JP3736574B2 - アスペルギルス オリゼ(Aspergillus oryzae)におけるβ−ガラクトシダーゼの生産増加 - Google Patents
アスペルギルス オリゼ(Aspergillus oryzae)におけるβ−ガラクトシダーゼの生産増加 Download PDFInfo
- Publication number
- JP3736574B2 JP3736574B2 JP50336496A JP50336496A JP3736574B2 JP 3736574 B2 JP3736574 B2 JP 3736574B2 JP 50336496 A JP50336496 A JP 50336496A JP 50336496 A JP50336496 A JP 50336496A JP 3736574 B2 JP3736574 B2 JP 3736574B2
- Authority
- JP
- Japan
- Prior art keywords
- lactase
- aspergillus oryzae
- dna
- sequence
- aspergillus
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Lifetime
Links
Images








Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/24—Hydrolases (3) acting on glycosyl compounds (3.2)
- C12N9/2402—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1)
- C12N9/2468—Hydrolases (3) acting on glycosyl compounds (3.2) hydrolysing O- and S- glycosyl compounds (3.2.1) acting on beta-galactose-glycoside bonds, e.g. carrageenases (3.2.1.83; 3.2.1.157); beta-agarase (3.2.1.81)
- C12N9/2471—Beta-galactosidase (3.2.1.23), i.e. exo-(1-->4)-beta-D-galactanase
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y302/00—Hydrolases acting on glycosyl compounds, i.e. glycosylases (3.2)
- C12Y302/01—Glycosidases, i.e. enzymes hydrolysing O- and S-glycosyl compounds (3.2.1)
- C12Y302/01023—Beta-galactosidase (3.2.1.23), i.e. exo-(1-->4)-beta-D-galactanase
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Enzymes And Modification Thereof (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Medicines Containing Material From Animals Or Micro-Organisms (AREA)
Description
本発明は、コウジカビ(アスペルギルス・オリゼ)(Aspergillus oryzae)におけるラクターゼの産生増加に関する。さらに特定すると、本発明は、アスペルギルス・オリゼ ラクターゼをコードしているDNA配列、ならびにラクターゼ産生の制御、発現および分泌に必要な付加的なDNAからなるDNAを用いて宿主細胞を形質転換することにより、アスペルギルス(コウジカビ)属(Aspergillus)宿主細胞内でアスペルギルス ラクターゼの産生を増加させることに関する。
発明の背景
ラクトースは牛乳やホエイ中に含まれる主な糖質である(約42%を占めている)。ホエイとは、チーズの生産工程においてクリームや牛乳から固形分を除去した後に残っている液体である。
β−D−ガラクトシダーゼ(β−D−ガラクトシドガラクトヒドロラーゼ、EC3.2.1.23、ラクターゼとも呼ばれる)は、二糖類であるラクトースをグルコースとガラクトースに加水分解する酵素である。牛乳およびホエイにラクトースが存在することにより、一部の人々にはいくつかの医学的問題が生じると考えられている。これらの医学的問題は一般的に「ラクトース不耐性」と呼ばれている。例えば、アジア人、アフリカ人、およびアフロアメリカ人では高い割合(〜90%)で、ならびにおよそ5%の白系アメリカ人および西ヨーロッパ人ではβ−ガラクトシダーゼ(ラクターゼ)が欠損しているといわれている。(The Lancet, Vol.338, 9月14日号, 1994, p.663-664参照。)ラクターゼ酵素の欠損による影響は、典型的なものとして、ラクトースを含有する製品、例えば、牛乳、アイスクリームまたはその他の乳製品を摂取した後の腹部膨満、腹鳴、および/または下痢として現れる。子供もある種のラクトース不耐性になりやすいが、これは、ガラクトースからグルコースへの変換がうまくできないためである。ガラクトースが血中に蓄積すると肝臓が肥大する。この状態をガラクトース血症とよぶ。
3000万から5000万人が何らかの形のラクトース不耐性に苦しんでいるといわれている。ラクトース不耐性の治療のために数種のラクターゼ製品が市販されており、それらの中には例えば、ラクタイド(Lactaid)(ジョンソン&ジョンソン(Johnson & Johnson)から発売)、デイリーイーズ(Dairy Ease)(スターリングウインスロップ(Sterling Winthrop,Inc.)から発売)、およびラクトゲスト(Lactogest)(トンプソンメディカル(Thompson Medical)から発売)などがある。これらのラクターゼ製品は、2種類のうちの一方に分類されるのが一般的である:錠剤またはカプレットの形でヒトが経口摂取できるような最適pHの低い(2.5〜3.5)ラクターゼ;牛乳、チーズ、アイスクリームおよびその他ラクトースやホエイを含有する食品への食品添加物として使用する。最適pHが中性(6.5〜7.0)のラクターゼである。一般的には、最適pHの低いラクターゼは麹ラクターゼ法により、菌体内で産生される。この方法は、ロール(Roehr),M.、クビセック(Kubicek), C.P.、コミネック(Kominek), J.、(1992年)(ベネット(Bennett), J.W.およびクリック(Klich), M.A.編)pp.116-119;バターワース−ハイネマン(Butterworth-Heinemann)、ストーンハム(Stoneham), M.A.およびロックウッド(Lockwood), L.B.(1979年)微生物テクノロジー(Microbiral Technology), 第1巻第2版(ペプラー(Peppler), H.J.編)pp.356-387、アカデミックプレス(Academic Press)社刊、ニューヨーク(New York)に記載されているクエン酸の生産に用いられている麹法と類似しているものと考えられている。中性pHのラクターゼは、ヨシダ(Yoshida), H.、アラキ(Araki), K.、カワイ(Kawai), M.、Agric. Biol. Chem. 52: 951-955に記載されているように、クルイヴェロミセス ラクティス((Kluyveromyces lactis)のような酵母を用いて生産されるのが一般的である。
現在の生産方法は、工業衛生あるいは規制/安全性に対する配慮と関連しており、および/または生産コストがかかりすぎているため、ラクターゼを安全かつ低コストで生産するシステムを開発する必要がある。従って、本発明の目的は、ラクトース不耐性に苦しむ人々に投与することができるラクターゼの産生増加の方法を提供することである。本発明のラクターゼは、ラクトース不耐性に苦しむほ乳類に対して既知の任意の方法、例えば、錠剤、ドロップ、該酵素を実際に含有する牛乳または乳製品として投与することができる。
発明の概要
従って、本発明の目的は、ラクターゼの産生を増加させる方法を提供することにある。すなわち、本発明の一つの実施態様は、アスペルギルス株(好ましくはアスペルギルス・オリゼ)においてラクターゼの産生を増加させる新規な方法からなる。本発明の方法は、アスペルギルスをDNAを用いて形質転換することにより行われ、ここで、このDNAには、ラクターゼ遺伝子(好ましくはアスペルギルス ラクターゼ、最も好ましくはアスペルギルス・オリゼ ラクターゼ)をコードしているDNA配列、および、そのようなアスペルギルス株内の分泌系で機能し、かつ、ラクターゼ遺伝子をコードしているDNA配列に作用的に連結しているシグナル配列をコードしているDNAが含まれる。そのようなシグナル配列は、ラクターゼに関連したシグナル配列であるかもしくはその他に由来するものである。
本発明においては、糸状菌内で発現可能な任意のラクターゼ遺伝子を用いることができる。ラクトバシルス・ブルガリクス(Lactobacillus bulgaricus)由来のラクターゼ遺伝子はクローニングされ、大腸菌(E. coli)内で発現している(シュミット(Schumidt), B.F.ら、Journal of Bacteriology, 1989年2月号, p.625-635)。アスペルギルス・ニガー(A. niger)由来のラクターゼ遺伝子およびcDNA配列は既知である(クマー(Kumar), V.ら、Bio/Technology 10:82-85, 1992年)。アスペルギルス・オリゼのラクターゼ遺伝子は出願人らによってクローニングされ、部分的に配列が解析されており、アスペルギルス・オリゼ内での発現については本明細書に詳述されている。該遺伝子のC末端部分は現在解析中である。かくして、本発明の別の態様は、アスペルギルス・オリゼ由来のラクターゼ遺伝子のすべてまたはほぼすべてをコードしているDNA断片(遺伝子)(配列番号1)、および該遺伝子によりコードされ、ラクトース加水分解活性を有するラクターゼタンパク質のアミノ酸配列(配列番号 2)に関する。ラクターゼ遺伝子全体が解析されているわけではないが、当業者であれば、本発明の以下の教示に従えば全遺伝子のDNA配列を確認することができ、該遺伝子によって発現したタンパク質のアミノ酸配列を推定することができる。それゆえ、本発明は該遺伝子配列の全体または一部を包含するものである。
本発明の方法の別の態様においては、アスペルギルス宿主細胞を、プロモーター配列、すなわち、アスペルギルス宿主細胞により機能的に認識され、かつ、シグナル配列をコードしているDNA配列に作用的に連結している配列をコードしているDNA配列により形質転換する。さらに、宿主細胞内で発現可能な選択マーカーをコードしているDNA配列を用いて該宿主細胞を形質転換する。例えば、アスペルギルス・オリゼ由来のpyrG DNA配列はアスペルギルス・オリゼ内で発現可能である(デリオター−ヤコブス(deRioter-Jacobs), Y.M.J.Tら, Curr. Genet.(1989)16:159-163)。
本発明のひとつの実施態様においては、ラクターゼ産物が発現し、その後、アスペルギルス宿主細胞から分泌されるように、必要なDNAを用いてアスペルギルス・オリゼ株を形質転換する。
【図面の簡単な説明】
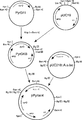
図1はCCC161からCCC28(CB587972としてオランダに寄託されている)に至る突然変異過程を模式的に示したものである。
図2は、ラクターゼ遺伝子を含むアスペルギルス・オリゼの染色体DNA由来の9kBのBglII断片を示すサザンハイブリダイゼーションのゲルである。
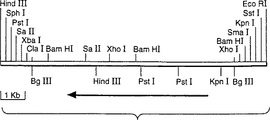
図3はアスペルギルス・オリゼのラクターゼ断片の制限地図を示している。
図4はアスペルギルス・オリゼのラクターゼ遺伝子の部分配列を示している(配列番号 1)。この図では、該遺伝子配列内に8個のイントロンが存在していることが示されている。ラクターゼ分泌のためのシグナル配列は314番から370番のコドンである(配列番号 4)。シグナル配列のアミノ酸配列は配列番号 3に示されている。
図5は、4kBのBamHI染色体断片からのアスペルギルス・オリゼ由来のpyrG遺伝子の存在を示すサザンハイブリダイゼーションのゲルである。
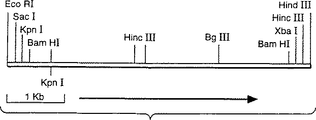
図6はアスペルギルス・オリゼpyrG断片の制限地図を示している。
図7は、pyrG11プラスミドおよびpUC218::A.o.lacプラスミドを用いたプラスミドpPyrLac4の形成を示している。
図8は、KpnIを有するpPyrLac4プラスミドが、アスペルギルス・オリゼ由来のDNAのみからなり、かつpyrGとラクターゼ遺伝子とを含有する12kBの断片をどのようにして放出するかを示している。
発明の詳細な説明
本発明は、糸状菌、好ましくはアスペルギルス・オリゼからのラクターゼの産生を増加させることに関する。特に、本発明は、アスペルギルス・オリゼ宿主株内においてアスペルギルス・オリゼ ラクターゼの産生を増加させることに関する。さらに特定すると、本明細書に記載されている方法を実施することにより、分泌ラクターゼ量は約10倍になる。ラクターゼ遺伝子の発現の制御についてはほとんどわかっていないため、この結果は驚くべきことである。というのも、遺伝子の複数(1以上)のコピーが挿入されることによりラクターゼの過剰産生を招き、特に10倍ものラクターゼの産生増は期待されていなかったのである。
そのようなラクターゼの産生増加は、ラクターゼ遺伝子の1以上のコピー(複数のコピー)が宿主株に形質転換されたために生じたものと考えられる。ラクターゼ遺伝子の複数のコピーは、ラクターゼ遺伝子および他の制御DNAをコードしているDNA配列を含むプラスミドまたはベクターにより導入され、あるいは本明細書において例示されているように、DNAの線状断片上に存在させることができる。ラクターゼ遺伝子のそのような複数のコピーによりアスペルギルス宿主が形質転換されると、該遺伝子は宿主細胞株の染色体に組み込まれるか、または染色体外に導入される。
本発明の教示に従えば、生物学的に活性なラクターゼはある種の糸状菌、好ましくはアスペルギルス、最も好ましくはアスペルギルス・オリゼ内で発現し、分泌させることができる。本明細書で用いている生物学的に活性なラクターゼとは、生化学的活性に関して天然に存在するものと同等の能力を示す活性型として分泌されるラクターゼを意味している。
一般的には、ラクターゼをコードしている配列に作用的に連結しているシグナル配列をコードしているDNAを含むDNA断片を用いる。図4(配列番号 1)はコドン番号314番から370番のシグナル配列を含んでいる。分泌シグナルに対するDNA配列を配列番号 4に示す。成熟タンパク質はコドン番号371番からはじまるDNA配列によってコードされている。
本発明のDNA断片はさらに、選択マーカーおよび/または機能性プロモーター、ならびにターミネーター配列を有している。このように構築したDNA断片、または該DNAを保有するプラスミドを用いて、アスペルギルス・オリゼを形質転換することができる。その後、ラクターゼの発現および分泌について、また別の方法としては発現可能な選択マーカーを用いてスクリーニングすることにより、活性な組換え体を確認することができる。好ましい選択マーカーの例としては、種々の抗生物質(例えば、アミノグリコシド系、フレオマイシン、ベノミルなど)に対して耐性を有し、アスペルギルス・オリゼ由来のpyrGをコードしているか、または、アスペルギルス属由来のpyrA、argB、trpCもしくはamdS、アカパンカビ(Neurospora crassa)由来のpyr4などをコードしている配列を含む;これらのおよび他の多くの選択マーカーについては当業者において既知である。
形質転換は、遺伝材料を宿主微生物に導入するための既知の方法である。すなわち、形質転換とは、DNAを保持した状態で生物内にDNAを導入することを意味しており、これは、染色体外因子としてまたは染色体に組み込まれた形のいずれであってもよい。本発明の実施例において使用した形質転換法は実質的にはキャンベル(Campbell), E.I.らによって報告されているものである(Curr. Genet. (1989)16:53-56)。
本発明において、方法に関する好ましい実施態様としては、宿主株には異種DNAを導入しないことである。商業的生産用に選ばれた株に、異種のまたは外来性のDNA配列を挿入すると、同種のDNAのみを挿入した場合よりも承認前に広範な試験を監督機関から要求される。同種DNAとは、宿主株(好ましくはアスペルギルス・オリゼ)由来のDNA、または宿主株(すなわちアスペルギルス・オリゼ)のDNA配列と一致するように合成され、かつ、50塩基対未満の連続した合成DNA、好ましくは25塩基対未満の連続した合成DNAを有しているものである。現在の指針によると、「25塩基対未満の完全に配列分析されたDNA配列を挿入する場合は宿主ベクター系を変性したとはみなさない」となっている。(アメリカ合衆国健康・教育・福祉省(U.S. Department of Health, Education and Welfare)、公衆衛生局(Public Health Service)、国立衛生研究所(National Institute of Health)「公認宿主−ベクター系の変性(Modification of Certified Host-Vector Ststems), Recombinant-DNA Technical Bulletin 2 (3):132, 1979)
本発明の宿主株は任意の糸状菌であり、好ましくはアスペルギルス・オリゼである。糸状菌は真核微生物であり、真菌亜門のすべての糸状体を含む(アレクソポウロス(Alexopoulos), C.J.、1962年、菌類学入門(Introductory Mycology)、ジョン ウィリー&サンズ(John Wiley & Sons)社刊、ニューヨーク(New York))。発現宿主(宿主株)としては、以下の属に含まれる種々の糸状菌を用いることができる:アスペルギルス属、トリコデルマ(Tricoderma)属、ニューロスポラ(Neurospora)属、ポドスポラ(Podospora)属、ムコール(Mucor)属、、アキヤ(Achlya)属、シゾフィルム(Schizophylum)属、ウスティラゴ(Ustilago)属およびコプリヌス(Coprinus)属。特定の発現宿主としては、アスペルギルス・ニデュランス(A. nidulans)、アスペルギルス・ニガー、アスペルギルス・アワモリ(A. awamori)、アスペルギルス・オリゼ、アスペルギルス・クラッサ(A. crassa)、トリコデルマ・レーセイ(T. reesei)(ロンゲブラキアトゥム(longebrachiatum))およびトリコデルマ・ヴィリデ(T. viride)が挙げられる。望ましい宿主株については米国特許出願第07/413,010号、および第07/770,049号に記載されており、これらを参照として本明細書に取り入れる。
ここで使用しているように、プロモーターまたはプロモーター配列とは、発現目的のために特定の糸状菌宿主株によって認識されるDNA配列(例えば、アスペルギルス・オリゼによって認識されるDNA配列)である。プロモーター配列は、所望するラクターゼ産物をコードしているDNA配列に作用的に連結している。そのような連結により、形質変換ベクターのシグナル配列をコードしているDNA配列の開始コドンに対してプロモーターは然るべき位置を占める。プロモーター配列は、シグナル配列および所望するラクターゼ産物の発現に関与している転写および翻訳制御配列を含む。該プロモーターの例としては、アスペルギルス・ニガーのグルコアミラーゼ由来のプロモーター、アスペルギルス・ニガーのアミラーゼプロモーター、およびその他当業者に知られているプロモーターが挙げられる。好ましいプロモーターは米国特許出願番号第07/413,010号に記載されているものであり、ここで参照として挙げておく。
シグナル配列はアミノ酸配列であり、これは、ラクターゼのアミノ末端に作用的に連結された場合に宿主株からのラクターゼの分泌を促すものである。そのようなシグナル配列は、ラクターゼに関連したシグナル配列(すなわち、生来のシグナル配列)であるか、または他を起源とするもの(すなわち、外来性のシグナル配列)である。シグナル配列は、生来のシグナル配列を介して、または、外来性のシグナル配列をコードしているDNAを正しい読み枠内にラクターゼをコードしているDNAに結合することを介して、ラクターゼに作用的に連結し、シグナル配列およびラクターゼ産物の翻訳を行う。本発明で使用する好ましいシグナル配列は、アスペルギルス・オリゼ ラクターゼ由来の生来のシグナル配列であり、これは配列番号 4に示されている。該シグナル(または分泌)配列のアミノ酸配列は配列番号 3に示されている。
本明細書において開示されている好ましい実施態様は例示であり、本発明の範囲を限定するものではない。
実施例
実施例1
アスペルギルス・オリゼ株の確立
ラクターゼ産生に使用した株は、オランダの細菌培養中央研究所(Centraal Bureau van Schimmelculture)を入手源とするCBS87972の誘導体である。これはCCC28と改名され、約5U/mlのラクターゼを産生する(1Uとは、FCC標準ラクターゼ活性分析に記載されている条件下において、37℃でONPGから1分間に1μmolのニトロフェノールを放出する酵素量である)。図1に概略を示しているように、CCC28をUVによる突然変異にかけ、選択を行い、CCC161株を得た(ATCC#74285としてメリーランド州ロックヴィルにあるアメリカンタイプカルチャーコレクション(American Type Culture Collection)に寄託している)。
培地
−継代培養はポテトデキストロース寒天(PDA)(ディフコ(DIFCO)社製)上で行った
−コロニーを単離するためには、対数増殖を防ぐ目的でPDAにデオキシコリン酸ナトリウム(180mg/ml)を添加して培養を行った(PDA Nadx)。
固体の産生培地
・小麦ふすま 50g
・寒天(ディフコ(Difco)社製) 17g
・水道水1:1
液体の産生培地
・小麦ふすま 50g
・水道水1:1
発芽用培地
・モルト抽出物(ディフコ(Difco)社製) 30g
・蒸留水1:1
溶液A:UV突然変異を行うための分生子懸濁液を調製するためのものであり、液体培地に接種したもの
・トゥイーン80(プロラボ(Prolabo)社製) 1ml
・グリセロール(プロラボ(Prolabo)社製) 150ml
・蒸留水 1:1
溶液B:NTG突然変異を行うための分生子懸濁液
・トゥイーン80 1ml
・モルト抽出物 30g
・蒸留水1:1
溶液C:NTG突然変異用リンス液
・モルト抽出物 30mg
・グリセロール 150ml
・蒸留水 1:1
溶液D:NTG突然変異用処置液
・トゥイーン80 1ml
・TAPS(シグマ(Sigma)社製) 0.1M
・蒸留水 1:1
・pHを9に調整
すべての培地および懸濁液は、グリセロールを含有するものを除き、120℃で20分滅菌し、グリセロールを含有するものは110℃で30分滅菌した。
UV突然変異
溶液Aの分生子懸濁液は、約1〜2週間経過して十分に芽胞形成しているPDAプレートから調製した。菌糸体断片を除去するためにデカンテーションを行った後、上清を採取した:4mlの該分生子懸濁液を小さいペトリ皿に移した。
この懸濁液を照射に用いた。ペトリ皿はフィリップス(Philips)社の殺菌灯(6W)の5cm下方に蓋を開けて置いた。照射時間は生存率が約1%となるように設定した。照射の間、該分生子懸濁液は均一になるように揺り動かした。
照射後、照射を受けた分生子懸濁液のサンプルを適切に希釈して培地上に塗布し、単離コロニーを得て生存率を確認した。残りの照射分生子は-20℃の冷凍庫内に保存し、さらに分析を行った。照射後は、光復帰系を阻害するため、全ての実験を暗黒下で行った。
この結果、CCC159株が得られ、この株は固体培地上では最高18U/ml、液体培地内では3U/mlのラクターゼを産生した。続いてCCC159をNTG突然変異過程に供し、選択を行い、CCC161株を得たが、この株は液体培地内で最高50U/mlのラクターゼを産生することができた。NTG突然変異は以下のように実施した:
NTG突然変異
分生子懸濁液はUV突然変異用の溶液B(上記)の場合と同様に調製した。この懸濁液5mlを20mlの発芽培地を含むエーレンマイヤーフラスコに入れ、ロータリーシェーカー上で30℃、3時間インキュベートした。
分生子はジョウアン(JOUAN)ベンチ式遠心分離機C400を用いて4500rpmで15分間遠心分離することによりハーベストした。分生子を25mlの処置溶液D(上記)に再懸濁した。次に、8mlの分生子懸濁液を適量のNTG結晶を含むエーレンマイヤーフラスコに入れた。各々の実験において、2つの異なる量のNTGについて試験を行い、NTGを含まないものを対照とした。エーレンマイヤーフラスコはロータリーシェーカー上で1時間インキュベートした。次に、分生子を遠心分離により2回洗浄し、8mlのリンス溶液C(上記)に再懸濁した。
最終懸濁液の一部をUV照射における場合と同様に生存率の確認に使用した。残りは-20℃の冷凍庫内に保存し、さらに分析を行った。処理を行った分生子は適切な希釈を行ってPDA Nadx上に塗布した、30℃で4日間インキュベートした後、コロニーを数えて生存率を確認した。生存率を確認した後、適切な量の処理分生子を希釈してPDA Nadx上に塗布し、ラクターゼ活性を調べるのに十分な量の生存細胞を得た。
突然変異を行った後、変異種から親種を分離するにはサブクローニング工程を経ることが必要である。このため、「変異」分生子懸濁液を適切に希釈して塗布し、単離コロニーを得るが、このとき、確実に一つのコロニーが一つの分生子から形成されるようにする。各々の変異に対して、100個のサブクローンについて固体倍地上でラクターゼ活性を調べた。最も良いクローンをPDA培地上に再び取り出し、液体培地で試験を行った。
実施例2
アスペルギルス・オリゼ ラクターゼのクローニング
アスペルギルス・ニガー由来のラクターゼのcDNAは既知であり、報告されている(クマー(Kumer), V.ら、Bio/Technology 10:82-85,1992)。転写、翻訳および分泌に必要な配列を有するラクターゼ遺伝子を保持するのに十分な大きさのアスペルギルス・オリゼCCC161の染色体のDNAの断片を同定するために、アスペルギルス・ニガーのDNAを鋳型として用い、アスペルギルス・ニガーの配列に基くPCRにより、プローブを作成した。これらのプローブを用いることにより、ラクターゼ遺伝子を含むアスペルギルス・オリゼの染色体DNA由来の9kbのBglII断片をサザンハイブリダイゼーションにより同定した(図2)。このサイズのクラスを含み、BglIIで切断された染色体DNAのサブゲノムライブラリーを、pUC218内で構築し、上述のプローブを用いて、ラクターゼ遺伝子を含む個々のクローン(pUC218::A.o.lac)をコロニーハイブリダイゼーションにより同定した。該断片の制限地図が得られ(図3)、遺伝子の配列は解析中である。該遺伝子の部分配列を図4(配列番号 1)に示すが、ここには該遺伝子に存在するイントロンが含まれている。アスペルギルス・オリゼのラクターゼ遺伝子により発現されたタンパク質の部分推定アミノ酸配列は、配列番号 2として示している。
実施例3
アスペルギルス・オリゼpyrGのクローニング
アスペルギルス・オリゼのpyrG遺伝子は、4kbのBamHI染色体断片上に存在することが知られている(デリオター−ヤコブス(deRioter-Jacobs),Y.M.J.Tら、 Curr. Genet. 16:159-163, 1989)。このことは、アスペルギルス・ニガーのDNAを鋳型として用い、アスペルギルス・ニガーの配列に基づくPCRにより作成したプローブを使用して、BamHIで消化したアスペルギルス・オリゼの染色体DNAにサザンハイブリダイゼーションを行うことにより確認した(図5)。このサイズのクラスを含み、BglIIで切断された染色体DNAのサブゲノムライブラリーをpUC218内で構築し、上述のプローブを用いて、pyrG遺伝子を含む個々のクローン(pyrG11)をコロニーハイブリダイゼーションにより同定した。断片の制限地図はそれが本物であることを示している(図6)。
実施例4
pPyrLac4の構築
上述の2つのプラスミド(pyrG11および pUC218::A.o.lac)を用いて、以下の方法によりプラスミドpPyrLac4を構築した(図7):
(1)pyrG11をKpbIおよびBamHIで切断し、pyrGを含む断片を単離し、pUC19を切断したKpnIおよびBamHI部位にクローニングすることによりプラスミドpyrGKBを得た。このプラスミドはアスペルギルス・オリゼpyrG断片内にあるが、pyrG遺伝子の外側に存在する唯一のBglII部位を有する。
(2)pUC218::A.o.lacをBglIIで消化し、ラクターゼ遺伝子を含むアスペルギルス・オリゼの断片を単離し、pyrKGLKBの唯一のBglII部位にクローニングすることにより、プラスミドpPyrLac4を得た。続いてpPyrLac4をKpnIで切断し、アスペルギルス・オリゼDNAのみからなり、pyrGとラクターゼ遺伝子とを含む12kbの断片を切り出した(図8)。この断片をアスペルギルス・オリゼのCCC161pyr6の形質転換に使用し、ラクターゼの産生を増加させた。
実施例5
アスペルギルス・オリゼCCC161の形質転換
本発明者らの産生株の形質転換に上述のDNA断片を使用するため、CCC161のpyrG変異体を調製した。pyrG変異体はマターン(Mattern, I.E.)らにより記載された方法(Mol. Gen. Genet. 210:460-461, 1987)を一部変更し、5−フッ化オロト酸(5−FOA)に耐性の変異体を選択し、続いて増殖にウリジンを必要とするコロニーをスクリーニングすることにより調製した。そのような変異体のうちのひとつであるCCC161pry6を次の段階に用いた。
キャンベル(Campbell, E.I.)らの方法(Curr. Genet. 16:53-56, 1989)を一部変更し、上述のKpnI断片を用いてCCC161pyr6を形質転換した。PyrG遺伝子を取り込み、増殖にもはやウリジンを必要としない形質転換体について、ラクターゼ産生能を調べた。ラクターゼ産生の増加をスクリーニングするにあたっては、X−gal(5−ブロモ−4−クロロ−3−インドール−β−D−ガラクトシド)指示プレート−これは大腸菌(E.Coli)におけるβ−ガラクトシダーゼ活性の確認に使用する標準法であり、以下および「高等細菌遺伝学(Advanced Bacterial Genetics)」(デーヴィス(Davis), R.W.、ボトステイン(Botstein), D.、およびロス(Roth), J.R.編、コールドスプリングハーバー研究所(Cold Spring Harbor Laboratory)刊、p.48、1980年)に記載されている−の培地を変更して用いた。

配列リスト
(1)一般情報
(i)出願人:ジェネンコア インターナショナル(Genencor International, Inc.)
(ii)発明の名称:アスペルギルス・オリゼにおける β−ガラクトシダーゼの産生増加
(iii)配列の数:4
(vi)問い合わせ先:
(A)名宛て人:ジェネンコア インターナショナル(Genencor International, Inc.)
(B)通り名:180 キンボール通り(Kimball Way)
(C)市名:南サンフランシスコ(South San Francisco)
(D)州名:カリフォルニア
(E)国名:アメリカ合衆国
(F)郵便番号:94080
(v)コンピュータによる読みとり可能な形式:
(A)媒体の形式:フロッピーディスク
(B)コンピューター:IBM PC コンパチブル
(C)操作システム:PC−DOS/MS−DOS
(D)ソフトウェア:パテントイン リリース(PatentIn Release)#1.0、バージョン#1.25
(vi)出願データ:
(A)出願番号:
(B)出願日:
(C)分類:
(viii)代理人情報:
(A)氏名:ホーン、マーガレット A(Horn, Mergaret)
(B)登録番号:33,401
(C)識別番号:GC250−PCT
(ix)電信情報:
(A)電話:(415) 742−7536
(B)テレファクス:(415) 742−7217
(2)配列番号 1に関する情報:
(i)配列の特徴:
(A)長さ:3515塩基対
(B)種類:核酸
(C)鎖:1本
(D)トポロジー:直線状
(ii)分子型:DNA(ゲノム)
(xi)配列の記載:配列番号 1:
(i)配列の特徴:
(A)長さ:911アミノ酸
(B)種類:アミノ酸
(C)鎖:1本
(D)トポロジー:直線状
(ii)分子型:タンパク質
(xi)配列の記載:配列番号 2:
(i)配列の特徴:
(A)長さ:19アミノ酸
(B)種類:アミノ酸
(C)鎖:1本
(D)トポロジー:直線状
(ii)分子型:タンパク質
(xi)配列の記載:配列番号 3:
(i)配列の特徴:
(A)長さ:57塩基対
(B)種類:核酸
(C)鎖:1本
(D)トポロジー:直線状
(ii)分子型:DNA(ゲノム)
(xi)配列の記載:配列番号 4:
Claims (6)
- 配列番号1の配列を含むアスペルギルス・オリゼから単離されたラクトース加水分解活性を有するタンパク質をコードするDNA断片を含む発現ベクターで、アスペルギルス・オリゼ宿主細胞(ATCC#74285)を形質転換して得られた細胞。
- アスペルギルス・オリゼ宿主細胞におけるラクターゼ産生を増加させる方法であって:
a)アスペルギルス・オリゼ ラクターゼをコードするDNA配列および該アスペルギルス・オリゼ ラクターゼをコードするDNA配列に作用的に連結している生来のアスペルギルス・オリゼ ラクターゼのシグナル配列をコードするDNA配列を含む配列番号1のDNA配列を含むDNAを用いて宿主細胞を形質転換し;
b)前記DNAを有する形質転換体を形成するのに適切な条件下で前記工程a)から得られた形質転換細胞を培養し;さらに、
c)ラクターゼ産生の増加に関して形質転換体をスクリーニングする;各工程を含み、
前記アスペルギルス・オリゼ宿主細胞がアスペルギルス・オリゼ細胞(ATCC#74285)であることを特徴とする方法。 - アスペルギルス・オリゼ ラクターゼをコードするDNAの1以上のコピーを用いて前記宿主細胞を形質転換することを特徴とする請求項2記載の方法。
- 前記宿主細胞を形質転換するのに用いるDNAが、アスペルギルス内で発現可能な選択マーカーをコードするDNA配列をさらに含むことを特徴とする請求項3記載の方法。
- 選択マーカーがアスペルギルス・オリゼpyrG遺伝子であることを特徴とする請求項4記載の方法。
- 配列番号1の配列から成るアスペルギルス・オリゼ ラクターゼおよびアスペルギルス・オリゼ シグナル配列をコードするDNA、ならびにアスペルギルス・オリゼの選択マーカーをコードするDNAを用いてアスペルギルス・オリゼ宿主細胞を形質転換することを特徴とする請求項2記載の方法。
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| US26763194A | 1994-06-29 | 1994-06-29 | |
| US08/267,631 | 1994-06-29 | ||
| PCT/US1995/008013 WO1996000786A1 (en) | 1994-06-29 | 1995-06-23 | INCREASED PRODUCTION OF β-GALACTOSIDASE IN ASPERGILLUS ORYZAE |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| JPH10504449A JPH10504449A (ja) | 1998-05-06 |
| JP3736574B2 true JP3736574B2 (ja) | 2006-01-18 |
Family
ID=23019572
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| JP50336496A Expired - Lifetime JP3736574B2 (ja) | 1994-06-29 | 1995-06-23 | アスペルギルス オリゼ(Aspergillus oryzae)におけるβ−ガラクトシダーゼの生産増加 |
Country Status (10)
| Country | Link |
|---|---|
| US (1) | US5736374A (ja) |
| EP (1) | EP0767837B1 (ja) |
| JP (1) | JP3736574B2 (ja) |
| AT (1) | ATE254176T1 (ja) |
| CA (1) | CA2193965C (ja) |
| DE (1) | DE69532122T2 (ja) |
| DK (1) | DK0767837T3 (ja) |
| ES (1) | ES2210297T3 (ja) |
| MX (1) | MX9606610A (ja) |
| WO (1) | WO1996000786A1 (ja) |
Families Citing this family (14)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US6833260B1 (en) | 1999-10-08 | 2004-12-21 | Protein Scientific, Inc. | Lactose hydrolysis |
| AU1996101A (en) * | 1999-12-30 | 2001-07-16 | Novozymes A/S | Fungal extracellular fam35 beta-galactosidases |
| ES2454167T3 (es) | 2005-11-28 | 2014-04-09 | Dsm Ip Assets B.V. | Preparación de enzima que produce un sabor puro |
| ES2591359T3 (es) | 2007-12-03 | 2016-11-28 | Novozymes A/S | Método para producir un producto lácteo de bajo contenido en lactosa |
| CN103881994B (zh) * | 2014-04-14 | 2020-03-27 | 中国农业科学院生物技术研究所 | 一种具有高转糖苷活性的β-半乳糖苷酶突变体及其制备方法和应用 |
| DK3228194T3 (da) | 2016-04-07 | 2021-06-07 | Dsm Ip Assets Bv | Fermenteret mælkeprodukt med diacetyl dannet ved hjælp af lactase |
| WO2018187524A1 (en) * | 2017-04-07 | 2018-10-11 | Dupont Nutrition Biosciences Aps | BACILLUS HOST CELLS PRODUCING β-GALACTOSIDASES AND LACTASES IN THE ABSENCE OF P-NITROBENZYLESTERASE SIDE ACTIVITY |
| CN107365713A (zh) * | 2017-08-23 | 2017-11-21 | 齐鲁工业大学 | 一种用于高产乳糖酶的米曲霉原生质体的制备与再生方法 |
| JP7706443B2 (ja) | 2020-04-17 | 2025-07-11 | 合同酒精株式会社 | 容器詰めラクターゼ溶液 |
| EP4000417A1 (en) | 2020-11-23 | 2022-05-25 | DSM IP Assets B.V. | Combination of lactase and a yeast cell wall derived taste modulator |
| WO2022223430A1 (en) | 2021-04-19 | 2022-10-27 | Dsm Ip Assets B.V. | A composition of enzymes and human milk oligosaccharides |
| JPWO2023106420A1 (ja) | 2021-12-09 | 2023-06-15 | ||
| CN116179368B (zh) * | 2023-02-10 | 2024-04-19 | 宁波希诺亚海洋生物科技有限公司 | 一株产酸性乳糖酶的米曲霉及其应用 |
| CN120005857B (zh) * | 2023-11-07 | 2025-10-17 | 中国农业大学 | 米曲霉β-半乳糖苷酶突变体的高效表达及在乳品中的应用 |
Family Cites Families (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3919049A (en) * | 1974-06-27 | 1975-11-11 | Tokyo Tanabe Co | Process for preparing {62 -galactosidase |
| US4885249A (en) * | 1984-12-05 | 1989-12-05 | Allelix, Inc. | Aspergillus niger transformation system |
| JPS63301794A (ja) * | 1987-05-30 | 1988-12-08 | Wakamoto Pharmaceut Co Ltd | 乳糖分解方法 |
| GB8905674D0 (en) * | 1989-03-13 | 1989-04-26 | Imperial College | Dna construct and modified yeast |
-
1995
- 1995-06-23 AT AT95924068T patent/ATE254176T1/de not_active IP Right Cessation
- 1995-06-23 ES ES95924068T patent/ES2210297T3/es not_active Expired - Lifetime
- 1995-06-23 JP JP50336496A patent/JP3736574B2/ja not_active Expired - Lifetime
- 1995-06-23 WO PCT/US1995/008013 patent/WO1996000786A1/en not_active Ceased
- 1995-06-23 MX MX9606610A patent/MX9606610A/es unknown
- 1995-06-23 EP EP95924068A patent/EP0767837B1/en not_active Expired - Lifetime
- 1995-06-23 DE DE69532122T patent/DE69532122T2/de not_active Expired - Lifetime
- 1995-06-23 DK DK95924068T patent/DK0767837T3/da active
- 1995-06-23 CA CA002193965A patent/CA2193965C/en not_active Expired - Lifetime
-
1996
- 1996-02-05 US US08/596,985 patent/US5736374A/en not_active Expired - Lifetime
Also Published As
| Publication number | Publication date |
|---|---|
| MX9606610A (es) | 1997-03-29 |
| EP0767837A1 (en) | 1997-04-16 |
| CA2193965C (en) | 2007-11-13 |
| ES2210297T3 (es) | 2004-07-01 |
| DE69532122D1 (de) | 2003-12-18 |
| DE69532122T2 (de) | 2004-07-08 |
| CA2193965A1 (en) | 1996-01-11 |
| EP0767837B1 (en) | 2003-11-12 |
| WO1996000786A1 (en) | 1996-01-11 |
| ATE254176T1 (de) | 2003-11-15 |
| DK0767837T3 (da) | 2004-03-15 |
| JPH10504449A (ja) | 1998-05-06 |
| US5736374A (en) | 1998-04-07 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP3736574B2 (ja) | アスペルギルス オリゼ(Aspergillus oryzae)におけるβ−ガラクトシダーゼの生産増加 | |
| Daba et al. | The ancient koji mold (Aspergillus oryzae) as a modern biotechnological tool | |
| CA1333777C (en) | Aspartic proteinase deficient filamentous fungi | |
| TW526268B (en) | Method for producing a fermented milk product | |
| JP4922524B2 (ja) | 糸状菌の分野における新規発現調節配列および発現産物 | |
| EP1334182B2 (en) | Method of producing non-bovine chymosin and use hereof | |
| CN1219971A (zh) | 丝状真菌的形态突变体 | |
| US8900647B2 (en) | Koji mold having large-scale genomic duplication | |
| EP2408910A2 (en) | Chrysosporium lucknowense protein production system | |
| JP2022110110A (ja) | 糸状真菌宿主細胞によって発現された組換えシュウ酸デカルボキシラーゼ | |
| CN109988714B (zh) | 一种里氏木霉及其应用 | |
| CN1108302A (zh) | 保加利亚乳杆菌,和含有该乳杆菌的食品组合物 | |
| JPH07123977A (ja) | 乳酸菌及び発酵乳製品 | |
| JP6335161B2 (ja) | 細胞表層発現用ポリヌクレオチド | |
| KR20170088964A (ko) | 진균 숙주 주, dna 구성물, 및 사용 방법 | |
| CN119823882B (zh) | 一株高蛋白转化率的威尼斯镰刀菌及其在提高菌丝蛋白饱腹感中的应用 | |
| JP2018516076A (ja) | Aldcの製造方法 | |
| JP2003516112A (ja) | 相同又は非相同タンパク質生産のための組換えペニシリウム フニクロスム | |
| JPH07222591A (ja) | 発現系、組込みベクター及びその組込みベクターで形質転換された細胞 | |
| Wöstemeyer et al. | Inter-mating type protoplast fusion in the zygomycete Absidia glauca | |
| EP0205506A1 (en) | METHOD FOR TRANSFORMING MUSHROOMS WITH A VECTOR. | |
| TW201002820A (en) | Method for the introduction of a heterologous polynucleotide into a mushroom | |
| JP2960775B2 (ja) | ノカルジオフォーム微生物由来のdna配列の発現 | |
| Kadiri et al. | Application of microbial enzyme in food biotechnology | |
| KR20160145411A (ko) | 바실러스 서브틸리스 1-d-5, 이를 이용한 비피도박테리아 증식방법 및 효소식품 제조방법 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A131 | Notification of reasons for refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A131 Effective date: 20041130 |
|
| A601 | Written request for extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A601 Effective date: 20050221 |
|
| A602 | Written permission of extension of time |
Free format text: JAPANESE INTERMEDIATE CODE: A602 Effective date: 20050404 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050413 |
|
| A02 | Decision of refusal |
Free format text: JAPANESE INTERMEDIATE CODE: A02 Effective date: 20050607 |
|
| A521 | Request for written amendment filed |
Free format text: JAPANESE INTERMEDIATE CODE: A523 Effective date: 20050829 |
|
| A911 | Transfer to examiner for re-examination before appeal (zenchi) |
Free format text: JAPANESE INTERMEDIATE CODE: A911 Effective date: 20050915 |
|
| TRDD | Decision of grant or rejection written | ||
| A01 | Written decision to grant a patent or to grant a registration (utility model) |
Free format text: JAPANESE INTERMEDIATE CODE: A01 Effective date: 20051004 |
|
| A61 | First payment of annual fees (during grant procedure) |
Free format text: JAPANESE INTERMEDIATE CODE: A61 Effective date: 20051018 |
|
| R150 | Certificate of patent or registration of utility model |
Free format text: JAPANESE INTERMEDIATE CODE: R150 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20091104 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20091104 Year of fee payment: 4 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20101104 Year of fee payment: 5 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20111104 Year of fee payment: 6 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121104 Year of fee payment: 7 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20121104 Year of fee payment: 7 |
|
| FPAY | Renewal fee payment (event date is renewal date of database) |
Free format text: PAYMENT UNTIL: 20131104 Year of fee payment: 8 |
|
| S111 | Request for change of ownership or part of ownership |
Free format text: JAPANESE INTERMEDIATE CODE: R313113 |
|
| S531 | Written request for registration of change of domicile |
Free format text: JAPANESE INTERMEDIATE CODE: R313531 |
|
| S533 | Written request for registration of change of name |
Free format text: JAPANESE INTERMEDIATE CODE: R313533 |
|
| R350 | Written notification of registration of transfer |
Free format text: JAPANESE INTERMEDIATE CODE: R350 |
|
| R250 | Receipt of annual fees |
Free format text: JAPANESE INTERMEDIATE CODE: R250 |
|
| EXPY | Cancellation because of completion of term |