/mL精製トリプトファンラセマーゼ、4mM MgCl2、50mM L−トリプトファン、0.5mgD−アミノトランスフェラーゼ(実施例14に記載のようにバチルス・スファエリクスから精製)、100mMピルビン酸ナトリウム、100mMリン酸カリウム緩衝液pH7.5、および0.05mM PLP。ピルビン酸は、広域特異性D−アミノトランスフェラーゼに対して許容し得るアミノ受容体であるので、α−ケトグルタル酸は使用しなかった。D−トリプトファンを開始基質とし、ラセマーゼを含まないコントロールを含んだ。試料は、緩やかに振盪させながら、30℃で、2時間または一晩(20時間)インキュベートした。試料は、実施例1に記載のように分析した。アッセイの結果を表18において下記に示す(nd=検出せず)。
突然変異誘発アラニンラセマーゼの数種の復帰変異体を作り出した。復帰変異体は、以下のプライマーを用いて、先に記載のように、QuikChange Multi Site−Directed Mutagenesis Kit(Stratagene社、ラ ホーヤ、CA)を用いて部位特異的突然変異誘発によって作製した。
5'-gccatttggaaacgatcaactatgaagtgccttgcacgatcag-3'(配列番号37);
5'-ctcccgcctggcggttgccttcttggatgaggcgctcgctttaag-3'(配列番号38);および
5'gccggacgacacgcacattatggcggtcgtgaaggcgaacgcc-3'(配列番号39)
突然変異リスト:
MF1:N41S(自然突然変異)、P197L、Y354A
MF2:F66E、P197L、Y354A
MY1:M35C、F66E、P197L
突然変異誘発ラセマーゼ:M35C、F66E、P197L、Y354A
実施例4AでpET30中にクローニングした野生型ジオバチルス ステアロサーモフィルスアラニンラセマーゼ(配列番号:41)を、部位特異的突然変異誘発のための鋳型として用い、Y354Aの変化を加えた。突然変異誘発は、QuickChange−Multi site−directed突然変異誘発キット(Stratagene、La Jolla、CA)を用いて行った。以下の突然変異誘発性プライマーは、Y354Aの変化を加えるために用いた、5’−gccatttggaaacgatcaacgcggaagtgccttgcacgatcag−3’(配列番号:107)。部位特異的突然変異誘発は、メーカーのプロトコールに記載されるように行った。数種の単離物は配列決定し(Agencourt、Beverly、MA)、適正な配列を有する単離物は、さらなる分析のために選択し、用いた。
数種の変異体を以下の通り単離した。プライマーは、位354で「NNK」ランダム突然変異誘発を進めるために作製した。プライマー、5’−gccatttggaaacgatcaacnnkgaagtgccttgcacgatcag−3’(配列番号108)は、QuickChange Multi Site Directed Mutagenesis Kit(Stratagene、La Jolla、CA)と共に用いた。反応に用いた鋳型は、pET30の中にクローニングした野生型G.ステアロサーモフィルスアラニンラセマーゼとした。突然変異誘発はメーカーのプロトコールに従って進めた。数種の変異体はこの方法を用いて単離した。以下の変異体はこの方法を用いて単離した:Y354D、Y354E、Y354L、Y354H、およびY354K。他のアミノ酸置換はQuickChange Multi Kitおよびこれらのプライマー:
Y354N 5’−ccatttggaaacgatcaacaacgaagtgccttgcacgatcag−3’(配列番号109)および
Y354G 5’−gccatttggaaacgatcaacggcgaagtgccttgcacgatcag−3’(配列番号110)を用いて作製した。
数種の変異体を以下の通り単離した。2つのプライマーは、鋳型としてY354A変異体(実施例4Bに記載)を用いて、アミノ酸位35および265での「NNK」ランダム突然変異誘発を進めるために作製した。Y254Aをコードする遺伝子は、NcoIおよびBamHI制限部位を用いて、pPRONcoベクターの中にサブクローニングし、この構築物を突然変異誘発反応に用いた。pPRONcoプラスミドは、NcoI部位のうちの一方を突然変異T1538Cによって除去した、pPROLar.A122ベクター(BD Biosciences Clontech、Palo Alto、CA)の誘導体である。以下のプライマー(5’−リン酸化)を、QuikChange Multi Site Directed Mutagenesis Kit(Stratagene、La Jolla、CA)と共に用いた:
Y265X:5'-CAA CCA GGC GAA AAG GTG AGC NNK GGT GCG ACG TAC ACT GCG CAG-3'(配列番号113)および
M35X:5'-GCC GGA CGA CAC GCA CAT TNN KGC GGT CGT GAA GGC GAA CGC C-3'(配列番号114)
NはG、C、T、またはAを示し、KはGまたはTを示す。
BAR(広域特異性アミノ酸ラセマーゼ)は、文献からの情報を用いて、P.プチダKT2440中で同定した。(Roise,D.、Soda,K.、Yagi,T.、Walsch,C.T.、Biochemistry 23、5195〜5201頁、(1984年))。P.プチダKT2440はまたATCC47054としても知られている。P.ストリアタからのBAR酵素の活性部位は、配列決定され、報告された(LTAVLKADAYGXGIGL(配列番号:116)、Xは任意の自然発生アミノ酸を示す)。この配列は、NCBIで入手可能なP.プチダKT2440ゲノム配列をBLASTするのに用いた。ほとんど同一のコンセンサス配列を有するタンパク質を同定した。プライマーは、American Type Culture Collection(ATCC、Manassas、VA)から得たゲノムDNAからの遺伝子をクローニングするために設計した、また、以下の通りとする。
5'-AGAAGACATATGCCCTTTCGCCGTAGGG-3'(配列番号117)および
5'-AGAAGAGGATCCTCAGTCGACGAGTATCTTCG-3')(配列番号118)。
atgccctttcgccgtacccttctggctgcatccctggcacttctgatcaccggacaggcccccctgtatgcggcaccaccgttgtcgatggacaacggcaccaacaccctgaccgtgcaaaacagcaatgcctgggtcgaagtcagcgccagcgccctgcagcacaacatccgcacgctgcaggccgagctggccggcaagtccaagctgtgcgccgtgctcaaggccgatgcctatggccacggtatcggcctggtaatgccatcgatcatcgcccaaggcgtgccctgcgtggcggtggccagcaacgaggaggcccgcgtggtccgcgccagtggcttcaccgggcaactggtgcgggtacgcctggccagcctcagcgagctggaagatggcttgcagtacgacatggaagagctggtgggcagcgcggaatttgcccgccaggccgatgccatcgccgcgcgccatggcaagaccttgcgcattcacatggcgctcaactccagcggcatgagccgcaacggggtggagatggccacctggtccggccgtggcgaagcgctgcagatcaccgaccagaagcacctcaagctggtcgcgctgatgacccacttcgccgtggaagacaaggacgatgtacgcaagggcctggcggcattcaacgagcagaccgactggttgatcaagcacgccaggctggaccgcagcaagctcaccctgcacgccgccaactcgttcgctacgctggaagtgccggaagcgcgcctggacatggtacgaacgggtggcgcgctgttcggcgacaccgtgccggcgcgcaccgagtacaaacgtgcgatgcagttcaaatcgcacgtggcggcggtgcacagctatccggccggcaacaccgtgggctatgaccgcaccttcaccctggcccgtgattcgcggctggccaacattacggtcgggtactccgatggctaccgccgggtattcaccaacaagggccatgtgctgatcaacggccaccgtgtgccggtcgtgggcaaggtgtcgatgaacacgctgatggtcgatgtcaccgacttccctgatgtgaaggggggtaacgaagtggtgctgttcggcaagcaggccgggggcgaaatcacccaggccgagatggaagaaatcaacggcgcgttgctcgccgatttgtacaccgtatggggcaattccaacccgaagatactcgtcgactga。
MPFRRTLLAASLALLITGQAPLYAAPPLSMDNGTNTLTVQNSNAWVEVSASALQHNIRTLQAELAGKSKLCAVLKADAYGHGIGLVMPSIIAQGVPCVAVASNEEARVVRASGFTGQLVRVRLASLSELEDGLQYDMEELVGSAEFARQADAIAARHGKTLRIHMALNSSGMSRNGVEMATWSGRGEALQITDQKHLKLVALMTHFAVEDKDDVRKGLAAFNEQTDWLIKHARLDRSKLTLHAANSFATLEVPEARLDMVRTGGALFGDTVPARTEYKRAMQFKSHVAAVHSYPAGNTVGYDRTFTLARDSRLANITVGYSDGYRRVFTNKGHVLINGHRVPVVGKVSMNTLMVDVTDFPDVKGGNEVVLFGKQAGGEITQAEMEEINGALLADLYTVWGNSNPKILVD。
上記に記載されるpET30 KT2440 BARプラスミドはBL21 DE3 pLysS(Invitrogen、Carlsbad、CA)中に形質転換した。結果として生じた株は、0.4〜0.6のOD600まで通気させながら37℃でLBまたはTerrific Broth中で成長させ、1mM IPTGを用いて誘発した。インキュベーションは、通気させながら37℃で3〜4時間継続した。細胞は、遠心分離によって採取し、細胞ペレットは使用まで−80℃で保存した。細胞ペレットは氷上で解凍した。細胞は、BugBusterおよびBenzonase(Novagen、Madison、WI)を用いて溶解した。細胞片は、遠心分離によって除去し、無細胞抽出物は直ちに用いるか、または−80℃で保存した。KT2440 BAR遺伝子は、さらに、pET28のNdeI−BamHI部位にクローニングし、BL21 DE3 pLysSコンピテント細胞中に形質転換した。この構築物は、可溶性タンパク質をあまり効率的に発現したとは考えられず、したがって、タグなしバージョン(pET30 KT2440 BAR)をその後の研究で用いた。
Amplex Redアッセイは本実施例に記載されるように設定した。P.プチダKT2440 BARは200μgで用いた(本実施例に記載されるように精製)。野生型G.ステアロサーモフィルスアラニンラセマーゼおよびY354Aは実施例4Bに記載されるように精製し、200μgまたは1000μgのいずれかで用いた。CEは、本実施例に記載されるように調製した無細胞抽出物である。60分の時点での結果を下記の表22に示す。全時間的経過を図11に示す。
1.P.プチダIFO12996、NBRC12996、P.プチダ12996、およびP.ストリアタとしても知られている(Nagata,Shinji;Esaki,Nobuyoshi;Tanizawa,Katsuyuki;Tanaka,Hidehiko;Soda,Kenji、Agricultural and Biological Chemistry(1985年)、49(4)、1137〜41頁)
2.P.プチダSCRC−744(Asano,Yasuhisa;Endo,Kaori、Applied Microbiology and Biotechnology(1988年)、29(6)、523〜7頁)
3.P.グラベオレンス(P.タエトロレンスとしても知られている)(Soda,Kenji;Yorifuji,Takamitsu;Ogata,Koichi、Journal of Biological Chemistry (1971年)、246(16)、5085〜92頁)
4.エロモナス・キャビエ、エロモナス・パンクタータ(Aeromonas punctata)亜種キャビエとしても知られている(Inagaki,Kenji;Tanizawa,Katsuyuki;Tanaka,Hidehiko;Soda,Kenji、Agricultural and Biological Chemistry(1987年)、51(1)、173〜80頁)
5.P.ストリアタAKU0813(Soda,Kenji;Osumi,Takaharu、Biochemical and Biophysical Research Communications(1969年)、35(3)、363〜8頁)
6.P.オーレオファシエンス(Salcher,Olga;Lingens,Franz、Journal of General Microbiology(1980年)、121(2)、465〜71頁)
7.P.プチダ12633(Wolf,Larissa B.;Sonke,Theo;Tjen,Kim C.M.F.;Kaptein,Bernard;Broxterman,Quirinus B.;Schoemaker,Hans E.;Rutjes,Floris P.J.T.、Advanced Synthesis & Catalysis(2001年)、343(6+7)、662〜674頁)
8.P.フルオレッセンス株(Ju,Jiansong;Yokoigawa,Kumio;Misono,Haruo;Ohnishi,Kouhei、Journal of Bioscience and Bioengineering(2005年)、100(4)、409〜417頁)
9.P.ミヤミズ(P.miyamizu)(Chibata,Ichiro;Tosa,Tetsuya;Sano,Ryujiro、Applied Microbiology(1965年)、13(4)、618〜24頁)
10.P.オレオボランス株(P.プチダとしても知られている)
11.シュードモナス株2150(Okazaki,Hiroshi、Agricultural and Biological Chemistry(1968年)、32(2)、254〜6頁)
モナチン生成アッセイは、精製P.プチダKT2440 BAR(実施例4Eで精製)(100μg)または精製Y354A(実施例4Bで精製)(500μg)、D−アミノトランスフェラーゼ(BioCatalytics AT−103(Pasadena、CA))(500μg)、および配列番号104のアルドラーゼ(実施例3B)(50μg)を用いて行った。上記の酵素に加えて、以下のものを1mLの反応混合物当たりに追加した:4mM MgCl2、50mM L−トリプトファン、100mMピルビン酸ナトリウム、100mMリン酸カリウム緩衝液pH7.5、および0.05mM PLP。コントロールとして、実験は、ラセマーゼなしでかつD−トリプトファンで始めて、行った。結果の概要を下記の表25に示す。
P.プチダKT2440(実施例4Eに記載)に由来するBAR酵素をコードする遺伝子を、Y396C突然変異を有する酵素を生成するために改変した。同様の突然変異は、「Tryptophan Racemase Derived from Broad Specificity Amino Acid Racemase by Directed Evolution」(Sato,M.ら、10th International Symposium on the Genetics of Industrial Microorganisms、Prague、June 24〜28 206)と題するポスターならびにK.Kino、M.Sato、M.Yoneyama、およびK.Kirimura、Appl Microbiol Biotechnol(2007年)73:1299〜1305頁に記載された。突然変異誘発は、タグなしタンパク質をもたらすpET30中のBAR遺伝子を用いて、QuickChange−Multi 部位特異的突然変異誘発キット(Stratagene、La Jolla、CA)を用いて行った。以下の突然変異誘発性プライマーはY396C変化を加えるために用いた。5'-TTGCTCGCCGATTTGTGCACCGTATGGGGCAATTC-3'(配列番号121)。
実験概要
実施例4Eに記載されるシュードモナス・プチダKT2440 BARは、pET28構築物からのN末端Hisタグが不溶性タンパク質を生成するように考えられたので、可溶性を改善し、精製を促進するために、様々な配列タグと共に発現させた。遺伝子は、サブクローニングし、発現させ、D−トリプトファンへのL−トリプトファンの変換での活性について試験した。
P.プチダKT2440 BARをコードする遺伝子は、実施例4Eに記載されるpET30構築物からサブクローニングした。ポリメラーゼ連鎖反応は、約100ngのプラスミドDNAを用いて、C末端Strepタグを生成するpET30aベクター(終止コドンなし、C末端Hisタグを生成する)(Novagen、Madison、WI)、pET22−b(+)(周辺質リーダー配列)、およびpASK−IBA3(IBA、Gottingen、Germany)中にクローニングするための5’制限部位およびオーバーハングを用いて設計されたプライマーを用いて行った。同じN末端プライマーを、実施例4Eでように、pET30aクローニングに用いた。用いるプライマー配列は以下の通りである。
pET30 C末端-XhoI:5'-AAGTCGCTCGAGGTCGACGAGTATCTTCGGG-3'(配列番号122);
pASK N末端:5'-ACGGTAGGTCTCAAATGCCCTTTCGCCGTACC-3'(配列番号123);
pASK C末端:5'-AACCGTGGTCTCAGCGCTGTCGAGGAGTATCTTCGGG-3'(配列番号124);
pET 22 N末端:5'-GCTCCACATGTCTCCCTTTCGCCGTACCCTTCTGGCTGCATC-3'(配列番号125);および
pET22 C末端:5'-CCGCCGGATCCTCAGTCGACGAGTATCTTCGGGTTGGAATTGC-3'(配列番号126)。
pET30構築物およびpASK−IBA3構築物についてのPCR産物を、Qiagen PCR精製キット(Qiagen、Valencia、CA)を用いて精製した。産物およびベクターを、NdeI/XhoI(pET30a)またはBsaI(pASK−IBA3)で消化し、その後、Qiagenゲル抽出キット(Qiagen、Valencia、CA)を用いて、0.8%TAE−アガロースゲルから精製した。消化したベクターおよび挿入断片は、Rapid(商標)DNA Ligation Kit(Roche、Indianapolis、IN)を用いてライゲーションした。約50ngの処理した挿入断片、100ngの処理したベクター(ベクターに対する挿入断片の3対1のモル比)、5UのT4 DNAリガーゼ、および1×ライゲーション緩衝液を室温で20分間インキュベートした。ライゲーション混合物を、TOP10化学的コンピテントイー・コリ細胞中に形質転換し、適切な抗生物質を含有するLB平板上で平板培養した。pET22構築物についてのPCR産物は、QIAquick(登録商標)Gel Extraction Kit(Qiagen、Valencia、CA)を用いて精製し、pCR−Blunt II−TOPO(Invitrogen)中にクローニングした。挿入断片配列は、検証し、pET−22b(+)(Novagen、Madison、WI)のNcoI制限部位およびBamHI制限部位の中へBamHI/PciI断片としてクローニングした。
プラスミドDNAは、イー・コリ発現宿主BL21(DE3)(Novagen、Madison、WI)中に形質転換した。BL21(DE3)中でのpET30構築物およびpET22構築物の誘発は、Overnight Express IIプロトコール(Novagen、Madison、WI)を用いて行った。pASK−IBA3構築物の誘発はメーカーのプロトコールに従って行った。細胞抽出物は、メーカーのプロトコールに従って、ベンゾナーゼヌクレアーゼを含有するNovagen BugBuster(商標)試薬中に細胞ペレットを懸濁させることによって一晩培養物から調製した。上清(細胞抽出物)は、可溶性タンパク質の分析および発現パーセントの評価のために4〜15%勾配ゲル上に装填した。全タンパク質アッセイは、標準物質としてBSAを用いて、Bio−Radキット(Bradfordアッセイ法)を用いて進めた。C末端HisタグBARは、十分に発現し、可溶性であり、N末端Hisタグタンパク質に対して有意に改善した。しかしながら、pASK−IBA3 Strepタグタンパク質は、全タンパク質または可溶性画分中で十分に発現するようには考えられなかった。C末端HisタグBARは、Novagen His−Bindカラムを用いて精製し、溶出の後になお可溶性であるように考えられた。しかしながら、一晩のイミダゾール中での−80℃での保存に際して、タンパク質は融解の際に沈殿した。したがって、タンパク質は細胞性抽出物中でアッセイした。タンパク質がイミダゾール溶出緩衝液からより適切に取り出された場合、タンパク質は可溶性のままであろうということが期待される。
P.プチダNBRC12996 BAR配列は、株KT2440に由来する上記に記載されるBARタンパク質とDNAレベルで91%同一およびタンパク質レベルで94%同一である。P.プチダのNBRC12996株は、日本のNITE(National Institute of Technology and Evaluation)Biological Resource Centerから得た。プライマーは、実施例4Eに記載されるP.プチダKT2440 BAR遺伝子のクローニングに用いたプライマーと同じものとした。株は、ペプトン10g、酵母抽出物2g、MgSO4・7H2O 1g、蒸留水1L、寒天15g、pH7.0を含有する培地上で成長させた。細胞はペトリ皿からこすり取り、400μLの脱イオン化し、加圧滅菌したH2O中に再懸濁させた。この細胞懸濁液を鋳型としてPCR反応に用いた。結果として生じたPCR産物はQIAquick(登録商標)PCR精製キット(Qiagen、Valencia、CA)を用いて精製し、BamHIおよびNdeIで消化した。消化DNAを1%アガロースゲル上で走らせ、最も目立つバンドをゲルから切り抜いた。DNAはQIAquick(登録商標)ゲル抽出キット(Qiagen、Valencia、CA)を用いて精製した。結果として生じた断片は、上記に記載されるようにNdeIおよびBamHIで消化し、精製したpET30にライゲーションした。ライゲーションしたものは、TOP10化学的コンピテント細胞(Invitrogen、Carlsbad、CA)中に形質転換し、50μg/mLカナマイシンを補足したLB平板上で平板培養した。単離コロニーは、画線培養し、精製し、プラスミドプレップの培養を開始するために用いた。挿入断片の存在を確認するために、プラスミドをゲル上で走らせた。挿入断片を有するプラスミドを、配列決定のためにAgencourt(Beverly、MA)に送付した。適正な配列を有する単離物を同定し、後の研究に用いた。
atgccctttcgccgtaccctcctggctgcatccctcgctctgctgatcactggccaggccccgctgtacgccgcaccgcccctgtcgatggacaacggcaccaccgccctgaccgcgcagaacagcaacgcctgggtcgaaatcagtgccggcgcactgcaacacaacatccgtaccttgcaggccgagttgggcggcaagtccaagctgtgcgccgtgctcaaggccgacgcctatggccacggtatcggcctggtgatgccgtcgatcatcgcccagggcgtgccctgcgtggcggtggccagcaacgaggaggcacgcgtggtccgcgccagtggcttcaccgggcaactggtgcgggtacgcctggccagcctcggcgaagtggaagatgccttgcagtacgacatggaagagctggttggcagcgccgagttcgcccgccagctcgatgccatcgccgaacgccacggcaagaccctgcgcattcacatggcgctcaattccagcggcatgagccgcaacggcgtggaaatgaccacctggtccggccggggtgaagcgctgcagatcactgaccagaagcacctccagctggtcgcgctgatgactcacttcgccgtggaagacaaggacgatgtgcgcaaaggcctggcagcgttcaacgaacagaccgactggctgatcaagcacgcgaagcttgatcgcagcaagctcaccctgcatgccgccaactccttcgctacgctggaagtgccggaagcgcacctggacatggtgcgtaccggtggcgcgctgttcggcgacaccgtgccgacgcgcaccgaataccaacgtgtcatgcagttcaagtcgcacgtggcggcggtgcacagctacccggcaggcaacaccgtcggctacgaccgcaccttcaccctggcgcgtgattcgcgcctggccaacatcaccgtgggttactccgatggctaccgccgggtgttcaccaacaagggccatgtgctgatcaacggccaccgagtgccagtggtgggcaaggtgtcgatgaacaccttgatggtcgatgtcaccgatttccccgatgtgaaggggggcaacgaagtggtgctgttcggcaaacaggccgggagggagatcacccaggccgagatagaagaaatcaacggcgcgctgctcgccgacctctacaccgtatggggcagttccaacccgaagatactcgtcgactgaである。
MPFRRTLLAASLALLITGQAPLYAAPPLSMDNGTTALTAQNSNAWVEISAGALQHNIRTLQAELGGKSKLCAVLKADAYGHGIGLVMPSIIAQGVPCVAVASNEEARVVRASGFTGQLVRVRLASLGEVEDALQYDMEELVGSAEFARQLDAIAERHGKTLRIHMALNSSGMSRNGVEMTTWSGRGEALQITDQKHLQLVALMTHFAVEDKDDVRKGLAAFNEQTDWLIKHAKLDRSKLTLHAANSFATLEVPEAHLDMVRTGGALFGDTVPTRTEYQRVMQFKSHVAAVHSYPAGNTVGYDRTFTLARDSRLANITVGYSDGYRRVFTNKGHVLINGHRVPVVGKVSMNTLMVDVTDFPDVKGGNEVVLFGKQAGREITQAEIEEINGALLADLYTVWGSSNPKILVDである。
pET30 NBRC12996 BARをBL21 DE3 pLysSコンピテント細胞中に形質転換し、タンパク質を発現させ、無細胞抽出物を、実施例4Eに記載されるように調製した。精製KT2440 BAR(実施例4Eに記載)(100μg)を12996BAR(100μLの無細胞抽出物)と比較した。結果を下記の表27に示す。
実験概要
ビブリオ・フィシェリの推定上のアラニンラセマーゼ(Genbankタンパク質受入番号AAW85230.1またはYP_204118)をコードする遺伝子をクローニングし、発現させ、D−トリプトファンへのL−トリプトファンの変換での活性について試験した。
ビブリオ・フィシェリ・ゲノムDNA(ATCC700601D)を鋳型としてアラニンラセマーゼ遺伝子のPCR増幅に用いた。プライマーは、株ES114(CP000020.1 GI:59478708の(NC_006840としても挙げられる)領域800842..802053)からの公開遺伝子配列に基づいて設計した。ポリメラーゼ連鎖反応は、pET28ベクターおよびpET30ベクター(Novagen、Madison、WI)中にクローニングするために5’制限部位およびオーバーハングを用いて設計したプライマーを用いてゲノムDNAに対して行った。
プライマー配列:
N末端:
5'-GCGGCCCATATGAAGTTTACTAAATGTGCAT-3'(配列番号130)および
C末端:
5'-GGCCGCGGATCCCTATTTGTAGATCTTAGGATTTG-3'(配列番号131)。
PCR産物は、Qiagenゲル抽出キット(Qiagen、Valencia、CA)を用いて0.8%TAE−アガロースゲルからゲル精製した。産物は、TOPOクローニングし、メーカーのプロトコール(Invitrogen、Carlsbad、CA)に従ってTOP10細胞中に形質転換した。プラスミドDNAは、Qiagenスピンミニプレップキット(Qiagen、Valencia、CA)を用いて結果として生じた形質転換体から精製し、NdeIおよびBamHIを用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するように考えられるプラスミドの配列は、汎発性のM13順方向プライマーおよびM13逆方向プライマーを用いてジデオキシ連鎖停止DNA配列決定によって検証し、配列は、上記に挙げた受入番号と同一であることが分かった。
プラスミドDNAは、イー・コリ発現宿主BL21(DE3)pLysS(Novagen、Madison、WI)中にサブクローニングした。培養物を成長させ、プラスミドは、Qiagenミニプレップキット(Qiagen、Valencia、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。
2種のP.プロファンダムBAR相同体のアミノ酸配列を公的データベース(NCBI)から得た。CR378673(ヌクレオチド342444..343658)の推定上のアラニンラセマーゼコード配列をプライマーから構築した。アミノ酸配列(NCBI受入番号CAG21670)は、GeneComposerソフトウェアパッケージ(バージョン1.0、Emerald Biosciences、Bainbridge Island、WA)を用いて最適なDNA配列を設計するために用いた。CR378673最適化DNA配列(配列番号132)は以下の通りである。
atgaagctgaagctgagcctggtcgccctggcactgatgggtcagactactgctaatgccgcaccactgctggtggacttcgataacaatgagcgtgaggaacgtgtgcaaagctctaatgcgtggctggagattgatacccaagcattcagtggcaatattcagttactgcagaaccaactgaaagccgacaccaagatctgtgcgattatgaaggcggatgcatacggtaatggcattgccggcttgatgcctagtatcattgctaaccaagtgccttgtgttggtatcaccagcaatgaggaagcgcgggtggttcgtaaacatggctttattgggaagatcatgcgtgtccgtgcagcctcgaagaatgaaattgagggtggcttgcagtaccagatggaagaattgatcggtacgaaggctcaagccgatcaaatcatcgaaattgcacgcgcaaatggcacgacgattccggttcatttagccttgaatacaagcggcatgggccgcaacggtctggacctgacgacctacgaaggccaagttgaaggtgtagagattgctggcgatccaaacctggagattgtcggcatgatgactcatttcccgaacgagggactggacgaaatcaaacggaaagtcaaacgtttcaaagtagaaacgaaatggttaatggattccactgacttgaagcgcaaagatgtgacgctccacgtcgcaaacagctatatcaccttgaatctgcctgaagcgcatctggatatggtacgcccaggtggcatgctgtatggcgactatccggcgacagcgccgtatcagcgtatcgtaagcttcaagacccacgttgcctctttgcaccactttccggctggctcaaccattgggtacggatctaccgctgttctggaacgtgattcagttctggctaatctgccgattggctattcggatggcttcgcgcgctcgttaggaaataaagccgaagtcctgattaacggccagcgtgcgcgcgtcatgggtatggtcagtatgaacacgacgatggtcgatgtaacggatattgtggatgttcagaccaatgaagaagtcgtgatctttggccgccagggtttcgaagagattacgggcgaggagacggaagagaagtctaatcgtattcttccggaacattacactgtgtggggcgccacaaacccgcgtatttatcgctaa。
ATGAACTTTAAGATGACTCTGTTAAGCCTGGCCATTACATTCCCGAGCTTCAGCATCTATAGCGCGCCACTGGTCATTGATCAGAACCTGCCAAGCGAACAGTCGATTCAGCAAAGCAACAGCTGGCTGGAAGTTAGCCTGGGCCAGTTTAAATCCAATATTGAACAATTTAAATCTCATATTAAAGCCGATACTAAGATTTGTGCCGTTATGAAAGCCGATGCATACGGCAATGGCATCTTCGGTCTGATGCCGACAATTCTGGAACAGCAAATCCCATGCGTGGCGATTGCAAGTAACGCGGAAGCTCGCGCTGTGCGTGAAAGCGGGTTTAAGGGCCAGCTGCTGCGTGTCCGCAGCGCGAGCTTAGGCGAGATTAAACAGTCACTGGACCTGAACATTGAAGAACTGATCGGCTCACATCAGCAGGCGAAGTTCATTGCAGAGCTGGGTGTAGAACGTAATCAGAAGATTAACGTTCATTTAGCTCTGAACGACGGAGGGATGGGTCGCAATGGGATCGATATGTCTACCGAACAAGGCAAAGCCGAGGCCCTCGACATCGCGACCCAGGCAAATCTGAACATTGTTGGTATTATGACTCACTTCCCGAACTATAATGCGGATAAAGTGCGTGTGAAGCTGAAAGACTTCCAGACAAACTCCAGCTGGCTGATCAAGCAGGCGGATCTGAAGCGCGATGAACTCACGCTCCACGTGGCCAACAGCTATGTGTCCATTAATGTTCCAGAAGCGCAACTGGATATGGTTCGCCCGGGCGGCGTGCTGTATGGCGATCTTCCGACCAATCCGGAATATCCGAGCATCGTATCGTTCAAGACGCGGATTGCGTCAATTCACCAGCTGCCAGCATCCCAGACCGTGGGCTACGATTCGACCTATATTACGAAACGTGATAGCGTTCTGGCAAACCTGCCAGTCGGCTACAGTGATGGCTATCCGCGCCGTATGGGTAATCAGGCTGATGTGATTATCAACGGACAACGCGCCAAAGTGGTGGGTGTGACCAGCATGAATACTAGTATCGTCGATATTACCGATATTAAAGGCGTTAAACAGGGTCAAGAAGTTACCCTGTTTGGCAAGCAGAAGAATGTGCAGATTAGCGTGGCCGAAATGGAGGATTATTCGAAGTTAATCTTCCCGGAACTGTACACCATGTGGGGTCAGGCGAATCCGCGTTACTACCTGAAGTAA(配列番号137)。
実験概要
シュードモナス・タエトロレンス(P.グラベオレンスとしても知られている)アルギニンラセマーゼ(Genbank受入番号AB096176、核酸配列)およびそのI384M変異体は、クローニングし、発現させ、D−トリプトファンへのL−トリプトファンの変換での活性について試験した。この遺伝子は、上記に記載される、KT2440に由来するP.プチダBAR遺伝子に72%同一であり、NBRC12996に由来するP.プチダBAR遺伝子に73%同一である。アミノ酸配列は双方のP.プチダBARタンパク質に72%同一である。
シュードモナス・タエトロレンス(ATCC4683)は、225rpmで振盪させながら28℃で栄養ブロス中で成長させた。ポリメラーゼ連鎖反応は、pET28ベクターおよびpET30ベクター(Novagen、Madison、WI)中にクローニングするために5’制限部位およびオーバーハングを用いて設計したプライマーを用いて全細胞に対して行った。
N末端:5'-ATAATACATATGCCCTTCTCCCGTACCC-3'(配列番号138)および
C末端:5'-GCGGCGGGATCCTTACTGATCTTTCAGGATT-3'(配列番号139)。
PCR産物は、Qiagenゲル抽出キット(Qiagen、Valencia、CA)を用いて0.8%TAE−アガロースゲルからゲル精製した。産物は、TOPOクローニングし、メーカーのプロトコール(Invitrogen、Carlsbad、CA)に従ってTOP10細胞中に形質転換した。プラスミドDNAは、Qiagenスピンミニプレップキット(Qiagen、Valencia、CA)を用いて結果として生じた形質転換体から精製し、NdeIおよびBamHIを用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するように考えられるプラスミドの配列は、汎発性のM13順方向プライマーおよびM13逆方向プライマーを用いてジデオキシ連鎖停止DNA配列決定によって検証した。
プラスミドDNAは、イー・コリ発現宿主BL21(DE3)pLysS(Novagen、Madison、WI)中に形質転換した。培養物を成長させ、プラスミドは、Qiagenミニプレップキット(Qiagen、Valencia、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。
ATGCCCTTCTCCCGTACCCTGCTCGCCCTTTCCCTTGGCATGGCATTGCTGCAAAACCCGGCCTTTGCTGCGCCACCCCTGTCGATGACCGACGGCGTAGCTCAAGTGAATACCCAGGACAGCAATGCCTGGGTCGAAATCAATAAAGCCGCGTTCGAGCACAACATACGGACTCTGCAAACCGCCCTCGCCGGCAAGTCGCAGATCTGCGCCGTACTCAAGGCGGATGCCTATGGCCACGGTATCGGCTTGTTGATGCCCTCGGTGATCGCCATGGGTGTTCCCTGTGTCGGTGTCGCCAGCAACGAAGAAGCCCGCGTCGTGCGCGAGAGCGGTTTCAAGGGTCAACTGATACGCGTGCGCACCGCTGCCCTGAGCGAACTGGAAGCTGCACTGCCGTACAACATGGAAGAGCTGGTGGGCAACCTGGACTTCGCGGTCAAGGCCAGCCTGATTGCCGAGGATCACGGTCGCCCGCTGGTGGTGCACCTGGGTCTGAATTCCAGCGGCATGAGCCGTAACGGAGTGGACATGACCACCGCTCAGGGCCGTCGTGATGCGGTAGCTATCACCAAGGTGCCAAACCTGGAAGTGCGGGCGATCATGACCCACTTCGCGGTCGAAGATGCTGCCGACGTGCGTGCCGGGCTCAAGGCCTTCAATCAGCAAGCCCAATGGCTGATGAACGTGGCCCAGCTTGATCGCAGCAAGATCACCCTGCACGCGGCCAACTCGTTCGCCACACTGGAGGTGCCCGAATCGCATCTGGACATGGTCCGCCCCGGCGGCGCGCTGTTCGGCGACACCGTACCGTCCCACACCGAGTACAAGCGGGTCATGCAGTTCAAGTCCCACGTGGCGTCGGTCAACAGCTACCCCAAGGGCAACACCGTCGGTTATGACCGCACGTACACCCTGGGCCGCGACTCGCGGCTGGCCAACATCACCGTCGGCTACTCTGACGGCTACCGCCGCGCGTTTACCAATAAAGGGATTGTGCTGATCAACGGCCATCGCGTGCCAGTGGTGGGCAAAGTCTCGATGAACACCCTGATGGTGGACGTCACTGACGCGCCGGATGTGAAAAGCGGCGATGAAGTGGTGCTGTTCGGGCACCAGGGCAAGGCCGAGATTACCCAGGCTGAGATCGAAGACATCAACGGTGCACTGCTTGCGGATCTGTATACCGTGTGGGGCAATTCCAACCCTAAAATCCTGAAAGATCAGTAA
(配列番号140)。
MPFSRTLLALSLGMALLQNPAFAAPPLSMTDGVAQVNTQDSNAWVEINKAAFEHNIRTLQTALAGKSQICAVLKADAYGHGIGLLMPSVIAMGVPCVGVASNEEARVVRESGFKGQLIRVRTAALSELEAALPYNMEELVGNLDFAVKASLIAEDHGRPLVVHLGLNSSGMSRNGVDMTTAQGRRDAVAITKVPNLEVRAIMTHFAVEDAADVRAGLKAFNQQAQWLMNVAQLDRSKITLHAANSFATLEVPESHLDMVRPGGALFGDTVPSHTEYKRVMQFKSHVASVNSYPKGNTVGYDRTYTLGRDSRLANITVGYSDGYRRAFTNKGIVLINGHRVPVVGKVSMNTLMVDVTDAPDVKSGDEVVLFGHQGKAEITQAEIEDINGALLADLYTVWGNSNPKILKDQ
(配列番号204)。
P.taetリーダーなし F NdeI:5'-GGTTAATTCATATGGCGCCACCCCTGTCGAT-3'(配列番号180)
P taetC末端Xho:5'-AAGTCGCTCGAGCTGATCTTTCAGGATTTTAG-3'(配列番号181)。
突然変異誘発は、タグなしタンパク質をもたらすpET30中のP.タエトロレンスBAR遺伝子を用いて、QuickChange−Multi部位特異的突然変異誘発キット(Stratagene、La Jolla、CA)を用いて行った。以下の突然変異誘発性プライマーはI384M変化を加えるために用いた。5'-TACCCAGGCTGAGATGGAAGACATCAACG-3'(配列番号141)。
ストレプトマイセス種は、D−トリプトファンのごときD−アミノ酸を含有する、カルシウム依存性の抗生物質を生成することで知られている。トリプトファンが抗生物質中への取り込み前またはその後にラセミ化されるかどうかは知られていない。公的なストレプトマイセスゲノムは、シュードモナス種に由来する活性を有するアラニンラセマーゼに対して高度に相同なアラニンラセマーゼを含有していないが、実際に酵素が同様に広域特異性を有するかどうかを決定するためにクローニング作業を試みた。
Y.シュードツベルクロシスBAR相同体は、St.Louis、MOのWashington UniversityのVirginia MillerおよびMatt Lawrenzから得たゲノムDNAからクローニングした(株YPIII)。用いたプライマーは以下の通りである:5’−gcggcgcatatgcacgttcgttttcgtc−3’(配列番号142)および5’−ggcggcgggatcccggtgaaataacttaatctac−3’(配列番号143)。遺伝子をPCRによってゲノムDNAから増幅し、QIAquick(登録商標)PCR精製キット(Qiagen、Valencia、CA)を用いて精製した。断片はNdeIおよびBamHIを用いて消化した。消化物を1%アガロースゲル上で走らせ、ラセマーゼ断片はゲルから切り取った。DNAはQIAquick(登録商標)ゲル抽出キット(Qiagen、Valencia、CA)を用いてゲルから精製した。断片は、上記に記載されるように消化し、精製したpET30にライゲーションした。ライゲーションしたものは、TOP10化学的コンピテント細胞(Invitrogen、Carlsbad、CA)に形質転換した。プラスミドDNAを単離し、挿入断片についてスクリーニングするために1%アガロースゲル上で走らせた。挿入断片を有する数種のプラスミド単離物を、配列決定のためにAgencourt(Beverly、MA)に送付した。適正な配列を有する単離物を選択し、発現株を作製するためにBL21 DE3 pLysSコンピテント細胞中に形質転換した。
atgcacgttcgttttcatcatttattcttattaccattaataactttggtcgcttgtagccaacccgtatcaaaaaaccatcttagcctgacctcactatctgccaacgcccagcaacctgtagtaaataatgcgtggcttgaaatctctcaaggtgcgctggatttcaatactaaaaagatgcttacactgctggataataaatccacactttgtgcaatattaaaaggtgatgcctatggacatgacctgaccttagtcacaccggtgatgctaaaaaacaatgtgcaatgtattggggttgccagcaatcaggaactaaaaacggtacgtgatctaggatttacggggcagttgatacgggtcagaagtgcaacattaaaagaaatgcaacaagctatggcttacgatgttgaagaacttattggcgataaaaccgtcgctgagcagttaaataatattgcaaaactgaatggaaaagttctgcgtatccatctggcactgaactccgcagggatgtctcgtaatgggctggaggtcagtaaggcccgcggtttaaatgacgcaaagacaattgtaggtttaaaaaatctgacaatcgttggcatcatgtcgcactacccggtggaagatgctagcgaaatcaaagcagacttggctcgattccagcaacaagccaaagatgttatcgcggtcacggggctaaaacgtgaaaagattaagctccacgtcgccaatacattcgcgaccttagcggtgcctgattcatggttggatatggtccgtgtgggaggggtgttttatggtgacaccatcgccagcacagagtataagcgggtcatgaccttcaaatctaacatcgcatcgctgaacaactaccctaagggcggtactgttggctatgaccggacctatacattgaaacgtgattccctgctggcgaatatccccgtgggttatgccgatgggtatcgccgagtatttagtaatgcggggcatgtgattattcaaggtcagcgcctgcccgtattaggcaaaacatcaatgaatacggtcatggtagacgtcaccgatctgaaaaaagtgagtttaggtgatgaagttgtcttgttcggtaagcaaggcaatgcggaaattcaggcagaagaaattgaagatctcagtggcgcactctttaccgaaatgtcaattctgtggggcgcaaccaataagcgtattctggtagattaa(配列番号144)。
MHVRFHHLFLLPLITLVACSQPVSKNHLSLTSLSANAQQPVVNNAWLEISQGALDFNTKKMLTLLDNKSTLCAILKGDAYGHDLTLVTPVMLKNNVQCIGVASNQELKTVRDLGFTGQLIRVRSATLKEMQQAMAYDVEELIGDKTVAEQLNNIAKLNGKVLRIHLALNSAGMSRNGLEVSKARGLNDAKTIVGLKNLTIVGIMSHYPVEDASEIKADLARFQQQAKDVIAVTGLKREKIKLHVANTFATLAVPDSWLDMVRVGGVFYGDTIASTEYKRVMTFKSNIASLNNYPKGGTVGYDRTYTLKRDSLLANIPVGYADGYRRVFSNAGHVIIQGQRLPVLGKTSMNTVMVDVTDLKKVSLGDEVVLFGKQGNAEIQAEEIEDLSGALFTEMSILWGATNKRILVD(配列番号145)。
実施例4Eに言及されるように、エロモナス種は、シュードモナスに由来する酵素に同様の特性を有する広域特異性アミノ酸ラセマーゼを含有するといった証拠が文献にある。この例は、A.ハイドロフィラ、A.ジャンデイ、A.ソブリアおよびA.キャビエの広域特異性アミノ酸ラセマーゼの同定および単離ならびにA.キャビエ細胞性抽出物中のトリプトファンラセマーゼ活性の同定を記載する。この例はまた、A.シューベルティーおよびA.サルモニシダのBAR相同体の部分的な核酸配列の単離をも記載する。
AVAAPYLPLASDHRNGEVQTASNAWLEVDLGAFEHNIQTLKDRLGDKGPKICAIMKADAYGHGIDLLVPSVVKAGIPCIGIASNEEARVAREKGFTGRLMRVRAATPAEVEQALPYKMEELIGSLVSAQGIADIAQRHHTNIPVHIALNSAGMSRNGIDLRLADSKEDALAMLKLKGITPVGIMTHFPVEEKEDVKMGLAQFKLDSQWLLEAGKLDRSKITIHAANSFATLEVPDAYFDMVRPGGLLYGDSIPSYTEYKRVMAFKTQVASVNHYPAGNTVGYDRTFTLKRDSWLANLPLGYSDGYRRALSNKAYVLIQGQKVPVVGKTSMNTIMVDVTDLKGVKPGDEVVLFGRQGEAEVKQADLEEYNGALLADMYTIWGYTNPKKIK(配列番号146)。
tcttggggtt ggtgtagccc cagatggtgt acatgtccgc cagcagggcg
ccgttgtact cttccagatc cgcctgtttc acctcagcct caccctggcg
gccgaacagc accacctcgt caccgggttt gacccctttc agatcggtca
cgtccaccat gatggtgttc atggaggtct tgcccaccac cggcaccttc
tggccctgga tcagcacata ggccttgttg ctcagcgccc ggcgatagcc
gtcggagtag cccagcggca ggttggcgag ccaggagtcg cgcttgaggg
tgaaggtgcg gtcataaccg acggtgttgc cggccgggta gtggttgacg
gaggcaacct gggtcttgaa cgccatcacc cgcttgtact cggtgtagga
ggggatggag tcaccgtaca gcaggccgcc cgggcgcacc atgtcgaagt
aggcgtccgg cacttccagg gtggcgaagg agttggcggc gtggatggtg
atcttgctgc gatccagctt gcccgcttcc agcagccact gggagtccag
tttgaactgg gccagcccca tcttgacgtc ctctttctcc tccaccggga
agtgggtcat gatgccgacc ggggtgatcc ccttgagctt gagcatggcc
agcgcgtctt ccttggagtc agccaggcgc agatcgatgc cgttgcggct
catgccggcg gagttgagcg cgatgtgcac cgggatattg gtgtggtggc
gctgggcgat gtcggcgatg ccctgagcac tcaccaggct gccgatgagc
tcttccatct tgtagggcag ggcctgttcc acttcggccg gggtggcggc
acgtacccgc atcaggcggc cggtgaagcc cttctcacgg gccacgcggg
cctcttcgtt gctggcgatg ccgatgcagg ggatgccggc cttgaccacc
gagggcacca gcaggtcgat gccgtggccg taggcgtcgg ccttcatgat
ggcgcagatc ttcggccctt tgtcaccgag gcgatccttg agggtctgga
tgttgtgctc gaaggcgccg agatcgactt ccagccaggc attgctggcg
gtctgcactt cgccgttgcg atgatcgctg gccagcggca ggtaaggggc
cgcgacggcc tgaccggcca gcaggcccag gatcagcgtg gccagcagtg t(配列番号147)。
ハイドロフィラFNde1(配列番号148):5'-TTCCAAGGCATATGCCCTTCTCCCGTACACTGCTGGCCACGCTGATCCT 3';および
ハイドロフィラRBamH1(配列番号149):5'-GGAACCTTGGATCCTCAATCTTTGATTTTCTTGGGGTTGGTGTAGCCCCAGATG 3'。
PCR産物は、Qiagenゲル抽出キット(Qiagen、Valencia、CA)を用いて0.8%TAE−アガロースゲルからゲル精製した。産物は、TOPOクローニングし、メーカーのプロトコール(Invitrogen、Carlsbad、CA)に従ってTOP10細胞中に形質転換した。プラスミドDNAは、Qiagenスピンミニプレップキット(Qiagen、Valencia、CA)を用いて結果として生じた形質転換体から精製し、NdeIおよびBamHIを用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するように考えられるプラスミドの配列は、汎発性のM13順方向プライマーおよびM13逆方向プライマーを用いてジデオキシ連鎖停止DNA配列決定によって検証した。
MPFSRTLLATLILGLLAGQAVAAPYLPLASDHRNGEVQTASNAWLEVDLTAFEQNLQTKTRLGDKGPQICAIMKADAYGHGIDLLVPSVIKAEIPCIGIASNEEARVAREKGFSGRLMRVRAATPIEVEQALPYKLEELVGSLVSAQGISDIALRHHTTIPVHVALNSAGMSRNGIDLRLADAKQDALAMLKLKGITPVGIMTHFPVEEKEDVKLGLAQFKLDSQWLLEAGKLDRSKITIHAANSFATLAVPDAYFDMVRPGGLLYGDSIPSYTEYKRVMAFKTQVASVNHYAAGNTVGYDRTFTLKRDSWLANLPLGYSDGYRRALSNKAYVLIQGQKVPVVGKTSMNTIMVDVTDLKGVKPGDEVVLFGRQGEAEVKQADLEEYNGALLADMYTIWGYTNPKKIKD(配列番号151)。
ATGCCCTTCTCCCGTACACTGCTGGCCACGCTGATCCTGGGCCTGCTGGCCGGTCAAGCCGTCGCAGCCCCCTATCTGCCTCTGGCAAGCGATCATCGCAACGGCGAAGTACAAACCGCCAGCAACGCCTGGCTGGAAGTAGATCTGACCGCGTTTGAACAGAATCTGCAGACCCTCAAGACCCGCCTCGGCGACAAGGGCCCGCAGATCTGCGCCATCATGAAGGCGGACGCCTACGGTCACGGTATCGATCTGCTGGTTCCCTCCGTCATCAAGGCCGAGATCCCCTGTATCGGCATCGCCAGCAACGAAGAGGCCCGCGTCGCCCGCGAGAAGGGGTTCAGCGGCCGCCTGATGCGGGTACGGGCCGCCACACCTATCGAAGTGGAACAGGCCCTGCCCTACAAGCTGGAAGAGCTGGTTGGCAGCCTGGTGAGTGCTCAGGGGATCTCCGACATCGCCCTGCGCCACCACACCACCATTCCGGTGCATGTCGCCCTCAACTCCGCCGGTATGAGCCGCAACGGCATCGACCTGCGTCTGGCCGATGCCAAGCAAGATGCGCTGGCCATGCTCAAGCTCAAGGGGATCACCCCGGTCGGCATCATGACCCACTTCCCGGTGGAGGAGAAAGAGGACGTCAAGCTGGGGCTGGCTCAGTTCAAGCTGGACTCCCAGTGGCTGCTGGAAGCAGGCAAGCTGGATCGCAGCAAGATCACCATCCATGCCGCCAACTCCTTCGCCACCCTGGCAGTGCCGGACGCCTACTTTGACATGGTGCGCCCGGGCGGCCTGCTCTACGGCGACTCCATCCCCTCCTACACCGAATACAAGCGGGTGATGGCATTCAAGACCCAGGTCGCCTCGGTCAACCACTATGCGGCGGGCAACACAGTCGGTTATGACCGCACCTTTACTCTCAAACGTGACTCCTGGCTCGCCAACCTGCCGCTCGGTTACTCCGACGGCTATCGCCGTGCGCTCAGCAACAAGGCCTATGTGCTGATCCAGGGTCAGAAGGTGCCGGTGGTCGGCAAGACCTCCATGAACACCATCATGGTGGACGTGACCGATCTCAAAGGGGTAAAGCCCGGTGATGAAGTGGTGCTGTTTGGCCGTCAGGGTGAGGCAGAAGTGAAACAGGCTGATCTGGAGGAGTACAACGGCGCCCTGTTGGCGGACATGTACACCATCTGGGGCTACACCAACCCCAAGAAAATCAAAGATTGA(配列番号150)。
プラスミドDNAは、イー・コリ発現宿主BL21(DE3)(Novagen、Madison、WI)中に形質転換した。培養物を成長させ、プラスミドは、Qiagenミニプレップキット(Qiagen、Valencia、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。BL21DE3での誘発は、メーカーの指示に従ってOvernight Express培地中で行った(Novagen、Madison、WI)。
atgcacaaga agacactgct ggccaccttg atcctgggcc tgctggccgg tcaagccgtc
gcagccccct atctgcctct ggcaagcgat catcgcaacg gcgaagtaca aaccgccagc
aacgcctggc tggaagtaga tctgaccgcg tttgaacaga atctgcagac cctcaagacc
cgcctcggcg acaagggccc gcagatctgc gccatcatga aggcggacgc ctacggtcac
ggtatcgatc tgctggttcc ctccgtcatc aaggccgaga tcccctgtat cggcatcgcc
agcaacgaag aggcccgcgt cgcccgcgag aaggggttca gcggccgcct gatgcgggta
cgggccgcca cacctatcga agtggaacag gccctgccct acaagctgga agagctggtt
ggcagcctgg tgagtgctca ggggatctcc gacatcgccc tgcgccacca caccaccatt
ccggtgcatg tcgccctcaa ctccgccggt atgagccgca acggcatcga cctgcgtctg
gccgatgcca agcaagatgc gctggccatg ctcaagctca aggggatcac cccggtcggc
atcatgaccc acttcccggt ggaggagaaa gaggacgtca agctggggct ggctcagttc
aagctggact cccagtggct gctggaagca ggcaagctgg atcgcagcaa gatcaccatc
catgccgcca actccttcgc caccctggca gtgccggacg cctactttga catggtgcgc
ccgggcggcc tgctctacgg cgactccatc ccctcctaca ccgaatacaa gcgggtgatg
gcattcaaga cccaggtcgc ctcggtcaac cactatgcgg cgggcaacac agtcggttat
gaccgcacct ttactctcaa acgtgactcc tggctcgcca acctgccgct cggttactcc
gacggctatc gccgtgcgct cagcaacaag gcctatgtgc tgatccaggg tcagaaggtg
ccggtggtcg gcaagacctc catgaacacc atcatggtgg acgtgaccga tctcaaaggg
gtaaagcccg gtgatgaagt ggtgctgttt ggccgtcagg gtgaggcaga agtgaaacag
gctgatctgg aggagtacaa cggcgccctg ttggcggaca tgtacaccat ctggggctac
accaacccca agaagatcaa acgctga
(配列番号176)。
1 mhkktllatl ilgllagqav aapylplasd hrngevqtas
41 nawlevdlta feqnlqtlkt rlgdkgpqic aimkadaygh
81 gidllvpsvi kaeipcigia sneearvare kgfsgrlmrv
121 raatpieveq alpykleelv gslvsaqgis dialrhhtti
161 pvhvalnsag msrngidlrl adakqdalam lklkgitpvg
201 imthfpveek edvklglaqf kldsqwllea gkldrskiti
241 haansfatla vpdayfdmvr pggllygdsi psyteykrvm
281 afktqvasvn hyaagntvgy drtftlkrds wlanlplgys
321 dgyrralsnk ayvliqgqkv pvvgktsmnt imvdvtdlkg
361 vkpgdevvlf grqgeaevkq adleeyngal ladmytiwgy
401 tnpkkikr
(配列番号177)。
A.hydroph F Nde1 5'-GGAACCTTCATATGCACAAGAAGACACTGCTGG-3'(配列番号182);
A.hydroph R BamH1(タグなし) 5'-GGTTCCAAGGATCCTCAGCGTTTGATCTTCTTGGG-3'(配列番号183);および
A.hydroph R Xho1(C末端タグ) 5'-GGCCAATTCTCGAGGCGTTTGATCTTCTTGGGGT-3'(配列番号184)。
エロモナス・キャビエATCC14486は37℃で栄養ブロス中で成長させた。培養物からの細胞(200mL)は、遠心分離し、0.85%NaClを用いて1回洗浄した。細胞ペレットは、5μL/mLプロテアーゼインヒビターカクテルセット#3(Calbiochem−Novabiochem Corp.、San Diego、CA)および1μL/mLベンゾナーゼヌクレアーゼを含有する5mL/湿細胞重量gのBugBuster(商標)(Novagen、Madison、WI)試薬中で再懸濁させた。試料は、オービタルシェーカー上で20分間室温でインキュベートした。不溶性細胞片は、4℃で20分間16,000xgでの遠心分離によって除去した。無細胞抽出物はPD−10カラム(GE Healthcare、Piscataway、NJ)で脱塩した。
Aer deg F2:5'-GCCAGCAACGARGARGCMCGCGT-3'(配列番号152);および
Aer deg R1:5'-TGGCCSTKGATCAGCACA-3'(配列番号153)
KはGまたはTを示し、RはAまたはGを示し、SはCまたはGを示し、MはAまたはCを示す。
PCR産物は、Qiagenゲル抽出キット(Qiagen、Valencia、CA)を用いて0.8%TAE−アガロースゲルからゲル精製した。産物は、TOPOクローニングし、メーカーのプロトコール(Invitrogen、Carlsbad、CA)に従ってTOP10細胞中に形質転換した。プラスミドDNAは、Qiagenスピンミニプレップキット(Qiagen、Valencia、CA)を用いて結果として生じた形質転換体から精製し、EcoR1を用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するように考えられるプラスミドの配列は、汎発性のM13順方向プライマーを用いたジデオキシ連鎖停止DNA配列決定によって検証した。
GCCAGCAACGARGARGCMCGCGTTGCCCGCGAGAAGGGCTTCGAAGGTCGCCTGATGCGGGTACGTGCCGCCACCCCGGATGAAGTGGAGCAGGCCCTGCCCTACAAGCTGGAGGAGCTCATCGGCAGCCTGGAGAGCGCCAAGGGGATCGCCGACATCGCCCAGCGCCATCACACCAACATCCCGGTGCACATCGGCCTGAACTCCGCCGGCATGAGCCGCAACGGCATCGATCTGCGCCAGGACGATGCCAAGGCCGATGCCCTGGCCATGCTCAAGCTCAAGGGGATCACCCCGGTCGGCATCATGACCCACTTCCCGGTGGAGGAGAAAGAGGACGTCAAGCTGGGGCTGGCCCAGTTCAAGCTGGACTACCAGTGGCTCATCGACGCCGGCAAGCTGGATCGCAGCAAGCTCACCATCCACGCCGCCAACTCCTTCGCCACCCTGGAAGTACCGGAAGCCTACTTTGACATGGTGCGCCCGGGCGGCATCATCTATGGCGACACCATTCCCTCCTACACCGAGTACAAGAAGGTGATGGCGTTCAAGACCCAGGTCGCCTCCGTCAACCACTACCCGGCGGGCAACACCGTCGGCTATGACCGCACCTTCACCCTCAAGCGCGACTCCCTGCTGGCCAACCTGCCGATGGGCTACTCCGACGGCTACCGCCGCGCCATGAGCAACAAGGCCTATGTGCTGATCMASGGCCA(配列番号154)、RはAまたはGを示し、SはCまたはGを示し、MはAまたはCを示す。
ASNEEARVAREKGFEGRLMRVRAATPDEVEQALPYKLEELIGSLESAKGIADIAQRHHTNIPVHIGLNSAGMSRNGIDLRQDDAKADALAMLKLKGITPVGIMTHFPVEEKEDVKLGLAQFKLDYQWLIDAGKLDRSKLTIHAANSFATLEVPEAYFDMVRPGGIIYGDTIPSYTEYKKVMAFKTQVASVNHYPAGNTVGYDRTFTLKRDSLLANLPMGYSDGYRRAMSNKAYVLIXG(配列番号155)。
XはH、Q、N、またはKである。
atgcacaaga aaacactgct cgcgaccctg atctttggcc tgctggccgg ccaggcagtc gccgccccct atctgccgct cgccgacgac caccgcaacg gtcaggaaca gaccgccgcc aacgcctggc tggaagtgga tctcggcgcc ttcgagcaca acatccagac cctgaagaat cgcctcggtg acaagggccc gcagatctgc gccatcatga aggcggacgc ctacggtcac ggcatcgacc tgctggtccc ttccgtggtc aaggcaggca tcccctgcat cggcatcgcc agcaacgaag aagcacgtgt tgcccgcgag aagggcttcg aaggtcgcct gatgcgggta cgtgccgcca ccccggatga agtggagcag gccctgccct acaagctgga ggagctcatc ggcagcctgg agagcgccaa ggggatcgcc gacatcgccc agcgccatca caccaacatc ccggtgcaca tcggcctgaa ctccgccggc atgagccgca acggcatcga tctgcgccag gacgatgcca aggccgatgc cctggccatg ctcaagctca aggggatcac cccggtcggc atcatgaccc acttcccggt ggaggagaaa gaggacgtca agctggggct ggcccagttc aagctggact accagtggct catcgacgcc ggcaagctgg atcgcagcaa gctcaccatc cacgccgcca actccttcgc caccctggaa gtaccggaag cctactttga catggtgcgc ccgggcggca tcatctatgg cgacaccatt ccctcctaca ccgagtacaa gaaggtgatg gcgttcaaga cccaggtcgc ctccgtcaac cactacccgg cgggcaacac cgtcggctat gaccgcacct tcaccctcaa gcgcgactcc ctgctggcca acctgccgat gggctactcc gacggctacc gccgcgccat gagcaacaag gcctatgtgc tgatccatgg ccagaaggcc cccgtcgtgg gcaagacttc catgaacacc accatggtgg acgtcaccga catcaagggg atcaaacccg gtgacgaggt ggtcctgttc ggacgccagg gtgatgccga ggtgaaacaa tctgatctgg aggagtacaa cggtgccctc ttggcggaca tgtacaccgt ctggggctat accaacccca agaagatcaa gcgctaa
(配列番号178)。
1 mhkktllatl ifgllagqav aapylpladd hrngqeqtaa
41 nawlevdlga fehniqtlkn rlgdkgpqic aimkadaygh
81 gidllvpsvv kagipcigia sneearvare kgfegrlmrv
121 raatpdeveq alpykleeli gslesakgia diaqrhhtni
161 pvhiglnsag msrngidlrq ddakadalam lklkgitpvg
201 imthfpveek edvklglaqf kldyqwlida gkldrsklti
241 haansfatle vpeayfdmvr pggiiygdti psyteykkvm
281 afktqvasvn hypagntvgy drtftlkrds llanlpmgys
321 dgyrramsnk ayvlihgqka pvvgktsmnt tmvdvtdikg
361 ikpgdevvlf grqgdaevkq sdleeyngal ladmytvwgy
401 tnpkkikr
(配列番号179)。
A.キャビエF Nde1 5'-GGAACCTTCATATGCACAAGAAAACACTGCTCGCGACC-3'(配列番号185);
A.キャビエR BamH1(タグなし) 5'-GGTTCCAAGGATCCTTAGCGCTTGATCTTCTTGGGGTTG-3'(配列番号186);および
A.キャビエR Xho1(C末端タグ) 5'-TTCCAAGGCTCGAGGCGCTTGATCTTCTTGGGGTTGGTA-3'(配列番号187)。
A.cavリーダーなし F NdeI 5' CCTTGGAACATATGGCCCCCTATCTGCCGCT 3(配列番号188).'
Aer deg F1 mod:5'-AAGGCSGAYGCCTAYGGYCACGG-3'(配列番号189)
SはCまたはGおよびYはCまたはTである、ならびに
Aer deg R3:5'-CGGCGRTAGCCRTCRGAGTA-3'(配列番号190)、RはAまたはGである。
ATGCACAAGAAAACGCTATTGGCCACCCTGATCTTCGGCCTGCTCGCGGGCCAAGCCGTTGCGGCTCCCTATCTGCCCCTTGCGACGGATCATCGCAACGGTCAGGAGCAAACCGCCAGCAACGCCTGGTTGGAAGTGGATCTGGGCGCCTTCGAACACAATATCCAGACCCTCAAGGATCGCCTCGGTGACAAGGGTCCGCAGATCTGCGCCATCATGAAGGCCGACGCCTATGGTCATGGCATCGACCTGCTGGTCCCCTCCGTGGTCAAGGCCAATATCCCCTGCATCGGCATCGCCAGCAACGAAGAGGCCCGCGTCGCCCGCGAGAAGGGCTTTACCGGCCGTCTGATGCGGGTGCGTGCCGCCACACCGGCCGAAGTGGAGCAGGCGCTGCCCTACAAGATGGAAGAGCTGATCGGCAGTCTGGTGAGTGCTCAGGGGATCGCCGACATCGCCCAGCGCCACCACACCAATATTCCGGTACACATTGGTCTCAACTCTGCTGGCATGAGCCGCAACGGTATCGACCTGCGTCTGGCCGATGCCAAGCAGGATGCGCTGGCCATGCTCAAGCTCAAGGGGATCACCCCGGTCGGCATCATGACCCACTTCCCGGTGGAGGAGAAAGAGGACGTCAAGATGGGGCTGGCCCAGTTCAAACTGGACTCTCAGTGGCTGCTGGAAGCGGGCAAGCTGGATCGCAGCAAGATCACCATCCACGCCGCCAACTCCTTCGCCACCCTGGAAGTGCCGGATGCCTACTTCGACATGGTGCGTCCGGGTGGCCTGCTCTACGGCGACTCCATCCCCTCCTACACCGAATACAAGCGGGTGATGGCATTCAAGACCCAGGTCGCCTCGGTCAACCACTACCCGGCGGGCAATACCGTTGGCTATGACCGTACCTTTACCCTCAAGCGTGAATCCTGGCTCGCCAACCTGCCGCTGGGCTACTCCGATGGCTACCGCCGTGCGCTCAGCAACAAGGCCTATGTGCTGATCCAGGGTCAGAAGGTGCCGGTGGTCGGCAAGACCTCCATGAACACCATCATGGTGGACGTCACTGATCTCAAAGGGGTGAAACCCGGTGATGAGGTGGTGCTGTTTGGCCGTCAGGGCGAGGCCGAGGTGAAACAGGCTGATCTGGAAGAGTACAACGGCGCCCTGTTAGCGGACATGTACACCATCTGGGGCTACACCAACCCCAAGAAGATCAAACGCTGA
(配列番号191)。
MHKKTLLATLIFGLLAGQAVAAPYLPLATDHRNGQEQTASNAWLEVDLGAFEHNIQTLKDRLGDKGPQICAIMKADAYGHGIDLLVPSVVKANIPCIGIASNEEARVAREKGFTGRLMRVRAATPAEVEQALPYKMEELIGSLVSAQGIADIAQRHHTNIPVHIGLNSAGMSRNGIDLRLADAKQDALAMLKLKGITPVGIMTHFPVEEKEDVKMGLAQFKLDSQWLLEAGKLDRSKITIHAANSFATLEVPDAYFDMVRPGGLLYGDSIPSYTEYKRVMAFKTQVASVNHYPAGNTVGYDRTFTLKRESWLANLPLGYSDGYRRALSNKAYVLIQGQKVPVVGKTSMNTIMVDVTDLKGVKPGDEVVLFGRQGEAEVKQADLEEYNGALLADMYTIWGYTNPKKIKR
(配列番号192)。
ATGCACAAGAAAACACTGCTGGCCACCCTGATCCTCGGCCTGCTGGCCGGGCAAGCGGTTGCAGCCCCCTACCTGCCGCTGGCCAGCGATCACCGCAACGGCGAAGTCCAGACCGCCAGCAATGCCTGGCTGGAAGTCGATCTCGGCGCCTTCGAGCACAATATCCAGACCCTCAAGGATCGTCTCGGTGACAAGGGGCCGAAGATCTGCGCCATCATGAAGGCGGATGCCTATGGCCACGGTATCGATCTGCTGGTTCCCTCGGTGGTGAAAGCGGGTATCCCCTGCATCGGTATCGCCAGCAATGAAGAAGCTCGTGTCGCCCGCGAGAAGGGCTTCACCGGTCGTCTGATGCGGGTACGTGCTGCCACCCCGGACGAAGTGGAGCAGGCCCTGCCCTACAAGATGGAGGAGCTGATCGGCAGTCTGGTGAGTGCTCAGGGCATCGCCGATATCGCCCAGCGCCACCACACCACCATTCCGGTGCATATCGCCCTCAACTCCGCCGGCATGAGCCGCAACGGCATCGATCTGCGGCTGGCCGACTCCAAGCAGGATGCGCTGGCCATGCTCAAGCTCAAGGGGATCACCCCGGTCGGCATCATGACCCACTTCCCGGTGGAGGAGAAAGAGGACGTCAAGATGGGTCTGGCCCAGTTCAAACTGGACTCCCAGTGGCTGCTGGAAGCGGGCAAGCTGGATCGCAGCAAGATCACCATCCACGCCGCCAACTCCTTCGCAACACTTGAAGTGCCGGATGCCTACTTCGACATGGTGCGCCCGGGTGGCCTGCTCTACGGTGACTCCATCCCCTCCTACACCGAGTACAAGCGGGTGATGGCGTTCAAGACCCAGGTTGCCTCCGTCAACCACTACCCGGCCGGCAACACCGTCGGTTATGACCGCACCTTCACCCTCAAGCGCGACTCCTGGCTCGCCAACCTGCCGCTCGGTTACTCCGATGGCTATCGCCGCTCCCTGAGCAACAAGGCCTATGTGCTGATCCAGGGCCAGAAGGTGCCGGTGGTCGGCAAGACCTCCATGAACACCATCATGGTGGATGTGACCGACCTGAAAGGGGTGAAACCCGGTGACGAAGTGGTGCTGTTCGGCCGTCAGGGAAATGCCGAGGTGAAGCAGGCGGATCTGGAGGAGTACAACGGCGCCCTGCTGGCGGACATGTACACCATCTGGGGCTACACCAACCCCAAGAAGATCAAGCACTAA
(配列番号193)。
MHKKTLLATLILGLLAGQAVAAPYLPLASDHRNGEVQTASNAWLEVDLGAFEHNIQTLKDRLGDKGPKICAIMKADAYGHGIDLLVPSVVKAGIPCIGIASNEEARVAREKGFTGRLMRVRAATPDEVEQALPYKMEELIGSLVSAQGIADIAQRHHTTIPVHIALNSAGMSRNGIDLRLADSKQDALAMLKLKGITPVGIMTHFPVEEKEDVKMGLAQFKLDSQWLLEAGKLDRSKITIHAANSFATLEVPDAYFDMVRPGGLLYGDSIPSYTEYKRVMAFKTQVASVNHYPAGNTVGYDRTFTLKRDSWLANLPLGYSDGYRRSLSNKAYVLIQGQKVPVVGKTSMNTIMVDVTDLKGVKPGDEVVLFGRQGNAEVKQADLEEYNGALLADMYTIWGYTNPKKIKH
(配列番号194)。
A.サルモニシダ(ATCC27013およびATCC14174)(配列番号195):
AAGGCSGATGCCTAYGGTCACGGTATCGACCTGCTGGTCCCCTCCGTGGTCAAGGCCAATATCCCCTGTATCGGCATCGCCAGCAACGAAGAGGCCCGCGTGGCGCGCGAGAAGGGGTTCAGCGGCCGCCTGATGCGGGTACGGGCCGCCACACCGATCGAAGTGGAACAGGCCCTGCCCTACAAGCTGGAAGAGCTGGTTGGCAGCCTGGTGAGTGCTCAGGGGATCTCCGACATCGCCCTGCGCCACCACACCACCATTCCGGTGCATGTCGCCCTCAACTCCGCCGGCATGAGCCGCAACGGCATCGACCTGCGTCTGGCCGATGCCAAGCAAGATGCGCTGGCCATGCTCAAGCTCAAGGGGATCACCCCGGTCGGCATCATGACCCACTTCCCGGTGGAGGAGAAAGAGGACGTCAAGCTGGGGCTGGCCCAGTTCAAGCTGGACTCCCAGTGGCTGCTGGAAGCAGGCAAGCTGGATCGCAGCAAGATCACCATCCATGCCGCCAACTCCTTCGCCACCCTGGCAGTGCCGGACGCCTACTTTGACATGGTGCGCCCGGGCGGCCTGCTCTACGGCGACTCCATCCCCTCCTACACCGAATACAAGCGGGTGATGGCATTCAAGACCCAGGTCGCCTCGGTCAACCACTATGCGGCGGGCAACACAGTCGGTTATGACCGCACCTTTACTCTCAAACGTGACTCCTGGCTCGCCAACCTGCCTCTCGGTTACTCCGAYGGCTAYCGCCG
(配列番号195)SはCまたはGおよびYはCまたはTである。
A.シューベルティー(ATCC43701)(配列番号196):
AAGGCGGATGCCTATGGTCACGGCATCGATCTGCTGGTCCCCTCCGTGATCAAGGCCGGCATTCCTTGCATCGGCATCGCCAGCAACGAAGAGGCTCGCGTCGCCCGTGAGAAGGGCTTCGAAGGCCGTCTGATGCGGGTGCGCGCCGCCACCCCGCAAGAGGTGGAAGCCGCCCTCCCCTACAAGATGGAGGAGCTGGTCGGCAGCCTGGAGAGCGCCCGTCTGATGTCGGAGATTGCCCTGCGTCACCACACCACCATTGCGTACCATCTGGGGCTCAACTCCGCCGGCATGAGCCGCAACGGCCTGGATCTGCGCCTCTCCGACGCCAAGCGCGACGCACTCGACCTGATGAAGCTCAAGGGGCTGCAGGTGGTCGGCATCATGACCCACTTCCCGGTCGAGGAGAAAGAGGACGTGAAGATGGGCTTCGCCCAGTTTCAGCTCGACACCCAGTGGCTCATCGAAGCCGCTCGTCTGGATCGCAGCAAGTTGACCCTGCACTGTGCCAACTCCTTTACCACCCTGGAGGTGCCCGAGGCCTATCTGGACATGGTCCGCCCGGGCGGCATCATCTATGGCGACACCATTCCCTCCTACACCGAATACAAGAAGGTGATGGCCTTCAAGACCCGGGTCGCCTCGGTCAATCACTACCCGAAGGGAAATAGCGTCGGCTATGACCGCACCTTCACCCTGGCACGCGACTCCTGGCTCGCCAACCTGCCGCTGGGCTACTCCGACGGCTACCGCCGGGCGCTGAGCAACAAGGCCTATGTGCTGGTGAATGGCCAGAAGGCCCCCGTGGTGGGCAAGACATCCATGAACACCATCATGGTGGACGTGACCGACATCAAGGGGGTCAAACCGGGTGACGAGGTGGTGCTGTTTGGCCGCCAGGGCAACGCCGAGGTGAAGCAGTCCGATCTCGAGGAGTACAACGGCGCCCTCCTGGCGGACATGTACACCATCTGGGGCTACACCAATCCACGTATCATCAAGCGCTGA
A.ジャンデイ:
順方向NdeI 5'-CCGGAACCTTCATATGCACAAGAAAACACTGCTGGCCAC-3'(配列番号197)および
逆方向XhoI 5'-TTCCAAGGCTCGAGGTGCTTGATCTTCTTGGGGTTGGT-3'(配列番号198)。
A.ソブリア
順方向NdeI 5'-CCGGAACCTTCATATGCACAAGAAAACGCTATTGGCCAC-3'(配列番号199)および
逆方向XhoI 5'-TTCCAAGGCTCGAGGCGTTTGATCTTCTTGGGGTTGGT-3'(配列番号200)。
イー・コリで生成した、P.タエトロレンス(配列番号204)、A.キャビエ(配列番号179)、およびA.ハイドロフィラ(配列番号177)に由来する精製C末端Hisタグ付き広域アミノ酸ラセマーゼ(BAR)の試料を逆相液体クロマトグラフィー/エレクトロスプレーイオン化四重飛行時間型質量分析(LC/ESI−QTOF−MS)によって特徴づけ、各タンパク質のリーダー配列がどの位置で開裂するかを決定した。質量スペクトルは、m/z 300〜m/z 1300の範囲にわたり収集した。各タンパク質について獲得した質量スペクトルのデコンボリューション(結合した異なる数のプロトンを有する同じ質量のタンパク質分子に起因するm/z値の包絡線の分子量への数学的な変換)はMS計測装置を備えた最大エントロピーデコンボリューションソフトウェアを用いて達成した。P.タエトロレンスBARについて観察された分子量は42651Daであり、このタンパク質がアミノ酸A23の後で開裂することを示した(理論的な分子量=42654Da)。A.キャビエBARについて観察された分子量は43565Daであり、このタンパク質がアミノ酸21の後で開裂することを示した(理論的な分子量=43568Da)。A.ハイドロフィラBARについて観察された分子量は43450Daであり、このタンパク質がアミノ酸21の後で開裂することを示した(理論的な分子量=43453Da)。配列番号204のアミノ酸2〜23をコードするコドンが除去され、部分的な遺伝子が上記の方法と同じようにイー・コリで発現する場合、精製発現産物について観察された分子量は42483Daである。この結果に基づいて、リーダーなし発現産物の成熟タンパク質は、最初の2つまたは3つのアミノ酸が除去されているように考えられ、それを、野生型産物の成熟タンパク質よりも1つまたは2つのアミノ酸分短くしている。配列番号179のアミノ酸2〜21をコードするコドンが除去された場合、精製発現産物について観察された分子量は43500であり、最初の2つのアミノ酸(MA)が除去されていることに相応する(理論的な分子量=43498)。したがって、成熟タンパク質は、野生型成熟タンパク質と比較して1つのさらなるアラニンを欠いており、実施例4Oに示されるように、周辺質中に分泌されるようには考えられない。A.キャビエおよびP.タエトロレンスの双方のリーダーなし発現産物から開裂するアラニンに加えて、P.タエトロレンス(配列番号204)リーダーなし発現産物に由来する成熟タンパク質がプロリンを欠けている場合、それは、活性の低下の増加について説明する可能性がある。しかしながら、SDS−PAGEによって判断されるように発現の劇的な低下によって証拠づけられるように、タンパク質は、決して適切に発現せず、折りたたまれない可能性がある。N末端Hisタグ付きタンパク質はニッケルアフィニティーカラム上で精製されないと考えられ、Signal P 3.0プログラムはリーダー配列を予測したので、上記に記載されるP.プチダBAR酵素は、イー・コリによって同様に処理されるであろう。
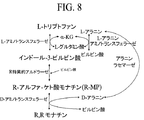
実施例4A(5)、9、および10(3)に記載され、かつ図1〜8に示されるプロセスの多くは、インドール−3−ピルビン酸およびピルビン酸からR−MPを優先的に生成するアルドラーゼで最適に働くであろう。従って、R−MPを優先的に生成するアルドラーゼをコードする核酸を含有するクローンを単離し、試験するための方法が記載される。炭素源としてのリボースを有するM9最少培地上で成長させる場合にピルビン酸の補足を必要とするエシェリキア・コリの株は先に記載された。Ponce,E.ら,「Cloning of the two pyruvate kinase isoenzymes structural genes from Escherichia coli.:The relative roles of these enzymes in pyruvate biosynthesis」,J.Bacteriol.177:5719−5722,(1995)。株の関連する遺伝子型は次の通りである:ΔpykA ΔpykF。ダブルノックアウトは、DatsenkoおよびWanner,Proceed.Natl.Acad.Sci.USA 97:6640−6645,(2000)の方法によって生成した。これらの株は、ピルビン酸生成アルドラーゼスクリーニングのためのおよびモナチンの特定の立体異性体、モナチン前駆体の特定の立体異性体、またはモナチンもしくはモナチン前駆体の類似体に対する活性が高いアルドラーゼをスクリーニングするための基礎を形成できる。モナチン前駆体の類似体は、4−ヒドロキシ−4−メチル−2−オキソグルタル酸、4−カルボキシ−4−ヒドロキシ−2−オキソアジペート(oxoadipate)、4−ヒドロキシ−4−メチル−2−オキソアジペート、またはアルドール反応においてピルビン酸に変換される他のカルボキシルリッチ化合物のごときProAアルドラーゼまたはKHGアルドラーゼの基質として同定された化合物を含む。用いることができるモナチンの類似体の例は、4−ヒドロキシ−4−メチルグルタミン酸であり、これは、試験細胞中の天然のアミノトランスフェラーゼによって4−ヒドロキシ−4−メチル−2−オキソグルタル酸(ProAの基質)に容易にアミノ基転移され得る。
以下のプライマーは、pykAノックアウトを生成するために用いた。
5'-ATGTCCAGAAGGCTTCGCAGAACAAAAATCGTTACCACGTTAGGTGTAGGCTGGAGCTGCTTC-3'(配列番号3)および
5'-CTCTACCGTTAAAATACGCGTGGTATTAGTAGAACCCACGGTACCATATGAATATCCTCCTTAG-3'(配列番号4)。
5'-AGGACGTGAACAGATGCGGTGTTAGTAGTGCCGCTCGGTACCAGCATATGAATATCCTCCTTAG-3'(配列番号5)および
5'-ATGAAAAAGACCAAAATTGTTTGCACCATCGGACCGAAAACCGGTGTAGGCTGGAGCTGCTTC-3'(配列番号6)。
ダブルノックアウトは、Balchのビタミン溶液、Balchの修正微量元素溶液(Balch,W.E.ら,「Methanogens:reevaluation of a unique biological group」,Microbiol.Rev.43:260−296,(1979))、および0.4%D−リボースを補足した最少培地(M9塩)(Difco社)上での成長について試験した。5mMピルビン酸が培地にさらに含まれていない限り、成長は、ダブル変異体について見られなかった。野生型MG1655は、ピルビン酸有りおよびなしの双方で上記の培地上で成長した。ダブルノックアウトは、リボースではなく0.4%グルコースを補足した上記に記載される最少培地上での成長について試験した。この培地上での成長は、野生型株を用いて見出されたものに類似していた。この培地で、ピルビン酸は、ptsI遺伝子産物(ホスホエノールピルビン酸からピルビン酸を作製し、グルコースにリン酸を移すホスホトランスフェラーゼ系の酵素)を介してグルコースから生成され得る。ダブルノックアウト株は、さらに、リボースではなく0.4%L−アラビノースまたは0.4%D−キシロースを補足した上記に記載される培地を用いて、成長について試験した。ピルビン酸は、これらの5−炭素含有(非PTS)基質での成長から生成されない。5mMピルビン酸を補足しない限り、ダブルノックアウトはこれらの条件下で成長せず、野生型株は、ピルビン酸有りおよびなしの双方で正常に成長した。
実験概要
イー・コリAspC(HEXaspC)のhexa変異体は、国際公開第03/091396A2号の実施例6に記載のようにS,Sモナチンの生成についてAspCと比較してより優れた活性を有することがわかった。HEX(受入番号:/AHFA gi:127190)は、AspCからの以下の突然変異を含有する(イー・コリ番号付け):V35L、K37Y、T43I、N64L、TI04S、およびN285S。構造分析および文献報告(Rothman,S.およびKirsch,J.,J.Mol.Biol.327:593−608,(2003);Rothman,S.ら,Protein Science 13:763−772,(2004))に基づき、モナチン生成経路に利用される基質:L−トリプトファン、S−MP、またはその双方に対して速度論的活性を増加させると予想された5種以上の変異体を作り出した。変異体のうちの2種は、トリプトファンおよびS,Sモナチンの双方のアミノ基転移速度を増加させた。変異体のうちの2種は、S,Sモナチンの形成に対する立体選択性の増加を示したが、一方はそれほど立体選択性ではなかった。これに基づき、同様の突然変異を有するバチルス種からの広域特異性D−アミノトランスフェラーゼは、図3に示され、かつ実施例4(4)に記載されるR,Rモナチン経路のD−アミノトランスフェラーゼとして有用であろうことが予想される。変異体のうちの1種(HEXaspCP9T/R122G)は、L−トリプトファンアミノ基転移に対する活性を増加させたが、S,Sモナチン生成またはS,Sモナチンアミノ基転移における活性は著しく減少した。従って、この酵素は、図1、2、4、5、6、7、および8に示され、かつ実施例9および10(3)に記載されるR,Rモナチン生成経路の第1の工程に有用であることが予想される。一般的に、L−トリプトファンに対するAspCの活性に類似する活性ならびにR−MPおよびS−MPに対する制限された活性を有するアミノトランスフェラーゼは、図1、2、4、5、6、7、および8に表すプロセスに有用であろう。
pUC19中でクローニングしたHEX遺伝子は、J.F.Kirsch教授(Department of Molecular and Cell Biology,University of California,バークリー,バークリー,CA 94720−3206)によって提供されたものであり、pET23a中への遺伝子のクローニングのために鋳型として用いた。Onuffer,J.J.およびKirsch,J.F.,「Redesign of the substrate specificity of Escherichia coli aspartate aminotransferase to that of Escherichia coli tyrosine aminotransferase by homology modeling and site−directed mutagenesis」,Protein Science 4:1750−1757(1995)を参照されたい。NCBI受入番号1AHF_A GI:1127190(HEXアミノ酸配列)もまた参照されたい。以下のプライマーは、pET23aベクター(Novagen社、マディソン、WI)中にHEX遺伝子をクローニングするために設計した。
HEXaspCプライマー:
N末端:5'-GCGGAACATATGTTTGAGAACATTACCGCC-3'(配列番号7);
C末端:5'-ATAACCGGATCCTTACAGCACTGCCACAATCG-3'(配列番号8)。
Novagen Overnight Express(商標)Autoinduction System 2(カタログ#71366−3;溶液1〜6の液体培養物(5mL)は、以下の株の新鮮な平板培養物または凍結グリセロールストックから接種した。
E.coli BL21(DE3)::HEXaspCpET23a
E.coli BL21(DE3)::HEXaspCW130FpET23a
E.coli BL21(DE3)::HEXaspCT156ApET23a
E.coli BL21(DE3)::HEXaspCP9T/T156ApET23a
E.coli BL21(DE3)::HEXaspCP9T/R122GpET23a
E.coli BL21(DE3)::HEXaspCR122G/T156ApET23a
変異体は、基質としてS,SモナチンおよびL−トリプトファンを用いるアミノ基転移活性について試験した。アミノトランスフェラーゼ活性は、実施例1に記載のように、OPAポストカラム誘導体化を用いたHPLCによって、反応の副産物であるグルタミン酸の形成に従って測定した。反応混合物は、1.0mL中、100mM HEPPS緩衝液、pH8.0、20mMアルファ−ケトグルタル酸、0.08mMピリドキサールリン酸、25mMトリプトファンまたはS,Sモナチン、および酵素(細胞抽出物タンパク質における2.5mgとして供給)を含有した。酵素以外の成分はすべて、一緒に混合した。酵素は、反応を開始するために追加し、反応溶液は、90分間、30℃(緩やかに振盪させる)でインキュベートした。反応は、酵素を追加しないネガティブコントロールと共に二通り行った。反応は、10%ギ酸(最終濃度)の追加によって停止させ、混合物は、21,000rpmで遠心分離し、上清は注意深く取り出し、濾過した。データは、グルタミン酸のバックグラウンドレベルについておよびタンパク質を沈殿させるための、酸の追加による希釈について補正し、次いで、追加した変異体アミノトランスフェラーゼの量によって標準化した。トリプトファンを基質として利用した場合、HEXaspCは、毎時1mgのアミノトランスフェラーゼ当たり13.0mMグルタミン酸を生成した。変異体の、百分率として表した相対的活性は、以下の通りである:HEXaspCW130F(156%)、HEXaspCT156A(151%)、HEXaspCP9T/T156A(63.7%)、HEXaspCP9T/R122G(116%)、およびHEXaspCR122G/T156A(107%)。S,Sモナチンを基質として利用した場合、HEXaspCは、毎時1mgのアミノトランスフェラーゼ当たり7.43mMグルタミン酸を生成した。変異体の、百分率として表した相対的活性は、以下の通りである:HEXaspCW130F(113%)、HEXaspCT156A(87.7%)、HEXaspCP9T/T156A(67.3%)、HEXaspCP9T/R122G(11.2%)、およびHEXaspCR122G/T156A(114%)。
実験概要
バチルス株ATCC4978からの新規なD−アミノトランスフェラーゼ遺伝子(実施例15に記載される)は、部位特異的方法を用いて突然変異誘発させた。変異体遺伝子を発現させ、モナチン生成経路について注目する活性についてアッセイした。
突然変異誘発のためのプライマーは、Stratagene Multi−Changeキット(La Jolla、CA)に挙げられた示唆に従って設計した。プライマーは5’−リン酸化した。突然変異誘発は、メーカーの指示に従って、Stratagene Multi−Changeキットを用いて行った。突然変異誘発のために用いた鋳型は、実施例15に記載されるpET30(タグなし)またはpET28(タグ付き)の4978DAT構築物のいずれかとした。プライマーは表37で下記に挙げる。
適正な突然変異を含有するプラスミドDNAをイー・コリBL21(DE3)コンピテント細胞中に形質転換し、実施例16に記載されるように誘発した。細胞抽出物は、BugBuster試薬およびBenzonase Nuclease(EMD Biosciences/Novagen、Madison、WI)を用いて調製した。1mlのアッセイは、緩やかに振盪させながら30℃で行った、また10.2mg D−トリプトファン、0.05mM PLP、4mM MgCl2、100mMリン酸カリウム緩衝液pH7.5、約50μgのアルドラーゼ、200mMピルビン酸、および細胞抽出物として供給された0.150〜0.5mg/mL D−アミノトランスフェラーゼを含有した。全タンパク質アッセイは、Bio−Rad(Hercules、CA)全タンパク質キット(Coomassie)またはPierce(Rockford、IL)BCAキットを用いて行い、D−アミノトランスフェラーゼの発現パーセントは、SDS−PAGEまたはBio−Rad Experion Automated Electrophoresis Systemによって評価した。試料は、3時間および一晩の時点で測った。
S180A/S181A/S182R;
L151F;
V34G
S181R
A153R/S181A/S182A;
A153R/S182A;
A153R/S182G;
S180R/S181A/S182G;
S180R/S181A/S182A;
S180R/S181G/S182G;
S180G/S181R/S182G;および
S180A/S181R/S182A。
F200M 4978 DAT変異体を作り出すために、実施例15からの野生型4978 DATオープンリーディングフレーム(タグ付き)は、プライマー73および80ならびにPfuTurbo(登録商標)DNA Polymerase(Stratagene)を用いて増幅し、pCRII−Blunt(Invitrogen、Carlsbad、CA)中にクローニングし、その配列を検証した(Agencourt、Beverly、MA)。5’領域および3’領域は、プライマー80および96および99および103をそれぞれ用いて増幅した(表42)。次いで、増幅したDNAは、Qiagen QIAquick(登録商標)Gel Extraction Kit(Qiagen、Valencia、CA)を用いてゲル精製した。それらを組み合わせ、プライマー80および99を用いてPCRにさらにかけた(表42)。増幅したDNAは、上記に記載されるようにゲル精製し、pCRII Blunt中にクローニングし、その配列を検証した。DATオープンリーディングフレームは、pET28b中にNdeI/XhoI制限消化物断片としてサブクローニングした。
500mLバッフル付きフラスコ中に、50μg/mLカナマイシンを有する100mL LBを含有する培養物に、1mL一晩培養物を接種し、約0.6の光学濃度(600nm)まで37℃で成長させた。タンパク質の生成は、1mMの最終濃度でIPTGによって誘発した。細胞は、IPTGの追加後、4.5時間30℃でインキュベートした。細胞は遠心分離し、−80℃で凍結した。細胞は、破壊し(Novagen推奨のプロトコールに従って、1μL/mLベンゾナーゼヌクレアーゼ、5μL/mLプロテアーゼインヒビターカクテルII、および0.033μL/mL rLysozymeを含有するNovagen BugBuster試薬を用いて調製)、SDS−PAGEによって分析した。変異体(141−LRcD−144→EYcY)および(243−TS−244→NR)はそれらが調製された条件下で不溶性のタンパク質をもたらした。変異体243−TS−244→NKは、試験した条件下で数量化できる活性を有していなかった、また、おそらく、S244K変異体のように、野生型と比較して活性の弱い酵素である。
実施例6Bに示されるように、T243N変異体DATの初期活性はB.スファエリクスDATよりも有意に高いが、活性はより急速に減少する。下記に記載される嫌気性プロトコールを用いるさらなる実験は、T243N変異体DATの初期活性はB.スファエリクスDATよりも8倍まで高かった、しかしながら活性は、嫌気条件下でさえ急速に減少したことを示した。以下の研究は、長期間のより高い活性の維持を試みるために行った。
AT−102およびAT−104は、BioCatalytics社(パサデナ、CA)から購入した分岐鎖L−トランスアミナーゼ(EC2.6.1.42)である。酵素は、化学的に生成したS,SおよびR,Rモナチンの基質を用いて、アミノ基転移活性について試験した。反応は、0.5mLの全容量で行い、二通り実行した。アッセイは、50mM Tris pH7.8、0.08mM PLP、10mM α−ケトグルタル酸(「α−KG」)、5mMモナチン、および1mg/mLアミノトランスフェラーゼ酵素を含有した。ネガティブコントロールは外来性アミノトランスフェラーゼ酵素を含有しなかった。試料は、100rpmの振盪で30℃で2時間インキュベートした。試料は濾過し、LC/MS/MS分析は、実施例1に記載のように、グルタミン酸レベルを確認するために実行した。グルタミン酸レベルは、MPの生成と理論量的に関連するはずである。R,Rを反応基質として用いた場合、非常に低いレベルのグルタミン酸がネガティブコントロール中に存在した。AT−104は、ネガティブコントロールよりもわずかに多くのグルタミン酸を生成し、R,Rモナチン基質(D−アミノ酸)との低いレベルの活性を示した。双方の分岐鎖L−アミノトランスフェラーゼはS,Sモナチンに対して活性を示した。AT−102は102μg/mLグルタミン酸を生成し、AT−104は64μg/mLグルタミン酸を生成した。比較については、広域特異性アミノトランスフェラーゼ(AT−101、同様にBioCatalytics社からのもの)は、これらの条件下で75μg/mLを生成した。モナチンは、広域特異性アミノトランスフェラーゼまたはアスパラギン酸アミノトランスフェラーゼに対する基質として通常役立つジカルボキシルアミノ酸および芳香族アミノ酸に対してより構造類似性を有するので、分岐鎖アミノトランスフェラーゼでの高活性はやや予想外である。しかしながら、モナチンのグルタミン酸主鎖により、アミノ供与体としてグルタミン酸を利用できる多くのアミノトランスフェラーゼもまたモナチンに対して活性を有する可能性がある。
AT−102およびAT−104は、C.テストステローニ(国際公開第03091396A2号に記載のように生成)からのProAアルドラーゼを用いる、共役反応におけるモナチンの生成について試験した。酵素および付加的な成分/基質を、100mMリン酸カリウム緩衝液pH7.5、100mM L−グルタミン酸、および0.1mM PLPを含有したキット中に提供された反応緩衝液に直接的に追加した。1mLの反応緩衝液に、以下のものを追加した:4mgインドール−3−ピルビン酸、20mgピルビン酸、細胞抽出物中に提供される約50μg ProA、1μL 2M MgCl2、および試験されることになる2mgのアミノトランスフェラーゼ酵素。反応はすべて二通り行い、ネガティブコントロール反応は、付加的なアミノトランスフェラーゼを追加せずに行った。ポジティブコントロール(AT−101)は比較のために利用した。この酵素は広域特異性L−アミノトランスフェラーゼである。モナチンのバックグラウンド生成は、組換えProA酵素の細胞性抽出物中に存在する天然のイー・コリアミノトランスフェラーゼによるものである。反応は、緩やかに振盪させながら(100rpm)、30℃で一晩インキュベートした。試料は、実施例1に記載のように濾過し、逆相LC/MS/MS分析にかけた。結果を下記の表47に示す。
バチルス・リケニホルミス(Bacillus licheniformis)は、イー・コリ分岐鎖アミノトランスフェラーゼよりも、D−アミノトランスフェラーゼとより密接に関連する推定上の分岐鎖アミノトランスフェラーゼを含有する。それを、D−アミノ基転移活性についてアッセイし、イー・コリBCATについて上記に言及した、予測された活性部位残基に基づいて突然変異誘発した。
B.リケニホルミス(ATCC番号14580)は、一晩30℃で栄養寒天上で成長させた。コロニーのグループを100μLの滅菌水中に置き、細胞を破壊するために95℃で10分間加熱した。3μLを、続くポリメラーゼ連鎖反応(PCR)による増幅において用いた。
プライマーは、NcoI部位ならびにSalI部位を用いて、pET28bベクターおよびpET30aベクター(Novagen社、マディソン、WI)ならびにpTRC99a(GE Healthcare Life Sciences社)にクローニングするためにB.リケニホルミス遺伝子(915bp)について設計した。pET30構築物はN末端HisタグおよびSタグを含有するが、pET28構築物はタグ付きではない。
N末端5'-GGTTAAGGCCATGGGGGACCAGAAAGACCA-3'(配列番号44);およびC末端:5'-GGCCTTCCGTCGACTCAGCTGACACTTAAGCT-3'(配列番号45)。
PCR生成物は、精製し、SalI緩衝液(New England Biolabs社、イプスウィッチ、MA)中でSalIおよびNcoIを用いて消化した。消化したベクター(pET28、pET30、およびpTRC99a)ならびに挿入断片は、Qiagen QIAquick(登録商標)Gel Extraction Kit(Qiagen社、バレンシア、CA)を用いて精製した。ライゲーションは、Roche社製Rapid DNA Ligation Kit(Roche社、インディアナポリス、IN)を用いて行い、精製した。ライゲーションしたものは、Bio−Rad社エレクトロポレーションマニュアルに記載のように、0.2cmキュベットおよびBio−Rad社製Gene Pulser(登録商標)IIシステムを用いてエシェリキア・コリDH10Bに形質転換した。細胞は、225rpmで振盪させながら37℃で30分間、900μL SOC培地中で回復させた。細胞は、カナマイシン(25μg/mL)を含有するLB寒天平板上で平板培養した。プラスミドDNAは、Qiagen社製スピンミニプレップキット(Qiagen社、バレンシア、CA)を用いて精製し、SalIおよびNcoIを用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するようであるプラスミドの配列は、Agencourt BioScience社(ビバリー、MA)でジデオキシ連鎖停止DNA配列決定によって検証した。配列決定により、NCBI受入番号AAU24468 GI:52004526に挙げられるアミノ酸配列を有するタンパク質を生成する受入番号CP000002 GI56160984 2851268..2850354に見られるコード配列を検証した。
プラスミドDNA(pETベクター)は、pETベクター中の構築物のために、イー・コリ発現宿主BL21(DE3)細胞(Novagen社、マディソン、WI)中に形質転換した。培養物を成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。誘発は、通常、カナマイシン(50μg/mL)を含有するLB培地中で行った。細胞は、37℃で0.4〜0.8のOD600まで成長させ、0.1mM IPTG(イソプロピルチオガラクトシド)を用いて誘発し、誘発の3〜4時間後にサンプリングした。細胞抽出物は、Novagen社製BugBuster(商標)試薬付属のプロトコールに従って調製した(ベンゾナーゼヌクレアーゼおよびRoche社製完全プロテアーゼインヒビターカクテルを追加)。高いレベルの可溶性タンパク質が、SDS−PAGEで判断されるように、予測した分子量で得られた。細胞性抽出物中の可溶性タンパク質はSDS−PAGEによって分離した。
本実施例は、L−グルタミン酸およびD−グルタミン酸(またはL−およびD−アスパラギン酸またはL−およびD−アラニン)の間で相互変換するために用いることができるアミノ酸ラセマーゼ酵素をクローニングし、試験するために用いる方法を記載する。グルタミン酸ラセマーゼ、アスパラギン酸ラセマーゼ、またはアラニンラセマーゼは、経路における工程がL−アミノ酸(例えば、L−グルタミン酸、L−アスパラギン酸、またはL−アラニン)を生成し、経路の他の工程がD−アミノ酸(例えば、D−グルタミン酸、D−アスパラギン酸、またはD−アラニン)を消費する場合、R,Rモナチンを生成するための生合成経路に有用である。図4は、L−トリプトファン特異的アミノトランスフェラーゼ、R特異的アルドラーゼ、D−アミノトランスフェラーゼ、およびグルタミン酸(またはアスパラギン酸またはアラニン)ラセマーゼを用いる、L−トリプトファンからR,Rモナチンを生成するための生合成経路を示す。
ラクトバチルス・ブレビス(Genbank受入番号D29627、核酸配列)およびペディオコッカス・ペントサセウス(murI遺伝子)(Genbank受入番号L22789)からのグルタミン酸ラセマーゼ(EC5.1.1.3)をコードする遺伝子は、イー・コリ中でクローニングし、発現させた:その抽出物は、D−グルタミン酸へのL−グルタミン酸の変換およびL−グルタミン酸へのD−グルタミン酸の変換における活性について試験した。BioCatalytics社製アスパラギン酸ラセマーゼ酵素(EC5.1.1.13)もまたL−およびD−アスパラギン酸の間の相互転換について試験した。
L.ブレビスゲノムDNA(ATCC8287D)は、American Type Culture Collectionから得た。P.ペントサセウス(ATCC25745)は、ラクトバチルスMRSブロス中で37℃で成長させ、2mlは、Mekalanos,J.J.,「Duplication and amplification of toxin genes in Vibrio cholerae」,Cell 35:253−263,(1983)の方法を用いるゲノムDNA単離に用いた。
プライマーは、pET28ベクターおよびpET30ベクター(Novagen社、マディソン、WI)中にクローニングするために5’制限部位およびオーバーハングを用いて設計した。
N末端:5'-GCGGCGCCATGGAAAATGATCCGATTGGTCTAATG-3'(配列番号15)、および
C末端:5'-GCGGCGGTCGACGCAATTACAATTGTGTTTGTC-3'(配列番号16)。
N末端:5'-GCGGCGCCATGGATGTATGTATAATTTTATTTAG-3'(配列番号17)、および
C末端:5'-GCGGCGGTCGACAAATTTCATTATTCATTCTAATTT-3'(配列番号18)。
PCR産物は、Qiagen社製ゲル抽出キット(Qiagen社、バレンシア、CA)を用いて、0.8%TAE−アガロースゲルからゲル精製した。PCR産物は、SmartSpec 3000(商標)分光光度計を用いて定量した。産物は、メーカー推奨のプロトコール(New England Biolabs社、ビバリー、MA)に従って制限酵素NcoIおよびSalIを用いて消化し、Qiagen社製ゲル抽出キット(Qiagen社、バレンシア、CA)を用いて、0.8%TAE−アガロースゲルからゲル精製した。ベクターpET28およびpET30は、制限酵素NcoIおよびSalIを用いた消化によって調製し、その後、エビアルカリフォスファターゼを用いる処理およびQiagen社製ゲル抽出キット(Qiagen社、バレンシア、CA)を用いる0.8%TAE−アガロースゲルからの精製が続いた。
配列分析によって検証したプラスミドDNAは、イー・コリ発現宿主BL21(DE3)(Novagen社、マディソン、WI)中にサブクローニングした。培養物を成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。
本実施例は、L−トリプトファン(L−チロシンまたは芳香族)アミノトランスフェラーゼ、ProAアルドラーゼ、アラニンラセマーゼ、グルタミン酸ラセマーゼ、またはアスパラギン酸ラセマーゼ、および広域特異性D−アミノ酸アミノトランスフェラーゼを用いて、L−トリプトファンから立体異性的に豊富なR,Rモナチンを生成する方法を記載する。図5は経路を示す図である。立体異性的に豊富なR,Rモナチンの生成へのこのアプローチは、モナチン前駆体(MP)からのモナチンの生成において低い活性を有する、工程1の酵素を必要とする。前の結果に基づき、発明者らは、国際公開第03/091396A2号の実施例1に記載されるシノリゾビウム・メリロティおよびロドバクター・スフェロイデスのtatA遺伝子産物を用いた。
L.ブレビスおよびP.ペントサセウスからのグルタミン酸ラセマーゼは、実施例8に記載のようにイー・コリ中で生成した。いくつかの場合では、これらの酵素のHis6−タグ付きバージョンは、メーカーのプロトコール(Novagen社、マディソン、WI)に従ってHis−Bind900カートリッジを用いて精製し、PD−10カラム(G25 Sephadex、Amersham−Pharmacia社)を用いて、脱塩し、イミダゾールを除去した。酵素は、25mMリン酸カリウムpH8.0中に溶出した。アスパラギン酸ラセマーゼ(ASPR−101)およびD−アミノトランスフェラーゼ(AT−103)はBioCatalytics社から購入し、アラニンラセマーゼはシグマ社(セントルイス、MO)(カタログ番号A8936)から購入した。S.メリロティおよびR.スフェロイデスのチロシン(芳香族)アミノトランスフェラーゼは、国際公開第03/091396A2号の実施例1に記載のように調製した。コマモナス・テストステローニProAアルドラーゼは国際公開第03/091396A2号の実施例4に記載のように調製した。全タンパク質アッセイは、メーカーのプロトコールに従ってBio−Rad社製タンパク質アッセイ(ヘラクレス、CA)を利用して行った。
反応混合物(1mL容量、二通り実行)は、100mMリン酸カリウム緩衝液(pH8)、2mM MgCl2、0.05mMピリドキサール−5’−リン酸(PLP)、200mMピルビン酸ナトリウム、5mM α−ケトグルタル酸ナトリウム、またはオキサロ酢酸塩、細胞抽出物において供給された約280μg/mL S.メリロティTatA、1mg/mL BioCatalytics社製D−アミノトランスフェラーゼ(AT−103)、100μL/mLのグルタミン酸ラセマーゼ細胞抽出物または1mg/mLアスパラギン酸ラセマーゼ、および細胞抽出物として提供された約100μg/mLのProAアルドラーゼを含有した。固体トリプトファンは10.2mg/mlの濃度で追加した。ネガティブコントロールはラセマーゼを含有しなかった。試料は、1時間、2時間、または一晩30℃(250rpmで振盪しながら)でインキュベートした。試料は、沈澱物を除去するために遠心分離し、シリンジで濾過し、実施例1に記載されるLC/MS/MS法を用いる、モナチンについての分析前に−80℃で保存した。
上記実験は、約54μgの精製L−アミノトランスフェラーゼ(S.メリロティまたはR.スフェロイデスのTatAのいずれか)、1mgアスパラギン酸アミノトランスフェラーゼ(BioCatalytics社)、1mgD−アミノトランスフェラーゼ、アミノ受容体としての5mMオキサロ酢酸塩、および75μg精製アルドラーゼを用いて繰り返した。反応は、2時間のサンプリング時間および一晩のインキュベーション期間で二通り実行した。ネガティブコントロールは、ラセマーゼを用いず、S.メリロティ L−アミノトランスフェラーゼを用いて行った。逆相クロマトグラフィーに基づくR,R/S,SおよびS,R/R,Sモナチンピーク定量化の定量化に加えて、各立体異性体の百分率は、実施例1に記載されるFDAA誘導体化技術を用いて決定した。結果を下記の表53に示す。
モナチン生成は、C.テストステローニからのProAアルドラーゼを用いて試験した。約50μgの精製L−アミノトランスフェラーゼ(S.メリロティ TatA)、1mg D−アミノトランスフェラーゼ(AT−103、BioCatalytics社)、アミノ受容体としてのピルビン酸、50μg精製アルドラーゼ、およびシグマ社(セントルイス、MO)から購入した70μgアラニンラセマーゼ(カタログ番号A8936)。反応は二通り実行し、一晩インキュベートした。各立体異性体の百分率は、実施例1に記載されるFDAA誘導体化技術を用いて決定した。ラセマーゼのないコントロールを含んだ。結果を下記の表56に示す。
図3に示されるように、立体反転アミノトランスフェラーゼは、モナチンの生成のための生合成経路で有用である。例えば、D−フェニルグリシンアミノトランスフェラーゼまたはその変異体は、アミノ供与体としてL−グルタミン酸を用いてR−MPからR,Rモナチンを生成できる。
本実施例は、アミノ供与体としてL−グルタミン酸を用いて、Rモナチン前駆体をR,Rモナチンに変換するために用いることができる立体反転酵素である4D−ヒドロキシフェニルグリシンアミノトランスフェラーゼを合成するために用いられる方法を記載する。
シュードモナス・スタッツェリ4D−ヒドロキシフェニルグリシンアミノトランスフェラーゼ(4D−HPG AT)についての公開配列(Genbank受入番号AY319935、核酸配列;Genbank受入番号AAQ8290、タンパク質配列)をPCRプライマー設計のための鋳型として用いた。あるいは、シュードモナス・プチダ(CAD42450(タンパク質)、AX467211(ヌクレオチド))からの4−D−ヒドロキシフェニルグリシンアミノトランスフェラーゼを配列鋳型として用いる。合計34個の順方向プライマーおよび35個の逆方向プライマーを設計した。順方向プライマーおよび逆方向プライマーは、40塩基長であり、20個のオーバーラップ塩基対を共有した。さらに、2つの外部プライマーは、pET28ベクターおよびpET30ベクター(Novagen社、マディソン、WI)中にクローニングするために5’制限部位およびオーバーハングを用いて設計した。
5'-GGCCGGCATATGTCGATCCTTAACGACTACAAACGT-3'(配列番号19)、およびC末端(XhoI部位を有する):
5'-GGAAGGCTCGAGTCATGATTGGTTTCCAGACAAATT-3'(配列番号20)。
P.スタッツェリからの遺伝子配列は、以下のプロトコールを用いて増幅した。一次100μL PCR反応は、0.05μMのそれぞれの69個の内部プライマー、0.4mM各dNTP、10U rTth PolymeraseXL(Roche社、インディアナポリス、IN)、0.625U Pfuポリメラーゼ(Stratagene社、ラ ホーヤ、CA)、1×XL緩衝液、および1mM Mg(OAc)2を含んだ。用いられるサーモサイクラープログラムは、3分間94℃でのホットスタートを含み、以下の工程を15回繰り返し:30秒間94℃、30秒間42℃、および15秒間68℃、その後、以下の工程を10回繰り返し:30秒間94℃、30秒間52℃、および30秒間68℃、その後、以下の工程を10回繰り返した:30秒間94℃、30秒間60℃、および1分15秒間68℃。最終の10サイクルの後、試料は、7分間68℃で維持し、次いで、4℃で保存した。このPCRプロトコールは、0.8%TAE−アガロースゲル上で約0.5kbに産物のスメアを生成した。
プラスミドDNAは、イー・コリ発現宿主BL21(DE3)(Novagen社、マディソン、WI)中に形質転換した。培養物を成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。
立体反転D−フェニルグリシンアミノトランスフェラーゼ(D−4−ヒドロキシフェニルグリシンアミノトランスフェラーゼとも呼ばれる)を有するシュードモナス属および同様な属の生物は、以下の様式で単離する。土壌試料は、以下の培地を有するペトリ皿上でインキュベートする:(1リットル当たり)15g寒天、3.4g KH2PO4、3.55g Na2HPO4、0.2g MgSO4・7H2O、8mg CaCl2・2H2O、10mg酵母抽出物、1ml 1000×微量元素溶液(Balch,W.E.ら,「Methanogens:reevaluation of a unique biological group」,Microbiol.Rev.43:260−296,(1979))、および1g D−フェニルグリシン(D−4−ヒドロキシフェニルグリシン)。
上記の(1)および(2)に記載されるD−ヒドロキシフェニルグリシンアミノトランスフェラーゼは粗無細胞タンパク質抽出物において用いるまたは上記の(1)に記載のように精製する。S.メリロティおよびR.スフェロイデスのチロシン(芳香族)アミノトランスフェラーゼは、国際公開第03/091396A2号の実施例1に記載のように調製する。コマモナス・テストステローニProAアルドラーゼは国際公開第03/091396A2号の実施例4に記載のように調製する。全タンパク質アッセイは、メーカーのプロトコールに従ってBio−Rad社製Protein Assayを利用して行う(ヘラクレス、CA)。
DPGATのHisタグなしバージョンは、上記の(1)に記載のように発現させ、抽出物はアッセイに用いた。アッセイを設定した、またアッセイは、100mMリン酸カリウムpH7.0、60mM D−フェニルグリシン、5mM α−ケトグルタル酸、および50μMピリドキサール−5’−リン酸を含んだ。アッセイは、アッセイ容量の1ml当たりに、本実施例において上記に記載のように調製した100μLの抽出物を追加することにより開始した。試料は、数回の時点(0、1、2、5、10、30、60、および120分)で取り、等容量の2Mギ酸を用いて停止した。試料はまた、一晩のインキュベーション(約1200分)の後にも取った。試料は、実施例1に記載されるLC/ポストカラム蛍光検出(OPA)法によってグルタミン酸生成について分析した。結果を下記の表57に要約する。
背景
D−メチオニン−ピルビン酸アミノトランスフェラーゼ(EC2.6.1.41)は、まれであるが、立体反転トランスアミナーゼの他の例であると考えられる。この酵素は、L−アラニンおよび4−メチルチオ−2−オキソブタノエートへのD−メチオニンおよびピルビン酸の可逆的な変換を触媒する。オキサロ酢酸、フェニルピルビン酸、2−オキソ酪酸、2−オキソ吉草酸、2−オキソヘプタン酸、グリオキシレート、およびオキソグルタル酸もまたアミノ受容体として役立ち得る。
D−メチオニンアミノトランスフェラーゼは、標準的なクロマトグラフィープロトコールおよびPharmacia社製AKTA Explorerシステムを用いて、カリフラワー小花および発芽ピーナッツ胚から部分的に精製する。相同タンパク質のタンパク質配列は、LC/MS/MSフィンガープリント技術およびHarvard Microchemistry機関によって行われるデータベース検索により決定する。植物遺伝子のコード領域は、実施例10(1)に記載のように、標準的なPCRプロトコールを用いることによってまたは遺伝子の合成によってcDNAライブラリーからクローニングする。
カリフラワーからの単離
400グラムの新たに摘んだカリフラワー小花は、浸漬およびブレンダーを用いる混合を交互にすることによって、400mLの4℃スクロース/緩衝液(0.4Mスクロースおよび0.1Mリン酸ナトリウム緩衝液pH7.4)を用いて抽出する。細胞片は、チーズクロスを用いて濾過により取り出し、結果として生じた溶液は4℃で30分間40,000×gで遠心分離する。固体材料(ミトコンドリア細胞器官を含有する)は、20mL 10mMリン酸ナトリウム緩衝液pH7.4中に再懸濁させ、酵素は、200mL冷(−30℃)アセトンを用いて抽出する。懸濁液は再遠心分離し、沈澱物は、Savant社製Speed Vacを用いて乾燥させる。固体材料は、10mMリン酸ナトリウム緩衝液pH7.4中に溶かし、残りのアセトンは、PD−10カラムを用いて除去する。
発芽ピーナッツ胚ホモジネート(子葉なし)からのD−メチオニンアミノトランスフェラーゼ酵素は、Durham,J.I.ら,Phytochemistry 12:2123−2126,(1973)の方法に従って精製する。還元剤は、酵素を安定化するために粗抽出物の調製の間に用い、細胞片は、33,000×gでの遠心分離によって取り出す。35〜50%硫安画分は、低温でのインキュベーションおよび沈澱物中のタンパク質の除去によってさらに精製する。上清は、アセトンを用いてさらに分画する。次いで、活性プールは、ゲル濾過クロマトグラフィー(Sephadex200、G.E. Healthcare社、ピスカタウェイ、NJ)によってさらに精製する。
図8は、L−アミノ酸アミノトランスフェラーゼ(L−芳香族アミノトランスフェラーゼ、L−アラニン−アミノトランスフェラーゼおよび/またはL−トリプトファン−アミノトランスフェラーゼのごとき)、R特異的アルドラーゼ、アラニンラセマーゼ、ならびにD−アラニンアミノトランスフェラーゼを用いて、L−トリプトファンから立体異性的に豊富なR,Rモナチンを生成するための生合成経路を示す。
生成物、R,R−モナチンは以下の反応混合物から精製した。0.33リットル中、50mM炭酸水素アンモニウム、pH8.2、4mM MgCl2、0.05mMピリドキサールリン酸(「PLP」)、200mMピルビン酸ナトリウム、および50mM D−トリプトファンを、トリプトファンが溶けるまで500mLガラスボトル中で室温で混合した。液体に数分間窒素を流し、次いで3.0mg/mL Biocatalytics社製(パサデナ、CA)広範囲D−トランスアミナーゼ(カタログ#AT−103)および配列番号22の0.1mg/mL精製アルドラーゼを追加した。反応混合物は室温で緩やかに撹拌した。アルドラーゼは、実施例3Aに記載のように精製した。50mM D−トリプトファンの付加的なアリコートを、混合物を最初に調製した15時間および22時間後に固体として追加した。ヘッドスペースに各追加の後に窒素を流した。追加したトリプトファンのすべてが溶けたわけではなかったが、濃度は約50mMに維持した。40時間後、残りの固体トリプトファンを濾過した。ポストカラム蛍光検出液体クロマトグラフィーによる反応混合物の分析(実施例1を参照されたい)は、溶液中のトリプトファンの濃度が49mMであり、モナチンの濃度が3.9mMであったことを示した。
B.スファエリクスHIS6−D−アラニンアミノトランスフェラーゼの精製のための標準的手順
BL21(DE3)::B.スファエリクス dat pET30aの新鮮な培養平板(50μg/mLカナマイシンを有するLB寒天)から開始して、細胞は、3〜5時間、225rpmで振盪させながら37℃で、50μg/mlカナマイシンを有する5mLのLuria−Bertaniブロス(「LB」)中で成長した。続いて、培養物は、Novagen社製Overnight Express System II溶液1〜6(EMD Bioscience社、マディソン、WI)に加えて50μg/mLカナマイシンを含有するフラスコ中に0.25%(容量/容量)で移した。細胞は、一晩(16〜18時間)、37℃および225rpmで成長させた。OD600が約8.0となると、細胞は、10分間10,000rpmでJS−16.25回転子を有するBeckman社製(フラートン、CA)J25II遠心分離機中で遠心分離によって採取した。細胞ペレットは、低温の50mM EPPS緩衝液(pH8.2)を用いて洗浄し、細胞をさらに遠心分離した。洗浄した細胞ペレットは、採取し、直ちに用いたまたは精製に必要とされるまで−80℃で凍結させた。
無細胞抽出物は、上記に記載のように調製した。His6−BsphDATタンパク質は、以下の変化を加えて同様に精製した:細胞破壊およびタンパク質精製に用いた緩衝液はすべて、50μM PLPを有する100mMリン酸カリウム、pH7.8を含有した。タンパク質は、GE Healthcare社製Chelating Sepharose(商標)Fast Flow樹脂(ニッケル(II)形態)のみを用いて精製した。
トリプトファンおよびピルビン酸からのインドール−3−ピルビン酸およびアラニンの形成は、双方の精製手順によって調製した酵素を用いてアッセイした。反応混合物は、100mMリン酸カリウム、pH7.8、0.05mMピリドキサールリン酸(PLP)、100mMピルビン酸ナトリウム、40mM D−トリプトファン、および0.03〜0.1mg/mL精製酵素を含有した。トリプトファンは固体として追加した。酵素以外の成分はすべて一緒に混合し、トリプトファンが溶けるまで30℃でインキュベートした。次いで、酵素を追加し、反応溶液は室温でインキュベートした。所定の時点で、反応物をサンプリングし、試料は、直ちに氷上に保存し、実施例1に記載されるポストカラム蛍光検出液体クロマトグラフィー法によるアラニン分析のために希釈した。毎分1mgの酵素当たりに形成されたアラニンの濃度として、下記の表59に酵素調製物の特異的活性を挙げる。
数種のバチルスD−アミノ酸アミノトランスフェラーゼ(EC2.6.1.21、D−アラニンアミノトランスフェラーゼまたはD−アスパラギン酸アミノトランスフェラーゼとしても知られている)は、実施例18に記載のように、R,Rモナチンの生成のための共役アッセイにおいて用いるために組換えで生成した。これらの酵素は、モナチンの生成について先に記載されるD−アミノトランスフェラーゼ(米国特許出願公開第20040063175号および米国特許出願公開第2005282260号)に相同である。新規なD−アミノ酸アミノトランスフェラーゼ(「DAAT」)を含有する候補とすることができる株の選択に用いられるアプローチは、ATCCに委託されたB.スファエリクス株のリストを評価し、異なる種名で先に委託されたいくつかについて分析するためのものであった。以下の生物をATCCにオーダーした:ATCC4978−−バチルス・ロタンス(Bacillus rotans)として初め委託されたバチルス・スファエリクスおよびATCC7063−−バチルス・セロシティディス(Bacillus serositidis)として初め委託されたバチルス・スファエリクスおよびATCC21538−バチルス・サーキュランスとして初め委託されたバチルス・スファエリクス。バチルス・スファエリクス、バチルス・ハロデュランス、ゲオバチルス・ステアロテルモフィルス、バチルス・セレウス、バチルス・スブチリス、およびバチルス・リケニホルミスからの知られているDAATタンパク質配列は、種々のDAATタンパク質中の保存された配列領域を得るために整列させた。プライマーは、タンパク質配列保存の領域において設計し、上記に言及したATCC株からのDAAT遺伝子配列のポリメラーゼ連鎖反応(「PCR」)増幅に用いた。
プライマーは、DAATのアラインメントにおける保存領域に基づいて上記に言及されるように設計した。オリゴヌクレオチドプライマー配列を下記に示す。5'-GAAGACCGTGGTTATCAATTT-3'(配列番号65)(順行プライマー)、5'-GATGGTATTTACGAAGTAATC-3'(配列番号66)(順行プライマー)、5'-AGATTTAATATCACAACGTAAC-3'(配列番号67)(逆行プライマー)、5'-GCCAAGTAAAATTTAAGATTTA-3'(配列番号68)(逆行プライマー)、5'-ATTTGCTGGGTGCGTATAAAG-3'(配列番号69)(逆行プライマー)。知られているDAATとのプライマー組合せのアラインメントに基づいたPCR断片の期待サイズ:配列番号65および配列番号67−およそ380bp;配列番号65および配列番号68−およそ395bp;配列番号65および配列番号69−およそ534bp;配列番号66および配列番号67−およそ336bp;配列番号66および配列番号68−およそ346bp;配列番号66および配列番号69−およそ510bp。
4978 DAT GSP1 Upstrm 5'-GACATGCTCCTCCGCTGTAAATAATTCACC-3'(配列番号70);
4978 DAT GSP1 Dwnstr 5'-CCCTGGTGATGAAGTGAAGCCAGTATTAAC-3'(配列番号71);
4978 DAT GSP2 Upstrm 5'-ATCGCCAAATTGATAACCACGGTCTTC-3'(配列番号72);
4978 DAT GSP2 Dwnstr 5'-ACGTCCCGTAGCAAACTTTGAAAAAGGTGT-3'(配列番号73);
7063 DAT GSP1 Upstrm 5'-TGCATAGAATCGGTCGATATGTTCAGTAGC-3'(配列番号74);
7063 DAT GSP1 Dwnstr 5'-GCGGAGAAACGATTACAGAAGGTTCTTCAA-3'(配列番号75);
7063 DAT GSP2 Upstrm 5'-GTCACCAAATTGATAACCACGGTCTTC-3'(配列番号76);および
7063 DAT GSP2 Dwnstr 5'-GGTGTACTTTATACGCACCCAGCAAAT-3'(配列番号77)。
AP1 5'-GTAATACGACTCACTATAGGGC-3'(配列番号78)および
AP2 5'-ACTATAGGGCACGCGTGGT-3'(配列番号79)。
ATCC4978DAATNde1F 5'-GGCCTTGGCATATGAGTTATAGCTTATGGAATGACC-3'(配列番号80);
ATCC4978DAATBamH1R 5'-GGCCTTAAGGATCCTTATGCGCGAATACCTTTTGGG-3'(配列番号81);
ATCC7063DAATNde1F 5'-GGCCTTGGCATATGAGCTACACTTTATGGAATGA-3'(配列番号82);および
ATCC7063DAATBamH1R2a 5'-GGCCAAGGATCCGCTACCCACTAATCATTAGA-3'(配列番号83)。
ATCC4987およびATCC7063からの新規なDAATは、実施例15に記載のように、(pETベクター中)、イー・コリ発現宿主BL21(DE3)(Novagen社、マディソン、WI)中に形質転換した。培養物は、上記に記載されるプロトコールを用いて成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、プラスミドの同一性を確認するために上記に記載のように制限消化によって分析した。
ATCC4978からのアミノトランスフェラーゼもまた、D−トリプトファンからモナチンを生成するための能力について試験した(実施例3Aでのように)。以下のものを反応混合物1mL当たりに追加した:約50μgアルドラーゼ(C.テストステローニProAアルドラーゼまたは配列番号22のアルドラーゼ、精製済み)、4mM MgCl2、50mM D−トリプトファン(固体として供給)、1.0mg D−アミノトランスフェラーゼ、100mMピルビン酸ナトリウム、100mMリン酸カリウム緩衝液pH7.5、および0.05mM PLP。実験は、アミノトランスフェラーゼを追加しないネガティブコントロールと共に二通り実行した。試料は、緩やかに振盪させながら30℃で種々の長さの期間インキュベートした。これらの方法を用いて、モナチンが生成された場合に検出された唯一の立体異性体はR,RおよびS,Rである。全モナチンおよびR,Rモナチンパーセントは、実施例1に記載のように検出された、またそれを下記の表62〜64に挙げる。表62〜64のそれぞれに示される結果は、二通りの反応からの平均値である。
数種のバチルスD−アミノ酸アミノトランスフェラーゼ(EC2.6.1.21、D−アラニンアミノトランスフェラーゼまたはD−アスパラギン酸アミノトランスフェラーゼとしても知られている)は、R,Rモナチンの生成のための共役アッセイにおいて用いるために組換えで生成した。この酵素は、モナチンの生成について先に記載されるD−アミノトランスフェラーゼ(米国特許出願公開第20040063175号および米国特許出願公開第2005282260号)に相同である。
B.スファエリクス(ATCC番号10208)およびB.リケニホルミス(ATCC番号10716)は、一晩30℃で栄養寒天上で成長させた。コロニーのグループを100μLの滅菌水中に置き、細胞を破壊するために95℃で5分間加熱した。3μLを、続くポリメラーゼ連鎖反応(PCR)による増幅において用いた。ゲノムDNAをB.ハロデュランス(ATCC番号BAA−125D)についてオーダーし、100ng/μLの濃度まで水中で再懸濁させた。バチルス・セレウスのゲノムDNA(ATCC番号1−987Dおよび14579D)は、同様にクローニングするためにオーダーした。
プライマーは、NcoI部位およびBamHI部位を用いて、pET28bベクターおよびpET30aベクター(Novagen社、マディソン、WI)にクローニングするためにB.スファエリクスdat遺伝子について設計した。pET30構築物はN末端HisタグおよびSタグを含有するが、pET28構築物はタグ付きではない。
N末端:5'-GATATACCATGGCATACTCATTATGGAATG-3'(配列番号88)およびC末端:5'-GTTATCGGATCCTTAGGCATTAATTGAAATTG-3'(配列番号89)。
B.リケニホルミスプライマーおよびB.ハロデュランスのプライマーは、NcoI部位およびBamHI部位を用いて、pET28bベクターおよびpET30aベクターにクローニングするために設計した。pET30構築物はこの場合タグなしであったが、pET28構築物は小さなN末端his−タグを含有した。
B.リケニホルミス datプライマー:
N末端5'-GGCCGGTTCATATGAAAGTTCTTTTTAACGGC-3'(配列番号90)およびC末端5'-CCTTCCGGATCCTTAAACCGTTTTGGCTGTCT-3'(配列番号91)。
B.ハロデュランスプライマー:
N末端5'-GATATACATATGGATTATTGCCTTTACCAA-3'(配列番号92)およびC末端5'-GAATCCGGATCCTCACTGCTTCATCGCTGTTTG-3'(配列番号93)。
B.セレウスプライマー:
N末端5'-TAAGAGGAATAACATATGGCATACGAAAGATTT-3'(配列番号94)およびC−末端5'-GAATTCGGATCCTTAAGAAGATGACATATTGG-3'(短鎖PCR産物)(配列番号95)。
N末端5-TAAGAGGAATAACATATGGGATCGAAATTGGCA-3'(長鎖PCR産物)(配列番号96)。
N−末端5'-GGCCTTGGCATATGGGATACACTTTATGGAATGACC-3'(配列番号97)およびC−末端5'-TTGGAACCGGATCCTTATATATGAAGCGGTTTTGG-3'(配列番号98)。
B.スファエリクスDATについてのPCR産物は、Qiagen社製QIAquick(登録商標)PCR精製キット(Qiagen社、バレンシア、CA)を用いて精製し、BamHI緩衝液(New England Biolabs社、イプスウィッチ、MA)中でBamHIおよびNcoIで消化した。消化したベクター(pET28およびpET30)ならびに挿入断片は、Qiagen QIAquick(登録商標)Gel Extraction Kit(Qiagen社、バレンシア、CA)を用いて精製した。ライゲーションは、Roche社製Rapid DNA Ligation Kit(Roche社、インディアナポリス、IN)を用いて行い、QIAquick(登録商標)PCR精製キットを用いて精製した。ライゲーションしたものは、Bio−Rad社エレクトロポレーションマニュアルに記載のように、0.2cmキュベットおよびBio−Rad社製Gene Pulser(登録商標)IIシステムを用いてエシェリキア・コリDH10Bに形質転換した。細胞は、225rpmで振盪させながら、37℃で30分間、900μL SOC培地中で回復させた。細胞は、カナマイシン(25μg/mL)を含有するLB寒天平板上で平板培養した。プラスミドDNAは、Qiagen社製スピンミニプレップキット(Qiagen社、バレンシア、CA)を用いて精製し、BamHIおよびNcoIを用いた制限消化によって正確な挿入断片についてスクリーニングした。正確な挿入断片を有するようであるプラスミドの配列は、Agencourt BioScience社(ビバリー、MA)でジデオキシ連鎖停止DNA配列決定によって検証した。配列決定により、NCBI受入番号AAC33964(gi:3513755)に挙げられるアミノ酸配列を有するタンパク質を生成する、受入番号AF081278領域:134..985(gi:3513754)に見られるコード配列を検証した。
プラスミドDNAは、pETベクター中の構築物のために、イー・コリ発現宿主BL21(DE3)(Novagen社、マディソン、WI)中にサブクローニングした。培養物を成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、同一性を確認するために制限消化によって分析した。誘発は、通常、カナマイシン(50μg/mL)を含有するLB培地中で行った。細胞は、37℃で0.4〜0.8のOD600まで成長させ、0.1mM IPTG(イソプロピルチオガラクトシド)を用いて誘発し、誘発の3〜4時間後にサンプリングした。細胞抽出物は、Novagen社製BugBuster(商標)試薬付属のプロトコールに従って調製した。(ベンゾナーゼヌクレアーゼおよびRoche社製完全プロテアーゼインヒビターカクテルを追加)。高いレベルの可溶性タンパク質が、双方のB.ハロデュランス遺伝子産物、双方のB.スファエリクス遺伝子産物、双方のG.ステアロテルモフィルス遺伝子産物、およびタグなしB.リケニホルミス遺伝子産物について、SDS−PAGEで判断されるように、予測した分子量で得られた。精製タンパク質を用いた反応については、Hisタグ遺伝子産物は、メーカーのプロトコール(Novagen社、マディソン、WI)に従ってHis−Bindカートリッジを用いて精製した。溶出液画分は、PD−10(Amersham Biosciences社、ピスカタウェイ、NJ)カラム上で脱塩し、25〜100mMリン酸カリウム緩衝液、pH7.5中に溶出させた。全タンパク質アッセイは、Pierce社製BCAキット(ロックフォード、IL)を用いて行い、発現パーセントはSDS−PAGEから評価した。あるいは、細胞抽出物中の可溶性タンパク質は、BioRad Laboratories社製Experion Automated Electrophoresis Stationで分離し、Experion Softwareバージョン1.1.98.0を用いて濃度および発現パーセントについて分析した。B.セレウス遺伝子を含有するpBAD−TOPO構築物はInvitrogen社によって推奨されるように発現させたが、DAATの発現のレベルは、SDS−PAGE分析の間に他のタンパク質と組換えタンパク質を識別できないものであった。
数種のバチルスD−アミノ酸アミノトランスフェラーゼは実施例18および15に記載された。G.ステアロテルモフィルス酵素は、モナチンに対する低いアミノ基転移活性を有し、D−トリプトファンからより少ない全モナチンが生成されたが、この酵素はなお注目する構造要素を有し、またこの酵素は熱安定酵素である。従って、ハイブリッドタンパク質をより高い活性の酵素(B.スファエリクス)およびゲオバチルス酵素の間で作り出した。
設計した標的タンパク質配列は配列番号99とする。配列番号100、配列番号99に対応するコード配列はイー・コリコドン使用頻度に基づいて設計した。
N−末端5'-GGCCTTGGCATATGGGATACACTTTATGGAATGACCA-3'(配列番号101)およびC−末端5'-TTGGAACCGGATCCTTAGCTGTTAAGGCTCAGTGGAA-3'(配列番号102)
PCR生成物は、Qiagen社製QIAquick(登録商標)Gel Extraction Kit(Qiagen社、バレンシア、CA)を用いてゲル精製し、メーカーのプロトコール(Invitrogen社)に従って、Zero Blunt TOPO(登録商標)クローニングキットを用いてクローニングした。プラスミドは、PCRによって最初のスクリーニングのためにTOP10化学的コンピテント細胞中に形質転換した。プラスミドDNAは制限消化によってスクリーニングし、DNA配列を検証した。
プラスミドDNAは、メーカーのプロトコールに従って、イー・コリ発現宿主BL21(DE3)(Novagen社、マディソン、WI)中に形質転換した。培養物を成長させ、プラスミドは、Qiagen社製ミニプレップキット(Qiagen社、バレンシア、CA)を用いて単離し、同一性を確認するためにPCRによって分析した。誘発は、カナマイシン(50μg/mL)を含有するLB培地中で行った。細胞は、0.1mM IPTG(イソプロピルチオガラクトシド)を用いて誘発して、37℃で0.5のOD600まで成長させ、誘発の3時間後にサンプリングした。細胞抽出物は、Novagen社製BugBuster(商標)試薬付属のプロトコールに従って調製した。(ベンゾナーゼヌクレアーゼおよびRoche社製完全プロテアーゼインヒビターカクテルを追加)。高いレベルの全タンパク質が、両遺伝子産物について、SDS−PAGEで判断されるように、予測した分子量で得られた。しかしながら、タンパク質の可溶性レベルはより低かった。遺伝子産物のタグなしバージョンはより発現し、細胞性抽出物としてアッセイした。細胞抽出物中の可溶性タンパク質は、BioRad Laboratories社製Experion Automated Electrophoresis Stationで分離し、Experion Softwareバージョン1.1.98.0を用いて濃度および発現パーセントについて分析し、比較アッセイに用いられるD−アミノトランスフェラーゼの量を標準化した。
D−アミノ酸デヒドロゲナーゼは、BioCatalytics社(パサデナ、CA)から購入したライブラリーの一部であった。
モナチンを形成するためのMPのアミノ化は、アミノトランスフェラーゼまたはNADHもしくはNADPHのごとき還元補因子を必要とするデヒドロゲナーゼによって触媒できる。これらの反応は可逆的であり、どちらかの方向で測定できる。デヒドロゲナーゼ酵素を用いる場合の方向性は、アンモニウム塩の濃度によって大部分はコントロールできる。
モナチンの酸化的脱アミノは、NAD+がより多くの発色団のNADHに変換されるにつれての、340nmでの吸光度の増加に従ってモニターした。
基質としてこのアッセイに用いられるR−MPは、アミノ受容体としてピルビン酸を用いて、リン酸カリウム緩衝液中のAT−103広範囲D−アミノトランスフェラーゼ(BioCatalytics社)を用い、R,Rモナチンのアミノ基転移によって生成した。S−MPは、アミノ受容体として2−オキソグルタル酸を用いて、リン酸カリウム緩衝液中のAT−102L−アミノトランスフェラーゼ(BioCatalytics社)を用い、S,Sモナチンのアミノ基転移によって生成した。両化合物は、調整用スケールのHPLCを用いて精製した。
配列番号22のアルドラーゼと共役する、BioCatalytics社製アミノ酸デヒドロゲナーゼ酵素を用いる、インドール−3−ピルビン酸およびピルビン酸からのモナチンの生成は、以下の条件下でアッセイした:1mg/mLデヒドロゲナーゼ酵素、10mM NADH、500μg/mLアルドラーゼ(精製済み)、50mMリン酸カリウム緩衝液pH7.5、4mM MgCl2、20mg/mL インドール−3−ピルビン酸、200mMギ酸アンモニウム、および200mMピルビン酸を20時間100rpmで30℃でインキュベートした。ネガティブコントロールはアミノ酸デヒドロゲナーゼ酵素を含有しなかった。実験は二通り行った。デヒドロゲナーゼのどれも、ネガティブコントロールと比較して、インドールピルビン酸およびピルビン酸から数量化できる量のモナチンを生成するようではなかった(実施例1で記載されるLC/MS/MSによって測定)。しかしながら、大量のアラニンおよびトリプトファンが生成された。デヒドロゲナーゼに対するアルドラーゼの比の増加はモナチン生成を改善すると考えられることが予想される。進化分子工学アプローチは、ピルビン酸およびインドール−3−ピルビン酸に対するMPに対しての還元的アミノ化活性の比を改善するために用いることができることもまた予想される。
バチルス・スファエリクスD−アラニンアミノトランスフェラーゼは、実施例14で記載のようにHIS6−タグ付きタンパク質として精製した。
シノリゾビウム・メリロティHMGアルドラーゼ(「proA」)は、HIS6−タグ付きB.スファエリクスD−アラニンアミノトランスフェラーゼについて実施例14で記載される手順に類似する手順を用いてHIS6−タグ付きタンパク質として精製した。
B.スファエリクスHIS6−タグ付きD−アラニンアミノトランスフェラーゼおよびR.メリロティHIS6−タグ付きproAアルドラーゼは、実施例21および22に記載のように、精製し、固定化した。
オペロンは、D−アミノトランスフェラーゼ(「DAT」)およびR特異的アルドラーゼを用いるイー・コリでのR,Rモナチンのin vivo生成を実証するために構築した。R,Rモナチンは、米国特許出願公開第2005/0282260A1号(図1および2ならびに実施例11)に記載される経路を用いてD−トリプトファンから作製した。簡潔に言えば、D−トリプトファンは、アルファ−ケト酸もまたD−アミノ酸に変換されるアミノ基転移反応によって in vivoでインドール−3−ピルビン酸に変換される。ピルビン酸は、R特異的アルドラーゼを用いて、アルドール縮合でin vivoでインドール−3−ピルビン酸と反応して、圧倒的にR−MPを生成する。終わりに、第1の反応からのR−MPおよびD−アミノ酸(または細胞中の任意のD−アミノ酸)はR,Rモナチンおよび対応するアルファ−ケト酸にin vivoで変換される。
プラスミドpCEC−Ndeは、pPRONdeプラスミド(ClontechからのpProLAR、米国特許出願公開第20040235123号に記載されるように修正)のp15A複製開始点をpPROTet.E133(Clontech Laboratories,Inc.、Mountain View、CA)からのCol E1複製開始点と交換することによって構築した。両プラスミドは、制限酵素Avr IIおよびAat II(New England Biolabs、Beverly、MA)を用いて処理し、適切な断片(Col E1開始点およびクロラムフェニコール抵抗性遺伝子を運ぶpPROTet.E133に由来する1731bp断片ならびにPlac/araプロモーター領域およびマルチクローニング部位を運ぶpPRONdeに由来する760bp)はアガロースゲル電気泳動によって精製し、その後、QIAquick(登録商標)Gel Extraction Kit(Qiagen、Valencia、CA)を用いる抽出および回収を続けた。精製DNA断片は、Quick Ligation Kit(New England Biolabs、Beverly、MA)を用いて共にライゲーションし、ライゲーション混合物は、化学的コンピテントイー・コリTOP10細胞(Invitrogen、Carlsbad、CA)に形質転換した。クローンは50μg/mLクロラムフェニコールを含有するLB寒天上で単離し、プラスミドの制限消化によって確認した。
プライマーは、R選択的アルドラーゼ活性を有する酵素をコードする配列番号172のDNA配列に基づいて設計した。配列番号173のアルドラーゼおよびそのアルドラーゼをコードする核酸配列を含有するプラスミド(特定のアルドラーゼの遺伝子配列は配列番号172として示される、はDiversa Corporation、San Diego、Californiaから得た。配列番号172は、アルドラーゼ遺伝子についてDiversa Corp.によってスクリーニングされたライブラリーの一部であった。しかしながら、配列番号172のアルドラーゼ遺伝子は、当業者に知られている任意の方法によって再構築されてもよい。例えば、配列番号172のアルドラーゼ遺伝子は、当業者に知られているアセンブリーPCR法を利用して再構築されてもよい。プライマーは、クローニングおよび発現のために、制限部位およびアルドラーゼ遺伝子の前にリボソーム結合部位を含有するように設計した。用いたオリゴヌクレオチドプライマー配列は次の通りであった。
5'-GGCCGGAACTGCAGAAGAAGGAGATATATAATGAAGCCGGTGGTGGTG-3'(配列番号174)および
アルドラーゼRxbaI
5'-GGCCAAGGTCTAGATTAGACATAGGTGAGCCC-3'(配列番号175)。
Oasis(登録商標)HLB 3cc(60mg)Extraction Cartridge(Waters Corp.、Milford、Massachusetts)を試料を濃縮するために用いた。発酵試料は、細胞物質を除去するために最初に遠心分離した。ギ酸を発酵試料に追加し、最終濃度を約1%とした。抽出カートリッジは、発酵試料の追加前に、少なくとも2mLメタノール、その後2mLの1%ギ酸を用いて調節した。各溶液は、抽出カートリッジを通して別々に流した。次いで、1%ギ酸を含有する無希釈の遠心分離した発酵試料の5mLまでを抽出カートリッジに追加した。発酵溶液は、カートリッジを通してゆっくり流した。カートリッジは、少なくとも2mLの1%ギ酸を用いてすすいだ。1mLのアセトニトリルまたはメタノールをカートリッジに追加し、カートリッジを通してゆっくり流した。この溶出液は、乾燥するまで、窒素の緩やかな流れの下に置いた。乾燥試料は、150〜200μLの水または移動相に再構成した。150μLを300μL HPLCプラスチックバイアル中に入れ、液体クロマトグラフに注入した。表73は、乾燥細胞重量当たりの全モナチンおよびR,R立体異性体またはS,S立体異性体である、合計のうちの割合を示す。
gaagccgg tggtggtgca gactatcgag cgggccgacc gagcgatcat cgagggtctg gccgcgtgtg gcgttgccac cgtccatgag gcgcaggggc gccgggggct gcttgcgtcc tacatgcgcc cgatctattc gggcgctgcg gttgcggcct cggccgtcac catcctctct ccaccctgcg acaactggat gctgcacgtc gccatcgagc agatccagcc gggcgacatt ctcgttctcg gcacgacctc tccgtccgat gccggctatt tcggtgatct gctggcgact tcggccaagg cgcgcggttg cgtcgggttg gtcatcgatg ccggcgtacg cgatatccgc gacctgacag cgatgcagtt tccggtctgg tccaaggccg tttcggccca gggcacgatc aaggagacgc tgggttcggt caacgtcccc gtcgtctgcg ccggtgctct ggtcaatccc ggcgacgtcg tcgtggccga tgacgacggt gtctgcgtgg tgcgccgcga ggaagccgcg gaaacgctgg aaaaggcccg ggcgcggatc gccaatgagg aggaaaagcg ccagcgcttt gccgctggcg aactcgggct cgacatctac aagatgcgcg aacgcctcgc tgccctgggg ctcacctatg tctga(配列番号172)。
Met Lys Pro Val Val Val Gln Thr Ile Glu Arg Ala Asp Arg Ala Ile Ile Glu Gly Leu Ala Ala Cys Gly Val Ala Thr Val His Glu Ala Gln Gly Arg Arg Gly Leu Leu Ala Ser Tyr Met Arg Pro Ile Tyr Ser Gly Ala Ala Val Ala Ala Ser Ala Val Thr Ile Leu Ser Pro Pro Cys Asp Asn Trp Met Leu His Val Ala Ile Glu Gln Ile Gln Pro Gly Asp Ile Leu Val Leu Gly Thr Thr Ser Pro Ser Asp Ala Gly Tyr Phe Gly Asp Leu Leu Ala Thr Ser Ala Lys Ala Arg Gly Cys Val Gly Leu Val Ile Asp Ala Gly Val Arg Asp Ile Arg Asp Leu Thr Ala Met Gln Phe Pro Val Trp Ser Lys Ala Val Ser Ala Gln Gly Thr Ile Lys Glu Thr Leu Gly Ser Val Asn Val Pro Val Val Cys Ala Gly Ala Leu Val Asn Pro Gly Asp Val Val Val Ala Asp Asp Asp Gly Val Cys Val Val Arg Arg Glu Glu Ala Ala Glu Thr Leu Glu Lys Ala Arg Ala Arg Ile Ala Asn Glu Glu Glu Lys Arg Gln Arg Phe Ala Ala Gly Glu Leu Gly Leu Asp Ile Tyr Lys Met Arg Glu Arg Leu Ala Ala Leu Gly Leu Thr Tyr Val(配列番号173)。
実施例18でクローニングしたバチルス・スファエリクス(ATCC株10208)D−アラニンアミノトランスフェラーゼをPLPを含有するリン酸カリウム緩衝液を用いて、改善された方法を用いて、実施例14に記載されるようにHIS6タグ付きタンパク質として精製した)。配列番号104のアルドラーゼ(実施例3Bでクローニング)は、実施例27に記載されるようにHIS6タグ付きタンパク質として精製した。
アミノ末端HIS6−精製タグを有するATCC株4978に由来するD−アミノトランスフェラーゼのT243N変異体(実施例6Bに記載される)は、振盪フラスコ中に50μg/mLカナマイシンを含有するEMD Biosciences Overnight Express System II(溶液1〜6)(La Jolla、CA)を用いて生成した。この発現系は、細胞成長をモニターする必要のないIPTG誘発性系の発現を誘発する。プラスミドpET28b上のATCC株4978に由来するT243N変異体D−アミノトランスフェラーゼについての遺伝子を運ぶイー・コリBL21(DE3)宿主細胞の液体培養物または平板培養物のいずれかからの培地の200mLアリコートの接種の後(1Lフラスコ中)、培養物は、225rpmで振盪させながら一晩30℃でインキュベートした。OD600が最小の6まで達すると、細胞は、遠心分離によって採取し、緩衝液を用いて1回洗浄した。
pET28bプラスミド上に配列番号104のアルドラーゼについての遺伝子(遺伝子は配列番号103として示される)を運ぶイー・コリBL21(DE3)pLysS宿主細胞のクローニングは実施例3Bに記載される。
Claims (69)
- L−トリプトファンからインドール−3−ピルビン酸(「I3P」)を生成し、
I3Pから少なくともRモナチン前駆体(「R−MP」)を含むモナチン前駆体(「MP」)を生成し、
次いでMPからモナチンを生成すること
を含む経路を介してモナチンまたはその塩を生成することを含むことを特徴とする方法であって、
モナチンは、少なくともR,Rモナチンを含み、
L−トリプトファンからのI3Pの生成は、モナチン前駆体、モナチン、またはその双方よりもL−トリプトファンに対するより高い活性、より高い特異性、またはその双方を有する1つまたは複数の第1の酵素によって促進され、該1つまたは複数の第1の酵素は、L−トリプトファンアミノトランスフェラーゼ、L−芳香族アミノトランスフェラーゼ、L−アスパラギン酸アミノトランスフェラーゼ、およびL−アミノ酸オキシダーゼならびにその組合せから選ばれ、
該R−MPからの該R,Rモナチンの生成は、D−アミノトランスフェラーゼおよびD−アミノ酸デヒドロゲナーゼまたはその組合せから選ばれる1つまたは複数の第3の酵素によって促進される該方法。 - 該1つまたは複数の第1の酵素が、シノリゾビウム・メリロティTatA、HEXAspCP9T/R122G、およびその組合せから選ばれることを特徴とする請求項1に記載の方法。
- 該1つまたは複数の第1の酵素が、モナチン前駆体、モナチン、またはその双方に対して制限された活性、制限された特異性、またはその双方を有し、該1つまたは複数の第2の酵素が、S−アルファ−ケト酸モナチンに対して制限された活性、制限された特異性、またはその双方を有していてもよいことを特徴とする請求項1に記載の方法。
- 該D−アミノ酸デヒドロゲナーゼが、D−AADH−101、D−AADH−102、D−AADH−103、D−AADH−105、D−AADH−106、D−AADH−107、D−AADH−108、およびその組合せから選ばれ、
該D−アミノトランスフェラーゼが、
a)T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異、F200M突然変異、F200Y突然変異、ならびにF200MおよびT243Nの突然変異を有する、配列番号86に対応するD−アミノトランスフェラーゼのうちの1つまたは複数から選ばれるD−アミノトランスフェラーゼの活性を有する酵素、
b)a)の酵素のいずれかに対して少なくとも90%の配列同一性パーセントを有する配列を含む酵素、
c)バチルス・ハロデュランスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼとバチルス・スファエリクスD−アミノトランスフェラーゼのハイブリッドであるハイブリッドD−アミノトランスフェラーゼ、バチルス・スファエリクスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼ、バチルス・リケニホルミスD−アミノトランスフェラーゼ、ATCC7063からのD−アミノトランスフェラーゼ、D−アミノトランスフェラーゼ活性を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、または
d)a〜cの組合せ
であることを特徴とする請求項1に記載の方法。 - 該バチルス・ハロデュランスD−アミノトランスフェラーゼが、受入番号NP−243677に挙げられるアミノ酸配列を含み;
該バチルス・リケニホルミス分岐鎖アミノトランスフェラーゼが、イー・コリF37Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリY96Fに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリY165Lに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL127Kに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリR98Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL108Rに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL110Hに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL127Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、およびイー・コリR41Kに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼから選ばれ;
該ハイブリッドD−アミノトランスフェラーゼが、配列番号99であり;
ATCC7063からの該D−アミノトランスフェラーゼが、配列番号87であることを特徴とする請求項4に記載の方法。 - 生成された該R,Rモナチンが、生成された全モナチンの少なくとも約60重量%であることを特徴とする請求項3に記載の方法。
- R−MPが、化学的アルドール縮合反応においてI3Pから生成されることを特徴とする請求項1に記載の方法。
- I3PからのR−MPの生成が、R−MPをS−モナチン前駆体(「S−MP」)に対して約1:1よりも大きな比で生成する1つまたは複数の第2の酵素によって促進されることを特徴とする請求項1に記載の方法。
- 該1つまたは複数の第2の酵素が、a)シノリゾビウム・メリロティHMGアルドラーゼからのアルドラーゼ、b)a)の相同体、c)a)に対して少なくとも90%の配列同一性パーセントを有する配列を含む酵素、d)配列番号22または配列番号104に示される配列を有するアルドラーゼ、およびe)その組合せから選ばれる、R特異的アルドラーゼ活性を有する酵素であることを特徴とする請求項8に記載の方法。
- 該1つまたは複数の第3の酵素が、S−アルファ−ケト酸モナチンまたはインドール−3−ピルビン酸についてよりもR−モナチン前駆体に対するより高い活性、より高い特異性、またはその双方を有し、該1つまたは複数の第2の酵素が、S−アルファ−ケト酸モナチンまたはインドール−3−ピルビン酸に対する制限された活性、制限された特異性、またはその双方を有していてもよいことを特徴とする請求項1に記載の方法。
- D−アミノトランスフェラーゼおよびD−アミノ酸デヒドロゲナーゼから選ばれる1つまたは複数の第1の酵素を用いてMP組成物からR,Rモナチンを生成することを含み、少なくともR−MPを含む該MP組成物および該1つまたは複数の酵素が、S−MPよりもR−MPからモナチンへの変換を選択的に促進することを特徴とする方法。
- 1つまたは複数の酵素が、バチルス・ハロデュランスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼおよびバチルス・スファエリクスD−アミノトランスフェラーゼのハイブリッド、ならびにその組合せから選ばれることを特徴とする請求項11に記載の方法。
- L−トリプトファンからインドール−3−ピルビン酸(「I3P」)を生成し、
I3PからMP組成物を生成すること
をさらに含み、
LトリプトファンからのI3Pの生成が、L−トリプトファンアミノトランスフェラーゼ、L−芳香族アミノトランスフェラーゼ、L−アスパラギン酸アミノトランスフェラーゼ、L−アミノ酸オキシダーゼ、およびその組合せから選ばれる1つまたは複数の第2の酵素によって促進され、
I3PからのMP組成物の生成が、1つまたは複数の非立体特異性アルドラーゼによって促進されることを特徴とする請求項12に記載の方法。 - モナチンを生成するための経路におけるある反応の生成物を、モナチンを生成するための経路における他の反応の反応物として再生するための方法であって、
L−トリプトファンが反応してI3Pを生成する場合に、アルファ−ケト基質からL−アミノ酸を生成し、
アミノ酸ラセマーゼを用いてL−アミノ酸をD−アミノ酸に変換し、
I3PからMPを生成し、
MPを反応させてモナチンを形成し、次いで
MPが反応してモナチンを形成する場合に、D−アミノ酸からアルファ−ケト基質を再生成することを含み、
アルファ−ケト基質は、アルファ−ケトグルタル酸、オキサロ酢酸、ピルビン酸、およびその組合せから選ばれ、
アミノ酸ラセマーゼは、アルファ−ケト基質がピルビン酸である場合、アラニンラセマーゼであり、アルファ−ケト基質がアルファ−ケトグルタル酸である場合、グルタミン酸ラセマーゼであり、アルファ−ケト基質がオキサロ酢酸である場合、アスパラギン酸ラセマーゼであることを特徴とする該方法。 - 該アラニンラセマーゼが、バチルス・ステアロサーモフィルスからのアラニンラセマーゼおよびゲオバチルス・ステアロテルモフィルスからのアラニンラセマーゼの熱安定性相同体から選ばれ、該グルタミン酸ラセマーゼが、L.ブレビスグルタミン酸ラセマーゼおよびP.ペントサセウスグルタミン酸ラセマーゼから選ばれ、該アスパラギン酸ラセマーゼはBioCatalytics社のASPR−101であることを特徴とする請求項14に記載の方法。
- I3PからのMPの生成が、R−MPをS−MPに対して約1:1よりも大きな比で優先的に生成する1つまたは複数のアルドラーゼ酵素によって促進されることを特徴とする請求項14に記載の方法。
- 1つまたは複数のアルドラーゼ酵素が、R特異的アルドラーゼ酵素であることを特徴とする請求項16に記載の方法。
- L−アミノ酸および立体反転アミノトランスフェラーゼの存在下でR−MPを反応させてR,Rモナチンを生成することを含み、L−アミノ酸は、L−アスパラギン酸、L−アラニン、およびL−グルタミン酸から選ばれることを特徴とする方法。
- トリプトファンからI3Pを生成し、I3PからR−MPを含むMPを生成することをさらに含むことを特徴とする請求項18に記載の方法。
- アミノトランスフェラーゼ酵素がトリプトファンからI3Pを生成することを促進し、アルドラーゼ酵素がI3PからR−MPを生成することを促進することを特徴とする請求項19に記載の方法。
- アルドラーゼ酵素が、R−MPをS−MPに対して1:1よりも大きな比で優先的に生成することを特徴とする請求項19に記載の方法。
- アルドラーゼ酵素がR特異的アルドラーゼ酵素であることを特徴とする請求項19に記載の方法。
- 平衡反応に対してモナチンの収量を増加させるための方法であって、
アルファ−ケト基質およびL−アミノトランスフェラーゼ酵素の存在下でトリプトファンを反応させてI3Pを生成し、アルファ−ケト基質が反応してL−アミノ酸を形成し、
I3Pを反応させてMPを生成し、
MPを反応させてモナチンを生成し、次いで
不可逆的反応でL−アミノ酸を反応させることによって、平衡経路からL−アミノ酸生成物を除去すること
を含むことを特徴とする該方法。 - アルファ−ケト基質がオキサロ酢酸であり、L−アミノ酸生成物がL−アスパラギン酸であり、不可逆的反応が、アスパラギン酸4−デカルボキシラーゼの存在下でL−アスパラギン酸を反応させてL−アラニンおよび二酸化炭素を生成するものであることを特徴とする請求項23に記載の方法。
- L−アミノ酸生成物を、R−MP反応のためのD−アミノ酸基質に変換することをさらに含むことを特徴とする請求項23に記載の方法。
- L−アミノ酸生成物がL−アラニンであり、L−アラニンが、アラニンラセマーゼ活性を有する酵素を用いてD−アラニンに変換されることを特徴とする請求項25に記載の方法。
- アルファ−ケト基質が、オキサロ酢酸およびアルファ−ケトグルタル酸ならびにその組合せから選ばれ、
アルファ−ケト基質がオキサロ酢酸である場合、L−アミノ酸生成物はL−アスパラギン酸であり、不可逆的反応が、アスパラギン酸デカルボキシラーゼの存在下でL−アスパラギン酸を反応させてベータアラニンおよび二酸化炭素を生成するものであり、
アルファ−ケト基質がアルファ−ケトグルタル酸である場合、L−アミノ酸生成物はL−グルタミン酸であり、不可逆的反応が、グルタミン酸デカルボキシラーゼの存在下でL−グルタミン酸を反応させて4−アミノブタン酸および二酸化炭素を生成するものであることを特徴とする請求項23に記載の方法。 - 経路におけるある工程で生成されたL−アミノ酸生成物を経路における他の工程で有用なD−アミノ酸基質に変換することを含む経路でモナチンを生成することを含み、経路は、
L−アミノトランスフェラーゼを用いてL−トリプトファンからI3Pを生成し、
アルファ−ケトグルタル酸の反応によってL−アミノ酸生成物としてL−グルタミン酸を生成し、該アルファ−ケトグルタル酸反応はL−トリプトファン反応と共役し、
L−アラニンアミノトランスフェラーゼを用いて、L−グルタミン酸とピルビン酸を反応させることによってL−アラニンを生成し、
I3PからR−MPを生成し、
アラニンラセマーゼを用いてL−アラニンからD−アミノ酸基質としてD−アラニンを生成し、
MPと共に基質としてD−アラニンを用いて、モナチンおよびピルビン酸を形成することを含むことを特徴とする方法。 - アルドラーゼ酵素がI3PからR−MPを生成することを促進することを特徴とする請求項28に記載の方法。
- アルドラーゼ酵素がR特異的アルドラーゼ酵素であり、R,Rモナチンが生成されることを特徴とする請求項29に記載の方法。
- アルドラーゼ酵素がR−MPをS−MPに対して約1:1よりも大きな比で生成し、R,Rモナチンが生成されることを特徴とする請求項29に記載の方法。
- L−トリプトファンからD−トリプトファンを生成し、
D−トリプトファンからI3Pを生成し、
I3PからR−MPを生成し、次いで
R−MPからモナチンを生成することを含む経路を介してR,Rモナチンまたはその塩を生成することを含むことを特徴とする方法であって、
L−トリプトファンからのD−トリプトファンの生成は、トリプトファンラセマーゼ、トリプトファンラセマーゼの活性を有するラセマーゼ、広域特異性アミノ酸ラセマーゼ、広域特異性アミノ酸ラセマーゼの活性を有するラセマーゼ、およびその組合せから選ばれる1つまたは複数の酵素によって促進され、
トリプトファンラセマーゼの活性を有するラセマーゼは、配列番号41に対応し、Y354C突然変異、Y354T突然変異、Y354G突然変異、Y354L突然変異、Y354N突然変異、Y354S突然変異、Y354I突然変異、Y354M突然変異、Y354P突然変異、ならびにY354A、M35V、およびR195Hの突然変異の組合せから選ばれる突然変異を有するアラニンラセマーゼ、配列番号43のアラニンラセマーゼ、配列番号41に対応し、M35C突然変異、F66E突然変異、Y354A突然変異、およびP197L突然変異ならびにその組合せから選ばれる突然変異を有するアラニンラセマーゼから選ばれ、
広域特異性アミノ酸ラセマーゼは、
a)配列番号120の広域特異性アミノ酸ラセマーゼ、Y396C突然変異を有する、配列番号120に対応する広域特異性アミノ酸ラセマーゼ、配列番号128の広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、配列番号128に対応する広域特異性アミノ酸ラセマーゼ、ビブリオ・フィシェリからのアラニンラセマーゼ、Genbank受入番号AAW85230.1のアラニンラセマーゼ、Genbank受入番号YP_204118のアラニンラセマーゼ、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼに対応する広域特異性アミノ酸ラセマーゼ、シュードモナス・プチダ(シュードモナス・ストリアタとしても知られている)から単離された広域特異性アミノ酸ラセマーゼ、配列番号116の配列を含む広域特異性アミノ酸ラセマーゼ、Genbank受入番号ZP_00898332.1 GI:82735470の広域特異性アミノ酸ラセマーゼ、ATCC4683からの広域特異性アミノ酸ラセマーゼ、配列番号204の広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、配列番号204に対応する広域特異性アミノ酸ラセマーゼ、P.プチダKT2440から単離された広域特異性アミノ酸ラセマーゼ、P.プチダNBRC12996から単離された広域特異性アミノ酸ラセマーゼ、ATCC7966からの広域特異性アミノ酸ラセマーゼ、シュードモナス株2150から単離された広域特異性アミノ酸ラセマーゼ、P.オレオボランスから単離された広域特異性アミノ酸ラセマーゼ、P.オーレオファシエンスから単離された広域特異性アミノ酸ラセマーゼ、P.プチダ12633から単離された広域特異性アミノ酸ラセマーゼ、P.フルオレッセンスから単離された広域特異性アミノ酸ラセマーゼ、P.プチダSCRC744から単離された広域特異性アミノ酸ラセマーゼ、P.グラベオレンスから単離された広域特異性アミノ酸ラセマーゼ、P.ストリアタAKU083から単離された広域特異性アミノ酸ラセマーゼ、配列番号204のアミノ酸24〜409を含有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号204のアミノ酸24〜409を含有する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ジャンデイから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC49572からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号194の広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ソブリアから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC35994からの広域特異性ラセマーゼの活性を有する酵素、配列番号192の広域特異性アミノ酸ラセマーゼの活性を有する酵素、L383M突然変異を有する、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼに対応する酵素、L383M突然変異を有する、ATCC14486からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号195を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号196を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号201を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号202を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、D76N突然変異を有する、エロモナス・キャビエからのアミノ酸ラセマーゼに対応する酵素、D76N突然変異を有する、配列番号179に対応する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ハイドロフィラから単離された広域特異性アミノ酸ラセマーゼ、配列番号177の広域特異性アミノ酸ラセマーゼ、配列番号177のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号146を含む広域特異性アミノ酸ラセマーゼ、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼ、配列番号179の広域特異性アミノ酸ラセマーゼ、配列番号179のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号155の部分配列を含有する広域特異性アミノ酸ラセマーゼ、配列番号151の広域特異性アミノ酸ラセマーゼ、およびその活性断片、
b)a)の広域特異性アミノ酸ラセマーゼのいずれかに対して少なくとも90%の配列同一性パーセントを有する配列およびその活性断片を含む酵素、
c)Genbank受入番号AB096176によってコードされた広域特異性アミノ酸ラセマーゼ、ラセマーゼがI384M突然変異を含むような置換を有する、Genbank受入番号AB096176に対応する核酸配列によってコードされた広域特異性アミノ酸ラセマーゼ、配列番号119によってコードされた広域特異性アミノ酸ラセマーゼ、配列番号127によってコードされた広域特異性アミノ酸ラセマーゼ、CP 000020.1 GI:59478708の領域800842..802053によってコードされた広域特異性アミノ酸ラセマーゼ、NC_006840によってコードされた広域特異性アミノ酸ラセマーゼ、KT2440 Bar DNAによってコードされた広域特異性アミノ酸ラセマーゼ、ラセマーゼがY396C突然変異を含むような置換を有する、配列番号119に対応する核酸配列によってコードされた広域特異性アミノ酸ラセマーゼ、ラセマーゼがI384M突然変異を含むような置換を有する、配列番号127に対応する核酸配列によってコードされた広域特異性アミノ酸ラセマーゼ、配列番号151を含む広域特異性アミノ酸ラセマーゼ、配列番号154を含む配列によってコードされた広域特異性アミノ酸ラセマーゼ、NZ_AALM01000002ヌクレオチド53173..54402によってコードされた広域特異性アミノ酸ラセマーゼ、配列番号140に対応する核酸配列によってコードされた広域特異性ラセマーゼ、ラセマーゼがI384M突然変異を含むような置換を有する、配列番号140に対応する核酸配列によってコードされた広域特異性ラセマーゼ、配列番号178に対応する核酸配列によってコードされた広域特異性ラセマーゼ、ラセマーゼがL383M突然変異を含むような置換を有する、配列番号178に対応する核酸配列によってコードされた広域特異性ラセマーゼ、配列番号193に対応する核酸配列によってコードされた広域特異性ラセマーゼ、配列番号176に対応する核酸配列によってコードされた広域特異性ラセマーゼ、ならびに
d)c)の広域特異性アミノ酸ラセマーゼをコードする遺伝子のいずれかに対して少なくとも90%の配列同一性パーセントを有する配列を含む酵素
から選ばれ、
R−MPからのR,Rモナチンの生成は、広域特異性を有するD−アミノトランスフェラーゼによって促進される該方法。 - D−トリプトファンからのインドール−3−ピルビン酸の生成が1つまたは複数の酵素によって促進され、R−モナチン前駆体からのR,Rモナチンの生成がD−トリプトファンからのインドール−3−ピルビン酸の生成を促進する酵素と同じ1つまたは複数の酵素によって促進されることを特徴とする請求項32に記載の方法。
- R−MPからのR,Rモナチンの生成が、
a)T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異、F200M突然変異、F200Y突然変異、ならびにF200MおよびT243Nの突然変異を有する、配列番号86に対応するD−アミノトランスフェラーゼのうちの1つまたは複数から選ばれるD−アミノトランスフェラーゼの活性を有する酵素、
b)a)の酵素のいずれかに対して少なくとも90%の配列同一性パーセントを有する配列を含む酵素、
c)バチルス・ハロデュランスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼとバチルス・スファエリクスD−アミノトランスフェラーゼのハイブリッドであるハイブリッドD−アミノトランスフェラーゼ、バチルス・スファエリクスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼ、バチルス・リケニホルミスD−アミノトランスフェラーゼ、ATCC7063からのD−アミノトランスフェラーゼ、D−アミノトランスフェラーゼ活性を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、または
d)a〜cの組合せ
から選ばれる広域特異性D−アミノトランスフェラーゼによって促進されることを特徴とする請求項32に記載の方法。 - I3PからのR−MPの生成が、R−MPをS−MPに対して1:1よりも大きな比で優先的に生成するアルドラーゼ酵素によって促進されることを特徴とする請求項32に記載の方法。
- アルドラーゼ酵素がR特異的アルドラーゼ酵素であることを特徴とする請求項35に記載の方法。
- a)T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異の組合せ、F200M突然変異、F200Y突然変異、ならびにF200MおよびT243Nの突然変異の組合せから選ばれる突然変異を有する、配列番号86に対応するD−アミノトランスフェラーゼ、
b)a)のいずれかに対して少なくとも90%の配列同一性パーセントを有し、突然変異を有する配列を含む酵素、
c)バチルス・ハロデュランスD−アミノトランスフェラーゼ、ハイブリッドD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼ、バチルス・リケニホルミスD−アミノトランスフェラーゼ、ATCC7063からのD−アミノトランスフェラーゼ、D−アミノトランスフェラーゼ活性を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、およびバチルス・スファエリクスD−アミノトランスフェラーゼから選ばれるD−アミノトランスフェラーゼ、ならびに
d)a)〜c)の組合せ
から選ばれるD−アミノトランスフェラーゼを用いてD−トリプトファンをアミノ基転移することを含むことを特徴とする方法。 - モナチンを生成することをさらに含み、少なくとも1つの精製工程をさらに含み、該R,Rモナチンが、全有機化合物の少なくとも約60重量%の純度の程度まで精製されることを特徴とする請求項37に記載の方法。
- 生成されたモナチンの少なくとも約75重量%が、R,Rモナチンであることを特徴とする請求項37に記載の方法。
- 生成されたモナチンの少なくとも約80重量%が、R,Rモナチンであることを特徴とする請求項37に記載の方法。
- a)配列番号86のD−アミノトランスフェラーゼ、
b)配列番号86に対して少なくとも90%の配列同一性パーセントを有する配列を含む酵素、
c)T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの二重突然変異、F200M突然変異、F200Y突然変異、ならびにF200MおよびT243Nの二重突然変異、またはその組合せから選ばれる1つまたは複数の突然変異を含む配列番号86のD−アミノトランスフェラーゼ
d)c)のD−アミノトランスフェラーゼに対して少なくとも90%の配列同一性パーセントを含み、1つまたは複数の突然変異を有するD−アミノトランスフェラーゼ、
e)配列番号87のD−アミノトランスフェラーゼ、ならびに
f)配列番号87に対して少なくとも90%の配列同一性パーセントを含むD−アミノトランスフェラーゼ
から選ばれる単離D−アミノトランスフェラーゼ。 - a)配列番号151、配列番号155、配列番号192、ならびにその活性断片および相同体、配列番号179ならびにその活性断片および相同体、配列番号177ならびにその活性断片および相同体、ならびに配列番号194ならびにその活性断片および相同体を含む広域特異性アミノ酸ラセマーゼ、ならびに
b)配列番号151、配列番号155、配列番号192およびその活性断片、配列番号179およびその活性断片、配列番号177およびその活性断片、ならびに配列番号194およびその活性断片に対して少なくとも90%の配列同一性パーセントを含む広域特異性アミノ酸ラセマーゼ酵素
から選ばれる単離広域特異性アミノ酸ラセマーゼ。 - 配列番号99を含むハイブリッドD−アミノトランスフェラーゼおよびその相同体ならびに配列番号99に対して少なくとも90%の配列同一性パーセントを含むハイブリッドD−アミノトランスフェラーゼ酵素から選ばれる単離ハイブリッドD−アミノトランスフェラーゼ。
- イー・コリF37Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリY96Fに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリY165Lに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL127Kに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリR98Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL108Rに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL110Hに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリL127Yに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、イー・コリR41Kに対応する突然変異を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼから選ばれた、D−アミノトランスフェラーゼ活性を有する単離されたバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ。
- 配列番号43を含むラセマーゼならびに配列番号43に対して少なくとも90%の配列同一性パーセントを含み、M35C、F66E、およびY354Aの突然変異を有するラセマーゼ酵素から選ばれる単離ラセマーゼ。
- 配列番号154を含む核酸、配列番号154に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号150を含む核酸、配列番号150に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号191を含む核酸、配列番号191に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号195を含む核酸、配列番号195に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号196を含む核酸、配列番号196に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号178を含む核酸、配列番号178に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号176を含む核酸、配列番号176に対して少なくとも90%の配列同一性パーセントを含む核酸、配列番号193を含む核酸、および配列番号193に対して少なくとも90%の配列同一性パーセントを含む核酸から選ばれる単離核酸。
- LトリプトファンおよびDトリプトファン、D−アミノトランスフェラーゼの活性を有する1つまたは複数の酵素、トリプトファンラセマーゼ、トリプトファンラセマーゼの活性を有するラセマーゼ、広域特異性アミノ酸ラセマーゼ、広域特異性アミノ酸ラセマーゼの活性を有するラセマーゼから選ばれる1つまたは複数の酵素、ならびにR特異的アルドラーゼの活性を有する1つまたは複数の酵素のうちの1つまたは複数を含む無細胞組成物。
- R−モナチン前駆体、L−トリプトファン、D−アミノ酸、およびモナチン前駆体またはモナチンのいずれかよりもL−トリプトファンに対するより高い活性、より高い特異性、またはその双方を有する1つまたは複数の酵素を含み、該1つまたは複数の第1の酵素は、L−トリプトファンアミノトランスフェラーゼ、L−芳香族アミノトランスフェラーゼ、L−アスパラギン酸アミノトランスフェラーゼ、L−アミノ酸オキシダーゼ、および立体反転アミノトランスフェラーゼから選ばれ、組成物は、D−アミノトランスフェラーゼおよびD−アミノ酸デヒドロゲナーゼから選ばれる1つまたは複数の酵素、R特異的アルドラーゼ活性を有する1つまたは複数の酵素、アラニンラセマーゼ、グルタミン酸ラセマーゼ、およびアスパラギン酸ラセマーゼから選ばれる1つまたは複数の酵素を含んでいていてもよい組成物。
- R,Rモナチンが、0.01%〜0.1%(容量/容量)界面活性剤および1%〜10%(容量/容量)ポリオールから選ばれる1つまたは複数の界面活性剤の存在下で生成されることを特徴とする請求項38に記載の方法。
- D−アミノトランスフェラーゼが、ピリドキサール−5’−リン酸の存在下で精製されることを特徴とする請求項32に記載の方法。
- 酵素が、固体担体上に固定化されることを特徴とする請求項1に記載の方法。
- 1つまたは複数のD−アミノトランスフェラーゼの存在下でモナチン前駆体を反応させて、モナチンを生成することを含み、1つまたは複数のD−アミノトランスフェラーゼは、
a)T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異の組合せ、F200M突然変異、F200Y突然変異、ならびにF200MおよびT243Nの突然変異の組合せまたはその組合せから選ばれる突然変異を有する、配列番号86に対応するD−アミノトランスフェラーゼ、
b)a)の酵素のいずれかに対して少なくとも90%の配列同一性パーセントを有し、突然変異を含むD−アミノトランスフェラーゼ酵素、ならびに
c)バチルス・ハロデュランスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼとバチルス・スファエリクスD−アミノトランスフェラーゼのハイブリッドであるハイブリッドD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼ、バチルス・リケニホルミスD−アミノトランスフェラーゼ、ATCC4978からのD−アミノトランスフェラーゼ、ATCC7063からのD−アミノトランスフェラーゼ、D−アミノトランスフェラーゼ活性を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、またはその組合せから選ばれる1つまたは複数のD−アミノトランスフェラーゼ
から選ばれることを特徴とする方法。 - (a)tnaT遺伝子、(b)改変フェニルアラニンパーミアーゼタンパク質、(c)一般的な芳香族アミノ酸パーミアーゼ、(d)一般的な芳香族アミノ酸パーミアーゼタンパク質の活性を付与する、配列の少なくともその部分およびフェニルアラニンパーミアーゼタンパク質の活性を付与する、配列の少なくともその部分を有するキメラタンパク質、(e)D−アミノ酸デアシラーゼ、ならびに(f)tRNAtrpおよび適切なD−アミノ酸デアシラーゼのうちの1つまたは複数を発現または過剰発現するように、モナチンを生成する微生物を遺伝子操作し、該微生物を用いて該モナチンを生成することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- 最小量のLアミノ酸のみを含有する培地中で、モナチンを生成する微生物を成長させ、該モナチンを生成することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- モナチンを生成している微生物からのインドール3ピルビン酸の分泌を阻止または低減することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- モナチンを生成している微生物によるインドール3ピルビン酸の取り込みを増加させることを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- インドール3ピルビン酸がモナチンを生成している微生物内で分解されるのを阻止または低減することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- モナチンを生成するように遺伝子操作された微生物のバイオマスを生成し、次いで該微生物でモナチンオペロンを誘導することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- モナチンを生成している微生物のための成長培地中でDトリプトファン以外のD−アミノ酸の使用を最小化することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- Dトリプトファンの取り込みの阻害が起こる閾値未満のレベルで、モナチンを生成する微生物のための成長培地中にDトリプトファンを提供し、該モナチンを生成することを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- (a)tnaT遺伝子、(b)改変フェニルアラニンパーミアーゼタンパク質、(c)一般的な芳香族アミノ酸パーミアーゼ、(d)一般的な芳香族アミノ酸パーミアーゼタンパク質の活性を付与する、配列の少なくともその部分およびフェニルアラニンパーミアーゼタンパク質の活性を付与する、配列の少なくともその部分を有するキメラタンパク質、(e)D−アミノ酸デアシラーゼ、ならびに(f)tRNAtrpおよび適切なD−アミノ酸デアシラーゼのうちの1つまたは複数を発現または過剰発現するように遺伝子操作された、モナチンを生成する微生物。
- モナチン前駆体と、T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異、F200M突然変異、F200Y突然変異、F200MおよびT243Nの突然変異、またはその組合せを有する、配列番号86に対応するD−アミノトランスフェラーゼから選ばれる1つまたは複数のD−アミノトランスフェラーゼとを反応させることを含むことを特徴とする、モナチンまたはその塩を生成するための方法。
- モナチン前駆体と、T243S突然変異、T243N突然変異、N100A突然変異、T243Q突然変異、T243NおよびN100Aの突然変異、F200M突然変異、F200Y突然変異、F200MおよびT243Nの突然変異、またはその組合せを有する、配列番号86に対応するD−アミノトランスフェラーゼのうちの1つまたは複数から選ばれるD−アミノトランスフェラーゼに対して少なくとも90%の配列同一性パーセントを有する配列を含む1つまたは複数のD−アミノトランスフェラーゼとを反応させること
を含むことを特徴とする、モナチンまたはその塩を生成するための方法。 - 該配列同一性パーセントが少なくとも95%であることを特徴とする請求項63に記載の方法。
- モナチン前駆体と、バチルス・ハロデュランスD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼとバチルス・スファエリクスD−アミノトランスフェラーゼのハイブリッドであるハイブリッドD−アミノトランスフェラーゼ、ジオバチルス・ステアロサーモフィルスD−アミノトランスフェラーゼ、バチルス・リケニホルミスD−アミノトランスフェラーゼ、ATCC4978からのD−アミノトランスフェラーゼ、ATCC7063からのD−アミノトランスフェラーゼ、D−アミノトランスフェラーゼ活性を有するバチルス・リケニホルミス分岐鎖アミノトランスフェラーゼ、またはその組合せから選ばれる1つまたは複数のD−アミノトランスフェラーゼとを反応させること
を含むことを特徴とする、モナチンまたはその塩を生成するための方法。 - ビブリオ・フィシェリからのアラニンラセマーゼ、Genbank受入番号AAW85230.1のアラニンラセマーゼ、Genbank受入番号YP_204118のアラニンラセマーゼ、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼに対応する広域特異性アミノ酸ラセマーゼ、配列番号116を含む広域特異性アミノ酸ラセマーゼ、Genbank受入番号ZP_00898332.1 GI:82735470の広域特異性アミノ酸ラセマーゼ、ATCC4683からの広域特異性アミノ酸ラセマーゼ、配列番号204の広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、配列番号204に対応する広域特異性アミノ酸ラセマーゼ、ATCC7966からの広域特異性アミノ酸ラセマーゼ、シュードモナス株2150から単離された広域特異性アミノ酸ラセマーゼ、P.オレオボランスから単離された広域特異性アミノ酸ラセマーゼ、P.オーレオファシエンスから単離された広域特異性アミノ酸ラセマーゼ、P.フルオレッセンスから単離された広域特異性アミノ酸ラセマーゼ、P.グラベオレンスから単離された広域特異性アミノ酸ラセマーゼ、P.ストリアタAKU083から単離された広域特異性アミノ酸ラセマーゼ、アミノ酸24〜409を含有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号204のアミノ酸24〜409を含有する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ジャンデイから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC49572からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号194の広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ソブリアから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC35994からの広域特異性ラセマーゼの活性を有する酵素、配列番号192の広域特異性アミノ酸ラセマーゼの活性を有する酵素、L383M突然変異を有する、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼに対応する酵素、L383M突然変異を有する、ATCC14486からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号195を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号196を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号201を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号202を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、D76N突然変異を有する、エロモナス・キャビエからのアミノ酸ラセマーゼに対応する酵素、D76N突然変異を有する、配列番号179に対応する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ハイドロフィラから単離された広域特異性アミノ酸ラセマーゼ、配列番号177の広域特異性アミノ酸ラセマーゼ、配列番号177のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号146を含む広域特異性アミノ酸ラセマーゼ、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼ、配列番号179の広域特異性アミノ酸ラセマーゼ、配列番号179のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号155を含む広域特異性アミノ酸ラセマーゼ、配列番号151の広域特異性アミノ酸ラセマーゼ、その活性断片、およびその組合せから選ばれる1つまたは複数の酵素を利用することを含む、
(a)LトリプトファンからDトリプトファンまたは(b)DトリプトファンからLトリプトファンを生成することを含むことを特徴とする方法。 - ビブリオ・フィシェリからのアラニンラセマーゼ、Genbank受入番号AAW85230.1のアラニンラセマーゼ、Genbank受入番号YP_204118のアラニンラセマーゼ、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼに対応する広域特異性アミノ酸ラセマーゼ、配列番号116を含む広域特異性アミノ酸ラセマーゼ、Genbank受入番号ZP_00898332.1 GI:82735470の広域特異性アミノ酸ラセマーゼ、ATCC4683からの広域特異性アミノ酸ラセマーゼ、配列番号204の広域特異性アミノ酸ラセマーゼ、I384M突然変異を有する、配列番号204に対応する広域特異性アミノ酸ラセマーゼ、ATCC7966からの広域特異性アミノ酸ラセマーゼ、シュードモナス株2150から単離された広域特異性アミノ酸ラセマーゼ、P.オレオボランスから単離された広域特異性アミノ酸ラセマーゼ、P.オーレオファシエンスから単離された広域特異性アミノ酸ラセマーゼ、P.フルオレッセンスから単離された広域特異性アミノ酸ラセマーゼ、P.グラベオレンスから単離された広域特異性アミノ酸ラセマーゼ、P.ストリアタAKU083から単離された広域特異性アミノ酸ラセマーゼ、アミノ酸24〜409を含有する、シュードモナス・タエトロレンスから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号204のアミノ酸24〜409を含有する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ジャンデイから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC49572からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号194の広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ソブリアから単離された広域特異性アミノ酸ラセマーゼの活性を有する酵素、ATCC35994からの広域特異性ラセマーゼの活性を有する酵素、配列番号192の広域特異性アミノ酸ラセマーゼの活性を有する酵素、L383M突然変異を有する、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼに対応する酵素、L383M突然変異を有する、ATCC14486からの広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号195を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号196を含む遺伝子によってコードされた広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号201を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、配列番号202を含む広域特異性アミノ酸ラセマーゼの活性を有する酵素、D76N突然変異を有する、エロモナス・キャビエからのアミノ酸ラセマーゼに対応する酵素、D76N突然変異を有する、配列番号179に対応する広域特異性アミノ酸ラセマーゼの活性を有する酵素、エロモナス・ハイドロフィラから単離された広域特異性アミノ酸ラセマーゼ、配列番号177の広域特異性アミノ酸ラセマーゼ、配列番号177のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号146を含む広域特異性アミノ酸ラセマーゼ、エロモナス・キャビエから単離された広域特異性アミノ酸ラセマーゼ、配列番号179の広域特異性アミノ酸ラセマーゼ、配列番号179のアミノ酸22〜408を含有する広域特異性アミノ酸ラセマーゼ、配列番号155を含む広域特異性アミノ酸ラセマーゼ、配列番号151の広域特異性アミノ酸ラセマーゼ、その活性断片から選ばれる1つまたは複数の広域特異性アミノ酸ラセマーゼに対して少なくとも90%の配列同一性パーセントを有する配列を含む1つまたは複数の酵素を利用することを含む、
(a)LトリプトファンからDトリプトファンまたは(b)DトリプトファンからLトリプトファンを生成することを含むことを特徴とする方法。 - 該配列同一性パーセントが少なくとも95%であることを特徴とする請求項67に記載の方法。
- ピリドキサール−5’−リン酸の存在下でD−アミノトランスフェラーゼを精製することを含むことを特徴とする方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US58401606A | 2006-10-20 | 2006-10-20 | |
| US11/714,279 US8076108B2 (en) | 2005-04-26 | 2007-03-06 | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
| PCT/US2007/081932 WO2008085575A2 (en) | 2006-10-20 | 2007-10-19 | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
Related Child Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015122370A Division JP2015211679A (ja) | 2006-10-20 | 2015-06-17 | モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JP2010507367A true JP2010507367A (ja) | 2010-03-11 |
| JP2010507367A5 JP2010507367A5 (ja) | 2011-01-13 |
Family
ID=39615760
Family Applications (2)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2009533566A Pending JP2010507367A (ja) | 2006-10-20 | 2007-10-19 | モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 |
| JP2015122370A Pending JP2015211679A (ja) | 2006-10-20 | 2015-06-17 | モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 |
Family Applications After (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP2015122370A Pending JP2015211679A (ja) | 2006-10-20 | 2015-06-17 | モナチンの立体異性体およびそれらの前駆体の生成のためのポリペプチドおよび生合成経路 |
Country Status (7)
| Country | Link |
|---|---|
| US (2) | US8076108B2 (ja) |
| EP (1) | EP2089529B1 (ja) |
| JP (2) | JP2010507367A (ja) |
| AR (1) | AR063348A1 (ja) |
| AU (1) | AU2007342275B2 (ja) |
| CA (1) | CA2666348A1 (ja) |
| WO (1) | WO2008085575A2 (ja) |
Cited By (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012147674A1 (ja) * | 2011-04-25 | 2012-11-01 | 味の素株式会社 | モナティンの製造方法 |
| CN106636227A (zh) * | 2016-09-22 | 2017-05-10 | 精晶药业股份有限公司 | 一种酶法转化l‑丙氨酸制备丙酮酸的方法 |
Families Citing this family (28)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US7297800B2 (en) | 2001-12-27 | 2007-11-20 | Ajinomoto Co., Inc. | Process of producing glutamate derivatives |
| US7572607B2 (en) * | 2002-04-23 | 2009-08-11 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of monatin and its precursors |
| US8372989B2 (en) * | 2002-04-23 | 2013-02-12 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of monatin and its precursors |
| RU2307871C2 (ru) | 2002-08-26 | 2007-10-10 | Адзиномото Ко., Инк. | НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ЗАМЕЩЕННЫХ α-КЕТОКИСЛОТ |
| AU2003289264A1 (en) * | 2002-12-09 | 2004-06-30 | Ajinomoto Co., Inc. | Mutant d-aminotransferase and process for producing optically active glutamic acid derivative using the same |
| WO2005014839A2 (en) * | 2003-08-01 | 2005-02-17 | Cargill, Incorporated | Monatin tabletop sweetener compositions and methods of making same |
| WO2005016022A1 (en) * | 2003-08-14 | 2005-02-24 | Cargill, Incorporated | Chewing gum compositions comprising monatin and methods of making same |
| WO2005020721A1 (en) * | 2003-08-25 | 2005-03-10 | Cargill, Incorporated | Beverage compositions comprising monatin and methods of making same |
| EP1678313B1 (en) * | 2003-10-21 | 2011-02-16 | Cargill, Incorporated | Production of monatin and monatin precursors |
| US8153405B2 (en) * | 2005-04-20 | 2012-04-10 | Cargill, Incorporated | Products and methods for in vivo secretion of monatin |
| US8158389B2 (en) * | 2005-04-20 | 2012-04-17 | Cargill, Incorporated | Products and methods for in vivo secretion of monatin |
| US8076108B2 (en) * | 2005-04-26 | 2011-12-13 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
| US7582455B2 (en) * | 2005-04-26 | 2009-09-01 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
| MX2008011477A (es) * | 2006-03-07 | 2008-09-23 | Cargill Inc | Aldolasas, acidos nucleicos que las codifican y metodos para hacerlas y usarlas. |
| AU2007223086B2 (en) * | 2006-03-07 | 2013-09-26 | Basf Enzymes Llc | Aldolases, nucleic acids encoding them and methods for making and using them |
| US20090198072A1 (en) * | 2006-05-24 | 2009-08-06 | Cargill, Incorporated | Methods and systems for increasing production of equilibrium reactions |
| WO2007140195A2 (en) * | 2006-05-24 | 2007-12-06 | Cargill, Incorporated | Methods and systems for increasing production of equilibrium reactions |
| CN101239941B (zh) * | 2007-02-08 | 2012-02-29 | 味之素株式会社 | 光学活性化合物的制造方法 |
| US8076107B2 (en) * | 2007-10-01 | 2011-12-13 | Cargill, Incorporated | Production of monatin stereoisomers |
| US8367847B2 (en) * | 2007-10-01 | 2013-02-05 | Cargill, Incorporated | Production of monatin enantiomers |
| US8003361B2 (en) * | 2007-10-01 | 2011-08-23 | Cargill Incorporated | Production of monatin enantiomers |
| CA2726928A1 (en) * | 2008-01-03 | 2009-07-16 | Cargill, Incorporated | Aminotransferase and oxidoreductase nucleic acids and polypeptides and methods of using |
| WO2009088442A2 (en) * | 2008-01-03 | 2009-07-16 | Cargill, Incorporated | Isomerases and epimerases and methods of using |
| EP2479272A4 (en) | 2010-10-14 | 2013-11-20 | Ajinomoto Kk | PROCESS FOR THE PRODUCTION OF MONATIN |
| US8771997B2 (en) | 2011-04-20 | 2014-07-08 | Ajinomoto Co., Inc. | Method for producing monatin using an L-amino acid aminotransferase |
| ES2538720B1 (es) * | 2013-12-20 | 2016-05-12 | Universidad Autónoma de Madrid | Racemasa BsrAh para la racemización de aminoácidos |
| WO2017151997A1 (en) * | 2016-03-04 | 2017-09-08 | President And Fellows Of Harvard College | Methods for making polypeptides including d-amino acids |
| CN113025602A (zh) * | 2021-03-18 | 2021-06-25 | 绵阳晟氏健康科技有限公司 | 固定化精氨酸消旋酶及其制备方法、应用 |
Citations (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000500981A (ja) * | 1995-11-30 | 2000-02-02 | マキシジェン,インコーポレイテッド | 反復的選択および組換えにより所望の特徴を有するポリヌクレオチドを作製する方法 |
| JP2001508660A (ja) * | 1997-01-24 | 2001-07-03 | バイオインヴェント インターナショナル アーベー | タンパク質機能のインビトロ分子進化の方法 |
| JP2004222657A (ja) * | 2003-01-24 | 2004-08-12 | Ajinomoto Co Inc | 2r−モナティンの製造方法 |
| WO2005016022A1 (en) * | 2003-08-14 | 2005-02-24 | Cargill, Incorporated | Chewing gum compositions comprising monatin and methods of making same |
| WO2005020721A1 (en) * | 2003-08-25 | 2005-03-10 | Cargill, Incorporated | Beverage compositions comprising monatin and methods of making same |
| JP2005523696A (ja) * | 2002-04-23 | 2005-08-11 | カーギル,インコーポレイティド | ポリペプチド及び生合成経路 |
| US20050282260A1 (en) * | 2002-04-23 | 2005-12-22 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of monatin and its precursors |
Family Cites Families (71)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3002889A (en) | 1960-06-20 | 1961-10-03 | Kyowa Hakko Kogyo Kk | Method of producing l-glutamic acid |
| US3128237A (en) | 1961-02-24 | 1964-04-07 | Ajinomoto Kk | Process for producing l-glutamic acid by bacterial fermentation |
| US3399114A (en) | 1964-08-22 | 1968-08-27 | Asahi Chemical Ind | Process for producing l-glutamic acid by using bacteria |
| US4371614A (en) | 1980-08-22 | 1983-02-01 | Ajinomoto Co., Inc. | E.Coli bacteria carrying recombinant plasmids and their use in the fermentative production of L-tryptophan |
| EP0232262A4 (en) | 1985-08-15 | 1989-09-19 | Stauffer Chemical Co | TRYPTOPHANE GENERATING MICROORGANISM. |
| GB2205834B (en) | 1987-06-15 | 1990-10-31 | South African Inventions | 3-(1-amino-1,3-dicarboxy-3-hydroxy-but-4-yl)-indole compounds |
| US5300437A (en) | 1989-06-22 | 1994-04-05 | Celgene Corporation | Enantiomeric enrichment and stereoselective synthesis of chiral amines |
| WO1989011212A2 (en) | 1989-08-30 | 1989-11-30 | Wm. Wrigley Jr. Company | Method of controlling release of cyclamate in chewing gum and gum produced thereby |
| ATE104961T1 (de) | 1990-01-19 | 1994-05-15 | Technology Finance Corp | Verfahren zur herstellung von 3-(1-amino-1,3dicarboxy-3-hydroxy-but-4-yl)-indol. |
| US5360724A (en) | 1992-12-18 | 1994-11-01 | Celgene Corporation | Process for the preparation of chiral 1-aryl-2-aminopropanes |
| US6264999B1 (en) | 1993-09-30 | 2001-07-24 | Wm. Wrigley Jr. Company | Chewing gum containing erythritol and method of making |
| JP3698742B2 (ja) | 1994-03-01 | 2005-09-21 | 協和醗酵工業株式会社 | 光学活性4−ヒドロキシ−2−ケトグルタル酸の製造法 |
| US5985617A (en) | 1997-02-18 | 1999-11-16 | Liao; James C. | Microorganisms and methods for overproduction of DAHP by cloned PPS gene |
| CA2234412C (en) | 1997-06-09 | 2008-09-02 | Kyowa Hakko Kogyo Co., Ltd. | Method for producing optically active compound |
| US6099848A (en) * | 1997-11-18 | 2000-08-08 | The Trustees Of The University Of Pennsylvania | Immunogenic compositions comprising DAL/DAT double-mutant, auxotrophic, attenuated strains of Listeria and their methods of use |
| DE19818541C2 (de) | 1998-04-24 | 2003-04-10 | Forschungszentrum Juelich Gmbh | Mikrobielle Herstellung von Substanzen aus dem aromatischen Stoffwechsel / III |
| US5994559A (en) | 1998-08-06 | 1999-11-30 | The Board Of Governors For Higher Education, State Of Rhode Island And Providence Plantations | Synthesis of monatin-A high intensity natural sweetener |
| JP2002060382A (ja) | 2000-08-22 | 2002-02-26 | Ajinomoto Co Inc | モナティンの立体異性体及びその使用、並びにモナティン類の製造方法及びそのための中間体 |
| DE10129711B4 (de) | 2001-06-22 | 2007-08-23 | Forschungszentrum Jülich GmbH | Verfahren zur fermentativen Herstellung von Pyruvat |
| AU2002349604A1 (en) | 2001-11-30 | 2003-06-10 | Ajinomoto Co., Inc. | Crystals of non-natural stereoisomer salts of monatin and utilization thereof |
| JP2003171365A (ja) | 2001-11-30 | 2003-06-20 | Ajinomoto Co Inc | モナティン類及びその製造中間体の製造方法並びに新規中間体 |
| CN100549182C (zh) | 2001-12-27 | 2009-10-14 | 味之素株式会社 | 谷氨酸衍生物的制备方法 |
| JP4196832B2 (ja) | 2001-12-27 | 2008-12-17 | 味の素株式会社 | グルタミン酸化合物及びそれらの製造中間体の製造方法並びにそのための新規中間体 |
| US7297800B2 (en) * | 2001-12-27 | 2007-11-20 | Ajinomoto Co., Inc. | Process of producing glutamate derivatives |
| WO2003074690A1 (en) | 2002-03-01 | 2003-09-12 | Kyowa Hakko Kogyo Co., Ltd. | Amino acid racemase having low substrate specificity and process for producing racemic amino acid |
| JP2003292484A (ja) | 2002-04-01 | 2003-10-15 | Ajinomoto Co Inc | γ−ヒドロキシアミノ酸誘導体及びモナティン類の製造方法 |
| US8372989B2 (en) | 2002-04-23 | 2013-02-12 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of monatin and its precursors |
| RU2307871C2 (ru) | 2002-08-26 | 2007-10-10 | Адзиномото Ко., Инк. | НОВАЯ АЛЬДОЛАЗА И СПОСОБ ПОЛУЧЕНИЯ ЗАМЕЩЕННЫХ α-КЕТОКИСЛОТ |
| AU2003289264A1 (en) | 2002-12-09 | 2004-06-30 | Ajinomoto Co., Inc. | Mutant d-aminotransferase and process for producing optically active glutamic acid derivative using the same |
| CN100408552C (zh) | 2003-01-09 | 2008-08-06 | 味之素株式会社 | 谷氨酸衍生物和焦谷氨酸衍生物的制造方法和新型的制造用中间体 |
| WO2004085624A2 (en) | 2003-03-24 | 2004-10-07 | Diversa Corporation | Transaminases, deaminases and aminomutases and compositions and methods for enzymatic detoxification |
| JP2004331650A (ja) | 2003-04-14 | 2004-11-25 | Kanazawa Univ Tlo Inc | モナチン類の製造法 |
| JP2004331644A (ja) | 2003-04-14 | 2004-11-25 | Kanazawa Univ Tlo Inc | モナチン類の製造法 |
| WO2005001105A1 (ja) | 2003-06-26 | 2005-01-06 | Ajinomoto Co., Inc. | モナティンの製造方法 |
| WO2005014839A2 (en) | 2003-08-01 | 2005-02-17 | Cargill, Incorporated | Monatin tabletop sweetener compositions and methods of making same |
| EP1678313B1 (en) | 2003-10-21 | 2011-02-16 | Cargill, Incorporated | Production of monatin and monatin precursors |
| JP4500977B2 (ja) | 2003-11-05 | 2010-07-14 | 味の素株式会社 | 新規アミノ酸誘導体及び甘味剤 |
| JP5113978B2 (ja) | 2003-11-21 | 2013-01-09 | 味の素株式会社 | グルタミン酸誘導体の有機アミン塩及びその利用 |
| KR101070183B1 (ko) | 2004-02-27 | 2011-10-05 | 아지노모토 가부시키가이샤 | 모나틴의 제조방법 |
| JP4604524B2 (ja) | 2004-03-16 | 2011-01-05 | 味の素株式会社 | 変異型アルドラーゼ、並びにこれを用いた光学活性ihog及び光学活性モナティンの製造方法 |
| JP4604537B2 (ja) | 2004-03-31 | 2011-01-05 | 味の素株式会社 | L−システイン生産菌及びl−システインの製造法 |
| US7935377B2 (en) | 2004-06-04 | 2011-05-03 | Ajinomoto Co., Inc. | Crystals of free (2R, 4R)-monatin and use thereof |
| CA2506247C (en) | 2004-06-07 | 2012-02-21 | Ajinomoto Co., Inc. | Novel aldolase, and method for producing optically active ihog and monatin |
| KR101216575B1 (ko) | 2004-07-14 | 2012-12-31 | 아지노모토 가부시키가이샤 | (2r,4r)-모나틴 칼륨 염 결정 및 이를 함유하는 감미료조성물 |
| JP2007284349A (ja) | 2004-07-27 | 2007-11-01 | Ajinomoto Co Inc | モナティンまたはその塩の製造方法 |
| ATE554178T1 (de) | 2004-10-05 | 2012-05-15 | Ajinomoto Kk | Verfahren zur herstellung von aminosäurederivaten aus hydroxyiminosäuren |
| CN101035442B (zh) | 2004-10-15 | 2012-04-11 | 味之素株式会社 | 甜味剂组合物 |
| JP2008530978A (ja) | 2005-03-03 | 2008-08-14 | 味の素株式会社 | 4−ハイドロキシ−l−イソロイシン又はその塩の製造法 |
| US8158389B2 (en) | 2005-04-20 | 2012-04-17 | Cargill, Incorporated | Products and methods for in vivo secretion of monatin |
| US8153405B2 (en) | 2005-04-20 | 2012-04-10 | Cargill, Incorporated | Products and methods for in vivo secretion of monatin |
| US7582455B2 (en) * | 2005-04-26 | 2009-09-01 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
| US8076108B2 (en) | 2005-04-26 | 2011-12-13 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
| US7524660B2 (en) | 2005-05-05 | 2009-04-28 | E.I. Du Pont De Nemours And Company | Utilization of fructose in microbial production strains |
| AU2007223086B2 (en) | 2006-03-07 | 2013-09-26 | Basf Enzymes Llc | Aldolases, nucleic acids encoding them and methods for making and using them |
| MX2008011477A (es) | 2006-03-07 | 2008-09-23 | Cargill Inc | Aldolasas, acidos nucleicos que las codifican y metodos para hacerlas y usarlas. |
| RU2338784C2 (ru) | 2006-03-24 | 2008-11-20 | Закрытое акционерное общество "Научно-исследовательский институт Аджиномото-Генетика" (ЗАО АГРИ) | Новая альдолаза, днк, кодирующая альдолазу, клетки, трансформированные днк, способ получения альдолазы и способ получения 4-гидрокси-l-изолейцина (варианты) |
| WO2007140195A2 (en) | 2006-05-24 | 2007-12-06 | Cargill, Incorporated | Methods and systems for increasing production of equilibrium reactions |
| US20090198072A1 (en) | 2006-05-24 | 2009-08-06 | Cargill, Incorporated | Methods and systems for increasing production of equilibrium reactions |
| US7354746B1 (en) | 2006-12-11 | 2008-04-08 | Ajinomoto Co., Inc. | Method for producing optically active IHOG and monatin |
| CN101239941B (zh) | 2007-02-08 | 2012-02-29 | 味之素株式会社 | 光学活性化合物的制造方法 |
| US8367847B2 (en) | 2007-10-01 | 2013-02-05 | Cargill, Incorporated | Production of monatin enantiomers |
| US8003361B2 (en) | 2007-10-01 | 2011-08-23 | Cargill Incorporated | Production of monatin enantiomers |
| US8076107B2 (en) | 2007-10-01 | 2011-12-13 | Cargill, Incorporated | Production of monatin stereoisomers |
| WO2011002469A1 (en) | 2009-07-02 | 2011-01-06 | Cargill, Incorporated | Isomerases and epimerases and methods of using |
| CA2726928A1 (en) | 2008-01-03 | 2009-07-16 | Cargill, Incorporated | Aminotransferase and oxidoreductase nucleic acids and polypeptides and methods of using |
| BRPI1009446A2 (pt) | 2009-03-12 | 2015-08-18 | Cargill Inc | Composições compreendendo monatina e cálcio |
| US20120076899A1 (en) | 2009-05-28 | 2012-03-29 | Cargill, Incorporated | Shelf stable monatin sweetened beverage |
| CN102686391A (zh) | 2009-12-30 | 2012-09-19 | 嘉吉公司 | 聚合物及制备和使用聚合物的方法 |
| WO2011082363A1 (en) | 2009-12-30 | 2011-07-07 | Cargill, Incorporated | Membrane separation and purification of monatin |
| WO2011082353A1 (en) | 2009-12-30 | 2011-07-07 | Cargill, Incorporated | Separation and purification of monatin |
| WO2011082365A2 (en) | 2009-12-30 | 2011-07-07 | Cargill, Incorporated | Improved bioreactor performance in the production of monatin |
-
2007
- 2007-03-06 US US11/714,279 patent/US8076108B2/en not_active Expired - Fee Related
- 2007-10-19 AR ARP070104644A patent/AR063348A1/es not_active Application Discontinuation
- 2007-10-19 AU AU2007342275A patent/AU2007342275B2/en not_active Ceased
- 2007-10-19 WO PCT/US2007/081932 patent/WO2008085575A2/en not_active Ceased
- 2007-10-19 JP JP2009533566A patent/JP2010507367A/ja active Pending
- 2007-10-19 EP EP07872326.9A patent/EP2089529B1/en not_active Not-in-force
- 2007-10-19 CA CA002666348A patent/CA2666348A1/en not_active Abandoned
-
2011
- 2011-11-02 US US13/287,900 patent/US8975046B2/en not_active Expired - Fee Related
-
2015
- 2015-06-17 JP JP2015122370A patent/JP2015211679A/ja active Pending
Patent Citations (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP2000500981A (ja) * | 1995-11-30 | 2000-02-02 | マキシジェン,インコーポレイテッド | 反復的選択および組換えにより所望の特徴を有するポリヌクレオチドを作製する方法 |
| JP2001508660A (ja) * | 1997-01-24 | 2001-07-03 | バイオインヴェント インターナショナル アーベー | タンパク質機能のインビトロ分子進化の方法 |
| JP2005523696A (ja) * | 2002-04-23 | 2005-08-11 | カーギル,インコーポレイティド | ポリペプチド及び生合成経路 |
| US20050282260A1 (en) * | 2002-04-23 | 2005-12-22 | Cargill, Incorporated | Polypeptides and biosynthetic pathways for the production of monatin and its precursors |
| JP2004222657A (ja) * | 2003-01-24 | 2004-08-12 | Ajinomoto Co Inc | 2r−モナティンの製造方法 |
| WO2005016022A1 (en) * | 2003-08-14 | 2005-02-24 | Cargill, Incorporated | Chewing gum compositions comprising monatin and methods of making same |
| WO2005020721A1 (en) * | 2003-08-25 | 2005-03-10 | Cargill, Incorporated | Beverage compositions comprising monatin and methods of making same |
Non-Patent Citations (5)
| Title |
|---|
| CSNG199901368002; 伏見 譲 Yuzuru HUSIMI: '生物学の立場から:進化分子工学 A-Life from a Biological Prespective: Evolutionary Molocular Enginee' 電子情報通信学会誌 第77巻 第2号 THE JOURNAL OF THE INSTITUTE OF ELECTRONICS,INFORMATION AND CO 第77巻, 社団法人電子情報通信学会 THE INSTITUTE OF ELECTRO * |
| CSNG200501163006; 伏見 譲 Yuzuru Husimi: '進化の周辺 Around Evolutinary Systems' 人工知能学会誌 第19巻 第6号 Journal of the Japanese Society for Artificial Intelligence 第19巻, (社)人工知能学会 The Japanese Society for Artif * |
| JPN6014009629; 伏見 譲 Yuzuru Husimi: '進化の周辺 Around Evolutinary Systems' 人工知能学会誌 第19巻 第6号 Journal of the Japanese Society for Artificial Intelligence 第19巻, (社)人工知能学会 The Japanese Society for Artif * |
| JPN6014009631; 伏見 譲 Yuzuru HUSIMI: '生物学の立場から:進化分子工学 A-Life from a Biological Prespective: Evolutionary Molocular Enginee' 電子情報通信学会誌 第77巻 第2号 THE JOURNAL OF THE INSTITUTE OF ELECTRONICS,INFORMATION AND CO 第77巻, 社団法人電子情報通信学会 THE INSTITUTE OF ELECTRO * |
| JPN6015005666; Appl. Microbiol. Biotechnol. vol.73, 20061007, pp.1299-1305 * |
Cited By (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2012147674A1 (ja) * | 2011-04-25 | 2012-11-01 | 味の素株式会社 | モナティンの製造方法 |
| CN106636227A (zh) * | 2016-09-22 | 2017-05-10 | 精晶药业股份有限公司 | 一种酶法转化l‑丙氨酸制备丙酮酸的方法 |
| CN106636227B (zh) * | 2016-09-22 | 2020-11-06 | 精晶药业股份有限公司 | 一种酶法转化l-丙氨酸制备丙酮酸的方法 |
Also Published As
| Publication number | Publication date |
|---|---|
| US8975046B2 (en) | 2015-03-10 |
| WO2008085575A2 (en) | 2008-07-17 |
| WO2008085575A3 (en) | 2009-03-19 |
| EP2089529A2 (en) | 2009-08-19 |
| EP2089529B1 (en) | 2019-03-20 |
| EP2089529A4 (en) | 2012-04-11 |
| CA2666348A1 (en) | 2008-07-17 |
| AR063348A1 (es) | 2009-01-21 |
| JP2015211679A (ja) | 2015-11-26 |
| US20120164697A1 (en) | 2012-06-28 |
| AU2007342275A2 (en) | 2009-07-09 |
| AU2007342275B2 (en) | 2013-06-20 |
| AU2007342275A1 (en) | 2008-07-17 |
| US8076108B2 (en) | 2011-12-13 |
| US20080020434A1 (en) | 2008-01-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| US8076108B2 (en) | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors | |
| US7582455B2 (en) | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors | |
| JP4988803B2 (ja) | ポリペプチド及び生合成経路 | |
| JP4829117B2 (ja) | モナチンおよびモナチン前駆体の製造 | |
| CN101374954B (zh) | 用于生产莫纳甜和其前体的多肽和生物合成途径 | |
| CN101365787B (zh) | 用于生产莫纳甜的立体异构体和其前体的多肽和生物合成途径 | |
| AU2012201740B2 (en) | Polypeptides and biosynthetic pathways for the production of stereoisomers of monatin and their precursors |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20101018 |
|
| A621 | Written request for application examination |
Free format text: JAPANESE INTERMEDIATE CODE: A621 Effective date: 20101018 |
|
| RD04 | Notification of resignation of power of attorney |
Free format text: JAPANESE INTERMEDIATE CODE: A7424 Effective date: 20110927 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20130319 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20130618 |
|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20140304 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140603 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140610 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20140704 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20140711 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20140724 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20150217 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20150617 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20150629 |
|
| A912 | Re-examination (zenchi) completed and case transferred to appeal board |
Free format text: JAPANESE INTERMEDIATE CODE: A912 Effective date: 20150828 |