KR20150143699A - L-아미노산을 생산하는 재조합균, 이의 구성 방법 및 l-아미노산 생산 방법 - Google Patents
L-아미노산을 생산하는 재조합균, 이의 구성 방법 및 l-아미노산 생산 방법 Download PDFInfo
- Publication number
- KR20150143699A KR20150143699A KR1020157032480A KR20157032480A KR20150143699A KR 20150143699 A KR20150143699 A KR 20150143699A KR 1020157032480 A KR1020157032480 A KR 1020157032480A KR 20157032480 A KR20157032480 A KR 20157032480A KR 20150143699 A KR20150143699 A KR 20150143699A
- Authority
- KR
- South Korea
- Prior art keywords
- gene
- promoter
- seq
- ala
- homology
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Images






Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/24—Proline; Hydroxyproline; Histidine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/0004—Oxidoreductases (1.)
- C12N9/0006—Oxidoreductases (1.) acting on CH-OH groups as donors (1.1)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/74—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora
- C12N15/77—Vectors or expression systems specially adapted for prokaryotic hosts other than E. coli, e.g. Lactobacillus, Micromonospora for Corynebacterium; for Brevibacterium
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1235—Diphosphotransferases (2.7.6)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
- C12N9/92—Glucose isomerase (5.3.1.5; 5.3.1.9; 5.3.1.18)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P13/00—Preparation of nitrogen-containing organic compounds
- C12P13/04—Alpha- or beta- amino acids
- C12P13/08—Lysine; Diaminopimelic acid; Threonine; Valine
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01043—Phosphogluconate 2-dehydrogenase (1.1.1.43)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y101/00—Oxidoreductases acting on the CH-OH group of donors (1.1)
- C12Y101/01—Oxidoreductases acting on the CH-OH group of donors (1.1) with NAD+ or NADP+ as acceptor (1.1.1)
- C12Y101/01049—Glucose-6-phosphate dehydrogenase (1.1.1.49)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Y—ENZYMES
- C12Y503/00—Intramolecular oxidoreductases (5.3)
- C12Y503/01—Intramolecular oxidoreductases (5.3) interconverting aldoses and ketoses (5.3.1)
- C12Y503/01009—Glucose-6-phosphate isomerase (5.3.1.9)
Landscapes
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biochemistry (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biomedical Technology (AREA)
- Molecular Biology (AREA)
- Medicinal Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
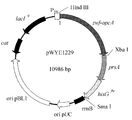
도 1은 재조합 플라스미드 pXMJ19-prsA - hisG fbr 의 도면.
도 2는 CG161 균주(pgi 유전자가 녹아웃됨) 게놈 DNA의 PCR 검증 전기 이동도.
도 3은 재조합 플라스미드 pXMJ19-zwf - opcA - prsA - hisG fbr 의 도면.
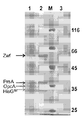
도 4는 L-히스티딘 공정균 CG171 발현 단백질의 SDS-PAGE 도면.
도 5는 L-히스티딘 공정균 CG171 중의 글루코스-6-포스페이트 데히드로게나아제 효소 활성 측정도.
도 6은 재조합 플라스미드 pXMJ19-zwf - opcA - prsA - hisG fbr - purH의 도면.
도 7은 CG328 균주가 플라스미드 DNA를 휴대한 PCR 검증 전기 이동도.
도 8은 CG353 균주(purF 유전자가 약화됨) 게놈 DNA의 PCR 검증 전기 이동도.
| 시간(min) | 유동상 A(%) | 유동상 B(%) |
| 0 | 86 | 14 |
| 2 | 88 | 12 |
| 4 | 86 | 14 |
| 10 | 70 | 30 |
| 20 | 30 | 70 |
| 21 | 10 | 90 |
| 24 | 0 | 100 |
| 균주 | 글루코스 소모량(g) | 최대 OD600 | 비성장 속도(h-1) | L-히스티딘 수율(g/L) |
| CG160 | 3.8 | 62.67 | 0.131 | 0.03 |
| CG176 | 3.8 | 60.97 | 0.127 | 1.18 |
| CG172 | 1.2 | 46.67 | 0.073 | 0.77 |
| CG173 | 3.8 | 59.07 | 0.120 | 1.50 |
| CG171 | 1.8 | 53.83 | 0.108 | 2.40 |
| 균주 | 히스티딘 수율(g/L) | 발효 시간(h) |
| CG171 | 10.87 | 52 |
| CG319 | 14.15 | 47 |
| CG328 | 15.96 | 50 |
| 균주 | 히스티딘 수율(g/L) |
| 야생형 | --- |
| CG350 | 0.65 |
| CG351 | 1.86 |
| CG352 | 2.23 |
| CG353 | 2.34 |
Claims (25)
- L-아미노산을 생산하는 재조합균에 있어서, 상기 재조합균은 시작균에 비하여 낮아진 글루코스-6-포스페이트 아이소메라아제 Pgi의 발현과 향상된 글루코스-6-포스페이트 데히드로게나아제 Zwf-OpcA의 발현을 가지며, 그 중에서 상기 시작균은 목표 아미노산을 누적시킬 수 있는 균주이며, 바람직하게는, 상기 목표 아미노산은 L-히스티딘, L-라이신, L-발린, L-트레오닌, L-프롤린 또는 L-히드록시프롤린인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제1항에 있어서, 상기 재조합균의 염색체 상의 pgi 유전자는 이미 불활성화되며 바람직하게는 이미 녹아웃되거나, 또는 pgi 유전자의 조절 요소가 이미 낮은 전사 또는 낮은 발현 활성의 조절 요소로 교체되고, 아울러 상기 재조합균 중에는 두 개 또는 더욱 많은 카피한 zwf - opcA 유전자를 가지거나, 또는 강한 프로모터로 상기 시작균 염색체 상의 tkt - tal - zwf - opcA - devB 오페론의 프로모터를 교체하는 바, 바람직하게는, 상기 강한 프로모터는 원시균의 P eftu 프로모터인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제1항에 있어서, 상기 시작균은 원시균에 비하여 L-히시티딘 합성 오페론의 hisEG 유전자와 hisDCB 유전자의 향상된 발현을 가지며, 바람직하게는, 강한 프로모터로 상기 hisEG 유전자와 hisDCB 유전자의 프로모터를 교체시키고, 더욱 바람직하게는 상기 원시균 염색체 상의 P glyA 프로모터로 각각 hisEG 유전자와 hisDCB 유전자의 프로모터를 교체시키며;
나아가 바람직하게는, 상기 시작균은 원시균에 비하여 향상된 PRPP 합성 효소 PrsA의 발현을 가지며, 더욱 바람직하게는, 상기 시작균 중에는 두 개 또는 더욱 많은 카피의 prsA 유전자를 가지거나, 또는 강한 프로모터로 prsA 유전자의 프로모터를 교체하는 바, 바람직하게는 상기 프로모터는 상기 원시균의 P sod 프로모터인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제1항에 있어서,
상기 시작균은 원시균에 비하여 향상된 dapA 유전자 또는 lysC 유전자의 발현을 가지거나; 또는
상기 시작균은 원시균에 비하여 향상된 발린 합성 유전자 ilvBNCE의 발현을 가지거나; 또는
상기 시작균은 원시균에 비하여 향상된 트레오닌 합성 유전자 hom 및 thrB의 발현을 가지거나; 또는
상기 시작균은 원시균에 비하여 향상된 ocd 유전자의 발현을 가지거나; 또는
상기 시작균은 원시균에 비하여 향상된 p4hD 유전자의 발현을 가지는; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제3항에 있어서, 상기 재조합균은 상기 시작균에 비하여 향상된 AICAR 트랜스메칠라제/IMP 환히드라제 PurH의 발현을 가지며;
바람직하게는, 상기 시작균 중에는 두 개 또는 더욱 많은 카피의 purH 유전자를 가지거나, 또는 강한 프로모터로 purH 유전자의 프로모터를 교체시키는 바, 바람직하게는, 상기 프로모터는 상기 원시균의 P eftu 프로모터인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제3항 또는 제5항에 있어서, 상기 재조합균은 상기 시작균에 비하여 약화된 아미도포스포리보실 전이효소 PurF의 발현을 가지며;
바람직하게는, 약한 프로모터로 purF 유전자의 프로모터를 교체하고, 더욱 바람직하게는, 상기 약한 프로모터는 상기 원시균 중의 P hom 프로모터인인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제1항 내지 제6항의 임의 항에 있어서, 상기 원시균은 코리네박테리움 속, 다이어리스터 속 또는 브레비박테리움 속으로부터 선택되는 한 가지 세균주이며;
바람직하게는, 상기 코리네박테리움 속의 세균은 Corynebacterium glutamicum, Corynebacterium pekinense, Corynebacterium efficiens, Corynebacterium crenatum, Corynebacterium thermoaminogenes, Corynebacterium aminogenes, Corynebacterium lilium, Corynebacterium callunae 및 Corynebacterium herculis으로부터 선택되는 한 가지 세균주이며;
상기 다이어리스터 속의 세규은 Microbacterium ammoniaphilum으로부터 선택되는 한 가지 세균주이며;
상기 브레비박테리움 속의 세균은 Brevibacteriaceae flvum, Brevibacteriaceae lactofermentum 및 Brevibacteriaceae ammoniagenes으로부터 선택되는 한 가지 세균주인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제7항에 있어서, 상기 원시균은 야생형 Corynebacterium glutamicum ATCC13032인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제8항에 있어서, 상기 시작균의 염색체 상에는 각각 상기 코리네박테리움 글루타미쿰 ATCC13032 염색체 상의 L-히스티딘 합성 오페론 hisEG와 hisDCB의 프로모터를 교체하는 서열 7 중의 5’ 말단 제863-1038번 뉴클레오티드 서열이 보여주는 P glyA 프로모터를 가지며;
상기 시작균은 돌연변이된 ATP-포스포리보실 전이효소를 발현할 수 있는 바, 상기 돌연변이된 ATP-포스포리보실 전이효소는 서열 6이 보여주는 ATP-포스포리보실 전이효소의 제215번의 아스파라진이 라이신으로 돌연변이되고, 제231번의 류신이 페닐알라닌으로 돌연변이되며, 제235번의 트레오닌이 알라닌으로 돌연변이된 효소이며, 바람직하게는, 상기 시작균의 염색체 상에는 상기 코리네박테리움 글루타미쿰 ATCC13032 염색체 상의 hisG 유전자를 교체하는 서열 4 중의 제1007-1852번 뉴클레오티드 서열이 보여주는 hisG fbr 유전자를 가지며;
상기 시작균의 염색체 상에는 상기 코리네박테리움 글루타미쿰 ATCC13032 염색체 상의 prsA 유전자의 프로모터를 교체하는 서열 11 중의 5' 말단 제656-847번 뉴클레오티드 서열이 보여주는 P sod 프로모터를 가지거나;
또는, 상기 시작균 중에는 두 개 또는 더욱 많은 카피의 prsA 유전자와 hisG fbr 유전자를 가지며;
그 중에서, 상기 prsA 유전자는 인코딩 서열 5가 보여주는 PrsA의 유전자에서 선택되며; 코드가 상기 PrsA와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 PrsA 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 prsA 유전자는 서열표 중의 서열 4가 보여주는 제15-992번 뉴클레오티드 서열인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제9항에 있어서, 상기 pgi 유전자는 인코딩 서열표 중의 서열 14가 보여주는 Pgi의 유전자에서 선택되며; 코드가 상기 Pgi와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 글루코스-6-포스페이트 아이소메라아제 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 pgi 유전자는 서열 13이 보여주는 뉴클레오티드 서열이며;
상기 zwf - opcA 유전자는 서열표 중의 인코딩 서열표 중의 서열 3이 보여주는 zwf-opcA의 유전자에서 선택되며; 코드가 상기 zwf-opcA와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 zwf-opcA 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 zwf - opcA 유전자는 서열 2가 보여주는 뉴클레오티드 서열이며;
상기 P eftu 프로모터는 서열 12가 보여주는 5’ 말단 제635-834번의 뉴클레오티드 서열인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제10항에 있어서, 상기 purH 유전자는 인코딩 서열표 중의 서열 16이 보여주는 purH의 유전자에서 선택되며; 코드가 상기 purH와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 purH 활성을 가지는 유전자 중의 하나이고, 상기 purH 유전자는 서열표 중의 서열 15가 보여주는 뉴클레오티드 서열인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- 제11항에 있어서, 상기 P hom 프로모터는 서열 18이 보여주는 5’ 말단 제736-865번의 뉴클레오티드 서열인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균.
- L-아미노산을 생산하는 재조합균의 구성 방법에 있어서, 하기 단계가 포함되는 바, 즉 시작균 중의 글루코스-6-포스페이트 아이소메라아제 Pgi의 발현을 낮추고, 또한 글루코스-6-포스페이트 데히드로게나아제 Zwf-OpcA의 발현을 향상시켜 상기 재조합균을 취득하며;
그 중에서 상기 시작균은 목표 아미노산을 누적시킬 수 있는 균주이며, 더욱 바람직하게는, 상기 목표 아미노산은 L-히스티딘, L-라이신, L-발린, L-트레오닌, L-프롤린 또는 L-히드록시프롤린인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제13항에 있어서,
상기 시작균 중의 Pgi의 발현을 낮추는 것은 하기 A) 또는 B) 방식을 통하여 구현하는 바, 즉
A) 상기 시작균 염색체 상의 pgi 유전자를 불활성화시킨 것으로서, 상기 불활성화는 녹아웃인 것이 바람직하며;
B) 상기 시작균 중의 pgi 유전자의 조절 요소를 낮은 전사와 낮은 발현 활성의 조절 요소로 교체하여 구현하며;
상기 시작균 중의 Zwf - OpcA의 발현을 향상시키는 것은 하기 C) 또는 D) 방식을 통하여 구현한다.
C) 상기 시작균 중의 zwf - opcA 유전자의 카피 수량을 증가시키며;
D) 상기 시작균 염색체 상의 tkt - tal - zwf - opcA - devB 오페론의 프로모터을 강한 프로모터로 교체시키는 바, 바람직하게는, 상기 강한 프로모터는 상기 원시균 염색체 상의 P eftu 프로모터인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제13항에 있어서, 상기 시작균을 취득하는 것은 원시균 염색체 상의 L-히스티딘 합성 오페론 hisEG와 hisDCB의 프로모터를 각각 강한 프로모터로 교체하는 단계가 포함되는 바, 바람직하게는, 상기 강한 프로모터는 상기 원시균 염색체 상의 P glyA 프로모터이며;
바람직하게는 상기 시작균을 취득하는 것에는 더우기 상기 시작균 중의 PRPP 합성 효소 PrsA의 발현을 향상시키는 단계를 포함하며;
더욱 바람작하게는, 상기 시작균 중의 PrsA의 발현을 향상시키는 것은 하기 E) 또는 F) 방식을 통하여 구현한다.
E) 상기 시작균 중의 prsA 유전자의 카피 수량을 증가시키며;
F) 상기 시작균 염색체 상의 prsA 유전자의 프로모터을 강한 프로모터로 교체시키는 바, 바람직하게는, 상기 강한 프로모터는 상기 원시균 염색체 상의 P sod 프로모터인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제13항에 있어서, 상기 시작균을 취득하는 것에는,
dapA 유전자 또는 lysC 유전자의 발현을 향상시키는 단계; 또는
발린 합성 유전자 ilvBNCE의 발현을 향상시키는 단계; 또는
트레오닌 합성 경로 유전자 hom 및 thrB의 발현을 향상시키는 단계; 또는
ocd 유전자의 발현을 향상시키는 단계; 또는
p4hD 전자의 발현을 향상시키는 단계가 포함되는; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제15항에 있어서, 상기 방법에는 나아가 상기 재조합균 중의 AICAR 트랜스메칠라제/IMP 환히드라제 PurH의 발현을 향상시키는 단계를 포함하며;
바람직하게는, 상기 재조합균 중의 PurH의 발현을 향상시키는 것은 하기 G) 또는 H) 방식을 통하여 구현하는 바, 즉
G) 상기 시작균 중의 purH 유전자의 카피 수량을 증가시키며;
F) 상기 시작균 염색체 상의 purH 유전자의 프로모터을 강한 프로모터로 교체시키는 바, 바람직하게는, 상기 강한 프로모터는 상기 원시균 염색체 상의 P eftu 프로모터인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제15항 또는 제17항에 있어서, 상기 방법에는 나아가 상기 재조합균 중의 아미도포스포리보실 전이효소 PurF의 발현을 약화시키는 단계가 포함되며;
상기 재조합균 중의 PurF의 발현을 약화시키는 것은 약한 프로모터로 purF 유전자의 프로모터를 교체시키는 방식으로 구현하며, 더욱 바람직하게는, 상기 시작균 염색체 상의 purF 유전자의 프로모터를 상기 원시균 중의 염색체 상의 P hom 프로모터로 교체시키는 것을 통하여 구현하는; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제13항 내지 제18항의 임의 항에 있어서, 상기 시작균을 취득하기 위한 상기 원시균은 코리네박테리움 속, 다이어리스터 속 또는 브레비박테리움 속으로부터 선택되는 한 가지 세균주이며;
바람직하게는, 상기 코리네박테리움 속의 세균은 Corynebacterium glutamicum, Corynebacterium pekinense, Corynebacterium efficiens, Corynebacterium crenatum, Corynebacterium thermoaminogenes, Corynebacterium aminogenes, Corynebacterium lilium, Corynebacterium callunae 및 Corynebacterium herculis으로부터 선택되는 한 가지 세균주이며;
상기 다이어리스터 속의 세규은 Microbacterium ammoniaphilum으로부터 선택되는 한 가지 세균주이며;
상기 브레비박테리움 속의 세균은 Brevibacteriaceae flvum, Brevibacteriaceae lactofermentum 및 Brevibacteriaceae ammoniagenes으로부터 선택되는 한 가지 세균주인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제19항에 있어서, 상기 원시균은 야생형 Corynebacterium glutamicum ATCC13032인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제20항에 있어서, 상기 시작균을 취득하는 것에는 하기 단계가 포함되는 바, 즉
상기 Corynebacterium glutamicum ATCC13032 염색체 상의 L-히스티딘 합성 오페론 hisEG와 hisDCB의 프로모터를 각각 서열 7 중의 5’ 말단 제863-1038번 뉴클레오티드가 보여주는 P glyA 프로모터로 교체하며;
상기 Corynebacterium glutamicum ATCC13032 이 발현하는 서열 6이 보여주는 ATP-포스포리보실 전이효소는 서열 6이 보여주는 ATP-포스포리보실 전이효소의 제215번의 아스파라진을 라이신으로 돌연변이시키고, 제231번의 류신이 페닐알라닌으로 돌연변이시키며, 제235번의 트레오닌이 알라닌으로 돌연변이시키며; 바람직하게는, 상기 돌연변이를 진행하기 위한 유전자는 서열 4 중의 제1007-1852번 뉴클레오티드 서열이 보여주는 hisG fbr 유전자이며;
바람직하게는, 상기 시작균을 취득하는 것에는 나아가 하기 단계가 포함되는 바, 즉
상기 Corynebacterium glutamicum ATCC13032 염색체 상의 prsA 프로모터를 서열 11 중의 5’ 말단 제656-847번 뉴클레오티드가 보여주는 P sod 프로모터로 교체하거나;
또는 나아가 하기 단계를 포함하는 바, 즉
상기 Corynebacterium glutamicum ATCC13032 중의 prsA 유전자의 카피 수량을 증가시키고,
상기 Corynebacterium glutamicum ATCC13032 중의 hisG fbr 유전자의 카피 수량을 증가시키며;
그 중에서, 상기 prsA 유전자는 인코딩 서열 5가 보여주는 PrsA의 유전자에서 선택되며; 코드가 상기 PrsA와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 PrsA 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 prsA 유전자는 서열표 중의 서열 4가 보여주는 제15-992번 뉴클레오티드 서열인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제21항에 있어서, 상기 pgi 유전자는 인코딩 서열표 중의 서열 14가 보여주는 Pgi의 유전자에서 선택되며; 코드가 상기 Pgi와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 또한 글루코스-6-포스페이트 아이소메라아제 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 pgi 유전자는 서열 13이 보여주는 뉴클레오티드 서열이며;
상기 zwf - opcA 유전자는 인코딩 서열표 중의 서열 3이 보여주는 zwf-opcA의 유전자에서 선택되며; 코드가 상기 zwf-opcA와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 zwf-opcA 활성을 가지는 유전자 중의 하나이고, 바람직하게는, 상기 zwf - opcA 유전자는 서열 2가 보여주는 뉴클레오티드 서열이며;
상기 P eftu 프로모터는 서열 12가 보여주는 5’ 말단 제635-834번의 뉴클레오티드 서열인; 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제22항에 있어서, 상기 purH 유전자는 인코딩 서열표 중의 서열 16이 보여주는 purH의 유전자에서 선택되며; 코드가 상기 purH와 비하여 적어도 60%의 상동성을 가지고, 바람직하게는 적어도 70%의 상동성을 가지며, 더욱 바람직하게는 적어도 80% 상동성을 가지고, 나아가 바람직하게는 적어도 95%의 상동성을 가지며, 더욱 바람직하게는 적어도 98% 내지는 99%의 상동성을 가지며 또한 purH 활성을 가지는 유전자 중의 하나이고, 상기 purH 유전자는 서열표 중의 서열 15가 보여주는 뉴클레오티드 서열인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- 제23항에 있어서, 상기 P hom 프로모터는 서열 18이 보여주는 5’ 말단 제736-865번의 뉴클레오티드 서열인 것을 특징으로 하는 L-아미노산을 생산하는 재조합균의 구성 방법.
- L-아미노산을 생산하는 방법에 있어서, 청구항 1 내지 12의 재조합균을 발효 배양하는 단계를 포함하는 바, 상기 L-아미노산은 L-히스티딘, L-라이신, L-발린, L-트레오닌, L-프롤린 또는 L-히드록시프롤린이 바람직한 것을 특징으로 하는 L-아미노산을 생산하는 방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201410050868.2A CN104845923B (zh) | 2014-02-14 | 2014-02-14 | 生产l‑组氨酸的方法及其专用重组菌 |
| CN201410050868.2 | 2014-02-14 | ||
| PCT/CN2015/072220 WO2015120775A1 (zh) | 2014-02-14 | 2015-02-04 | 一种产l-氨基酸的重组菌、其构建方法及l-氨基酸生产方法 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| KR20150143699A true KR20150143699A (ko) | 2015-12-23 |
| KR101937682B1 KR101937682B1 (ko) | 2019-01-14 |
Family
ID=53799587
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020157032480A Expired - Fee Related KR101937682B1 (ko) | 2014-02-14 | 2015-02-04 | L-아미노산을 생산하는 재조합균, 이의 구성 방법 및 l-아미노산 생산 방법 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US9796991B2 (ko) |
| EP (1) | EP3009505B1 (ko) |
| JP (1) | JP6188930B2 (ko) |
| KR (1) | KR101937682B1 (ko) |
| CN (2) | CN104845923B (ko) |
| WO (1) | WO2015120775A1 (ko) |
Cited By (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019190192A1 (ko) * | 2018-03-27 | 2019-10-03 | 씨제이제일제당 (주) | 신규한 프로모터 및 이를 이용한 l-아미노산 생산 방법 |
| KR102281360B1 (ko) * | 2021-01-26 | 2021-07-22 | 씨제이제일제당 (주) | 신규한 atp 포스포리보실트랜스퍼라제 변이체 및 이를 이용한 l-발린 생산 방법 |
| WO2022231042A1 (ko) * | 2021-04-30 | 2022-11-03 | 씨제이제일제당 (주) | 신규한 변이체 및 이를 이용한 l-발린 생산 방법 |
| WO2025159448A1 (ko) * | 2024-01-26 | 2025-07-31 | 씨제이제일제당 (주) | 신규한 리보스 포스페이트 디포스포키나제 변이체 및 이를 이용한 l-아미노산 생산 방법 |
Families Citing this family (30)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN107012161A (zh) * | 2017-04-03 | 2017-08-04 | 天津大学 | 利用秸秆水解液高产琥珀酸的谷氨酸棒杆菌及构建及应用 |
| CN107227283B (zh) * | 2017-05-26 | 2021-01-15 | 廊坊梅花生物技术开发有限公司 | 一种谷氨酸棒杆菌及其构建方法与应用 |
| KR101904666B1 (ko) | 2017-08-02 | 2018-11-29 | 씨제이제일제당 (주) | Atp 포스포리보실기 전이효소 변이체 및 이를 이용한 l-히스티딘 생산방법 |
| CN107867759A (zh) * | 2017-11-07 | 2018-04-03 | 周晓斌 | 一种新型污水处理外加营养培菌剂及其制作方法 |
| KR101904675B1 (ko) * | 2017-12-15 | 2018-10-04 | 씨제이제일제당 (주) | 5'-이노신산을 생산하는 미생물 및 이를 이용한 5'-이노신산의 생산 방법 |
| CN110117568B (zh) * | 2018-02-06 | 2021-03-30 | 中国科学院微生物研究所 | 生产l-组氨酸的重组菌、其构建方法以及l-组氨酸的生产方法 |
| CN110656074B (zh) * | 2018-06-28 | 2022-06-28 | 中国科学院青岛生物能源与过程研究所 | 一种合成次黄嘌呤的重组菌及其构建方法与应用 |
| CN109554324A (zh) * | 2018-12-14 | 2019-04-02 | 江南大学 | 一株产l-异亮氨酸的黄色短杆菌重组菌及其构建方法 |
| CN109971800A (zh) * | 2019-05-17 | 2019-07-05 | 南通普悦生物医药有限公司 | 酶解法制l-羟基脯氨酸的方法 |
| CN110184230A (zh) * | 2019-05-30 | 2019-08-30 | 天津科技大学 | 一株高产l-组氨酸的基因工程菌及其构建方法与应用 |
| CN110564662B (zh) * | 2019-09-30 | 2022-03-25 | 南京农业大学 | 一种整合型高效表达乙醛脱氢酶枯草杆菌的构建方法 |
| WO2021126961A1 (en) * | 2019-12-16 | 2021-06-24 | Ginkgo Bioworks, Inc. | Enhanced production of histidine, purine pathway metabolites, and plasmid dna |
| CN113278567A (zh) * | 2020-02-20 | 2021-08-20 | 杭州立丞生物科技有限公司 | 可改善菌株以提升生产重组蛋白质的方法 |
| CN113322218B (zh) * | 2020-02-28 | 2022-11-22 | 廊坊梅花生物技术开发有限公司 | 重组谷氨酸棒杆菌及生产l-苏氨酸的方法 |
| CN113637699B (zh) * | 2020-04-27 | 2023-05-30 | 中国科学院分子植物科学卓越创新中心 | 一种提高氨基酸产生菌生产能力的方法 |
| CN112063571B (zh) * | 2020-08-14 | 2022-05-06 | 廊坊梅花生物技术开发有限公司 | 高产l-氨基酸的工程菌及其构建方法与应用 |
| CN111996155B (zh) * | 2020-09-08 | 2022-02-11 | 浙江华睿生物技术有限公司 | 一种提高l-组氨酸产生菌生产能力的方法 |
| CN112625992B (zh) * | 2020-12-30 | 2022-07-08 | 宁夏伊品生物科技股份有限公司 | 一种改造基因bbd29_11265产l-谷氨酸的重组菌株及其构建方法与应用 |
| CN112695036B (zh) * | 2021-03-23 | 2021-07-06 | 中国科学院天津工业生物技术研究所 | 一种天冬氨酸激酶基因表达调控序列及其应用 |
| CN113583929B (zh) * | 2021-07-26 | 2024-04-05 | 中国科学院微生物研究所 | 发酵生产嘌呤核苷的重组菌及其构建方法与应用 |
| CN113957073B (zh) * | 2021-10-19 | 2023-09-01 | 山东寿光巨能金玉米开发有限公司 | 一种tkt基因启动子突变体及其在生产L-赖氨酸中的应用 |
| CN116179586A (zh) * | 2021-11-26 | 2023-05-30 | 上海凯赛生物技术股份有限公司 | 用于发酵生产l-赖氨酸的表达盒、菌株及其应用 |
| CN116411007A (zh) * | 2021-12-31 | 2023-07-11 | 上海凯赛生物技术股份有限公司 | 用于制备l-赖氨酸的表达盒、基因工程菌及其应用 |
| CN116411009A (zh) * | 2021-12-31 | 2023-07-11 | 上海凯赛生物技术股份有限公司 | 用于制备l-赖氨酸的表达盒、基因工程菌及其应用 |
| CN116555136A (zh) * | 2022-01-30 | 2023-08-08 | 廊坊梅花生物技术开发有限公司 | 一种修饰的棒状杆菌属微生物及其构建方法与应用 |
| CN117051029A (zh) * | 2022-05-05 | 2023-11-14 | 中国科学院微生物研究所 | 一种顺反子元件库的构建及其在棒杆菌中的应用 |
| CN115948313A (zh) * | 2022-11-10 | 2023-04-11 | 清华大学 | 提高赖氨酸和戊二胺发酵产量的方法 |
| CN116254242B (zh) * | 2022-12-21 | 2024-01-30 | 江南大学 | 一种atp磷酸核苷转移酶突变体及产l-组氨酸的谷氨酸棒杆菌 |
| CN115960874B (zh) * | 2023-02-14 | 2024-09-20 | 山东润德生物科技有限公司 | 一种谷氨酸棒杆菌内源GlcNAc6P磷酸酶及提高GlcNAc产量的方法 |
| KR20250052519A (ko) * | 2023-10-11 | 2025-04-21 | 고려대학교 산학협력단 | 카라기난 분해능 및 갈락토오즈 이용능이 도입된 코리네박테리움 속 균주 및 이를 이용한 해조류로부터 히스티딘을 생산하는 방법 |
Citations (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001004322A1 (en) * | 1999-07-09 | 2001-01-18 | Degussa Ag | Nucleotide sequences which code for the opca gene |
| JP2008523834A (ja) * | 2004-12-22 | 2008-07-10 | ビーエーエスエフ ソシエタス・ヨーロピア | 遺伝子発現のための多重プロモーターおよびその使用 |
| US20090137010A1 (en) * | 2007-09-27 | 2009-05-28 | Rustem Saidovich Shakulov | Method for producing amino acids using bacterium of the enterobacteriaceae family |
Family Cites Families (11)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| ATE335083T1 (de) * | 1993-02-26 | 2006-08-15 | Novozymes As | Methode und system zur erhöhten produktion von kommerziell wichtigen exoproteinen in gram- positiven bakterien |
| US6586214B1 (en) * | 1999-09-15 | 2003-07-01 | Degussa Ag | Method for increasing the metabolic flux through the pentose phosphate cycle in coryneform bacteria by regulation of the phosphoglucose isomerase (pgi gene) |
| JP4623825B2 (ja) * | 1999-12-16 | 2011-02-02 | 協和発酵バイオ株式会社 | 新規ポリヌクレオチド |
| SK287997B6 (sk) | 2000-03-20 | 2012-09-03 | Evonik Degussa Gmbh | Isolated gene of Corynebacterium glutamicum, recombinant coryneform bacteria and process for preparation of L-lysine or L-threonine |
| AU2002247644A1 (en) * | 2001-02-16 | 2002-09-04 | Degussa Ag | Nucleotide sequences of the ribosomal protein s12 gene (rpsl) from corynebacterium glutamicum |
| ATE423843T1 (de) | 2001-12-03 | 2009-03-15 | Kyowa Hakko Kogyo Kk | Mutierte 6-phosphogluconat-dehydrogenase |
| AU2004299729A1 (en) * | 2003-12-18 | 2005-06-30 | Basf Aktiengesellschaft | Methods for the preparation of lysine by fermentation of corynebacterium glutamicum |
| US20070026505A1 (en) | 2005-06-17 | 2007-02-01 | Madden Kevin T | Amino acid and metabolite biosynthesis |
| CN102124119A (zh) | 2008-04-30 | 2011-07-13 | 巴斯夫欧洲公司 | 使用具有降低的异柠檬酸脱氢酶活性的微生物生产精细化学药品的方法 |
| KR101166027B1 (ko) * | 2009-04-01 | 2012-07-19 | 씨제이제일제당 (주) | 5'-이노신산 생산성이 향상된 코리네박테리움 속 미생물 및 이를 이용한 핵산의 생산방법 |
| CN102676509B (zh) * | 2012-05-07 | 2013-12-18 | 中国科学院微生物研究所 | 一种阿拉伯糖诱导的表达载体及其构建方法和应用 |
-
2014
- 2014-02-14 CN CN201410050868.2A patent/CN104845923B/zh active Active
-
2015
- 2015-02-04 EP EP15748508.7A patent/EP3009505B1/en active Active
- 2015-02-04 CN CN201580000936.7A patent/CN106459886B/zh active Active
- 2015-02-04 JP JP2016515651A patent/JP6188930B2/ja active Active
- 2015-02-04 US US14/891,622 patent/US9796991B2/en active Active
- 2015-02-04 KR KR1020157032480A patent/KR101937682B1/ko not_active Expired - Fee Related
- 2015-02-04 WO PCT/CN2015/072220 patent/WO2015120775A1/zh not_active Ceased
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2001004322A1 (en) * | 1999-07-09 | 2001-01-18 | Degussa Ag | Nucleotide sequences which code for the opca gene |
| JP2003504065A (ja) * | 1999-07-09 | 2003-02-04 | デグサ アクチエンゲゼルシャフト | Opca遺伝子をコードするヌクレオチド配列 |
| JP2008523834A (ja) * | 2004-12-22 | 2008-07-10 | ビーエーエスエフ ソシエタス・ヨーロピア | 遺伝子発現のための多重プロモーターおよびその使用 |
| US20090137010A1 (en) * | 2007-09-27 | 2009-05-28 | Rustem Saidovich Shakulov | Method for producing amino acids using bacterium of the enterobacteriaceae family |
Non-Patent Citations (4)
| Title |
|---|
| Mogens Kilstrup 등. FEMS Microbiology Reviews. Vol. 29, No. 3, 페이지 555-592 (First published online 2005.08.28.) 1부. * |
| Robert K. Kulis-Horn 등. Microbial Biotechnology. Vol. 7, 페이지 5-25 (First published 2013.04.25.) 1부. * |
| 일본 공표특허공보 특표2003-504065호(2003.02.04.) 1부. * |
| 일본 공표특허공보 특표2008-523834호(2008.07.10.) 1부. * |
Cited By (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| WO2019190192A1 (ko) * | 2018-03-27 | 2019-10-03 | 씨제이제일제당 (주) | 신규한 프로모터 및 이를 이용한 l-아미노산 생산 방법 |
| WO2019190193A1 (ko) * | 2018-03-27 | 2019-10-03 | 씨제이제일제당 (주) | 글라이신 생산능이 증가된 미생물 및 이를 이용한 발효 조성물 생산 방법 |
| US10961554B2 (en) | 2018-03-27 | 2021-03-30 | Cj Cheiljedang Corporation | Promoter and a method for producing L-amino acid using the same |
| US11661616B2 (en) | 2018-03-27 | 2023-05-30 | Cj Cheiljedang Corporation | Microorganism having increased glycine productivity and method for producing fermented composition using the same |
| KR102281360B1 (ko) * | 2021-01-26 | 2021-07-22 | 씨제이제일제당 (주) | 신규한 atp 포스포리보실트랜스퍼라제 변이체 및 이를 이용한 l-발린 생산 방법 |
| WO2022163923A1 (ko) * | 2021-01-26 | 2022-08-04 | 씨제이제일제당 (주) | 신규한 atp 포스포리보실트랜스퍼라제 변이체 및 이를 이용한 l-발린 생산 방법 |
| WO2022231042A1 (ko) * | 2021-04-30 | 2022-11-03 | 씨제이제일제당 (주) | 신규한 변이체 및 이를 이용한 l-발린 생산 방법 |
| WO2025159448A1 (ko) * | 2024-01-26 | 2025-07-31 | 씨제이제일제당 (주) | 신규한 리보스 포스페이트 디포스포키나제 변이체 및 이를 이용한 l-아미노산 생산 방법 |
Also Published As
| Publication number | Publication date |
|---|---|
| JP6188930B2 (ja) | 2017-08-30 |
| CN104845923B (zh) | 2018-03-23 |
| CN104845923A (zh) | 2015-08-19 |
| EP3009505B1 (en) | 2019-07-03 |
| WO2015120775A1 (zh) | 2015-08-20 |
| KR101937682B1 (ko) | 2019-01-14 |
| US20160326556A1 (en) | 2016-11-10 |
| US9796991B2 (en) | 2017-10-24 |
| EP3009505A4 (en) | 2017-06-07 |
| JP2016519946A (ja) | 2016-07-11 |
| CN106459886B (zh) | 2019-08-20 |
| EP3009505A1 (en) | 2016-04-20 |
| CN106459886A (zh) | 2017-02-22 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| KR101937682B1 (ko) | L-아미노산을 생산하는 재조합균, 이의 구성 방법 및 l-아미노산 생산 방법 | |
| JP6679803B2 (ja) | 新規プロモーター及びその用途 | |
| KR102028554B1 (ko) | 신규한 프로모터 및 이를 이용한 l-아미노산 생산 방법 | |
| CN112795582B (zh) | 一种适合在微生物中高效合成nad衍生物的酶基因 | |
| CN108913642A (zh) | 大肠杆菌基因工程菌及其发酵同步生产l-色氨酸与l-缬氨酸的用途 | |
| CN110117568B (zh) | 生产l-组氨酸的重组菌、其构建方法以及l-组氨酸的生产方法 | |
| KR102078732B1 (ko) | 변형된 막 투과성 | |
| KR20230002331A (ko) | L-라이신을 생산하는 재조합 균주 및 이의 구축 방법과 응용 | |
| US20210324391A1 (en) | Recombinant microorganism, preparation method therefor and application thereof in producing coenzyme q10 | |
| CN106574239B (zh) | 生产l-色氨酸的埃希氏菌属微生物和使用其生产l-色氨酸的方法 | |
| JP3734466B2 (ja) | コバラミンおよび/またはコバミドの生合成に関与するポリペプチド、それらのポリペプチドをコードするdna配列、それらの調製および使用 | |
| KR102712136B1 (ko) | 바이오틴 신타제 활성을 갖는 폴리펩티드 변이체 및 이를 이용한 바이오틴 생산 방법 | |
| JP4898441B2 (ja) | 物質の製造法 | |
| EP4538283A1 (en) | L-histidine export protein and method for producing l-histidine by using same | |
| CN108587991B (zh) | 一种高产环肽类化合物的菌株 | |
| JP2002051790A (ja) | コリネ型細菌のアルギニンリプレッサー欠失株及びl−アルギニンの製造法 | |
| US12351846B2 (en) | Microorganism for producing L-amino acid having increased cytochrome C activity, and L-amino acid production method using same | |
| WO2006025477A1 (ja) | 工業的に有用な微生物 | |
| RU2819270C1 (ru) | Микроорганизм для продуцирования L-аминокислоты, обладающий повышенной активностью цитохрома С, и способ получения L-аминокислоты с его использованием | |
| CN111032874A (zh) | 表达活性d-脯氨酸还原酶的微生物以及生产活性d-脯氨酸还原酶的方法 | |
| CN117070584A (zh) | 烟酰胺腺嘌呤二核苷酸类化合物的合成方法 | |
| KR102016050B1 (ko) | 신규한 프로모터 및 이의 용도 | |
| AU2025268916A1 (en) | Microorganism into which exogenous nad-dependent isocitrate dehydrogenase is introduced and method for producing l-amino acid using same | |
| EP4541808A1 (en) | L-histidine export protein and method of producing l-histidine using same | |
| WO2024144000A1 (ko) | 신규한 폴리뉴클레오티드 및 이를 이용한 l-알라닌 생산 방법 |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| A201 | Request for examination | ||
| E13-X000 | Pre-grant limitation requested |
St.27 status event code: A-2-3-E10-E13-lim-X000 |
|
| P11-X000 | Amendment of application requested |
St.27 status event code: A-2-2-P10-P11-nap-X000 |
|
| P13-X000 | Application amended |
St.27 status event code: A-2-2-P10-P13-nap-X000 |
|
| PA0105 | International application |
St.27 status event code: A-0-1-A10-A15-nap-PA0105 |
|
| PA0201 | Request for examination |
St.27 status event code: A-1-2-D10-D11-exm-PA0201 |
|
| PG1501 | Laying open of application |
St.27 status event code: A-1-1-Q10-Q12-nap-PG1501 |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
St.27 status event code: A-1-2-D10-D21-exm-PE0902 |
|
| P11-X000 | Amendment of application requested |
St.27 status event code: A-2-2-P10-P11-nap-X000 |
|
| P13-X000 | Application amended |
St.27 status event code: A-2-2-P10-P13-nap-X000 |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
St.27 status event code: A-1-2-D10-D21-exm-PE0902 |
|
| E13-X000 | Pre-grant limitation requested |
St.27 status event code: A-2-3-E10-E13-lim-X000 |
|
| P11-X000 | Amendment of application requested |
St.27 status event code: A-2-2-P10-P11-nap-X000 |
|
| P13-X000 | Application amended |
St.27 status event code: A-2-2-P10-P13-nap-X000 |
|
| P22-X000 | Classification modified |
St.27 status event code: A-2-2-P10-P22-nap-X000 |
|
| E14-X000 | Pre-grant third party observation filed |
St.27 status event code: A-2-3-E10-E14-opp-X000 |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
St.27 status event code: A-1-2-D10-D21-exm-PE0902 |
|
| E13-X000 | Pre-grant limitation requested |
St.27 status event code: A-2-3-E10-E13-lim-X000 |
|
| P11-X000 | Amendment of application requested |
St.27 status event code: A-2-2-P10-P11-nap-X000 |
|
| P13-X000 | Application amended |
St.27 status event code: A-2-2-P10-P13-nap-X000 |
|
| E701 | Decision to grant or registration of patent right | ||
| PE0701 | Decision of registration |
St.27 status event code: A-1-2-D10-D22-exm-PE0701 |
|
| GRNT | Written decision to grant | ||
| PR0701 | Registration of establishment |
St.27 status event code: A-2-4-F10-F11-exm-PR0701 |
|
| PR1002 | Payment of registration fee |
St.27 status event code: A-2-2-U10-U12-oth-PR1002 Fee payment year number: 1 |
|
| PG1601 | Publication of registration |
St.27 status event code: A-4-4-Q10-Q13-nap-PG1601 |
|
| PR1001 | Payment of annual fee |
St.27 status event code: A-4-4-U10-U11-oth-PR1001 Fee payment year number: 4 |
|
| PR1001 | Payment of annual fee |
St.27 status event code: A-4-4-U10-U11-oth-PR1001 Fee payment year number: 5 |
|
| R18-X000 | Changes to party contact information recorded |
St.27 status event code: A-5-5-R10-R18-oth-X000 |
|
| PR1001 | Payment of annual fee |
St.27 status event code: A-4-4-U10-U11-oth-PR1001 Fee payment year number: 6 |
|
| PC1903 | Unpaid annual fee |
St.27 status event code: A-4-4-U10-U13-oth-PC1903 Not in force date: 20250108 Payment event data comment text: Termination Category : DEFAULT_OF_REGISTRATION_FEE |
|
| H13 | Ip right lapsed |
Free format text: ST27 STATUS EVENT CODE: N-4-6-H10-H13-OTH-PC1903 (AS PROVIDED BY THE NATIONAL OFFICE); TERMINATION CATEGORY : DEFAULT_OF_REGISTRATION_FEE Effective date: 20250108 |
|
| PC1903 | Unpaid annual fee |
St.27 status event code: N-4-6-H10-H13-oth-PC1903 Ip right cessation event data comment text: Termination Category : DEFAULT_OF_REGISTRATION_FEE Not in force date: 20250108 |