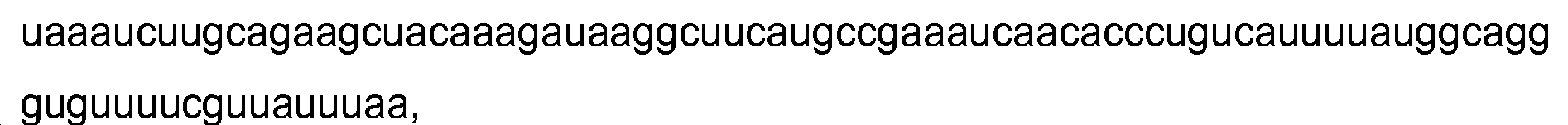모든 인용된 특허 및 비특허 문헌의 개시 내용은 전체적으로 본 명세서에 참고로 포함된다.
본 명세서에 사용되는 바와 같이, 용어 "발명" 또는 "개시된 발명"은 제한적인 것으로 의미되지 않고 청구범위에 정의되거나 본 명세서에 기재된 발명들 중 임의의 것에 일반적으로 적용된다. 이들 용어는 본 명세서에서 상호교환가능하게 사용된다.
본 명세서에서 용어 "통상적이지 않은 효모"는 사카로마이세스 (예를 들어, S. 세레비시아) 또는 스키조사카로마이세스(Schizosaccharomyces) 효모 종이 아닌 임의의 효모를 지칭한다. 통상적이지 않은 효모는, 본 명세서에 참고로 포함되는 문헌[Non-Conventional Yeasts in Genetics, Biochemistry and Biotechnology: Practical Protocols (K. Wolf, K.D. Breunig, G. Barth, Eds., Springer-Verlag, Berlin, Germany, 2003)]에 기재되어 있다. 소정 실시 형태에서 통상적이지 않은 효모는 추가로 (또는 대안적으로) 상동 재조합 (HR)에 의해 매개되는 수복 공정보다 비상동 말단 결합 (non-homologous end-joining; NHEJ) DNA 수복 공정을 선호하는 효모일 수 있다. 이러한 맥락에서 -HR보다 NHEJ를 선호- 통상적이지 않은 효모의 정의가 본 명세서에 참고로 포함되는 문헌 [Chen et aL. (PLoS ONE 8:e57952)]에 또한 개시되어 있다. 본 명세서에서 바람직한 통상적이지 않은 효모는 야로위아 속 (예를 들어, 야로위아 리폴리티카)의 것들이다. 본 명세서에서 용어 "효모"는 주로 단세포 형태로 존재하는 진균종을 지칭한다. 본 명세서에서 효모는 대안적으로 "효모 세포"로 지칭될 수 있다.
본 명세서에서 용어 "RNA-유도 엔도뉴클레아제" (RGEN)는 적어도 하나의 크리스퍼 (규칙적으로 사이 간격을 두고 분포하는 짧은 회문구조 반복 서열)-관련 (Cas) 단백질 및 적어도 하나의 RNA 성분을 포함하는 복합체를 지칭한다. 간략하게, RGEN의 RNA 성분은 표적 부위 서열의 DNA 서열에 상보적인 서열을 함유한다. 이러한 상보성에 기초하여, RGEN은 특정 DNA 표적 부위는 서열을 특이적으로 인식하여 절단할 수 있다. 본 명세서에서 RGEN은 4개의 알려진 크리스퍼 시스템, 예컨대 타입 I, II 또는 III 크리스퍼 시스템 (문헌[Horvath and Barrangou, Science 327:167-170]) 중 어느 하나의 Cas 단백질(들)과 적합한 RNA 성분(들)을 포함할 수 있다. 바람직한 실시 형태의 RGEN은 Cas9 엔도뉴클레아제 (크리스퍼 II 시스템)와 적어도 하나의 RNA 성분 (예를 들어, crRNA 및 tracrRNA, 또는 gRNA)을 포함한다.
용어 "크리스퍼" (규칙적으로 사이 간격을 두고 분포하는 짧은 회문구조 반복 서열)는, 예를 들어, 박테리아 및 고세균 세포에 의해 외래 DNA를 파괴하는데 사용되는 유형 I, II 또는 III DNA 절단 시스템의 인자를 암호화하는 소정 유전자좌(genetic loci)를 지칭한다 (문헌[Horvath and Barrangou, Science 327:167-170]). 크리스퍼 시스템의 성분은 본 명세서에서 통상적이지 않은 효모 세포에서 DNA를 표적화하기 위해 이용된다.
용어 "타입 II 크리스퍼 시스템" 및 "타입 II 크리스퍼-Cas 시스템"은 본 명세서에서 상호교환가능하게 사용되며, 적어도 하나의 RNA 성분과 복합체인 Cas9 엔도뉴클레아제를 이용하는 DNA 절단 시스템을 지칭한다. 예를 들어, Cas9는 크리스퍼 RNA (crRNA) 및 트랜스-활성화 크리스퍼 RNA (tracrRNA)와 복합체일 수 있다. 또 다른 예에서, Cas9는 가이드 RNA와 복합체일 수 있다. 따라서, crRNA, tracrRNA 및 가이드 RNA는 본 명세서에서 RNA 성분의 비제한적인 예이다.
용어 크리스퍼-관련 ("Cas") 엔도뉴클레아제는 본 명세서에서 Cas 유전자에 의해 암호화되는 Cas 단백질을 지칭한다. 적합한 RNA 성분과 복합체인 경우, Cas 엔도뉴클레아제는 소정 실시 형태에서 특정 DNA 표적 서열의 전부 또는 일부를 절단할 수 있다. 예를 들어, 특정 DNA 표적 서열에 단일- 또는 이중 가닥 절단을 도입할 수 있거나; 대안적으로 특정 DNA 표적 서열의 하나 또는 두 가닥을 절단할 수 있는 것을 특징으로 할 수 있다. Cas 엔도뉴클레아제는 표적 서열에서 DNA 이중 가닥을 풀고, Cas와 복합체인 crRNA 또는 가이드 RNA에 의한 표적 서열의 인식에 의해 매개되는 적어도 하나의 DNA 가닥을 절단한다. Cas 엔도뉴클레아제에 의한 이러한 표적 서열의 인식 및 절단은 전형적으로 정확한 프로토스페이서-인접 모티프 (PAM)가 DNA 표적 서열의 3' 말단에 위치하거나 인접한 경우 발생한다. 대안적으로, 본 명세서에서 Cas 단백질은 DNA 절단 또는 닉킹(nicking) 활성이 부족할 수 있지만, 적합한 RNA 성분과 복합체를 형성한 경우 여전히 DNA 표적 서열에 특이적으로 결합할 수 있다. 본 명세서에서 바람직한 Cas 단백질은 Cas9이다.
본 명세서에서 "Cas9" (이전에 Cas5, Csn1 또는 Csx12로 지칭됨)는 DNA 표적 서열의 전부 또는 일부를 특이적으로 인식하고 절단하기 위해, crRNA 및 tracrRNA와, 또는 가이드 RNA와 복합체를 형성하는 타입 II 크리스퍼 시스템의 Cas 엔도뉴클레아제를 지칭한다. Cas9 단백질은 RuvC 뉴클레아제 도메인 및 HNH (H-N-H) 뉴클레아제 도메인을 포함하며, 이들 각각은 표적 서열에서 단일 DNA 가닥을 절단한다 (두 도메인의 협동 작용은 DNA 이중 가닥 절단을 유도하는 반면, 하나의 도메인의 활성은 닉(nick)을 유도한다). 일반적으로, RuvC 도메인은 서브도메인(subdomain) I, II 및 III을 포함하며, 여기서 도메인 I은 Cas9의 N-말단 근처에 위치하고, 서브도메인 II 및 III은 HNH 도메인에 플랭킹한 단백질의 중간에 위치한다 (문헌[Hsu et al, Cell 157:1262-1278]). "Apo-Cas9"는 RNA 성분과 복합체가 형성되지 않은 Cas9를 지칭한다. Apo-Cas9는 DNA를 결합할 수 있지만, 비특이적 방식으로 결합하여, DNA를 절단할 수 없다 (문헌[Sternberg et al., Nature 507:62-67]).
일부 실시 형태에서, Cas 엔도뉴클레아제는 Cas9 폴리펩티드의 변형된 형태를 포함할 수 있다. Cas9 폴리펩티드의 변형된 형태는 Cas9 단백질의 자연 발생 뉴클레아제 활성을 감소시키는 아미노산 변화 (예를 들어, 결실, 삽입 또는 치환)를 포함할 수 있다. 예를 들어, 일부 경우에, Cas9 단백질의 변형된 형태는 상응하는 야생형 Cas9 폴리펩티드의 뉴클레아제 활성의 50% 미만, 40% 미만, 30% 미만, 20% 미만, 10% 미만, 5% 미만 또는 1% 미만을 갖는다 (2014년 3월 6일자로 출원된 미국 특허 출원 제US20140068797 A1호). 일부 경우에서, Cas9 폴리펩티드의 변형된 형태는 실질적인 뉴클레아제 활성을 갖지 않으며, 촉매적으로 "불활성화된 Cas9" 또는 "비활성화된(deactivated) cas9 (dCas9)"로 지칭된다. 촉매적으로 불활성화된 Cas9 변이체는 HNH 및 RuvC 뉴클레아제 도메인에서 돌연변이를 함유하는 Cas9 변이체를 포함한다. 이러한 촉매적으로 불활성화된 Cas9 변이체는 sgRNA와 상호작용하고 생체 내 표적 부위에 결합할 수 있지만, 표적 DNA의 어느 한 가닥을 절단할 수는 없다. DNA에 결합하지만 절단하지 않는 이러한 작용 방식은 영구적인 유전적 변화를 일으키지 않으면서, 염색체의 특정 유전자좌의 발현을 일시적으로 감소시키는 데 사용될 수 있다.
촉매적으로 불활성인 Cas9는 이종 서열에 융합될 수 있다 (2014년 3월 6일자로 출원된 미국 특허 출원 제US20140068797 A1호). 적합한 융합 파트너에는 표적 DNA 상에 또는 표적 DNA와 결합한 폴리펩티드 (예를 들어, 히스톤 또는 다른 DNA-결합 단백질) 상에 직접 작용함으로써 간접적으로 전사를 증가시키는 활성을 제공하는 폴리펩티드가 포함되나, 이에 한정되지 않는다. 추가의 적합한 융합 파트너에는 메틸트랜스퍼라제 활성, 데메틸라제 활성, 아세틸트랜스퍼라제 활성, 데아세틸라제 활성, 키나제 활성, 포스파타제 활성, 유비퀴틴 리가제 활성, 탈유비퀴틴화(deubiquitinating) 활성, 아데닐화 활성, 탈아데닐화 활성, 수모화(SUMOylating) 활성, 탈수모화 활성, 리보실화 활성, 탈리보실화 활성, 미리스토일화 활성 또는 탈미리스토일화 활성을 제공하는 폴리펩티드가 포함되나, 이에 한정되지 않는다 추가로 적합한 융합 파트너에는 표적 핵산의 증가된 전사를 직접적으로 제공하는 폴리펩티드 (예를 들어, 전사 활성 인자 또는 이의 단편, 전사 활성 인자를 동원하는(recruit) 단백질 또는 이의 단편, 소분자/약물-반응성 전사 조절자 등)가 포함되나, 이에 한정되지 않는다. 촉매적으로 불활성인 Cas9는 또한 FokI 뉴클레아제에 융합되어 이중 가닥 절단을 생성할 수 있다 (문헌[Guilinger et aL. Nature biotechnology, volume 32, number 6, June 2014]).
본 명세서에서 용어 "RNA 성분"은 DNA 표적 서열의 가닥에 상보적인 리보핵산 서열을 함유하는 RGEN의 RNA 성분을 지칭한다. 본 명세서에서 이러한 상보적인 서열은 "가이드 서열" 또는 "가변 표적화 도메인" 서열로 지칭된다. 본 명세서에서 적합한 RNA 성분의 예는 crRNA 및 가이드 RNA를 포함한다. 또한, 본 명세서에서 RNA 성분은 5'-캡을 가지지 않는다.
본 명세서에서 용어 "크리스퍼 RNA" (crRNA)는 하나 이상의 Cas 단백질 (예를 들어, Cas9)과 복합체를 형성할 수 있어, 복합체에 DNA 결합 특이성을 제공하는 RNA 서열을 지칭한다. crRNA는 DNA 표적 서열의 가닥에 상보적인 "가이드 서열" ("가변 표적화 도메인" [VT])을 함유하기 때문에 DNA 결합 특이성을 제공하다. crRNA는 crRNA가 유래된 크리스퍼 유전자좌의 반복 영역에 의해 암호화되는 "반복 서열" ("tracr RNA 메이트 서열")을 추가로 포함한다. crRNA의 반복 서열은 tracrRNA의 5'-말단에서 서열에 어닐링할 수 있다. 고유 크리스퍼 시스템의 crRNA는 크리스퍼 유전자좌로부터 전사된 "예비-crRNA"로부터 유래된다. 예비-crRNA는 스페이서 영역과 반복 영역을 포함하고; 스페이서 영역은 DNA 표적 부위는 서열에 상보적인 특유의 서열을 함유한다. 고유 시스템의 예비-crRNA는 다수의 상이한 crRNA로 처리되며, 각각은 반복 서열의 일부와 함께 가이드 서열을 갖는다. 크리스퍼 시스템은, 예를 들어, DNA 표적화 특이성을 위해 crRNA를 이용한다.
본 명세서에서 용어 "트랜스-활성화 크리스퍼 RNA" (tracrRNA)는 타입 II 크리스퍼 시스템에서 사용되며, 5'에서 3' 방향으로 (i) 크리스퍼 타입 II crRNA의 반복 영역과 어닐링하는 서열 및 (ii) 스템 루프-함유하는 부분을 함유하는 비암호화 RNA를 지칭한다 (문헌[Deltcheva et al., Nature 471:602-607]).
용어 "가이드 RNA" (gRNA) 및 "단일 가이드 RNA" (sgRNA)는 본 명세서에서 상호교환가능하게 사용된다. 본 명세서에서 gRNA는 tracrRNA에 작동가능하게 연결된 crRNA를 함유하는 키메라 서열을 지칭할 수 있다. 대안적으로, gRNA는, 예를 들어 crRNA와 tracrRNA의 합성 융합을 지칭할 수 있다. 문헌[Jinek et aL. (Science 337:816-821)]은 일부 gRNA 특징을 개시한다. gRNA는 또한 가이드 서열 (가변 표적화 도메인) 다음에 Cas 엔도뉴클레아제 인식 (CER) 도메인을 갖는 것을 특징으로 할 수 있다 [모두 본 명세서에 전체적으로 참고로 포함되는 2015년 2월 26일자로 출원된 WO2015026883, 2015년 3월 19일자로 출원된 미국 특허 출원 제US 2015-0082478 A1호 및 2015년 2월 26일자로 출원된 제US 2015-0059010 A1호]. CER 도메인은 tracrRNA 메이트 서열 다음에 tracrRNA 서열을 포함한다.
본 명세서에서 용어 "표적 부위 서열", "표적 부위", "표적 서열", "표적 DNA", "DNA 표적 서열", "표적 유전자좌", "게놈 표적 부위", "게놈 표적 서열", "게놈 표적 유전자좌" 및 "프로토스페이서"는 상호교환가능하게 사용된다. 표적 부위 서열은 본 명세서의 RGEN가 인식하고, 결합하며, 임의로 닉을 형성하거나 절단하는 통상적이지 않은 효모의 게놈 내의 염색체, 에피솜 또는 임의의 다른 DNA 분자 상의 폴리뉴클레오티드 서열을 지칭한다. 표적 부위는 (i) 효모 내의 내인성/고유 부위이거나, (ii) 효모에 이종성이어서 게놈에서 자연적으로 발생하지 않거나 (iii) 이가 고유하게 발생하는 위치에 비교하여 이종성인 게놈 위치에서 발견되는 것일 수 있다.
본 명세서에서 표적 부위 서열은 길이가 적어도 13 뉴클레오티드이고, 가이드 서열과 혼성화되고, (소정 실시 형태에서, 적합한 PAM이 표적 서열에 인접한 경우) 표적 서열에 Cas 단백질 또는 Cas 단백질 복합체를 직접 서열-특이적으로 결합시킬 수 있는 (crRNA 또는 gRNA의) 가이드 서열에 대한 충분한 상보성을 갖는 가닥을 갖는다. (엔도뉴클레오리틱(endonucleolytic) 또는 닉킹 Cas와 적용가능한) 절단/닉 부위는 (예를 들어, Cas9를 사용) 표적 서열 내에 있거나, 절단/닉 부위는 (예를 들어, FokI 효소로부터 유래된 것과 같은 이종 엔도뉴클레아제 도메인에 융합된 Cas9를 사용) 표적 서열 외부에 있을 수 있다.
본 명세서에서 "인공 표적 부위" 또는 "인공 표적 서열"은 통상적이지 않은 효모의 게놈에 도입된 표적 서열을 지칭한다. 일부 실시 형태에서, 인공 표적 서열은 효모의 게놈 내의 고유 표적 서열과 서열이 동일할 수 있지만, 게놈의 상이한 위치 (이종 위치)에 위치할 수 있거나, 효모의 게놈 내의 동일한 위치에 위치하는 경우 고유 표적 서열과 상이할 수 있다.
본 명세서에서 "에피솜"은 효모 세포의 염색체와 별도로 효모 세포에 자율적으로 존재할 수 있는 (복제할 수 있고 딸 세포로 전달할 수 있는) DNA 분자를 지칭한다. 에피솜 DNA는 효모세포에 고유하거나 이종성일 수 있다. 본 명세서에서 고유 에피솜의 예에는 미토콘드리아 DNA (mtDNA)가 포함된다. 본 명세서에서 이종 에피솜의 예에는 플라스미드 및 효모 인공 염색체 (YAC)가 포함된다.
본 명세서에서 "프로토스페이서 인접 모티프" (PAM)는 본 명세서의 RGEN에 의해 인식되는 짧은 서열을 지칭한다. 본 명세서의 PAM의 서열과 길이는 사용되는 Cas 단백질 또는 Cas 단백질 복합체에 따라 다를 수 있지만, 전형적으로는, 예를 들어 2, 3, 4, 5, 6, 7 또는 8개의 뉴클레오티드 길이이다.
용어 "5'-캡" 및 "7-메틸구아닐레이트 (m7G) 캡"은 본 명세서에서 상호교환가능하게 사용된다. 7-메틸구아닐레이트 잔기는 진핵생물에서 메신저 RNA (mRNA)의 5′ 말단 상에 위치한다. RNA 폴리머라제 II (Pol II)는 진핵생물에서 mRNA를 전사한다. 메신저 RNA 캡핑은 일반적으로 다음과 같이 발생한다: mRNA 전사물의 최말단 5' 포스페이트기는 RNA 말단 포스파타제에 의해 제거되어, 2개의 말단 포스페이트를 남긴다. 구아노신 모노포스페이트 (GMP)는 구아닐릴 트랜스퍼라제에 의해 전사물의 말단 포스페이트에 첨가되어, 전사물 말단에 5′-5′ 트라이포스페이트-연결된 구아닌을 남긴다. 마지막으로, 이러한 말단 구아닌의 7-질소가 메틸 트랜스퍼라제에 의해 메틸화된다.
본 명세서에서 용어 "5'-캡을 갖지 않는"은, 예를 들어, 5'-캡 대신에 5'-하이드록실기를 갖는 RNA를 지칭하는데 사용된다. 이러한 RNA는, 예를 들어 "캡핑되지 않은 RNA"로 지칭될 수 있다. 캡핑되지 않은 RNA는 5'-캡핑된 RNA가 핵외수송(nuclear export)의 대상이기 때문에 전사 후 핵에 더 잘 축적될 수 있다. 본 명세서에서 하나 이상의 RNA 성분이 캡핑되지 않는다
용어 "리보자임" 및 "리보핵산 효소"는 본 명세서에서 상호교환가능하게 사용된다. 리보자임은 특정 부위에서 RNA를 절단할 수 있는 2차, 3차, 및/또는 4차 구조(들)를 형성하는 하나 이상의 RNA 서열을 지칭한다. 리보자임은 리보자임 서열에 대하여 시스-부위에서 RNA를 절단할 수 있는 "자가 절단(self-cleaving) 리보자임"을 포함한다 (즉, 자가 촉매적(auto-catalytic) 또는 자가 절단). 리보자임 핵산 분해(nucleolytic) 활성의 일반적인 성질이 기재되어 있다 (예를 들어, 문헌[Lilley, BiocheM. SoC. TranS. 39:641-646]). 본 명세서에서 "망치머리 리보자임" (HHR)은 촉매 작용에 관여하는 3개의 염기쌍 형성된 스템 및 고도로 보존된, 비상보적 뉴클레오티드의 코어로 구성되는 작은 촉매 RNA 모티프를 포함할 수 있다. 본 명세서에 참고로 포함되는 문헌[Pley et aL. (Nature 372:68-74) and Hammann et aL. (RNA 18:871-885)]은 망치머리 리보자임 구조와 활성을 개시하고 있다. 본 명세서에서 망치머리 리보자임은, 예를 들어, 본 명세서에 참고로 포함되는 문헌[by Scott et aL. (Cell 81:991-1002])에 개시된 바와 같은 "최소 망치머리" 서열을 포함할 수 있다.
본 발명의 일 실시 형태에서, 방법은 통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법을 포함하며, 상기 방법은 상기 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 RNA 성분의 상류에 리보자임을 암호화하는 DNA 서열을 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하고, 여기서 제2 재조합 DNA 구조물로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하며, RNA 성분과 Cas9 엔도뉴클레아제는 표적 부위 서열의 전부 또는 일부에 결합할 수 있는 RGEN을 형성할 수 있다.
본 발명의 일 실시 형태에서, 통상적이지 않은 효모는 적어도 하나의 뉴클레오티드 서열에 작동가능하게 연결된 프로모터를 포함하는 폴리뉴클레오티드 서열을 포함하며, 여기서 상기 뉴클레오티드 서열은 RNA 성분을 암호화하는 DNA 서열의 상류에 리보자임을 암호화하는 DNA 서열을 포함하고, 상기 RNA 성분은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열에 상보적인 가변 표적화 도메인을 포함하며, RNA 성분은 RNA-유도 엔도뉴클레아제 (RGEN)를 형성할 수 있고, 상기 RGEN는 표적 부위 서열의 전부 또는 일부에 결합할 수 있으며, 뉴클레오티드 서열로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하고, 상기 RNA 성분은 5' 캡을 포함하지 않는다.
리보자임은 또한 임의의 선행 전사물을 제거하지만 리보자임 서열은 온전히 남겨두는 자체 서열의 5'을 절단하는 리보자임을 포함한다.
본 발명의 일 실시 형태에서, 통상적이지 않은 효모는 적어도 하나의 뉴클레오티드 서열에 작동가능하게 연결된 프로모터를 포함하는 폴리뉴클레오티드 서열을 포함하며, 여기서 상기 뉴클레오티드 서열은 RNA 성분을 암호화하는 DNA 서열의 상류에 리보자임을 암호화하는 DNA 서열을 포함하고, 상기 RNA 성분은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열에 상보적인 가변 표적화 도메인을 포함하며, RNA 성분은 RNA-유도 엔도뉴클레아제 (RGEN)를 형성할 수 있고, 상기 RGEN는 표적 부위 서열의 전부 또는 일부에 결합할 수 있으며, 뉴클레오티드 서열로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하고, 뉴클레오티드 서열로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하지 않아, 5' 캡이 없이 리보자임-RNA 성분 융합 분자를 생성한다.
용어 "표적화", "유전자 표적화", "DNA 표적화", "편집", "유전자 편집" 및 "DNA 편집"은 본 명세서에서 상호교환가능하게 사용된다. 본 명세서에서 DNA 표적화는 통상적이지 않은 효모의 염색체 또는 에피솜에서와 같은 특정 DNA 서열에서의 삽입-결실, 녹아웃 또는 녹인의 특이적 도입일 수 있다. 일반적으로, DNA 표적화는 본 명세서에서 적합한 RNA 성분과 결합한 Cas 단백질을 사용하여 통상적이지 않은 효모의 특정 DNA 서열에서 하나 또는 두 가닥을 절단함으로써 수행될 수 있다. 이러한 DNA 절단은, 이중 가닥 절단 (DSB)의 경우 표적 부위에서 삽입-결실을 형성할 수 있는 NHEJ 공정을 유도할 수 있다. 또한, 절단이 단일-가닥 절단 (SSB) 또는 DSB인지 여부에 관계없이, DNA 닉 또는 절단 부위에 적합한 공여 DNA 폴리뉴클레오티드가 제공되는 경우 HR 공정은 유도될 수 있다. 이러한 HR 공정은 공여 DNA 폴리뉴클레오티드의 서열에 따라 표적 부위에서 녹아웃 또는 녹인을 도입하는 데 사용될 수 있다.
대안적으로, 본 명세서에서 DNA 표적화는 본 명세서의 Cas/RNA 성분 복합체와 표적 DNA 서열의 특정 결합을 지칭할 수 있으며, 여기서 Cas 단백질은 DNA 가닥을 절단하거나 절단하지 않는다 (Cas 단백질의 엔도뉴클레오리틱 도메인의 상태에 따라).
본 명세서에서 용어 "삽입-결실"은 염색체 또는 에피솜 내의 표적 DNA 서열에서의 뉴클레오티드 염기의 삽입 또는 결실을 지칭한다. 이러한 삽입 또는 결실은, 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 그 이상의 염기일 수 있다. 소정 실시 형태에서, 삽입-결실은 심지어 더 클 수 있고, 적어도 약 20, 30, 40, 50, 60, 70, 80, 90 또는 100개의 염기일 수 있다. 삽입-결실이 유전자의 오픈 리딩 프레임 (ORF) 내에 도입되는 경우, 종종 삽입-결실은 프레임시프트 돌연변이를 생성함으로써 ORF에 의해 암호화되는 단백질의 야생형 발현을 방해한다.
용어 "녹아웃", "유전자 녹아웃" 및 "유전적 녹아웃"은 본 명세서에서 상호교환가능하게 사용된다. 본 명세서에서 녹아웃은 Cas 단백질로 표적화함으로써 부분적으로 또는 완전히 작동하지 않게 된 통상적이지 않은 효모의 DNA 서열을 나타내며; 녹아웃 이전의 이러한 DNA 서열은, 예를 들어, 아미노산 서열을 암호화할 수 있거나 조절 기능 (예를 들어, 프로모터)을 가질 수 있다. 녹아웃은 (NHEJ에 의한) 삽입-결실에 의해 또는, 표적화 부위 또는 그 부근에서 서열의 기능을 감소시키거나 완전히 파괴시키는 서열의 특이적 제거에 의해 생성될 수 있다. 본 명세서에서 녹아웃된 DNA 폴리뉴클레오티드 서열은 대안적으로, 예를 들어, 부분적으로 또는 완전히 파괴되거나 하향조절되는 것을 특징으로 할 수 있다.
일 실시 형태에서, 본 발명은 Cas9 엔도뉴클레아제 및 적어도 하나의 뉴클레오티드 서열에 작동가능하게 연결된 프로모터를 포함하는 폴리뉴클레오티드 서열을 포함하는 통상적이지 않은 효모에 관한 것이며, 여기서 상기 뉴클레오티드 서열은 RNA 성분을 암호화하는 DNA 서열의 상류에 리보자임을 암호화하는 DNA 서열을 포함하고, 상기 RNA 성분은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열에 상보적인 가변 표적화 도메인을 포함하며, RNA 성분은 Cas 엔도뉴클레아제와 함께, 표적 부위 서열에 결합할 수 있는 RNA-유도 엔도뉴클레아제 (RGEN)를 형성할 수 있다. Cas9 엔도뉴클레아제는 효모에서 단백질로서 도입될 수 있거나 재조합 DNA 구조물을 통해 도입될 수 있다. Cas9 엔도뉴클레아제는 본 기술 분야에 알려진 임의의 방법에 의해 안정적이거나 일과성 방식으로 발현될 수 있다.
용어 "녹인", "유전자 녹인" 및 "유전적 녹인"은 본 명세서에서 상호교환가능하게 사용된다. 녹인은 Cas 단백질을 사용하여 표적화함으로써 통상적이지 않은 효모의 특정 DNA 서열에서 DNA 서열을 대체하거나 삽입하는 것을 나타낸다. 녹인의 예는 유전자의 암호화 영역에 이종 아미노산 암호화 서열의 특이적 삽입 또는 유전자좌에 전사 조절 요소의 특이적 삽입이다.
용어 "공여 폴리뉴클레오티드", "공여 DNA", "표적화 폴리뉴클레오티드" 및 "표적화 DNA"는 본 명세서에서 상호교환가능하게 사용된다. 공여 폴리뉴클레오티드는 DNA 표적 부위 (예를 들어, 본 명세서의 Cas 단백질에 의해 특이적으로 표적화된 서열)에 또는 그 부근의 서열에 상동성인 적어도 하나의 서열을 포함하는 DNA 서열을 지칭한다. 편집될 뉴클레오티드 서열과 비교하는 경우, 적어도 하나의 뉴클레오티드 변형을 포함하는 폴리뉴클레오티드를 포함하는 공여 DNA 폴리뉴클레오티드는 "폴리뉴클레오티드 변형 주형", "폴리뉴클레오티드 변형 주형 DNA" 또는 "주형 DNA"로도 지칭된다. 뉴클레오티드 변형은 적어도 하나의 뉴클레오티드 치환, 첨가 또는 결실일 수 있다. 임의로, 폴리뉴클레오티드 변형 주형은 적어도 하나의 뉴클레오티드 변형에 플랭킹된 상동성 뉴클레오티드 서열을 추가로 포함할 수 있으며, 여기서 플랭킹 상동성 뉴클레오티드 서열은 편집될 원하는 뉴클레오티드 서열에 대한 충분한 상동성을 제공한다.
본 명세서에서 공여 폴리뉴클레오티드 내의 "상동성 서열"은 표적 부위 또는 그 부근의 서열과 100% 동일성, 또는 표적 부위 또는 그 부근의 서열과 적어도 약 95%, 96%, 97%, 98% 또는 99% 동일성을 갖는 적어도 약 25개의 뉴클레오티드의 서열을 포함하거나 이로 구성될 수 있다.
소정 실시 형태에서, 공여 DNA 폴리뉴클레오티드는 표적 부위에서 서열과 이종성인 서열에 의해 분리된 2개의 상동성 서열을 가질 수 있다. 이러한 공여 폴리뉴클레오티드의 이들 2개의 상동성 서열은 이종 서열에 플랭킹된 "상동성 암"으로 지칭될 수 있다. 2개의 상동성 암을 갖는 표적 부위와 공여 폴리뉴클레오티드 사이의 HR은 전형적으로 표적 부위에서 서열을 공여 폴리뉴클레오티드의 이종 서열로 대체시킨다 (공여 폴리뉴클레오티드의 상동성 암에 상동성인 DNA 서열 사이에 위치한 표적 부위 서열은 공여 폴리뉴클레오티드의 이종 서열에 의해 대체된다). 2개의 상동성 암을 갖는 공여 폴리뉴클레오티드에서, 암은 1개 이상의 뉴클레오티드에 의해 분리될 수 있다 (즉, 공여 폴리뉴클레오티드의 이종 서열은 길이가 적어도 1개의 뉴클레오티드일 수 있다). 본 명세서에서 통상적이지 않은 효모에서 수행될 수 있는 다양한 HR 절차가, 예를 들어, 본 명세서에 참고로 포함되는 문헌[DNA Recombination: Methods and Protocols: 1st Edition (H. Tsubouchi, Ed., Springer-Verlag, New York, 2011)]에 개시되어 있다.
일 실시 형태에서, 공여 DNA 구조물은 Cas 엔도뉴클레아제의 표적 부위로 삽입될 관심 폴리뉴클레오티드를 포함하며, 여기서 공여 DNA 구조물은 관심 폴리뉴클레오티드에 플랭킹된 제1 및 제2 상동성 영역을 추가로 포함한다. 공여 DNA의 제1 및 제2 상동성 영역은 각각 식물 게놈의 표적 부위에 존재하거나 플랭킹된 제1 및 제2 게놈 영역과 상동성을 공유한다.
용어 "부피 %", "부피 백분율", "vol %" 및 "v/v %"는 본 명세서에서 상호교환가능하게 사용된다. 용액 중의 용질의 부피%는 하기 식을 사용하여 결정될 수 있다: [(용질의 부피)/(용액의 부피)] × 100%.
용어 "중량%", "중량 백분율 (wt%)" 및 "중량-중량 백분율(% w/w)"은 본 명세서에서 상호교환가능하게 사용된다. 중량%는 재료가 조성물, 혼합물 또는 용액 중에 포함될 때 질량을 기준으로 한 재료의 백분율을 지칭한다.
용어 "폴리뉴클레오티드", "폴리뉴클레오티드 서열" 및 "핵산 서열"은 본 명세서에서 상호교환가능하게 사용된다. 이들 용어는 뉴클레오티드 서열 등을 포함한다. 폴리뉴클레오티드는 합성된, 비천연 뉴클레오티드 염기 또는 변경된 뉴클레오티드 염기를 임의로 함유하는 단일- 또는 이중-가닥인 DNA 또는 RNA 의 중합체일 수 있다. 폴리뉴클레오티드는 cDNA, 게놈 DNA, 합성 DNA, 또는 이의 혼합물의 하나 이상의 세그먼트로 이루어질 수 있다. 뉴클레오티드 (리보뉴클레오티드 또는 데옥시리보뉴클레오티드)는 다음과 같이 단일 문자 표기로 지칭될 수 있다: (각각 RNA 또는 DNA에 있어서) 아데닐레이트 또는 데옥시아데닐레이트는 "A", (각각 RNA 또는 DNA에 있어서) 시티딜레이트 또는 데옥시시티딜레이트는 "C", (각각 RNA 또는 DNA에 있어서) 구아닐레이트 또는 데옥시구아닐레이트는 "G", (RNA에 있어서) 유리딜레이트는 "U", (DNA에 있어서) 데옥시티미딜레이트는 "T", 퓨린 (A 또는 G)은 "R", 피리미딘 (C 또는 T)은 "Y", G 또는 T는 "K", A 또는 C 또는 T는 "H", 이노신은 "I", A 또는 T는 "W" 및 임의의 뉴클레오티드는 "N" (예를 들어, DNA 서열을 말하는 경우 N은 A, C, T 또는 G일 수 있고; RNA 서열을 말하는 경우 N은 A, C, U 또는 G일 수 있다). 본 명세서에 개시된 임의의 RNA 서열 (예를 들어, crRNA, tracrRNA, gRNA)은 적합한 DNA 서열에 의해 암호화될 수 있다.
본 명세서에서 사용되는 바와 같은 용어 "단리된"은 이의 천연 공급원으로부터 완전히 또는 부분적으로 정제된 폴리뉴클레오티드 또는 폴리펩티드 분자를 지칭한다. 일부 경우에, 단리된 폴리뉴클레오티드 또는 폴리펩티드 분자는 더 큰 조성물, 완충 시스템 또는 시약 믹스의 일부이다. 예를 들어, 단리된 폴리뉴클레오티드 또는 폴리펩티드 분자는 이종성 방식으로 세포 또는 유기체 내에 포함될 수 있다.
본 명세서에서 사용되는 바와 같은 용어 "유전자"는 암호화 영역으로부터 RNA (RNA는 DNA 폴리뉴클레오티드 서열로부터 전사됨)를 발현하는 DNA 폴리뉴클레오티드 서열을 지칭하며, RNA는 (단백질을 암호화하는) 메신저 RNA 또는 비-단백질-암호화 RNA (예를 들어, 본 명세서에서 crRNA, tracrRNA 또는 gRNA)일 수 있다. 유전자는 암호화 영역만을 지칭할 수 있거나, 암호화 영역 (예를 들어, 프로모터, 5'-비번역 영역, 3'-전사 종결 영역)의 상류 및/또는 하류의 조절 서열을 포함할 수 있다. 단백질을 암호화하는 암호화 영역은 본 명세서에서 대안적으로 "오픈 리딩 프레임" [ORF]으로 지칭될 수 있다. "고유" 또는 "내인성"인 유전자는 자연에서 그 자체의 조절 서열을 갖는 것으로 발견되는 유전자를 지칭하며; 이러한 유전자는 숙주 세포의 게놈에서 이의 자연적 위치에 위치한다. "키메라" 유전자는 자연에서 함께 발견되지 않는 조절 및 암호화 서열을 포함하는, 고유 유전자가 아닌 임의의 유전자를 말한다 (즉, 조절 영역과 암호화 영역이 서로 이종성이다). 따라서, 키메라 유전자는 상이한 공급원으로부터 유래된 조절 서열과 암호화 서열, 또는 동일한 공급원으로부터 유래되었지만, 자연에서 발견되는 것과 상이한 방식으로 배열된 조절 서열과 암호화 서열을 포함할 수 있다. "외래" 또는 "이종" 유전자는 유전자 전달에 의해 숙주 유기체 내로 도입된 유전자를 지칭한다. 외래 유전자는 비-천연 유기체에 삽입된 천연 유전자, 천연 숙주 내의 새로운 위치로 도입된 천연 유전자, 또는 키메라 유전자를 포함할 수 있다. 본 명세서에 개시된 소정 실시 형태에서, 폴리뉴클레오티드 서열은 이종성이다. "트랜스유전자(transgene)"는 형질전환 절차에 의해 게놈 내로 도입된 유전자이다. "코돈-최적화된" 오픈 리딩 프레임은 이의 코돈 사용 빈도가 숙주 세포의 바람직한 코돈 사용 빈도를 모방하도록 설계된 유전자이다.
고유 아미노산 서열 또는 폴리뉴클레오티드 서열은 자연 발생적인 반면, 비천연 아미노산 서열 또는 폴리뉴클레오티드 서열은 자연에서 발생하지 않는다.
본 명세서에서 사용되는 바와 같은 "조절 서열"은 유전자의 전사 시작 부위 (예를 들어, 프로모터), 5' 비번역 영역 및 3' 비암호화 영역의 상류에 위치하며, 전사, 프로세싱 또는 안정성 또는 유전자로부터 전사된 RNA의 번역에 영향을 줄 수 있는 뉴클레오티드 서열을 지칭한다. 본 명세서에서 조절 서열은 프로모터, 인핸서, 사일런서(silencer), 5' 비번역 선도 서열, 인트론, 폴리아데닐화 인식 서열, RNA 프로세싱 부위, 이펙터 결합 부위, 스템-루프 구조 및 유전자 발현의 조절에 관여하는 다른 요소를 포함할 수 있다. 본 명세서에서 하나 이상의 조절 요소는 본 명세서의 암호화 영역에 이종성일 수 있다.
본 명세서에서 사용되는 바와 같은 "프로모터"는 유전자로부터의 RNA의 전사를 조절할 수 있는 DNA 서열을 지칭한다. 일반적으로, 프로모터 서열은 유전자의 전사 시작 부위의 상류에 있다. 프로모터는 고유 유전자로부터 그 전체가 유래될 수도 있거나, 자연에서 발견되는 상이한 프로모터로부터 유래된 상이한 요소로 구성될 수도 있거나, 심지어 합성 DNA 세그먼트를 포함할 수도 있다. 대부분의 세포 유형에서 대부분의 경우에 유전자가 발현되도록 하는 프로모터는 통상적으로 "항시성 프로모터(constitutive promoter)"라고 지칭된다. 본 명세서에서 하나 이상의 프로모터는 본 명세서의 암호화 영역에 이종성일 수 있다.
본 명세서에서 사용되는 바와 같은 "강력한 프로모터"는 단위 시간당 상대적으로 많은 수의 생산 개시를 이끌 수 있는 프로모터를 지칭하고/하며 효모 내 유전자의 평균 전사 수준보다 더 높은 수준의 유전자 전사를 유도하는 프로모터이다.
본 명세서에서 사용되는 바와 같은 용어 "3' 비암호화 서열", "전사 종결자" 및 "종결자"는 암호화 서열의 하류에 위치한 DNA 서열을 지칭한다. 이는 폴리아데닐화 인식 서열 및 mRNA 프로세싱 또는 유전자 발현에 영향을 줄 수 있는 조절 신호를 암호화하는 다른 서열을 포함한다.
본 명세서에서 사용되는 바와 같은 용어 "카세트"는 단백질 또는 비-단백질-암호화 RNA를 암호화하는 서열에 작동가능하게 연결된 프로모터를 지칭한다. 카세트는 임의로 3' 비암호화 서열에 작동가능하게 연결될 수 있다.
폴리뉴클레오티드와 관련하여 본 명세서에서 사용되는 바와 같은 용어 "상류" 및 "하류"는 각각 "이의 5'" 및 "이의 3'"을 지칭한다.
본 명세서에서 사용되는 바와 같은 용어 "발현"은 (i) 암호화 영역으로부터 RNA의 전사 (예를 들어, mRNA, 또는 crRNA, tracrRNA 또는 gRNA와 같은 비-단백질 암호화 RNA) 또는 (ii) mRNA로부터의 폴리펩티드의 번역을 지칭한다.
유전자 또는 폴리뉴클레오티드 서열의 발현을 설명하는 데 사용되는 경우, 용어 "하향-조절", "파괴", "억제", "불활성화" 및 "침묵화"는 폴리뉴클레오티드 서열의 전사가 감소되거나 제거되는 경우를 지칭하도록 본 명세서에서 상호교환가능하게 사용된다. 이는 폴리뉴클레오티드 서열로부터의 RNA 전사물의 감소 또는 제거를 초래하며, 이는 (유전자가 ORF를 포함하는 경우) 폴리뉴클레오티드 서열로부터 유래된 단백질 발현의 감소 또는 제거를 초래한다. 대안적으로, 하향-조절은 폴리뉴클레오티드 서열에 의해 생성된 전사물로부터의 단백질 번역이 감소되거나 제거된 경우를 지칭할 수 있다. 또한 대안적으로, 하향-조절은 폴리뉴클레오티드 서열에 의해 발현된 단백질의 활성이 감소된 경우를 지칭할 수 있다. 세포에서 상기 공정 (전사, 번역, 단백질 활성) 중 어느 하나의 감소는 적합한 대조 세포의 전사, 번역 또는 단백질 활성에 대하여 약 40%, 50%, 60%, 70%, 80%, 90%, 95% 또는 100%일 수 있다. 하향-조절은, 예를 들어, 본 명세서에 개시된 바와 같은 표적화 이벤트 (예를 들어, 삽입-결실, 녹아웃)의 결과일 수 있다.
용어 "대조 세포" 및 "적합한 대조 세포"는 본 명세서에서 상호교환가능하게 사용되며, 특정 변형 (예를 들어, 폴리뉴클레오티드의 과발현, 폴리뉴클레오티드의 하향-조절)이 만들어진 세포 (즉, "실험 세포")와 관련하여 언급될 수 있다. 대조 세포는 실험 세포의 특정 변형을 가지지 않거나 발현하지 않는 임의의 세포일 수 있다. 따라서, 대조 세포는 형질전환되지 않은 야생형 세포일 수 있거나, 유전적으로 변형될 수 있지만 유전적 형질전환을 발현하지 않는다. 예를 들어, 대조 세포는 실험 세포의 직접적인 부모일 수 있으며, 직접적인 부모 세포는 실험 세포에 있는 특정 변형을 가지지 않는다. 대안적으로, 대조 세포는 하나 이상의 세대에 의해 제거되는 실험 세포의 부모일 수 있다. 또한 대안적으로, 대조 세포는 실험 세포의 형제일 수 있으며, 형제 세포는 실험 세포에 존재하는 특정 변형을 포함하지 않는다.
본 명세서에서 사용되는 바와 같은 용어 "증가된"은 증가된 양 또는 활성이 비교되는 양 또는 활성보다 적어도 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19% 또는 20% 초과인 양 또는 활성을 지칭할 수 있다. 용어 "증가된", "보다 많은" 및 "개선된"은 본 명세서에서 상호교환가능하게 사용된다. 용어 "증가된"은 단백질을 암호화하는 폴리뉴클레오티드의 발현을 특성화하는 데 사용될 수 있는데, 예를 들어, "증가된 발현"은 또한 "과발현"을 의미한다.
본 명세서에서 사용되는 바와 같은 용어 "작동가능하게 연결된"은 하나의 기능이 다른 것에 의해 영향을 받는 둘 이상의 핵산 서열의 결합을 지칭한다. 예를 들어, 프로모터는 그것이 암호화 서열의 발현에 영향을 미칠 수 있는 경우에 암호화 서열에 작동가능하게 연결된다. 다시 말하면, 암호화 서열은 프로모터의 전사적 조절 하에 있다. 암호화 서열은, 예를 들어, 조절 서열에 작동가능하게 연결될 수 있다. 또한, 예를 들어, crRNA는 crRNA의 tracrRNA 메이트 서열이 tracrRNA의 5' 서열과 어닐링하도록 본 명세서의 tracrRNA에 작동가능하게 연결 (융합)될 수 있다. 이러한 작동가능한 연결은 적합한 루프 형성 서열, 예컨대 GAAA (서열 번호:43), CAAA (서열 번호:44) 또는 AAAG (서열 번호:45)를 포함할 수 있다.
본 명세서에서 사용되는 바와 같은 용어 "재조합"은, 예를 들어, 화학적 합성에 의한 또는 유전자 조작 기술에 의해 분리된 핵산 세그먼트의 조작에 의한, 그렇지 않다면 분리되어 있는 2개의 서열 세그먼트의 인공적인 조합을 지칭한다. 용어 "재조합", "트랜스제닉(transgenic)", "형질전환된", "유전자 조작된" 또는 "외인성 유전자 발현을 위해 변형된"은 본 명세서에서 상호교환가능하게 사용된다.
본 명세서의 재조합 구조물/벡터 (예를 들어, 본 명세서의 리보자임-RNA 성분 카세트를 암호화하는 DNA 폴리뉴클레오티드 또는 본 명세서의 Cas 단백질을 암호화하는 DNA 폴리뉴클레오티드)를 제조하는 방법은 문헌[by J. Sambrook and D. Russell (Molecular Cloning: A Laboratory Manual, 3rd Edition, Cold Spring Harbor Laboratory PRess, Cold Spring Harbor, NY, 2001)]; 문헌[T.J. Silhavy et al. (Experiments with Gene Fusions, Cold Spring Harbor Laboratory PRess, Cold Spring Harbor, NY, 1984)]; 및 문헌[F.M. Ausubel et al. (Short Protocols in Molecular Biology, 5th Ed. Current Protocols, John Wiley and Sons, Inc., NY, 2002)]에 기재된 바와 같은 표준 재조합 DNA 및 분자 클로닝 기술을 따를 수 있다.
본 명세서에서 사용되는 바와 같은 용어 "형질전환"은 숙주 유기체 또는 숙주 세포로의 핵산 분자의 전달을 지칭한다. 예를 들어, 핵산 분자는 세포에서 자율적으로 복제하거나, 숙주 유기체/세포의 게놈에 통합되거나, 또는 복제되거나 통합되지 않고 세포에 일시적으로 존재하는 것일 수 있다. 플라스미드 및 선형 DNA 분자와 같은 형질전환에 적합한 핵산 분자의 비제한적인 예가 본 명세서에 개시되어 있다. 형질전환된 핵산 단편을 함유하는 숙주 유기체/세포 (예를 들어, 본 명세서의 통상적이지 않은 효모)는 "트랜스제닉", "재조합", "형질전환된" 또는 "형질전환체"로 지칭될 수 있다.
폴리뉴클레오티드 또는 폴리펩티드 서열과 관련하여 본 명세서에서 사용되는 바와 같은 용어 "서열 동일성" 또는 "동일성"은 특정 비교창 상에서 최대 상응도로 정렬되는 경우에 동일한 두 서열 내의 핵산 잔기 또는 아미노산 잔기를 지칭한다. 따라서, "서열 동일성 %" 또는 "동일성 %"은 비교창 상에서 최적으로 정렬된 2개의 서열을 비교함으로써 결정된 값을 지칭하며, 여기서 비교창 내의 폴리뉴클레오티드 또는 폴리펩티드 서열의 부분은 2개의 서열의 최적 정렬을 위한 기준 서열 (삽입 또는 결실을 포함하지 않음)과 비교하여 삽입 또는 결실 (즉, 갭)을 포함할 수 있다. 두 서열에서 동일한 핵산 염기 또는 아미노산 잔기가 나타나는 위치의 개수를 결정하여 일치하는 위치의 개수를 산출하고, 일치하는 위치의 개수를 비교창 내의 위치의 총 개수로 나누고, 그 결과에 100을 곱하여 서열 동일성의 백분율을 산출함으로써 백분율을 계산한다. DNA 서열과 RNA 서열 사이의 서열 동일성을 계산하는 경우, DNA 서열의 T 잔기가 RNA 서열의 U 잔기와 정렬되며, 이와 "동일"하게 간주될 수 있음을 이해할 것이다. 제1 폴리뉴클레오티드와 제2 폴리뉴클레오티드의 % 상보성을 결정하기 위해, 예를 들어 (i) 제1 폴리뉴클레오티드와 제2 폴리뉴클레오티드의 보체 서열 사이의 % 동일성 (또는 그 반대) 및/또는 (ii) 표준(canonical) 왓슨과 크릭 염기쌍을 생성할 제1 폴리뉴클레오티드와 제2 폴리뉴클레오티드 사이의 염기 %를 결정함으로써 이를 얻을 수 있다.
예를 들어, 국립 생물공학 정보 센터 (National Center for Biotechnology Information; NCBI) 웹사이트에서 온라인으로 이용가능한 기본 국소 정렬 검색 도구 (Basic Local Alignment Search Tool; BLAST) 알고리즘을 사용하여, 본 명세서에 개시된 둘 이상의 폴리뉴클레오티드 서열 (BLASTN 알고리즘) 또는 폴리펩티드 서열 (BLASTP 알고리즘) 사이의 % 동일성을 측정할 수 있다. 대안적으로, 서열 사이의 % 동일성은 클러스탈(Clustal) 알고리즘 (예를 들어, 클러스탈W 또는 클러스탈V)을 사용하여 수행될 수 있다. 클러스탈 정렬 방법을 사용하는 다중 정렬의 경우, 디폴트(default) 값은 갭 페널티(GAP PENALTY)=10 및 갭 길이 페널티(GAP LENGTH PENALTY)=10에 상응할 수 있다. 클러스탈 방법을 사용하는 단백질 서열의 % 동일성의 계산 및 쌍정렬을 위한 디폴트 파라미터는 케이터플 (KTUPLE)=1, 갭 페널티=3, 윈도우 (WINDOW)=5 및 다이아고날스 세이브드 (DIAGONALS SAVED)=5일 수 있다. 핵산의 경우, 이들 파라미터는 케이터플=2, 갭 페널티=5, 윈도우=4 및 다이아고날스 세이브드=4일 수 있다. 또한 대안적으로, 서열 사이의 % 동일성은 블로섬 (BLOSUM) 매트릭스 (예를 들어, 블로섬62)을 사용하는 갭 오픈(GAP OPEN)=10, 갭 연장(GAP EXTEND)=0.5, 엔드 갭 페널티(END GAP PENALTY)=false, 엔드 갭 오픈=10, 엔드 갭 연장=0.5와 같은 파라미터를 갖는 엠보스(EMBOSS) 알고리즘 (예를 들어, 니들(needle))을 사용하여 수행될 수 있다.
본 명세서에서, 제2 서열에 "상보적인" 제1 서열은, 대안적으로 제2 서열에 대해 "안티센스(antisense)" 배향인 것으로 지칭될 수 있다.
다양한 폴리펩티드 아미노산 서열 및 폴리뉴클레오티드 서열은 개시된 본 발명의 특정 실시 형태의 특징으로서 본 명세서에 개시되어 있다. 본 명세서에 개시된 서열과 적어도 약 70 내지 85%, 85 내지 90% 또는 90% 내지 95% 동일한 이들 서열의 변이체가 사용될 수 있다. 대안적으로, 변이체 아미노산 서열 또는 폴리뉴클레오티드 서열은 본 명세서에 개시된 서열과 적어도 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일성을 가질 수 있다. 변이체 아미노산 서열 또는 폴리뉴클레오티드 서열은 개시된 서열과 동일한 기능/활성, 또는 개시된 서열의 기능/활성의 적어도 약 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%를 갖는다.
본 명세서의 Cas9 단백질의 각각의 아미노산 위치에서의 본 명세서에 개시된 모든 아미노산 잔기는 예이다. 특정 아미노산이 서로 유사한 구조 및/또는 전하 특징을 공유한다고 (즉, 보존된) 고려하면, Cas9의 각 위치에서의 아미노산은 개시된 서열에서 제공된 것과 같거나, 다음과 같이 보존된 아미노산 잔기로 치환될 수 있다 ("보존적 아미노산 치환"):
1.
다음과 같은 작은 지방족의 비극성 또는 약간 극성인 잔기가 서로를 대체할 수 있다: Ala (A), Ser (S), Thr (T), Pro (P), Gly (G);
2.
다음과 같은 극성의 음으로 하전된 잔기 및 그들의 아미드가 서로를 대체할 수 있다: Asp (D), Asn (N), Glu (E), Gln (Q);
3.
다음과 같은 극성의 양으로 하전된 잔기가 서로를 대체할 수 있다: His (H), Arg (R), Lys (K);
4.
다음과 같은 지방족의 비극성 잔기가 서로를 대체할 수 있다: Ala (A), Leu (L), Ile (I), Val (V), Cys (C), Met (M); 및
5.
다음과 같은 큰 방향족 잔기가 서로를 대체할 수 있다: Phe (F), Tyr (Y), Trp (W).
하기 실시예 1에 나타낸 바와 같이, Pol III 프로모터-전사된 gRNA를 사용하여 통상적이지 않은 효모, 예컨대 야로위아 리폴리티카에서 Cas9-매개 DNA 표적화를 수행하는 것이 어려운 것으로 입증되었다. 따라서, Cas9에 대한 RNA 성분을 생성하는 다른 방법은 통상적이지 않은 효모에서 Cas9-매개 DNA 표적화를 제공하는 데 관심이 있다.
개시된 발명의 실시 형태는 5'-캡을 갖지 않는 적어도 하나의 RNA 성분을 포함하는 적어도 하나의 RNA-유도 엔도뉴클레아제 (RGEN)를 포함하는 통상적이지 않은 효모에 관한 것이다. 이러한 캡핑되지 않은 RNA 성분은 효모 내의 염색체 또는 에피솜의 표적 부위 서열에 상보적인 서열을 포함한다. RGEN은 표적 부위 서열의 전부 또는 일부에 결합하고, 임의로 절단할 수 있다.
의미 있게도, RGEN-매개 DNA 표적화는 RGEN 표적 부위 서열과 외인성으로 공급된 공여 DNA 서열 사이의 상동 재조합 (HR) 수준의 증가 또는 삽입-결실 형성에 의해 명시되는 바와 같이 이러한 통상적이지 않은 효모에서 발생한다. 본 발명 이전에, 통상적이지 않은 효모는 일반적으로 HR에 의한 유전자 표적화로 다루기 힘들며, 전형적으로 표적 부위에 랜덤하고 드문 DNA 절단에 의존하여 공여 DNA와 이의 HR을 촉진하였다. 이는 낮은 HR 활성을 가지며 대신에 비상동 말단 결합 (NHEJ) 활성을 선호하는 통상적이지 않은 효모에 기인한다. 따라서, 통상적이지 않은 효모에서 HR에 의한 유전 표적화는 이제 NHEJ 공정보다 HR을 선호하는 S. 세레비시아와 같은 통상적인 효모에서와 같이 실현 가능할 수 있다. 임의의 이론에 구속되기를 바라지는 않지만, 통상적이지 않은 효모 세포에서 5'-캡이 없는 적어도 하나의 RNA 성분을 제공하는 것은 RGEN-매개 DNA 표적화에 참여할 수 있는 핵에서 RNA 성분을 더 잘 축적시킨다고 여겨진다.
Csy4 (Cas6)-기반 RNA 프로세싱 도구와 같은 RNA 프로세싱 도구가 기재되어 있다 (문헌[Nissim et aL. 2014.분자 세포 54:698-710]). Csy4는 예비-crRNA 스템-루프 반복 서열을 결합하고, 이의 동족 기질을 특이적으로 절단하여 반복 서열의 단편에 의해 플랭킹된 스페이서 서열을 포함하는 성숙 crRNA를 생성한다 (문헌[Sternberg et aL. 2012. RNA,18(4):661-72]). 본 명세서에 개시된 (실시예 12)는 5' 캡을 갖지 않는 RNA 성분 (가이드 RNA)을 생성하도록 Csy4를 사용하여 가이드 RNA를 처리하여, 여기서 RNA 성분은 통상적이지 않은 효모의 게놈에서 표적 부위에 결합하여 절단할 수 있는 RGEN을 형성할 수 있다.
본 명세서의 통상적이지 않은 효모는 "통상적인" ("모델") 효모, 예컨대 사카로마이세스 (예를 들어, 출아 효모, 빵 효모 및/또는 맥주 효모로도 알려진 S. 세레비시아) 또는 스키조사카로마이세스 (예를 들어, 분열 효모로도 알려진 S. 폼베(pombe)) 종이 아니다. 소정 실시 형태에서, 통상적인 효모는 NHEJ에 매개되는 수복 공정보다 HR DNA 수복 공정을 선호하는 효모이다.
소정 실시 형태에서, 통상적이지 않은 효모는 HR에 의해 매개되는 수복 공정보다 NHEJ DNA 수복 공정을 선호하는 효모일 수 있다. 통상적인 효모, 예컨대 사카로마이세스 세레비시아 및 스키조사카로마이세스 폼베는 전형적으로 짧은 플랭킹 상동성 암 (30 내지 50 bp)과 일반적으로 70% 초과의 효율로 공여 DNA의 특이적 통합을 나타내는 반면, 통상적이지 않은 효모, 예컨대 피키아 파스토리스(Pichia pastoris), 한세눌라 폴리모르파(Hansenula polymorpha), 야로위아 리폴리티카, 피키아 스티피티스(Pichia stipitis) 및 클루이베로마이세스 락티스(Kluyveromyces lactis)는 보통 유사한 구조의 공여 DNA와 1% 미만의 효율로 특이적 통합을 나타낸다 (문헌[Chen et al., PLoS ONE 8:e57952]). 따라서, HR 공정에 대한 선호도는, 예를 들어, 효모를 적합한 공여 DNA로 형질전환시키고 공여 DNA에 의해 표적화될 것으로 예측되는 게놈 부위와 특이적으로 재조합되는 정도를 측정함으로써 측정될 수 있다. 예를 들어, 이러한 검정이 효모 게놈에서 공여 DNA의 고도의 랜덤 통합을 산출하는 경우, NHEJ에 대한 선호도 (또는 HR에 대한 낮은 선호도)가 명백할 것이다. 효모에서 DNA의 특이적 (HR-매개) 및/또는 랜덤 (NHEJ-매개) 통합의 비율을 측정하기 위한 검정이 본 기술 분야에 알려져 있다 (예를 들어, 문헌[Ferreira and Cooper, Genes Dev. 18:2249-2254]; 문헌[Corrigan et al., PLoS ONE 8:e69628]; 문헌[Weaver et al., ProC. NatL. AcaD. Sci. U.S.A. 78:6354-6358]; 문헌[Keeney and Boeke, Genetics 136:849-856]).
낮은 수준의 HR 활성을 고려하면, 본 명세서의 통상적이지 않은 효모는 (i) 예를 들어, 약 1%, 2%, 3%, 4%, 5%, 6%, 7% 또는 8% 미만의 30 내지 50 bp의 플랭킹 상동성 암을 갖는 적합한 공여 DNA에 의한 특이적인 표적화 비율을 나타내고/내거나 (ii) 예를 들어, 약 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74% 또는 75% 초과의 상기 공여 DNA의 랜덤 통합 비율을 나타낼 수 있다. 적합한 공여 DNA의 (i) 특이적 표적화 및/또는 (ii) 랜덤 통합의 이러한 비율은 본 명세서에 개시된 바와 같은 RGEN이 제공되기 전에 존재하는 그대로의 통상적이지 않은 효모를 특성화할 수 있다. 소정 실시 형태에서, 통상적이지 않은 효모에 RGEN을 제공하는 목적은 특정 부위에서 효모를 HR쪽으로 편향시키기 위한 부위 특이적 DNA 단일 가닥 절단 (SSB) 또는 이중 가닥 절단 (DSB)을 생성하는 것이다. 따라서, 본 명세서의 적합한 RGEN을 포함하는 통상적이지 않은 효모는 전형적으로 특정 공여 DNA와의 HR 비율의 증가를 나타내야 한다. 이러한 증가된 비율은 적합한 대조 (예를 들어, 동일한 공여 DNA로 형질전환되었지만, 적합한 RGEN가 결여된 동일한 통상적이지 않은 효모)에서의 HR 비율보다 적어도 약 2-, 3-, 4-, 5-, 6-, 7-, 8-, 9- 또는 10-배 더 높을 수 있다.
본 명세서의 소정 측면에서, 통상적이지 않은 효모는 무성으로 (무성생식형(anamorphic)) 또는 유성으로 (유성생식형(teleomorphic)) 생식할 수 있다. 본 명세서의 통상적이지 않은 효모는 전형적으로 단세포 형태로 존재하지만, 이들 효모의 특정 유형은 임의로 가성균사(pseudohyphae) (연결된 발아 세포의 줄)를 형성할 수 있다. 또 다른 측면에서, 통상적이지 않은 효모는 반수체 또는 이배체일 수 있고/있거나 이들 배수성 형태 중 어느 하나로 존재하는 능력을 가질 수 있다.
본 명세서의 통상적이지 않은 효모는 이들 모두가 본 명세서에 참고로 포함되는 문헌[Non-Conventional Yeasts in Genetics, Biochemistry and Biotechnology: Practical Protocols (K. Wolf, K.D. Breunig, G. Barth, Eds., Springer-Verlag, Berlin, Germany, 2003)], 문헌[Yeasts in Natural and Artificial Habitats (J.F.T. Spencer, D.M. Spencer, Eds., Springer-Verlag, Berlin, Germany, 1997)], 및/또는 문헌[Yeast Biotechnology: Diversity and Applications (T. Satyanarayana, G. Kunze, Eds., Springer, 2009)]에 기재된 바와 같은 본 기술 분야에 알려진 임의의 방법에 따라 배양될 수 있다.
본 명세서의 통상적이지 않은 효모의 비제한적인 예는 다음과 같은 속의 효모를 포함한다: 야로위아, 피키아, 쉬반니오마이세스, 클루이베로마이세스, 아르술라, 트리코스포론, 칸디다, 유스틸라고, 토룰롭시스, 자이고사카로마이세스, 트리고놉시스, 크립토코커스, 로도토룰라, 파피아, 스포로볼로마이세스 및 파치솔렌. 야로위아 종의 적합한 예는 야로위아 리폴리티카이다. 피키아 종의 적합한 예에는 P. 파스토리스, P. 메타놀리카(methanolica), P. 스티피티스, P. 아노말라(anomala) 및 P. 앙구스타(angusta)가 포함된다. 쉬반니오마이세스 종의 적합한 예에는 S. 카스텔리이(castellii), S. 알루비우스(alluvius), S. 호미니스 (hominis), S. 옥시덴탈리스(occidentalis), S. 카프리오티이(capriottii), S. 에트첼시이(etchellsii), S. 폴리모르푸스(polymorphus), S. 슈도폴리모르푸스(pseudopolymorphus), S. 반리자에(vanrijiae) 및 S. 야마다에(yamadae)가 포함된다. 루이베로마이세스 종의 적합한 예에는 K. 락티스, K. 마르시아누스(marxianus), K. 프라길리스(fragilis), K. 드로소필라룸(drosophilarum), K. 써모톨레란스(thermotolerans), K. 파세올로스포루스(phaseolosporus), K. 바누데니이(vanudenii), K. 왈티이(waltii), K. 아프리카누스(africanus) 및 K. 폴리스포루스(polysporus)가 포함된다. 아르술라 종의 적합한 예에는 A. 아데니니보란스(adeninivorans) 및 A. 테레스트레(terRestre)가 포함된다. 트리코스포론 종의 적합한 예에는 T. 쿠타네움(cutaneum), T. 카피타툼(capitatum), T. 인킨(inkin) 및 T. 비메리(beemeri)가 포함된다. 칸디다 종의 적합한 예에는 C. 알비칸스(albicans), C. 아스칼라피다움(ascalaphidarum), C. 암피시애(amphixiae), C. 앤트아티카(antarctica), C. 아르젠티아(argentea), C. 아틀란티카(atlantica), C. 아트모스패리카(atmosphaerica), C. 블랏타에(blattae), C. 브로멜리아세아룸(bromeliacearum), C. 카르포필리아(carpophila), C. 카르바잘리스(carvajalis), C. 세람비시다룸(cerambycidarum), C. 차우리오데스(chauliodes), C. 코리달리(corydali), C. 도스세이(dosseyi), C. 듀블리니엔시스(dubliniensis), C. 얼가텐시스(ergatensis), C. 프룩투스(fructus), C. 글라브라타(glabrata), C. 퍼멘타티(fermentati), C. 귈리어몬디이(guilliermondii), C. 해뮬로니이(haemulonii), C. 인섹타멘스(insectamens), C. 인섹토룸(insectorum), C. 인터메디아(intermedia), C. 제프레시이(jeffResii), C. 케피르(kefyr), C. 케로세네아에(keroseneae), C. 크루세이(krusei), C. 루시타니애(lusitaniae), C. 릭소소필리아(lyxosophila), C. 말토사(maltosa), C. 마리나(marina), C. 멤브라니파시엔스(membranifaciens), C. 밀러리(milleri), C. 모기이(mogii), C. 올레필리아(oleophila), C. 오레고네시스(oregonensis), C. 파라프실로시스(parapsilosis), C. 퀴어시트루사(quercitrusa), C. 루고사(rugosa), C. 사케(sake), C. 쉐하테아(shehatea), C. 템노칠라에(temnochilae), C. 테누이스(tenuis), C. 테아에(theae), C. 톨러란스(tolerans), C. 트로피칼리스(tropicalis), C. 츠시이아에(tsuchiyae), C. 시놀라보란티움(sinolaborantium), C. 소자에(sojae), C. 수브하쉬이(subhashii), C. 비스와나치이(viswanathii), C. 유틸리스(utilis), C. 우바투벤시스(ubatubensis) 및 C. 젬플리니나(zemplinina)가 포함된다. 유스틸라고 종의 적합한 예에는 U. 아베나에(avenae), U. 에스쿨렌타(esculenta), U. 호르데이(hordei), U. 마이디스(maydis), U. 누다(nuda) 및 U. 트리티치(tritici)가 포함된다. 토룰롭시스 종의 적합한 예에는 T. 게오차레스(geochaRes), T. 아지마(azyma), T. 글라브라타(glabrata) 및 T. 칸디다가 포함된다. 자이고사카로마이세스 종의 적합한 예에는 Z. 바일리이(bailii), Z. 비스포루스(bisporus), Z. 시드리(cidri), Z. 퍼멘타티(fermentati), Z. 플루오렌티누스(florentinus), Z. 콤부차엔시스(kombuchaensis), Z. 렌투스(lentus), Z. 멜리스(mellis), Z. 미크로엘리프소이데스(microellipsoides), Z. 엠라키이(mrakii), Z. 슈도로추시이(pseudorouxii) 및 Z. 로우시이가 포함된다. 트리고놉시스 종의 적합한 예에는 T. 바리아빌리스(variabilis)가 포함된다. 크립토코커스 종의 적합한 예에는 C. 라우렌티이(laurentii), C. 알비두스(albidus), C. 네오포르만스(neoformans), C. 가티이(gattii), C. 유니구툴라투스 (uniguttulatus), C. 아델리엔시스(adeliensis), C. 아에리우스(aerius), C. 알비도시밀리스(albidosimilis), C. 안타르크티쿠스(antarcticus), C. 아쿠아티쿠스(aquaticus), C. 아테르(ater), C. 부타네시스(bhutanensis), C. 콘소르티오니스(consortionis), C. 쿠르바투스(curvatus), C. 페놀리쿠스(phenolicus), C. 스킨네리(skinneri), C. 테레우스(terreus) 및 C. 비쉬니마치(vishniacci)가 포함된다. 로도토룰라 종의 적합한 예에는 R. 아체니오룸(acheniorum), R. 툴라(tula), R. 아쿠타(acuta), R. 아메리카나(americana), R. 아라우차리아에(araucariae), R. 아르티카(arctica), R. 아르메니아카(armeniaca), R. 아우란티아카(aurantiaca), R. 아우리쿨라리아에(auriculariae), R. 바카룸(bacarum), R. 벤티카(benthica), R. 비오우르게이(biourgei), R. 보고리엔시스(bogoriensis), R. 브론치알리스(bronchialis), R. 부포니이(buffonii), R. 칼립토게나에(calyptogenae), R. 충나멘시스(chungnamensis), R. 클란디엔시스(cladiensis), R. 코랄리나(corallina), R. 크레솔리카(cResolica), R. 크로세아(crocea), R. 사이클로클라스티카(cycloclastica), R. 다이레넨시스(dairenensis), R. 디플루덴스(diffluens), R. 에베르글라디엔시스(evergladiensis), R. 페룰리카(ferulica), R. 폴리오룸(foliorum), R. 프라가리아(fragaria), R. 푸지사넨시스(fujisanensis), R. 푸트로넨시스(futronensis), R. 겔라티노사(gelatinosa), R. 글라시알리스(glacialis), R. 글루티니스(glutinis), R. 그라실리스(gracilis), R. 그라미니스(graminis), R. 그린베르그시이(grinbergsii), R. 히말라옌시스(himalayensis), R. 힌눌레아(hinnulea), R. 히스톨리티카(histolytica), R. 힐로필리아(hylophila), R. 인카르나타(incarnata), R. 인게니오사(ingeniosa), R. 자바니카(javanica), R. 코이쉬카웬시스(koishikawensis), R. 락토사(lactosa), R. 라멜리브라치아에(lamellibrachiae), R. 라린기스(laryngis), R. 리그노필라(lignophila), R. 리니(lini), R. 론기스시마(longissima), R. 루드위기이(ludwigii), R. 리시노필라(lysinophila), R. 마리나(marina), R. 마르티니아에-프라간티스(martyniae -fragantis), R. 마트리텐시스(matritensis), R. 멜리(meli), R. 미누타(minuta), R. 무실라기노사(mucilaginosa), R. 니텐스(nitens), R. 노쏘파기(nothofagi), R. 오리자에(oryzae), R. 파시피카(pacifica), R. 팔리다(pallida), R. 페네아우스(peneaus), R. 필릴라(philyla), R. 필로플라나(phylloplana), R. 필라티이(pilatii), R. 필리마나에(pilimanae), R. 피니콜라(pinicola), R. 플리카타(plicata), R. 폴리모르파(polymorpha), R. 사이츠로페놀리카(psychrophenolica), R. 사이츠로필라(psychrophila), R. 푸스툴라(pustula), R. 레티노필라(retinophila), R. 로사세아(rosacea), R. 로술라타(rosulata), R. 루베파시엔스(rubefaciens), R. 루벨라(rubella), R. 루베센스(rubescens), R. 루브라(rubra), R. 루브로루고사(rubrorugosa), R. 루풀라(rufula), R. 루틸라(rutila), R. 산구이네아(sanguinea), R. 산니에이(sanniei), R. 사르토리이(sartoryi), R. 실베스트리스(silvestris), R. 실플렉스(simplex), R. 시넨시스(sinensis), R. 슬루피아에(slooffiae), R. 순키이(sonckii), R. 스트라미네아(straminea), R. 수베리콜라(subericola), R. 수가니이(suganii), R. 타이와넨시스(taiwanensis), R. 타이와니아나(taiwaniana), R. 테르페노이달리스(terpenoidalis), R. 테레아(terrea), R. 텍센시스(texensis), R. 토쿄엔시스(tokyoensis), R. 울자마에(ulzamae), R. 바닐리카(vanillica), R. 부일레미니이(vuilleminii), R. 야로위이, R. 윤나넨시스(yunnanensis ) 및 R. 졸티이(zsoltii)가 포함된다. 파피아 종의 적합한 예에는 P. 로도지마(rhodozyma)가 포함된다. 스포로볼로마이세스 종의 적합한 예에는 S. 알보루베센스(alborubescens), S. 반나엔시스(bannaensis), S. 베이징엔시스(beijingensis), S. 비스초리아에(bischofiae), S. 클라바투스(clavatus), S. 코프로스마에(coprosmae), S. 코프로미콜라(coprosmicola), S. 코랄리누스(corallinus), S. 디메나에(dimmenae), S. 드라코필리(dracophylli), S. 에론가투스(elongatus), S. 그라실리스(gracilis), S. 이노시토필루스(inositophilus), S. 존소니이(johnsonii), S. 코알라에(koalae), S. 마그니스포루스(magnisporus), S. 노보제알란이쿠스(novozealandicus), S. 오도루스(odorus), S. 파타고니쿠스(patagonicus), S. 프로덕투스(productus), S. 로세우스(roseus), S. 사시콜라(sasicola), S. 쉴바타누스(shibatanus), S. 신굴라이스(singularis), S. 수브룬네우스(subbrunneus), S. 시메트리쿠스(symmetricus), S. 시지기이(syzygii), S. 타우포엔시스(taupoensis), S. 수가에(tsugae), S. 잔투스(xanthus) 및 S. 윤나넨시스가 포함된다. 파치솔렌 종의 적합한 예에는 P. 탄노필루스(tannophilus)가 포함된다.
야로위아 리폴리티카는 본 명세서에 개시된 소정 실시 형태에서 바람직하다.
적합한 야로위아 리폴리티카의 예에는 다음과 같은 아메리칸 타입 컬쳐 컬렉션(American Type Culture Collection) (버지니아주 매나사스 소재의 ATCC)로부터 입수가능한 분리주가 포함된다: 균주명 ATCC #20362, #8862, #8661, #8662, #9773, #15586, #16617, #16618, #18942, #18943, #18944, #18945, #20114, #20177, #20182, #20225, #20226, #20228, #20327, #20255, #20287, #20297, #20315, #20320, #20324, #20336, #20341, #20346, #20348, #20363, #20364, #20372, #20373, #20383, #20390, #20400, #20460, #20461, #20462, #20496, #20510, #20628, #20688, #20774, #20775, #20776, #20777, #20778, #20779, #20780, #20781, #20794, #20795, #20875, #20241, #20422, #20423, #32338, #32339, #32340, #32341, #34342, #32343, #32935, #34017, #34018, #34088, #34922, #34922, #38295, #42281, #44601, #46025, #46026, #46027, #46028, #46067, #46068, #46069, #46070, #46330, #46482, #46483, #46484, #46436, #60594, #62385, #64042, #74234, #76598, #76861, #76862, #76982, #90716, #90811, #90812, #90813, #90814, #90903, #90904, #90905, #96028, #201241, #201242, #201243, #201244, #201245, #201246, #201247, #201249 및/또는 #201847.
본 명세서 임의의 다른 통상적이지 않은 효모뿐만 아니라 야로위아 리폴리티카는 유지성일 수 있고/있거나 (예를 들어, 이의 건조 세포 중량의 적어도 25%를 오일로서 생성함), 하나 이상의 다중불포화 지방산 (예를 들어, 오메가-6 또는 오메가-3)을 생성할 수 있다. 이러한 유지성은 효모가 이의 야생형 형태와 비교하여 증가된 양의 지질을 생산하도록 유전적으로 조작된 결과일 수 있다. 유지성 야로위아 리폴리티카 균주의 예가 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2009/0093543호, 제2010/0317072호, 제2012/0052537호 및 제2014/0186906호에 개시되어 있다.
통상적이지 않은 효모에 대해 본 명세서에 개시된 실시 형태는 또한 다른 미생물, 예컨대 진균류에 적용될 수 있다. 소정 실시 형태에서, 진균류는 HR에 의해 매개되는 수복 공정보다 NHEJ DNA 수복 공정을 선호하는 진균류일 수 있다. 본 명세서의 진균은 담자균(Basidiomycetes), 접합균(Zygomycetes), 호상균(Chytridiomycetes) 또는 자낭균 진균류일 수 있다. 본 명세서의 사상진균의 예에는 트리코데르마, 크리소스포리움(Chrysosporium), 티엘라비아(Thielavia), 뉴로스포라(Neurospora) (예를 들어, N. 크라사(crassa), N. 시토실라(sitophila)), 크리포넥트리아(Cryphonectria) (예를 들어, C. 파라시티카(parasitica)), 아우레오바시디움(Aureobasidium) (예를 들어, A. 풀루란스(pullulans)), 필리바시디움(Filibasidium), 피로마이세스(Piromyces), 크립토콕쿠스(Cryplococcus), 아크레모니움(Acremonium), 톨리포클라디움(Tolypocladium), 신탈리디움(Scytalidium), 스키조필룸(Schizophyllum), 스포로트리쿰(Sporotrichum), 페니실리움(Penicillium) (예를 들어, P. 빌라이아에(bilaiae), P. 카멤베르티(camemberti), P. 칸디둠(candidum), P. 크리소게눔(chrysogenum), P. 엑스펜숨(expansum), P. 퍼니쿨로섬(funiculosum), P. 글라우컴(glaucum), P. 마네페이(marneffei), P. 로커포르티(roqueforti), P. 베르루코숨(verrucosum), P. 비리디카툼(viridicatum)), 기베렐라(Gibberella) (예를 들어, G. 아쿠미나타(acuminata), G. 아베나세아(avenacea), G. 바카타(baccata), G. 키르키나타(circinata), G. 시아노게나(cyanogena), G. 푸지쿠로이(fujikuroi), G. 인트리칸스(intricans), G. 풀리카리스(pulicaris), G. 스틸보이데스(stilboides), G. 트리킨크타(tricincta), G. 제에(zeae)), 미셀리오프토라(Myceliophthora), 뮤코(Mucor) (예를 들어, M. 로우시(rouxii), M. 시르시넬로이데스(circinelloides)), 아스퍼질러스(Aspergillus) (예를 들어, A. 니거(niger), A. 오리자에(oryzae), A. 니둘란스(nidulans), A. 플라버스(flavus), A. 렌툴루스(lentulus), A. 테레우스, A. 클라바투스, A. 푸미가투스(fumigatus)), 푸사리움(Fusarium) (예를 들어, F. 그라미네아룸(graminearum), F. 옥시스포룸(oxysporum), F. 부비게눔(bubigenum), F. 솔라니(solani), F. 옥시스포룸, F. 버티실리오이데스(verticillioides), F. 프로리페라툼(proliferatum), F. 베네나툼(venenatum)) 및 후미콜라(Humicola) 속의 것들 및 이들의 무성생식형과 유성생식형이 포함된다. 본 명세서에 진균류의 속 및 종은 문헌[Barnett and Hunter (Illustrated Genera of Imperfect Fungi, 3rd Edition, Burgess Publishing Company, 1972)]에 개시된 바와 같은 형태에 의해 원하는 경우 정의될 수 있다. 진균은 임의로 해충/병원균, 예컨대 동물 (예를 들어, 인간)의 해충/병원균으로 특징지어질 수 있다.
본 명세서의 소정 측면에서 트리코데르마 종에는 T. 아그레시범(aggRessivum), T. 아마조니쿰(amazonicum), T. 아스페렐룸(asperellum), T. 아트로비리데(atroviride), T. 아우레오비리데(aureoviride), T. 아우스트로코닌기이(austrokoningii), T. 브레비콤팩툼(brevicompactum), T. 칸디둠, T. 카리 배움(caribbaeum), T. 카톱트론(catoptron), T. 크레메움(cremeum), T. 세라미쿰(ceramicum), T. 세리넘(cerinum), T. 클로로스포룸(chlorosporum), T. 크로모스페르뭄(chromospermum), T. 신나모메움(cinnamomeum), T. 시트리노비리데 (citrinoviride), T. 크라숨(crassum), T. 크레메움(cremeum), T. 딩글레예아(dingleyeae), T. 도로테아에(dorotheae), T. 에푸숨(effusum), T. 에리나세움(erinaceum), T. 에스토니쿰(estonicum), T. 페르틸레(fertile), T. 겔라티노수스(gelatinosus), T. 하넨세(ghanense), T. 하마툼(hamatum), T. 하르지아눔(harzianum), T. 헬리쿰(helicum), T. 인트리칸스, T. 코니랑브라(konilangbra), T. 코닌기이(koningii), T. 코닌지옵시스(koningiopsis), T. 론지브라치아툼(longibrachiatum), T. 론지필레(longipile), T. 미누티스포룸(minutisporum), T. 오블론지스포룸(oblongisporum), T. 오발리스포룸(ovalisporum), T. 페테르세니(petersenii), T. 필로스타히디스(phyllostahydis), T. 필루리페룸 (piluliferum), T. 플레우로티콜라(pleuroticola), T. 플레우로툼(pleurotum), T. 폴리스포룸(polysporum), T. 슈도코닌기(pseudokoningii), T. 푸베센스(pubescens), T. 레세이(reesei), T. 로게르소니(rogersonii), T. 로시쿰(rossicum), T. 사투르니스포룸(saturnisporum), T. 시넨시스(sinensis), T. 시누오숨(sinuosum), T. 스피랄레(spirale), T. 스트라미네움(stramineum), T. 스트리고숨(strigosum), T. 스트로마티쿰(stromaticum), T. 수로툰둠(surrotundum), T. 타이와넨세(taiwanense), T. 타일랜디쿰(thailandicum), T. 텔레포리콜룸(thelephoricolum), T. 테오브로미콜라(theobromicola), T. 토멘토숨(tomentosum), T. 벨루티눔(velutinum), T. 베렌스(virens), T. 비리데(viride) 및 T. 비리데넨스(viridescens)가 포함된다. 본 명세서의 트리코데르마 종은, 예를 들어, 본 명세서에 참고로 포함되는 문헌[Trichoderma: Biology and Applications (P.K. Mukherjee et al., Eds., CABI, Oxfordshire, UK, 2013)]에 기재된 바와 같이 배양되고/거나 조작될 수 있다.
소정 실시 형태에서, 미생물 세포는 조류 세포이다. 예를 들어, 조류 세포는 다음 중 어느 하나의 유래일 수 있다: 녹조류(green algae), 홍조류(red algae), 갈조류(brown algae), 규조류(diatoms) 및 와편모충류 (와편모조류). 다른 측면에서, 조류 세포는 미세조류 (예를 들어, 식물성 플랑크톤, 미소식물 또는 플랑크톤 조류) 또는 거대조류 (켈프, 해초)일 수 있다. 추가의 예로서, 본 명세서의 조류 세포는 포프리라(Porphyra) (김), 팔마리아(Palmaria) 종, 예컨대 P. 팔마타(palmata) (덜스(dulse)), 아르트로스피라(Arthrospira) 종, 예컨대 A. 플라텐시스(platensis) (스피룰리나(spirulina)), 클로렐라(Chlorella) (예를 들어, C. 프로토테코이데스(protothecoides)), 콘드루스(Chondrus) 종, 예컨대 C. 크리스푸스(crispus) (아이리쉬 모스(Irish moss)), 아파니조메논(Aphanizomenon), 사르가쑴(Sargassum), 코차유요(Cochayuyo), 보트리코쿠스(Botryococcus) (예를 들어, B. 브라우니이(braunii)), 두날리엘라(Dunaliella) (예를 들어, D. 테르티올렉타(tertiolecta), 그라실라리아(Gracilaria), 플레우로크리시스(Pleurochrysis) (예를 들어, P. 카르테아에(carterae)), 안키스트로데스무스(Ankistrodesmus), 시클로텔라(Cyclotella), 한츠슈이아(Hantzschia), 난노클로리스(Nannochloris), 난노클로롭시스(Nannochloropsis), 니츠키아(Nitzschia), 패오닥틸룸(Phaeodactylum) (예를 들어, P. 트리코누툼(tricornutum)), 세네데스무스(Scenedesmus), 스티코코커스(Stichococcus), 테트라셀미스(Tetraselmis) (예를 들어, T. 수에시카(suecica)), 탈라시오시라(Thalassiosira) (예를 들어, T. 슈도나나((pseudonana)), 크립테코디니움(Crypthecodinium) (예를 들어, C. 코흐니(cohnii)), 네오클로리스(Neochloris) (예를 들어, N. 올레오아분단스(oleoabundans)) 또는 치오키트리움(Schiochytrium)일 수 있다. 본 명세서의 조류 종은, 예를 들어, 본 명세서에 참고로 포함되는 문헌[Thompson (Algal Cell CulturE. Encyclopedia of Life Support System (EOLSS), Biotechnology Vol 1, available at eolss.net/sample-chapters internet site)]에 기재된 바와 같이 배양되고/거나 조작될 수 있다.
본 명세서의 5'-캡을 갖지 않는 적어도 하나의 RNA 성분을 포함하는 적어도 하나의 RGEN을 포함하는 통상적이지 않은 효모는 자연계에서 발생하지 않는다. 임의의 특정 이론에 구애되고자 함이 없이, 본 명세서의 RGEN이, 예를 들어 원핵생물에서만 발생하는 것으로 알려져왔기 때문에 이러한 효모는 자연적으로 발생하지 않는 것으로 여겨진다. 또한, 여겨진다 효모의 소정 실시 형태는 crRNA와 tracrRNA의 이종 결합을 나타내는 gRNA를 포함하는 RNA 성분을 갖는 RGEN을 포함함으로 인해 자연적으로 발생하지 않는다.
본 명세서에서 RGEN은 적어도 하나의 Cas 단백질과 적어도 하나의 RNA 성분을 포함하는 복합체를 지칭한다. 적합한 Cas 단백질의 예에는 하나 이상의 타입 I, II 또는 III 크리스퍼 시스템의 Cas 엔도뉴클레아제가 포함된다 (본 명세서에 참고로 포함되는 문헌[(Bhaya et al., Annu. Rev. GeneT. 45:273-297]). 타입 I 크리스퍼 Cas 단백질은, 예를 들어 Cas3 또는 Cas4 단백질일 수 있다. 타입 II 크리스퍼 Cas 단백질은, 예를 들어 Cas9 단백질일 수 있다. 타입 III 크리스퍼 Cas 단백질은, 예를 들어 Cas10 단백질일 수 있다. Cas9 단백질이 바람직한 실시 형태에서 사용된다. 소정 실시 형태에서, Cas 단백질은 박테리아 또는 고세균 단백질일 수 있다. 본 명세서에서, 타입 I 내지 타입 III 크리스퍼 Cas 단백질은 전형적으로 원핵생물 기원이며; 예를 들어, 타입 I 및 타입 III Cas 단백질은 박테리아 또는 고세균 종으로부터 유래될 수 있는 반면, II Cas 단백질은 (즉, Cas9) 박테리아 종으로부터 유래될 수 있다. 다른 실시 형태에서, 적합한 Cas 단백질은 Cas1, Cas1B, Cas2, Cas3, Cas4, Cas5, Cas6, Cas7, Cas8, Cas9, Cas10, Csy1, Csy2, Csy3, Cse1, Cse2, Csc1, Csc2, Csa5, Csn2, Csm2, Cs㎥, Csm4, Csm5, Csm6, Cmr1, Cmr3, Cmr4, Cmr5, Cmr6, Csb1, Csb2, Csb3, Csx17, Csx14, Csx10, Csx16, CsaX, Csx3, Csx1, Csx15, Csf1, Csf2, Csf3, Csf4, 이들의 상동체 또는 이들의 변형된 버전 중 하나 이상을 포함한다.
개시된 발명의 다른 측면에서, 본 명세서의 Cas 단백질은 다음의 속 중 어느 하나의 유래일 수 있다: 아에로피룸(Aeropyrum), 피로바쿨룸(Pyrobaculum), 술폴로부스(Sulfolobus), 아캐오글로부스( Archaeoglobus ), 할로카르쿨라( Haloarcula ), 메타노박테리움(Methanobacteriumn), 메타노코커스( Methanococcus ), 메타노사르시나(Methanosarcina), 메타노피러스( Methanopyrus ), 피로코커스( Pyrococcus ), 피크로필러스(Picrophilus), 써모플라스마( Thernioplasnia ), 코리네박테리움(Corynebacterium), 마이코박테리움(Mycobacterium), 스트렙토마이세스(Streptomyces), 아퀴펙스( Aquifrx ), 포르피로모나스( Porphvromonas ), 클로로비움(Chlorobium), 써머스( Thermus ), 바실러스, 리스테리아( Listeria ), 스타필로코커스 (Staphylococcus), 클로스트리디움(Clostridium), 써모아나에로박터(Thermoanaerobacter), 마이코플라스마( Mycoplasma ), 푸소박테리움(Fusobacterium), 아자쿠스( Azarcus ), 크로모박테리움( Chromobacterium ), 네이세리아(Neisseria), 니트로소모나스( Nitrosomonas ), 디설포비브리오( Desulfovibrio ), 게오박터(Geobacter), 믹소코커스(Myrococcus), 캄필로박터(Campylobacter), 볼리넬라(Wolinella), 아시네토박터(Acinetobacter), 에르위니아(Erwinia), 에스케리키아(Escherichia), 레지오넬라(Legionella), 메틸로코커스(Methylococcus), 파스퇴렐라(Pasteurella), 포토박테리움(Photobacterium), 살모넬라(Salmonella), 잔토모나스(Xanthomonas), 예르시니아(Yersinia), 스트렙토코커스, 트레포네마(Treponema), 프란시셀라(Francisella) 또는 써모토가(Thermotoga). 대안적으로, 본 명세서의 Cas 단백질은, 예를 들어, 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010/0093617호에 개시된 바와 같이 서열 번호:462-465, 467-472, 474-477, 479-487, 489-492, 494-497, 499-503, 505-508, 510-516 또는 517-521 중 어느 하나에 의해 암호화될 수 있다.
소정 실시 형태에서 RGEN은 Cas9 아미노산 서열을 포함한다. 본 명세서의 Cas9 단백질의 아미노산 서열과 본 명세서의 소정 다른 Cas 단백질은, 예를 들어, 스트렙토코커스 (예를 들어, S. 피오제네스, S. 뉴모니애(pneumoniae), S. 써모필러스(thermophilus), S. 아갈락티아(agalactiae), S. 파라상귀니스(parasanguinis), S. 오랄리스(oralis), S. 살리바리우스(salivarius), S. 마카카(macacae), S. 디스갈락티애(dysgalactiae), S. 안지노서스(anginosus), S. 콘스텔라투스(constellatus), S. 슈도포르시누스(pseudoporcinus), S. 뮤탄스(mutans)), 리스테리아 (예를 들어, L. 인노쿠아(innocua)), 스피로플라즈마(Spiroplasma) (예를 들어, S. 아피스(apis), S. 시르피디콜라(syrphidicola)), 펩토스트렙토코카세(Peptostreptococcaceae), 아토포비움(Atopobium), 포르피로모나스(Porphyromonas) (예를 들어, P. 카토니아(catoniae)), 프레보텔라(Prevotella) (예를 들어, P. 인테르메디아(intermedia)), 베일로넬라(Veillonella), 트레포네마(Treponema) (예를 들어, T. 소크란스키(socranskii), T. 티콜라(denticola)), 카프노사이토파가(Capnocytophaga), 피네골디아(Finegoldia) (예를 들어, F. 마그나(magna)), 코리오박테리아세아에(Coriobacteriaceae) (예를 들어, C. 박테리움(bacterium)), 올스넬라(Olsenella) (예를 들어, O. 프로푸사(profusa)), 헤모필루스(Haemophilus) (예를 들어, H. 스푸토룸(sputorum), H. 피트마니아에(pittmaniae)), 파스튜렐라(Pasteurella) (예를 들어, P. 베티아에(bettyae)), 올리비박터(Olivibacter) (예를 들어, O. 시티엔시스(sitiensis)), 에필리토니모나스(Epilithonimonas) (예를 들어, E. 테낙스(tenax)), 메소니아(Mesonia) (예를 들어, M. 모빌리스(mobilis)), 락토바실루스(Lactobacillus) (예를 들어, L. 플란타룸(plantarum)), 바실루스 (예를 들어, B. 세레우스(cereus)), 아퀴마리나(Aquimarina) (예를 들어, A. 무엘레리(muelleri)), 크리세오박테리(Chryseobacterium) (예를 들어, C. 파루스트레(palustre)), 박테로이데스(Bacteroides) (예를 들어, B. 그라미니솔벤스(graminisolvens)), 네이세리아 (예를 들어, N. 메닝기티디스(meningitidis)), 프란시셀라(Francisella) (예를 들어, F. 노비시다(novicida)) 또는 플라보박테리움(Flavobacterium) (예를 들어, F. 프리기다리움(frigidarium), F. 솔리(soli)) 종으로부터 유래될 수 있다. 본 명세서의 소정 측면에서 S. 피오제네스 Cas9가 바람직하다. 또 다른 예로서, Cas9 단백질은 본 명세서에 참고로 포함되는 문헌[Chylinski et aL. (RNA Biology 10:726-737)]에 개시된 Cas9 단백질 중 어느 하나 일 수 있다.
따라서, 본 명세서의 Cas9 단백질의 서열은, 예를 들어, 참고로 포함되는 진뱅크(GenBank) 기탁 번호 G3ECR1 (S. 써모필러스), WP_026709422, WP_027202655, WP_027318179, WP_027347504, WP_027376815, WP_027414302, WP_027821588, WP_027886314, WP_027963583, WP_028123848, WP_028298935, Q03JI6 (S. 써모필러스), EGP66723, EGS38969, EGV05092, EHI65578 (S. 슈도포르시누스), EIC75614 (S. 오랄리스), EID22027 (S. 콘스텔라투스), EIJ69711, EJP22331 (S. 오랄리스), EJP26004 (S. 안지노서스), EJP30321, EPZ44001 (S. 피오제네스), EPZ46028 (S. 피오제네스), EQL78043 (S. 피오제네스), EQL78548 (S. 피오제네스), ERL10511, ERL12345, ERL19088 (S. 피오제네스), ESA57807 (S. 피오제네스), ESA59254 (S. 피오제네스), ESU85303 (S. 피오제네스), ETS96804, UC75522, EGR87316 (S. 디스갈락티애), EGS33732, EGV01468 (S. 오랄리스), EHJ52063 (S. 마카카), EID26207 (S. 오랄리스), EID33364, EIG27013 (S. 파라상귀니스), EJF37476, EJO19166 (스트렙토코커스 종 BS35b), EJU16049, EJU32481, YP_006298249, ERF61304, ERK04546, ETJ95568 (S. 아갈락티아), TS89875, ETS90967 (스트렙토코커스 종 SR4), ETS92439, EUB27844 (스트렙토코커스 종 BS21), AFJ08616, EUC82735 (스트렙토코커스 종 CM6), EWC92088, EWC94390, EJP25691, YP_008027038, YP_008868573, AGM26527, AHK22391, AHB36273, Q927P4, G3ECR1 또는 Q99ZW2 (S. 피오제네스),)에 개시된 Cas9 아미노산 서열 중 어느 하나를 포함할 수 있다. 이러한 Cas9 단백질 서열의 임의의 변이체가 사용될 수 있지만, 본 명세서의 RNA 성분과 결합할 경우 DNA에 대한 특이적 결합 활성 및 임의로 엔도뉴클레오리틱 활성을 가져야 한다. 이러한 변이체는 참고 Cas9의 아미노산 서열과 적어도 약 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일한 아미노산 서열을 포함할 수 있다.
대안적으로, 본 명세서의 Cas9 단백질은, 예를 들어, 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010/0093617호에 개시된 바와 같은 서열 번호:462 (S. 써모필러스), 474 (S. 써모필러스), 489 (S. 아갈락티아), 494 (S. 아갈락티아), 499 (S. 뮤탄스), 505 (S. 피오제네스) 또는 518 (S. 피오제네스) 중 어느 하나에 의해 암호화될 수 있다. 또한 대안적으로, 본 명세서의 Cas9 단백질은, 예를 들어 서열 번호:11 또는 서열 번호:11의 잔기 1 내지 1368의 아미노산 서열을 포함할 수 있다. 또한 대안적으로, Cas9 단백질은, 예를 들어, 전술한 아미노산 서열 중 어느 하나와 적어도 약 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99% 동일한 아미노산 서열을 포함할 수 있다. 이러한 변이체인 Cas9 단백질은 본 명세서의 RNA 성분과 결합할 경우 DNA에 대한 특이적 결합 활성 및 임의로 절단 또는 닉킹 활성을 가져야 한다.
본 명세서에서 사용된 Cas 단백질 (예를 들어, Cas9)의 기원은 RNA 성분(들)이 유래된 것과 동일한 종 또는 상이한 종으로부터 유래될 수 있다. 예를 들어, 스트렙토코커스 종 (예를 들어, S. 피오제네스 또는 S. 써모필러스)으로부터 유래된 Cas9 단백질을 포함하는 RGEN은 동일한 스트렙토코커스 종으로부터 유래된 서열 (예를 들어, crRNA 반복 서열, tracrRNA 서열)을 갖는 적어도 하나의 RNA 성분과 복합체를 형성할 수 있다. 대안적으로, 본 명세서에서 사용된 Cas 단백질 (예를 들어, Cas9)의 기원은 RNA 성분(들)이 유래된 종과 상이한 종으로부터 유래될 수 있다 (Cas 단백질 및 RNA 성분(들)은 서로 이종성일 수 있음); 이러한 이종 Cas/RNA 성분 RGEN은 DNA 표적화 활성을 가져야 한다.
특정 표적 DNA 서열에 대한 본 명세서의 Cas 단백질의 결합 활성 및/또는 엔도뉴클레오리틱 활성을 결정하는 것은 본 명세서에 참고로 개시되는 미국 특허 제8697359호에 개시된 바와 같은 본 기술 분야에 알려진 임의의 적합한 검정에 의해 평가될 수 있다. 예를 들어, 통상적이지 않은 효모에서 Cas 단백질과 적합한 RNA 성분을 발현한 후, 삽입-결실의 존재가 예측되는 DNA 표적 부위를 조사함으로써 측정을 행할 수 있다 (이러한 특정 검정의 Cas 단백질은 완전한 엔도뉴클레오리틱 활성 [이중 가닥 절단 활성]을 가질 것이다). 예측되는 표적 부위에서 삽입-결실의 존재를 조사하는 것은, 예를 들어, DNA 서열 분석법을 통해 또는 표적 서열의 기능 상실을 분석하여 삽입-결실 형성을 추정함으로써 수행될 수 있다. 또 다른 예에서, Cas 단백질 활성은 표적 부위 또는 그 부근의 서열에 상동성인 서열을 포함하는 공여 DNA를 제공하는 통상적이지 않은 효모에서 Cas 단백질과 적합한 RNA 성분을 발현시킴으로써 측정될 수 있다. 표적 부위에서의 공여 DNA 서열의 존재는 (공여자와 표적 서열 사이의 성공적인 HR에 의해 예측되는 바와 같은) 표적화가 일어났음을 나타낼 것이다.
본 명세서의 Cas 단백질, 예컨대 Cas9는 전형적으로 이종 핵 국재화 서열 (NLS)을 추가로 포함한다. 본 명세서에서, 이종 NLS 아미노산 서열은, 예를 들어, 본 명세서의 효모 세포의 핵에 검출가능한 양으로 Cas 단백질의 축적을 유도하기 충분한 강도일 수 있다. NLS는 염기성의 양으로 하전된 잔기 (예를 들어, 라이신 및/또는 아르기닌)의 하나 (1부분(monopartite)) 또는 그 이상 (예를 들어, 2부분(bipartite))의 짧은 서열 (예를 들어, 2 내지 20개의 잔기)을 포함할 수 있으며, Cas 아미노산 서열 중 어디에도 위치할 수 있지만 단백질 표면 상에 노출되어야 한다. 본 명세서에서, NLS는, 예를 들어, Cas 단백질의 N-말단 또는 C-말단에 작동가능하게 연결될 수 있다. 둘 이상의 NLS 서열은 Cas 단백질 연결될 수 있는데, 예를 들어 Cas 단백질의 N- 및 C-말단 모두에 연결될 수 있다. 본 명세서에서 적합한 NLS 서열의 비제한적인 예에는 모두 본 명세서에 참고로 포함되는 미국 특허 제6660830호 및 제7309576호 (예를 들어, 그 안의 표 1)에 개시된 것들이 포함된다. 본 명세서에서 유용한 NLS의 또 다른 예에는 서열 번호:11의 아미노산 잔기 1373-1379가 포함된다.
소정 실시 형태에서, Cas 단백질에 의한 DNA-특이적 표적화를 유도하는 Cas 단백질 및 이의 각각의 RNA 성분 (예를 들어, crRNA)은 개시된 통상적이지 않은 효모에 이종성이다. 이러한 RGEN 성분의 이종성은 Cas 단백질 및 이들의 각각의 RNA 성분이 알려진 원핵생물 (박테리아 및 고세균)에만 존재한다는 사실에 기인한다.
본 명세서에서, Cas 단백질은 효모 세포에서의 발현을 위해 코돈-최적화된 오픈 리딩 프레임 (ORF)을 사용하여 통상적이지 않은 효모 세포에서 임의로 발현될 수 있다. 본 명세서에서, "코돈-최적화된" 서열은 이의 코돈 사용 빈도가 숙주 세포의 바람직한 코돈 사용 빈도를 모방하도록 설계된 유전자이다. 야로위아 리폴리티카가 통상적이지 않은 효모 세포인 측면에서, ORF의 코돈 최적화는 본 명세서에 참고로 포함되는 미국 특허 제7125672호에 제공된 바와 같은 야로위아 리폴리티카 코돈 사용 프로파일에 따라 수행될 수 있다.
일부 실시 형태에서, Cas 단백질은 하나 이상의 이종 단백질 도메인 (예를 들어, Cas 단백질에 더하여 1, 2, 3 또는 그 이상의 도메인)을 포함하는 융합 단백질의 일부이다. 이러한 융합 단백질은 임의의 추가의 단백질 서열 및 임의로 임의의 2개의 도메인 사이, 예컨대 Cas와 제1 이종 도메인 사이의 링커 서열을 포함할 수 있다. 본 명세서에서 Cas 단백질에 융합될 수 있는 단백질 도메인의 예에는 에피토프 태그 (예를 들어, 히스티딘 [His], V5, FLAG, 인플루엔자 혈구응집소 [HA], myc, VSV-G, 티오레독신 [Trx]), 리포터 (예를 들어, 글루타티온-5-트랜스퍼라제 [GST], 고추냉이 퍼옥시다제 [HRP], 클로람페니콜 아세틸트랜스퍼라제 [CAT], 베타-갈락토시다제, 베타-글루쿠로니다제 [GUS], 루시퍼라제, 녹색 형광 단백질 [GFP], HcRed, DsRed, 시안 형광 단백질 [CFP], 황색 형광 단백질 [YFP], 청색 형광 단백질 [BFP]) 및: 메틸라제 활성, 데메틸라제 활성, 전사 활성화 활성 (예를 들어, VP16 또는 VP64), 전사 억제 활성, 전사 방출 인자 활성, 히스톤 변형 활성, RNA 절단 활성 및 핵산 결합 활성 중 하나 이상의 활성을 갖는 도메인을 제한 없이 포함된다. 다른 실시 형태에서, Cas 단백질은 DNA 분자 또는 다른 분자, 예컨대 말토스 결합 단백질 (MBP), S-태그, Lex A DNA 결합 도메인 (DBD), GAL4A DNA 결합 도메인 및 단순 헤르페스 바이러스 (HSV) VP16에 결합하는 단백질과 융합될 수 있다. 본 명세서에서, Cas 단백질을 포함하는 융합 단백질의 일부가 될 수 있는 추가의 도메인은 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2011/0059502호에 개시되어 있다. Cas 단백질이 이종 단백질 (예를 들어, 전사 인자)에 융합되는 소정 실시 형태에서, Cas 단백질은 (본 명세서의 적합한 RNA 성분과 복합체인 경우) DNA 인식 및 결합 활성을 가지나, DNA 닉킹 또는 절단 활성은 없다.
본 명세서에서, RGEN은 DNA 표적 서열에서 DNA 가닥에 결합할 수 있고, 임의로 절단할 수 있다. 소정 실시 형태에서, RGEN은 DNA 표적 서열의 하나 또는 두 가닥을 절단할 수 있다. RGEN은, 예를 들어, DNA 표적 서열의 두 가닥을 절단할 수 있다.
본 명세서에서, DNA 표적 서열의 두 가닥을 절단할 수 있는 RGEN은 전형적으로 이의 엔도뉴클레아제 도메인 모두를 기능적 상태(functional state)로 포함하는 Cas 단백질을 포함한다 (예를 들어, 야생형 엔도뉴클레아제 도메인 또는 각각의 엔도뉴클레아제 도메인에서 일부 또는 모든 활성을 보유하는 이들의 변이체). 따라서, 야생형 Cas 단백질 (예를 들어, 본 명세서에 개시된 Cas9 단백질) 또는 Cas 단백질 각각의 엔도뉴클레아제 도메인에서 일부 또는 모든 활성을 보유하는 이들의 변이체가 DNA 표적 서열의 두 가닥을 절단할 수 있는 RGEN의 적합한 예이다. 기능적 RuvC 및 HNH 뉴클레아제 도메인을 포함하는 Cas9 단백질은 DNA 표적 서열의 두 가닥을 절단할 수 있는 Cas 단백질의 예이다. 본 명세서에서 DNA 표적 서열의 두 가닥을 절단할 수 있는 RGEN은 전형적으로 절단 부위에 평활 말단 (즉, 뉴클레오티드 돌출부가 없음)을 형성하도록 동일한 위치에서 두 가닥을 절단한다.
본 명세서에서 DNA 표적 서열의 한 가닥을 절단할 수 있는 RGEN은 본 명세서에서 닉카아제 활성 (예를 들어, 부분 절단 능력)을 갖는 것으로 특징지어질 수 있다. 본 명세서에서, Cas 닉카아제 (예를 들어, Cas9 닉카아제)는 전형적으로 Cas가 DNA 표적 서열 중 한 가닥만 절단하도록 하는 (즉, 닉을 형성) 하나의 기능적 엔도뉴클레아제 도메인을 포함한다. 예를 들어, Cas9 닉카아제는 (i) 기능장애 돌연변이인 RuvC 도메인 및 (ii) 기능적 HNH 도메인 (예를 들어, 야생형 HNH 도메인)을 포함할 수 있다. 또 다른 예로서, Cas9 닉카아제는 (i) 기능적 RuvC 도메인 (예를 들어, 야생형 RuvC 도메인) 및 (ii) 기능장애 돌연변이인 HNH 도메인을 포함할 수 있다.
본 명세서에 사용하기에 적합한 Cas9 닉카아제의 비제한적인 예가 본 명세서에 참고로 포함되는 문헌[Gasiunas et aL. (ProC. NatL. AcaD. Sci. U.S.A. 109:E2579-E2586)], 문헌[Jinek et aL. (Science 337:816-821)], 문헌[Sapranauskas et aL. (Nucleic Acids Res. 39:9275-9282)] 및 미국 특허 출원 공개 제2014/0189896호에 의해 개시되어 있다. 예를 들어, 본 명세서에서 Cas9 닉카아제는 Asp-31 치환 (예를 들어, Asp-31-Ala) (돌연변이 RuvC 도메인의 예) 또는 His-865 치환 (예를 들어, His-865-Ala), Asn-882 치환 (예를 들어, Asn-882-Ala) 또는 Asn-891 치환 (예를 들어, Asn-891-Ala) (HNH 도메인 돌연변이의 예)을 갖는 S. 써모필러스 Cas9를 포함할 수 있다. 또한 예를 들어, 본 명세서에서 Cas9 닉카아제는 Asp-10 치환 (예를 들어, Asp-10-Ala), Glu-762 치환 (예를 들어, Glu-762-Ala) 또는 Asp-986 치환 (예를 들어, Asp-986-Ala) (돌연변이 RuvC 도메인의 예) 또는 His-840 치환 (예를 들어, His-840-Ala), Asn-854 치환 (예를 들어, Asn-854-Ala) 또는 Asn-863 치환 (예를 들어, Asn-863-Ala) (돌연변이 HNH 도메인의 예)을 갖는 S. 피오제네스 Cas9를 포함할 수 있다. S. 피오제네스 Cas9에 관하여, 3개의 RuvC 서브도메인은 일반적으로 각각 아미노산 잔기 1-59, 718-769 및 909-1098에 위치하고, HNH 도메인은 아미노산 잔기 775-908에 위치한다 (문헌[Nishimasu et al., Cell 156:935-949]).
본 명세서에서, Cas9 닉카아제는 개시된 발명의 통상적이지 않은 효모에서 다양한 용도로 사용될 수 있다. 예를 들어, Cas9 닉카아제는 적합한 공여 폴리뉴클레오티드를 갖는 DNA 표적 부위 서열에 또는 그 부근에서 HR을 촉진하는 데 사용될 수 있다. 닉이 있는 DNA는 NHEJ 공정을 위한 기질이 아니고, HR 공정에 의해 인식되기 때문에, 특정 표적 부위에서 DNA에 닉을 형성하는 것은 적합한 공여 폴리뉴클레오티드를 사용하는 HR을 더 잘 수용할 수 있게 해야 한다.
또 다른 예로서, DNA 표적화의 특이성을 높이기 위해 한 쌍의 Cas9 닉카아제가 사용될 수 있다. 일반적으로, 이는 상이한 가이드 서열을 갖는 RNA 성분과 관련되어 있기 때문에, 원하는 표적화를 위한 영역에서 반대 가닥 상의 가까운 DNA 서열을 표적화하고 닉을 형성하는 2개의 Cas9 닉카아제를 제공함으로써 수행될 수 있다. 각각의 DNA 가닥의 이러한 가까운 절단은 DSB (즉, 단일-가닥 돌출부를 갖는 DSB)을 생성하고, 이는 이어서 NHEJ (삽입-결실 형성을 유도함) 또는 HR (제공된다면, 적합한 공여 폴리뉴클레오티드와의 재조합을 유도함)의 기질로 인식된다. 이러한 실시 형태에서 각각의 닉은, 예를 들어, 서로 적어도 약 5, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90 또는 100 (또는 5 내지 100 사이의 임의의 정수)개의 염기만큼 이격될 수 있다. 본 명세서에서, 상기한 바와 같이 하나 또는 2개의 Cas9 닉카아제 단백질이 Cas9 닉카아제 쌍에서 사용될 수 있다. 예를 들어, 돌연변이 RuvC 도메인을 가지나 기능하는 HNH 도메인 (즉, Cas9 HNH+/RuvC-)을 갖는 Cas9 닉카아제 (예를 들어, S. 피오제네스 Cas9 HNH+/RuvC-)를 사용할 수 있다. 각각의 닉카아제를 각각의 특정 DNA 부위로 표적화하는 가이드 RNA 서열을 갖는 본 명세서의 적합한 RNA 성분을 사용하여, 서로 가까운 (100개의 염기쌍 이하로 이격된) 특정 DNA 부위로 각각의 Cas9 닉카아제 (예를 들어, Cas9 HNH+/RuvC-)를 유도할 것이다.
소정 실시 형태에서 RGEN은 DNA 표적 부위는 서열에 결합할 수 있지만, 표적 부위 서열에서 임의의 가닥을 절단하지 않는다. 이러한 RGEN은 이의 모든 뉴클레아제 도메인이 기능장애 돌연변이인 Cas 단백질을 포함할 수 있다. 예를 들어, 본 명세서에서 DNA 표적 부위 서열에 결합할 수 있지만, 표적 부위 서열에서 임의의 가닥을 절단하지 않는 Cas9 단백질은 기능장애 돌연변이인 RuvC 도메인 및 기능장애 돌연변이인 HNH 도메인을 포함할 수 있다. 이러한 Cas9 단백질의 비제한적인 예에는 상기 개시된 RuvC 및 HNH 뉴클레아제 도메인 돌연변이 (예를 들어, Asp-10 치환, 예컨대 Asp-10-Ala 및 His-840 치환, 예컨대 His-840-Ala을 갖는 S. 피오제네스 Cas9) 중 어느 하나가 포함된다. 본 명세서에서, 표적 DNA 서열에 결합하지만 절단하지 않는 Cas 단백질은 유전자 발현을 조절하는 데 사용될 수 있으며, 예를 들어, 이 경우 Cas 단백질은 전사 인자 (또는 이의 일부) (예를 들어, 억제 인자 또는 활성 인자, 예컨대 본 명세서에 개시된 것 중 어느 하나)와 융합될 수 있다. 예를 들어, Asp-10 치환 (예를 들어, Asp-10-Ala) 및 His-840 치환 (예를 들어, His-840-Ala)을 갖는 S. 피오제네스 Cas9를 포함하는 Cas9는 VP16 또는 VP64 전사 활성 인자 도메인에 융합될 수 있다. 이러한 RGEN의 RNA 성분에서 사용되는 가이드 서열은, 예를 들어 유전자 프로모터 또는 다른 조절 요소 (예를 들어, 인트론) 내의 DNA 서열에 상보적일 것이다.
소정 측면의 효모는 (i) DNA 표적 서열의 DNA 가닥 중 하나 또는 둘 모두를 절단할 수 있는 RGEN 및 (ii) DNA 표적 부위 또는 그 부근의 서열에 상동성인 적어도 하나의 서열을 포함하는 공여 폴리뉴클레오티드 (본 명세서의 Cas 단백질에 의해 특이적으로 표적화된 서열)을 포함할 수 있다. 적합한 공여 폴리뉴클레오티드는 표적 부위가 (예를 들어, 본 명세서의 Cas 단백질을 사용하여 도입될 수 있는) SSB 또는 DSB를 함유하는 경우, DNA 표적 부위 또는 그 부근의 서열과 HR을 수행할 수 있다. 본 명세서의 공여 폴리뉴클레오티드 내의 "상동성 서열"은, 예를 들어, 서열 표적 부위 또는 그 부근의 서열과 100% 동일성 또는 서열 표적 부위 또는 그 부근의 서열과 적어도 약 95%, 96%, 97%, 98% 또는 99% 동일성을 갖는 적어도 약 25, 50, 75, 100, 150, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000 또는 10000개의 뉴클레오티드 또는 약 50 내지 500, 50 내지 550, 50 내지 600, 50 내지 650 또는 50 내지 700개의 뉴클레오티드의 서열을 포함할 수 있거나 이로 이루어질 수 있다.
본 명세서의 공여 폴리뉴클레오티드는, 예를 들어, 표적 부위 서열 또는 그 부근의 서열에 이종성인 서열에 의해 이격된 2개의 상동성 서열 (상동성 암)을 가질 수 있다. 이러한 공여 폴리뉴클레오티드와 표적 부위 서열 사이의 HR은 전형적으로 표적 부위에서 서열을 공여 폴리뉴클레오티드의 이종 서열로 대체시킨다 (공여 폴리뉴클레오티드의 상동성 암에 상동성인 표적 부위 서열 사이에 위치한 표적 부위 서열은 공여 폴리뉴클레오티드의 이종 서열에 의해 대체된다). 2개의 상동성 암을 갖는 공여 폴리뉴클레오티드에서, 암은, 예를 들어, 적어도 약 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 75, 100, 250, 500, 1000, 2500, 5000, 10000, 15000, 20000, 25000 또는 30000개의 뉴클레오티드 만큼 이격될 수 있다 (즉, 공여 폴리뉴클레오티드의 이종 서열은 길이가 적어도 약 1, 2, 3, 4, 5, 10, 20, 30, 40, 50, 75, 100, 250, 500, 1000, 2500, 5000, 10000, 15000, 20000, 25000 또는 30000개 뉴클레오티드이다). 각각의 상동성 암의 길이 (예를 들어, 상동성 서열에 대해 상기 개시된 임의의 길이)는 동일하거나 상이할 수 있다. 각각의 표적 부위 또는 그 부근의 상동성 서열을 갖는 각각의 암의 % 동일성 (예를 들어, 상동성 서열에 대해 상기 개시된 임의의 % 동일성)은 동일하거나 상이할 수 있다.
공여 폴리뉴클레오티드의 상응하는 상동성 서열에 상동성인 표적 부위 서열 또는 그 부근 (대안적으로, 인근 또는 이의 근접에 있는)의 DNA 서열은, 예를 들어, 표적 서열 내의 예측되는 Cas 단백질 절단 부위 (DSB 또는 닉)로부터 약 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 450, 500, 750, 1000, 2000, 3000, 4000, 5000, 6000, 7000, 8000, 9000, 10000, 20000, 30000, 40000, 50000 또는 60000 (또는 1 내지 60000 사이의 임의의 정수)개의 뉴클레오티드 (예를 들어, 약 1 내지 1000, 100 내지 1000, 500 내지 1000, 1 내지 500 또는 100 내지 500 뉴클레오티드) 내에 있을 수 있다. 이러한 뉴클레오티드 거리는 절단 부위로부터 상동성 서열의 첫번째 뉴클레오티드까지로 표시될 수 있으며, 절단 부위로부터 상류 또는 하류 방향으로 진행될 수 있다. 예를 들어, 공여 폴리뉴클레오티드의 상응하는 서열에 상동성인 표적 서열 근처의 서열은 표적 서열에서 예측되는 Cas 단백질 절단 부위의 500개의 뉴클레오티드 염기쌍 하류에서 시작할 수 있다. 2개의 상동성 암 (예를 들어, 이종 서열에 의해 이격된 제1 및 제2 상동성 암)을 갖는 공여 폴리뉴클레오티드를 사용하는 본 명세서의 실시 형태에서, 예를 들어, (공여자의 제1 상동성 암과 상동성에 해당하는) 상동성 서열은 예측되는 Cas 절단 부위의 상류에 있을 수 있고 (공여자의 제2 상동성 암과 상동성에 해당하는) 상동성 서열은 예측되는 Cas 절단 부위의 상류에 있을 수 있다. 예측되는 절단 부위로부터 이러한 상류 및 하류 상동성 서열 각각의 뉴클레오티드 거리는 동일하거나 상이할 수 있으며, 예를 들어, 상기 개시된 뉴클레오티드 거리 중 어느 하나일 수 있다. 예를 들어, (공여자의 제1 상동성 암과 상동성에 해당하는) 상동성 서열의 3' 말단은 예측되는 Cas 절단 부위의 600개의 뉴클레오티드 염기쌍 상류에 위치할 수 있고, (공여자의 제2 상동성 암과 상동성에 해당하는) 상동성 서열의 5' 말단은 예측되는 Cas 절단 부위의 400개의 뉴클레오티드 염기쌍 하류에 위치할 수 있다.
본 명세서에서, RGEN은 통상적이지 않은 효모의 게놈 내의 염색체, 에피솜 또는 임의의 다른 DNA 분자의 표적 부위 서열에서 DNA 가닥에 결합하고, 임의로 절단할 수 있다. RGEN의 RNA 성분이 표적 서열의 가닥에 상보적인 서열 (가이드 서열)을 포함하는 경우, 이러한 표적 서열의 인식 및 결합은 특이적이다. 소정 실시 형태에서, 표적 부위는 유일할 수 있다 (즉, 대상의 게놈에서 표적 부위 서열은 한번만 발생한다).
본 명세서에서, 표적 서열의 길이는, 예를 들어, 적어도 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 또는 30개의 뉴클레오티드; 13 내지 30개의 뉴클레오티드; 17 내지 25개의 뉴클레오티드; 또는 17 내지 20개의 뉴클레오티드일 수 있다. 이러한 길이는 PAM 서열을 포함하거나 제외할 수 있다. 또한, 본 명세서에서 표적 서열의 가닥은 가이드 서열과 혼성화되고, (적합한 PAM이 표적 서열에 인접한 경우, 아래 참조) 표적 서열에 Cas 단백질 또는 Cas 단백질 복합체를 직접 서열-특이적으로 결합시키기 위한 (crRNA 또는 gRNA의) 가이드 서열과의 충분한 상보성을 갖는다. 가이드 서열과 이의 상응하는 DNA 표적 서열의 가닥 사이의 상보성 정도는, 예를 들어, 적어도 약 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%이다. 본 명세서에서, 표적 부위는, 예를 들어, 유전자 산물 (예를 들어, 단백질 또는 RNA)을 암호화하는 서열 또는 비암호화 서열 (예를 들어, 조절 서열 또는 "정크(junk)" 서열) 내에 위치할 수 있다.
PAM (프로토스페이서-인접 모티프) 서열은 표적 부위 서열에 인접할 수 있다. PAM 서열은 본 명세서의 RGEN에 의해 인식되는 짧은 DNA 서열이다. 관련된 PAM 및 DNA 표적 서열의 처음 11개의 뉴클레오티드는 Cas9/gRNA 표적화 및 절단에 중요할 수 있다 (문헌[Jiang et al., NaT. Biotech. 31:233-239]). 본 명세서에서, PAM 서열의 길이는 사용되는 Cas 단백질 또는 Cas 단백질 복합체에 따라 다를 수 있지만, 전형적으로는, 예를 들어 2, 3, 4, 5, 6, 7 또는 8개의 뉴클레오티드 길이이다. PAM 서열은, 예를 들어, 결과적으로 RNA 성분 가이드 서열에 상보적인 표적 부위의 가닥에 상보적인 표적 부위 서열로부터 바로 하류에, 또는 이의 하류에 있는 2 또는 3개의 뉴클레오티드 내에 있다. RGEN이 RNA 성분과 복합체를 형성한 핵산 내부 분해 방식으로(endonucleolytically) 활성인 Cas9 단백질인 본 명세서의 실시 형태에서, Cas9는 RNA 성분에 의해 유도된 바와 같이 표적 서열에 결합하고, PAM 서열의 상류 세 번째 뉴클레오티드 위치의 5' 부근에서 두 가닥을 절단한다. 표적 부위:PAM 서열의 다음의 예를 고려하라:
이러한 예의 서열에서, N은 A, C, T 또는 G일 수 있고, X는 A, C, T 또는 G일 수 있다 (X는 또한 NPAM로도 지칭됨). 이러한 예에서, PAM 서열은 (밑줄친) XGG이다. 적합한 Cas9/RNA 성분 복합체는 이러한 표적을 이중 밑줄로된 N의 5' 부근에서 절단할 것이다. 서열 번호:46에서 N으로 이루어진 열(string of N's)은 본 명세서의 RNA 성분의 가이드 서열과, 예를 들어, 적어도 약 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100% 동일한 표적 서열을 나타낸다 (여기서, DNA 표적 서열의 임의의 T는 RNA 가이드 서열의 임의의 U와 정렬될 것임). (본 명세서에서 표적 부위를 나타내는) 이러한 표적 서열을 인식하고 결합하는데 있어서, Cas9 복합체의 RNA 성분의 가이드 서열은, N으로 이루어진 열의 보체 서열과 어닐링될 것이며; 가이드 서열과 표적 부위 보체 사이의 % 상보성은, 예를 들어, 적어도 약 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%이다. Cas9 닉카아제가 게놈에서 서열 번호:46을 표적으로 하는데 사용되는 경우, 닉카아제는 닉카아제의 어느 엔도뉴클레아제 도메인가 기능장애인지에 따라 이중 밑줄로된 N의 5' 부근에서 또는 상보성 가닥의 동일한 위치에서 닉을 형성할 것이다. (RuvC와 HNH 도메인이 모두 기능장애) 핵산 분해 활성이 없는 Cas9가 게놈에서 서열 번호:46을 표적으로 하는데 사용되는 경우, 표적 서열을 인식하고 결합하지만, 어떤 서열도 절단하지 않는다.
본 명세서에서, PAM은 전형적으로 사용되는 RGEN의 타입을 고려하여 선택된다. 본 명세서에서, PAM 서열은, 예를 들어, Cas가 유래될 수 있는 본 명세서에 개시된 임의의 종으로부터 유래된 Cas, 예컨대 Cas9를 포함하는 RGEN에 의해 인식되는 것일 수 있다. 소정 실시 형태에서, PAM 서열은 S. 피오제네스, S. 써모필러스, S. 아갈락티아, N. 메닝기티디스, T. 덴티콜라 또는 F. 노비시다로부터 유래된 Cas9를 포함하는 RGEN에 의해 인식되는 것일 수 있다. 예를 들어, S. 피오제네스로부터 유래된 적합한 Cas9는 NGG의 PAM 서열 (서열 번호:47; N은 A, C, T 또는 G일 수 있음)을 갖는 표적 게놈 서열을 표적화하는 데 사용될 수 있다. 다른 예로서, 적합한 Cas9는 다음과 같은 PAM 서열을 갖는 DNA 서열을 표적화하는 경우 하기의 종 중 어느 하나로부터 유래될 수 있다: S. 써모필러스 (NNAGAA [서열 번호:48]), S. 아갈락티아 (NGG [서열 번호:47]), NNAGAAW [서열 번호:49, W는 A 또는 T임], NGGNG [서열 번호:50]), N. 메닝기티디스 (NNNNGATT [서열 번호:51]), T. 덴티콜라 (NAAAAC [서열 번호:52]) 또는 F. 노비시다 (NG [서열 번호:53]) (여기서, 이러한 모든 특정 PAM 서열에서 N으로 이루어진 서열은 A, C, T 또는 G임). 본 명세서에서 유용한 Cas9/PAM의 다른 예에는, 본 명세서에 참고로 포함되는 문헌[Shah et aL. (RNA Biology 10:891-899)] 및 문헌[Esvelt et aL. (Nature Methods 10:1116-1121)]에 개시된 것들이 포함된다. 본 명세서에서, 표적 서열의 예는 서열 번호:46을 따르지만, 'XGG' PAM은 전술한 PAM 중 어느 하나에 의해 대체된다.
본 명세서의 실시 형태에서, 5'-캡을 갖지 않는 적어도 하나의 RNA 성분은 RGEN에 포함된다. 이러한 캡핑되지 않은 RNA 성분은 통상적이지 않은 효모 내의 염색체 또는 에피솜의 표적 부위 서열에 상보적인 서열을 포함한다. RGEN은 이러한 서열 상보성을 기초하여 표적 부위에서 DNA 가닥에 특이적으로 결합하고, 임의로 절단한다. 따라서, 개시된 발명의 실시 형태에서 RNA 성분의 상보적인 서열은 가이드 서열 또는 가변 표적화 도메인로도 지칭될 수 있다.
본 명세서에서, RNA 성분의 가이드 서열 (예를 들어, crRNA 또는 gRNA)은 길이가, 예를 들어, 적어도 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 또는 30개의 리보뉴클레오티드; 13 내지 30개의 리보뉴클레오티드; 17 내지 25개의 리보뉴클레오티드; 또는 17 내지 20개의 리보뉴클레오티드일 수 있다. 일반적으로, 본 명세서에서 가이드 서열은 표적 서열과 혼성화되고, (적합한 PAM이 표적 서열에 인접한 경우) 표적 서열에 Cas 단백질 또는 Cas 단백질 복합체를 직접 서열-특이적으로 결합시키기 위한 (crRNA 또는 gRNA의) 가이드 서열과의 충분한 상보성을 갖는다. 가이드 서열과 이의 상응하는 DNA 표적 서열 사이의 상보성 정도는, 예를 들어, 적어도 약 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%이다. 가이드 서열은 효모 세포의 DNA 표적 서열로 RGEN을 표적화하도록 적절하게 유전자 조작될 수 있다.
본 명세서에서, RNA 성분은, 예를 들어, 가이드 서열을 포함하는 crRNA 및 반복 (tracrRNA 메이트) 서열을 포함할 수 있다. 가이드 서열은 전형적으로 crRNA의 5' 말단에 또는 그 부근 (1, 2, 3, 4, 5, 6, 7, 8, 9, 10 또는 그 이상의 염기 내에)에 위치한다. crRNA의 가이드 서열의 하류는 tracrRNA의 5' 말단에서 서열에 상보적이고 이와 혼성화될 수 있는 "반복" 또는 "tracrRNA 메이트" 서열이다. 가이드 및 tracrRNA 메이트 서열은, 예를 들어, 바로 인접하거나, 1, 2, 3, 4 또는 그 이상의 염기에 의해 이격될 수 있다. tracrRNA 메이트 서열은 tracrRNA의 5' 말단에, 예를 들어, 적어도 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% 또는 99% 서열 상보성을 갖는다. 일반적으로, 상보성 정도는 tracrRNA 메이트 서열과 tracrRNA 서열의 최적 정렬을 참고하여, 두 서열 중 더 짧은 길이에 따를 수 있다. 본 명세서에서, tracrRNA 메이트 서열의 길이는, 예를 들어 길이가 적어도 8, 9, 10, 11, 12, 13, 14, 15, 16, 17 또는 18개의 리보뉴클레오티드일 수 있고, tracrRNA의 5' 말단에서 동일하거나 유사한 길이 (예를 들어, +/- 1, 2, 3, 4 또는 5개의 염기)의 서열과 혼성화된다. 본 명세서에서, tracrRNA 메이트 서열의 적합한 예에는 서열 번호:54 (guuuuuguacucucaagauuua), 서열 번호:55 (guuuuuguacucuca), 서열 번호:56 (guuuuagagcua, 실시예 참조) 또는 서열 번호:57 (guuuuagagcuag) 또는 (i) 적어도 약 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 서열 동일성을 갖고 (ii) tracrRNA의 5'-말단 서열과 어닐링할 수 있는 이들의 변이체가 포함된다. 본 명세서에서, crRNA의 길이는, 예를 들어, 적어도 약 18, 20, 22, 24, 26, 28, 30, 32, 34, 36, 38, 40, 42, 44, 46 또는 48개의 리보뉴클레오티드; 또는 약 18 내지 48개의 리보뉴클레오티드; 또는 약 25 내지 50개의 리보뉴클레오티드일 수 있다.
tracrRNA는 타입 II 크리스퍼 시스템의 Cas9 단백질이 RGEN에 포함되는 실시 형태에서 crRNA와 함께 포함되어야 한다. 본 명세서에서, tracrRNA는 5'에서 3' 방향으로 (i) crRNA의 반복 영역 (tracrRNA 메이트 서열)과 어닐링하는 서열 및 (ii) 스템 루프-함유 부분을 포함한다. (i)의 서열의 길이는, 예를 들어, 상기 개시된 임의의 tracrRNA 메이트 서열의 길이와 동일하거나 유사할 수 있다 (예를 들어, +/- 1, 2, 3, 4 또는 5개의 염기). 본 명세서에서, tracrRNA의 총 길이 (즉, 서열 성분 [i] 및 [ii])는, 예를 들어, 적어도 약 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85 또는 90개의 (또는 30 내지 90 사이의 임의의 정수) 리보뉴클레오티드일 수 있다. tracrRNA는 3'-말단에 1, 2, 3, 4, 5 또는 그 이상의 우라실 잔기를 추가로 포함할 수 있으며, 이는 전사 종결 서열을 갖는 tracrRNA를 발현함으로써 존재할 수 있다.
본 명세서에서, tracrRNA는, 예를 들어, Cas9 서열이 유래될 수 있는, 상기 열거된 임의의 박테리아 종으로부터 유래될 수 있다. 적합한 tracrRNA 서열의 예에는 본 명세서에 참고로 포함되는 미국 특허 제8697359호 및 문헌[Chylinski et aL. (RNA Biology 10:726-737)]에 개시된 것들이 포함된다. 본 명세서에서 바람직한 tracrRNA는 스트렙토코커스 종 tracrRNA (예를 들어, S. 피오제네스, S. 써모필러스)로부터 유래될 수 있다. 본 명세서의 tracrRNA의 다른 적합한 예는 하기를 포함할 수 있다:
서열 번호:58:
서열 번호:59:
서열 번호:60:
(S. 피오제네스 tracrRNA로부터 유래됨). 본 명세서의 tracrRNA의 다른 적합한 예는 하기를 포함할 수 있다:
서열 번호:61:
서열 번호:63:
(S. 써모필러스 tracrRNA로부터 유래됨).
본 명세서의 tracrRNA의 또 다른 예는 (i) 이와 적어도 약 80%, 85%, 90, 91, 92, 93, 94, 95, 96, 97, 98 또는 99%의 서열 동일성을 갖고 (ii) tracrRNA로서 작용할 수 있는 이들 tracrRNA 서열 번호의 변이체이다 (예를 들어, 5'-말단 서열은 crRNA의 tracrRNA 메이트 서열에 어닐링할 수 있고, 5'-말단 서열의 하류 서열은 하나 이상의 헤어핀을 형성할 수 있으며, 변이체 tracrRNA는 Cas9 단백질과 복합체를 형성할 수 있다).
본 명세서에 개시된 RGEN의 RNA 성분은, 예를 들어, tracrRNA에 작동가능하게 연결되거나 융합된 crRNA를 포함하는 가이드 RNA (gRNA)를 포함할 수 있다. 특정 바람직한 실시 형태에서, gRNA의 crRNA 성분은 tracrRNA 성분의 상류에 있다 (즉, 이러한 gRNA는 5'에서 3' 방향으로 tracrRNA에 작동가능하게 연결된 crRNA를 포함한다). 본 명세서에 개시된 바와 같은 (예를 들어, 상기 실시 형태) 임의의 crRNA 및/또는 tracrRNA (및/또는 이들의 부분, 예컨대 crRNA 반복 서열, tracrRNA 메이트 서열 또는 tracrRNA 5'-말단 서열)는, 예를 들어, gRNA에 포함될 수 있다.
본 명세서에서, gRNA의 crRNA 성분의 tracrRNA 메이트 서열은 tracrRNA 성분의 5'-말단과 어닐링함으로써, 헤어핀 구조를 형성할 수 있어야 한다. (crRNA 성분의) tracrRNA 메이트 서열과 (tracrRNA 성분의) 5'-말단 서열 사이의 길이 및 % 상보성에 관한 상기 개시 중 임의의 것은, 예를 들어 gRNA의 crRNA 및 tracrRNA 성분을 특징지을 수 있다. 이러한 어닐링을 용이하게 하기 위해, crRNA와 tracrRNA 성분의 작동가능한 결합 또는 융합은 바람직하게는 적합한 루프 형성 리보뉴클레오티드 서열을 포함한다 (즉, 루프 형성 서열은 crRNA와 tracrRNA 성분을 함께 연결하여 gRNA를 형성할 수 있음). RNA 루프 형성 서열의 적합한 예에는 GAAA (서열 번호:43, 실시예 참조), CAAA (서열 번호:44) 및 AAAG (서열 번호:45)가 포함된다. 그러나, 대체 루프 서열이 사용되는 바와 같이 더 길거나 더 짧은 루프 서열이 사용될 수 있다. 루프 서열은 바람직하게는 리보뉴클레오티드 트리플렛 (예를 들어, AAA) 및 트리플렛의 어느 하나의 말단에 추가의 리보뉴클레오티드 (예를 들어, C 또는 G)를 포함한다.
본 명세서에서, gRNA는 (crRNA 성분의) 이의 tracrRNA 메이트 서열과 tracrRNA 5'-말단 서열 부분을 어닐링하여 헤어핀 ("제1 헤어핀")을 형성한다. gRNA의 tracrRNA 성분의 서열에 따라, 하나 이상의 (예를 들어, 1, 2, 3 또는 4개) 추가의 헤어핀 구조가 이러한 제1 헤어핀의 하류를 형성할 수 있다. 따라서, gRNA는, 예를 들어 최대 5개의 헤어핀 구조를 가질 수 있다. gRNA는 gRNA 서열의 말단에 이어지는 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 또는 그 이상의 잔기를 추가로 포함할 수 있는데, 이는, 예를 들어 전사 종결 서열을 갖는 gRNA를 발현함으로써 존재할 수 있다. 이러한 추가의 잔기는, 예를 들어, 종결 서열의 선택에 따라 모두 U 잔기 또는 적어도 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 U 잔기일 수 있다.
개시된 발명에 유용한 적합한 gRNA의 비제한적인 예는 다음을 포함할 수 있다:
서열 번호:64:
서열 번호:70:
서열 번호:64 내지 70의 각각에서, 단일-밑줄로된 서열은 gRNA의 crRN 부분을 나타낸다. 각각의 "N"은 적합한 가이드 서열의 리보뉴클레오티드 염기 (A, U, G 또는 C)를 나타낸다. 소문자의 첫 번째 블록은 tracrRNA 메이트 서열을 나타낸다. 소문자의 두 번째 블록은 gRNA의 tracrRNA 부분을 나타낸다. 이중 밑줄로된 서열은 tracrRNA 메이트 서열과 어닐링하는 tracrRNA 서열 부분에 근접하여 제1 헤어핀을 형성한다. 루프 서열 (GAAA, 서열 번호:43)은 대문자로 표시되며, 이는 각각의 gRNA의 crRNA와 tracrRNA 부분을 작동가능하게 연결한다. 본 명세서의 gRNA의 다른 예는 (i) 이러한 서열과 적어도 약 80%, 85%, 90, 91, 92, 93, 94, 95, 96, 97, 98 또는 99%의 서열 동일성 (이 계산에서 가이드 서열은 제외함)을 갖고 (ii) Cas9 단백질을 특이적으로 표적화하는 gRNA로서 작용하여 표적 DNA 서열에 결합하고, 임의로 닉을 형성하거나 절단할 수 있는 전술한 gRNA의 변이체를 포함한다.
본 명세서에서, gRNA는 또한 가이드 서열 (VT 도메인)에 이어서 Cas 엔도뉴클레아제 인식 (CER) 도메인을 갖는 것으로 특징지어질 수 있다. CER 도메인은 tracrRNA 메이트 서열 다음에 tracrRNA 서열을 포함한다. 본 명세서에서 유용한 CER 도메인의 예는 상기 서열 번호:64 내지 70에 포함되는 것들을 포함한다 (각각의 CER 도메인은 VT 도메인의 N으로 이루어진 서열에 이어지는 서열이다). CER 도메인의 또 다른 적합한 예는 서열 번호:1 (실시예 참조)이며, 이는 5'에서 3' 방향으로 서열 번호:56의 tracrRNA 메이트 서열, 서열 번호:43의 루프 형성 서열 (GAAA) 및 서열 번호:58의 tracrRNA 서열을 포함한다.
개시된 발명의 RGEN의 RNA 성분은 5'-캡 (7-메틸구아닐레이트 [m7G] cap)을 갖지 않는다. 따라서, 본 명세서에서 RNA 성분은 이의 5'-말단에 7-메틸구아닐레이트 (m7G) 캡을 갖지 않는다. 본 명세서에서, RNA 성분은, 예를 들어, 5'-캡 대신에 5'-하이드록실기를 가질 수 있다. 대안적으로, 본 명세서에서 RNA 성분은, 예를 들어, 5'-캡 대신에 5' 포스페이트를 가질 수 있다. 5'-캡핑된 RNA (즉, 5' m7G 캡을 갖는 RNA)가 핵외수송의 대상이기 때문에 RNA 성분이 전사 후 핵에 더 잘 축적될 수 있다고 여겨진다. 본 명세서에서, 캡핑되지 않은 RNA 성분의 바람직한 예는 적합한 gRNA, crRNA 및/또는 tracrRNA를 포함한다. 소정 실시 형태에서, 본 명세서의 RNA 성분은 RNA 성분의 전구체의 5'-말단에서 리보자임 서열에 의한 RNA 자가처리에 의해 5'-캡이 없고 대신에 임의로 5'-하이드록실기를 갖는다 (즉, gRNA와 같은 RNA 성분의 상류에 리보자임 서열을 포함하는 전구체 RNA는 리보자임 서열을 제거하기 위해 리보자임-매개 자가처리를 거침으로써, 5'-캡이 없는 RNA 성분의 하류를 남긴다). 소정 다른 실시 형태에서, 본 명세서의 RNA 성분은 RNA 폴리머라제 III (Pol III) 프로모터로부터의 전사에 의해 생성되지 않는다.
소정 실시 형태에서, 효모는 (ii) RNA 성분을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 추가로 포함한다. 이러한 폴리뉴클레오티드 서열은 효모에 의해 Cas 단백질과 복합체를 형성하여 RGEN을 형성하는 RNA 성분을 발현하는 데 사용된다. 이러한 폴리뉴클레오티드 서열은, 예를 들어, 플라스미드, 효모 인공 염색체 (YAC), 코스미드, 파스미드, 박테리아 인공 염색체 (BAC), 바이러스 또는 선형 DNA (예를 들어, 선형 PCR 생성물) 또는 폴리뉴클레오티드 서열을 통상적이지 않은 효모 세포로 전달하는 데 유용한 임의의 다른 타입의 벡터 또는 구조물의 형태일 수 있다. 이러한 폴리뉴클레오티드 서열은 본 명세서의 효모 세포 에서 일시적으로 존재할 수 있거나 (즉, 게놈으로 통합되지 않음) 안정적으로 존재할 수 있다 (즉, 게놈으로 통합됨). 또한, 이러한 폴리뉴클레오티드 서열은 하나 이상의 적합한 마커 서열 (예를 들어, 선택 또는 표현형 마커)를 포함하거나 포함하지 않을 수 있다.
본 명세서에서, RNA 성분을 발현하기 위해 폴리뉴클레오티드 서열에 포함되는 적합한 프로모터는 통상적이지 않은 효모 세포에서 작동가능하며, 예를 들어, 구성적일 수 있거나 유도성일 수 있다. 소정 측면에서, 프로모터는 단위 시간당 상대적으로 많은 수의 생산 개시를 이끌 수 있는 프로모터 및/또는 효모를 포함하는 효모에서 유전자의 평균 전사 수준보다 높은 전사 수준을 유도하는 프로모터인 강력한 프로모터를 포함할 수 있다.
본 명세서에서 유용한 강력한 프로모터의 예에는 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2012/0252079호 (DGAT2), 제2012/0252093호 (EL1), 2013/0089910호 (ALK2), 제2013/0089911호 (SPS19), 제2006/0019297호 (GPD 및 GPM), 제2011/0059496호 (GPD 및 GPM), 제2005/0130280호 (FBA, FBAIN, FBAINm), 제2006/0057690호 (GPAT) 및 제2010/0068789호 (YAT1)에 개시된 것들이 포함된다. 강력한 프로모터의 다른 적합한 예에는 표 2에 열거된 것들이 포함된다.
[표 2]
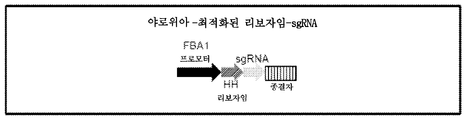
상기 열거된 강력한 프로모터는 야로위아 리폴리티카로부터 유래되지만, 예를 들어, 본 명세서에 개시된 임의의 통상적이지 않은 효모로부터의 이들의 상응하는 프로모터 (예를 들어, 상동체)가 강력한 프로모터로 작용할 수 있다고 여겨진다. 따라서, 강력한 프로모터는, 예를 들어, XPR2, TEF, GPD, GPM, GPDIN, FBA, FBAIN, FBAINm, GPAT, YAT1, EXP1, DGAT2, EL1, ALK2 또는 SPS19 프로모터를 포함할 수 있다. 대안적으로, 전술한 것 중 어느 하나에 상응하는 것과 같은 강력한 프로모터는 다른 유형의 효모 (예를 들어, S. 세레비시아, S. 폼베)로부터 유래될 수 있다 (예를 들어, 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010/0150871호에 개시된 임의의 강력한 프로모터). 본 명세서에서 유용한 강력한 프로모터의 다른 예는 PGK1, ADH1, TDH3, TEF1, PHO5, LEU2 및 GAL1 프로모터 및 본 명세서에 참고로 포함되는 문헌[Velculescu et aL. (Cell 88:243-251)]에 개시된 강력한 효모 프로모터를 포함한다. 본 명세서에서 유용한 강력한 프로모터의 또 다른 예는 서열 번호:12 (야로위아 FBA1 프로모터 서열)을 포함할 수 있다.
소정 실시 형태에서, 본 명세서의 프로모터는 RNA 폴리머라제 II (Pol II) 프로모터를 포함할 수 있다. 상기 열거된 모든 강력한 프로모터는 적합한 예 Pol II 프로모터의 예라고 여겨진다. Pol II 프로모터로부터의 전사는, 예를 들어, 적어도 약 12개의 단백질 (예를 들어, RPB1-RPN12 단백질)의 RNA 폴리머라제 II 복합체를 포함할 수 있다. 본 명세서에서, Pol II 프로모터로부터 전사된 RNA는 전형적으로 5'-캡핑된다 (예를 들어, 5'-말단에 m7G 기를 함유함). 본 명세서의 RNA 성분은 5'-캡을 갖지 않기 때문에, RNA 성분으로부터 5'-캡을 제거하는 방법은 본 명세서의 Pol II 프로모터로부터 발현되는 경우 사용되어야 한다. 본 명세서에서, Pol II-전사된 RNA 성분으로부터 5'-캡을 효과적으로 제거하는 적합한 방법은, 예를 들어, 하나 이상의 리보자임 (하기 참조), 그룹 1 자가-스플라이싱 인트론 및 그룹 2 자가-스플라이싱 인트론의 적절한 사용을 포함한다.
본 명세서에서 RNA 성분을 암호화하는 뉴클레오티드 서열은, 예를 들어, RNA 성분을 암호화하는 서열의 상류에 있는 리보자임을 추가로 암호화할 수 있다. 따라서, 소정 실시 형태에서, 효모는 (ii) 5'에서 3' 방향으로 리보자임과 RNA 성분을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 추가로 포함한다. 이러한 폴리뉴클레오티드 서열로부터 발현된 전사물은 자체 촉매 작용으로 리보자임 서열을 제거하여, 5'-캡을 갖지 않지만 RNA 성분 서열을 포함하는 RNA를 생성한다. 이러한 "자가처리된" RNA는 crRNA 또는 gRNA를 포함할 수 있으며, 예를 들어 Cas 단백질, 예컨대 Cas9와 복합체를 형성하여 RGEN을 형성할 수 있다.
본 명세서에서, 리보자임은, 예를 들어, 망치머리 (HH) 리보자임, 델타 간염 바이러스 (HDV) 리보자임, 그룹 I 인트론 리보자임, RnaseP 리보자임 또는 헤어핀 리보자임일 수 있다. 본 명세서에서, 리보자임의 다른 비제한적인 예는 VS(Varkud satellite) 리보자임, 글루코사민-6-포스페이트 활성화 리보자임 (glmS) 및 CPEB3 리보자임을 포함한다. 문헌[Lilley (BiocheM. SoC. TranS. 39:641-646)]은 리보자임 구조와 활성에 관한 정보를 개시하고 있다. 본 명세서에 사용하기에 적합한 리보자임의 예에는 본 명세서에 참고로 포함되는 EP0707638 및 미국 특허 제6063566호, 제5580967호, 제5616459호 및 제5688670호에 개시된 리보자임이 포함된다.
망치머리 리보자임이 특정 바람직한 실시 형태에서 사용된다. 이러한 유형의 리보자임은, 예를 들어, 본 명세서에 참고로 포함되는 문헌[Hammann et aL. (RNA 18:871-885)]에 개시된 바와 같은 타입 I, 타입 II 또는 타입 III 망치머리 리보자임일 수 있다. 본 명세서에서 적절하게 사용될 수 있는, 망치머리 리보자임을 암호화하는 DNA를 동정하는 다수의 방법이 문헌[Hammann et al.]에 개시되어 있다. 본 명세서에서, 망치머리 리보자임은, 예를 들어, 바이러스, 바이로이드, 식물 바이러스 위성 RNA, 원핵생물 (예를 들어, 고세균, 시아노박테리아, 아시도박테리아) 또는 진핵생물, 예컨대 식물 (예를 들어, 아라비돕시스 탈리아나(Arabidopsis thaliana), 카네이션), 원생생물 (예를 들어, 아메바, 유글레나류), 진균류 (예를 들어, 아스퍼질러스, 야로위아 리폴리티카), 양서류 (예를 들어, 영원(newt), 개구리), 주혈흡충류(schistosome), 곤충 (예를 들어, 귀뚜라미), 연체 동물, 포유류 (예를 들어, 마우스, 인간) 또는 선충류로부터 유래될 수 있다.
본 명세서에서, 망치머리 리보자임은 전형적으로 보존된 서열의 짧은 링커에 의해 이격된, 각각 나선구조 I, II 및 III으로 지칭되는 3가지의 염기쌍 형성된 나선 구조를 포함한다. 망치머리 리보자임의 3가지 타입 (I 내지 III)은 일반적으로 리보자임의 5' 및 3' 말단이 어느 나선 구조에 포함되는 지에 기초한다. 예를 들어, 망치머리 리보자임 서열의 5' 및 3' 말단에 의해 스템 I이 형성된다면, 이는 타입 I 망치머리 리보자임으로 지칭될 수 있다. 3가지 가능한 위상학적(topological) 타입 중에서, 타입 I은 원핵생물, 진핵생물 및 RNA 식물 병원균의 게놈에서 발견될 수 있는 반면, 타입 II 망치머리 리보자임은 원핵생물에서만 기재되어 왔으며, 타입 III 망치머리 리보자임은 주로 식물, 식물 병원균 및 원핵생물에서 발견된다. 소정 실시 형태에서, 망치머리 리보자임은 타입 I 망치머리 리보자임이다.
소정 실시 형태에서, 망치머리 리보자임을 암호화하는 서열은 적어도 약 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140 또는 150 (또는 40 내지 150 사이의 임의의 정수)개의 뉴클레오티드, 40 내지 100개의 뉴클레오티드 또는 40 내지 60개의 뉴클레오티드를 포함할 수 있다.
망치머리 리보자임을 암호화하는 서열은 RNA 성분을 암호화하는 서열의 상류에 있다. 본 명세서에서, 망치머리 리보자임을 암호화하는 서열은, 예를 들어, RNA 성분의 가이드 서열을 암호화하는 서열의 5' 부근이거나, 이의 5'의 적어도 2, 3, 4, 5, 6, 7, 8, 9 또는 10개의 뉴클레오티드일 수 있다 (예를 들어, 가이드 서열은 crRNA 또는 gRNA의 것일 수 있다). 망치머리 리보자임의 처음 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 또는 15번째의 리보뉴클레오티드는 전형적으로 망치머리 리보자임 서열의 바로 하류에 있는 서열의 리보뉴클레오티드의 처음 동일한 번호에 각각 상보적이어야 한다. 예를 들어, 본 명세서에서, 폴리뉴클레오티드 서열이 RNA 성분의 가이드 서열의 바로 상류에서 망치머리 리보자임 서열을 포함하는 RNA를 암호화하는 경우, 리보자임의 처음 6개의 리보뉴클레오티드는, 예를 들어, 가이드 서열의 처음 6개의 리보뉴클레오티드에 상보적일 수 있다. 이러한 예에서, 망치머리 리보자임은 가이드 서열의 첫 번째 위치의 바로 상류에서 RNA 전사물을 절단할 것이다 (또는 또 다른 방식으로 말하면, 망치머리 리보자임은 리보자임 서열의 바로 하류에서 RNA 전사물을 절단할 것이다). 이러한 논리는 다른 전술한 예시적인 실시 형태에 유사하게 적용된다. 예를 들어, 본 명세서에서, 폴리뉴클레오티드 서열이 RNA 성분의 가이드 서열의 상류에 있는 8개의 잔기인 망치머리 리보자임 서열을 포함하는 RNA를 암호화하는 경우 (예를 들어, 8-잔기 스페이서 서열이 있음), 예를 들어, 리보자임의 처음 6개의 리보뉴클레오티드는 리보자임 서열의 3' 부근의 6개의 리보뉴클레오티드에 상보적일 수 있다. 이러한 예에서, 망치머리 리보자임은 리보자임 서열의 바로 하류에서 RNA 전사물을 절단할 것이다. 또 다른 예로서, 본 명세서에서, 폴리뉴클레오티드 서열이 RNA 성분의 가이드 서열의 바로 상류에서 망치머리 리보자임 서열을 포함하는 RNA를 암호화하는 경우, 리보자임의 처음 10개의 리보뉴클레오티드는, 예를 들어, 가이드 서열의 처음 10개의 리보뉴클레오티드에 상보적일 수 있다. 이러한 예에서, 망치머리 리보자임은 가이드 서열의 첫 번째 위치의 바로 상류에서 RNA 전사물을 절단할 것이다 (또는 또 다른 방식으로 말하면, 망치머리 리보자임은 리보자임 서열의 바로 하류에서 RNA 전사물을 절단할 것이다).
망치머리 리보자임 서열의 예를 다음과 같이 나타낼 수 있다:
(서열 번호:15, N은 A, U, C 또는 G일 수 있다; 실시예 참조). 서열 번호:15의 처음 6개의 잔기는 본 명세서의 DNA 폴리뉴클레오티드로부터 발현된 RNA 전사물에서 서열 번호:15의 바로 다음에 오는 (예를 들어, 본 명세서에 개시된 crRNA 또는 gRNA의 가이드 서열의) 처음 6개의 잔기에 상보적이도록 (에 어닐링되도록) 설계될 수 있다. 리보자임은 서열 번호:15 바로 다음의 전사물을 절단할 것이다. 서열 번호:15를 바로 뒤따르는 서열 잔기와 어닐링하기 위해 서열 번호:15가 6개의 잔기 ("N")를 갖는 것으로 도시되어 있지만, 이러한 목적을 위한 이러한 리보자임의 시작에는 5 내지 15개의 "N" 잔기가 있을 수 있다. 서열 번호:15를 포함하는 RNA 전사물에 있어서, (i) 망치머리 리보자임의 나선 구조 I은 전사물에서 서열 번호:15 바로 다음의 처음 6개의 잔기와 N 잔기의 어닐링에 의해 형성될 것이고, (ii) 나선 구조 II는 단일-밑줄로 나타낸 상보적인 서열의 어닐링에 의해 형성될 것이며, (iii) 나선 구조 III은 이중-밑줄로 나타낸 상보적인 서열의 어닐링에 의해 형성될 것이라는 점에 유의해야 한다. 따라서, 소정 실시 형태에서, 망치머리 리보자임은 (i) 서열 번호:15와 적어도 약 80%, 85%, 90, 91, 92, 93, 94, 95, 96, 97, 98 또는 99%의 서열 동일성 (이 계산에서 "N" 서열은 제외함)을 갖고 (ii) 서로 어닐링되어 나선 구조 II와 III (나선 구조 I은 "N" 잔기의 적절한 선택으로 형성됨)을 형성하는 서열 번호:15의 단일 밑줄로된 영역 및 이중 밑줄로된 영역과 정렬되는 영역을 갖는 서열 번호:15의 변이체일 수 있다.
서열 번호:15 및 (상기) 이들의 다양한 실시 형태에 연결될 수 있는 서열의 예에는 서열 번호:64 내지 70 중 하나를 포함하는 gRNA가 포함된다.
본 명세서에서, RNA 성분 (본 명세서에서 "리보자임-RNA 성분 카세트")에 연결된 5' 망치머리 리보자임을 포함하는 RNA 서열을 암호화하는 DNA 폴리뉴클레오티드는 망치머리 리보자임 서열로 바로 시작되는 5'-말단을 갖는 전사물의 전사를 유도하도록 설계될 수 있다 (즉, 전사 시작 부위에서 리보자임 서열이 시작된다). 대안적으로, DNA 폴리뉴클레오티드는 리보자임-RNA 성분 카세트의 상류에 비-리보자임(non-ribozyme) 서열을 갖는 전사물의 전사를 유도하도록 설계될 수 있다. 이러한 5' 비-리보자임 전사물 서열은, 예를 들어, 단지 수 개의 뉴클레오티드 (1 내지 10)만큼 짧거나, 최대 5000 내지 20000개의 뉴클레오티드 만큼 길 수도 있다 (이러한 리보자임의 5' 서열은 리보자임이 그 자체를 RNA 성분으로부터 절단하는 경우, RNA 성분으로부터 제거된다).
소정 실시 형태에서, 리보자임-RNA 성분 카세트를 포함하는 DNA 폴리뉴클레오티드는 RNA 성분 서열의 하류에 적합한 전사 종결 서열을 포함할 수 있다. 본 명세서에서 유용한 전사 종결 서열의 예가 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2014/0186906호에 개시되어 있다. 예를 들어, S. 세레비시아 Sup4 유전자 전사 종결 서열 (예를 들어, 서열 번호:8)이 사용될 수 있다. 이러한 실시 형태는 전형적으로 리보자임-RNA 성분 카세트의 하류에 위치한 리보자임 서열을 포함하지 않는다. 또한, 이러한 실시 형태는 전형적으로 종결 서열의 선택에 따라 RNA 성분 서열의 말단에 이어지는 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30개 또는 그 이상의 잔기를 포함한다. 이러한 추가의 잔기는, 예를 들어, 종결 서열의 선택에 따라 모두 U 잔기 또는 적어도 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 U 잔기일 수 있다. 대안적으로, 리보자임 서열 (예를 들어, 망치머리 또는 HDV 리보자임)은 (예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 그 이상의 뉴클레오티드) RNA 성분 서열의 3'일 수 있으며; 이러한 실시 형태에서 RNA 성분 서열은 상류 및 하류 리보자임에 의해 플랭킹된다. 3' 리보자임 서열은 RNA 성분 서열로부터 그 자체를 절단하도록 적절하게 위치될 수 있고; 이러한 절단은, 예를 들어, RNA 성분 서열의 말단에서 정확하게, 또는 RNA 성분 서열의 말단에 이어지는 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15개 또는 그 이상의 잔기를 갖도록 전사물을 종료시킬 것이다.
소정 실시 형태에서, DNA 폴리뉴클레오티드는 (ii) 하나 초과의 리보자임-RNA 성분 카세트 (즉, 탠덤 카세트)를 포함하는 서열에 (i) 작동가능하게 연결된 프로모터를 포함할 수 있다. 이러한 DNA 폴리뉴클레오티드로부터 발현된 전사물은, 예를 들어, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 그 이상의 리보자임-RNA 성분 카세트를 가질 수 있다. 3' 리보자임 서열은 전사물 서열의 하류로부터 RNA 성분의 절단 및 분리를 허용하기 위해 각각의 RNA 성분 서열 다음에 임의로 (예를 들어, 상기와 같이) 포함될 수 있다. 이러한 실시 형태에서 각각의 RNA 성분은 전형적으로 본 명세서의 RGEN을 특유의 DNA 표적 부위 유도하도록 설계된다. 따라서, 이러한 DNA 폴리뉴클레오티드는, 예를 들어 통상적이지 않은 효모에서 동시에 다수의 상이한 표적 부위를 표적화하는데 적절하게 사용될 수 있고; 이러한 사용은 임의로 다중화 방법으로 특징지어질 수 있다. 3' 리보자임에 연결된 RNA 성분에 연결된 5' 망치머리 리보자임은 본 명세서에서 "리보자임-RNA 성분-리보자임 카세트"로 지칭될 수 있다. 본 명세서에서, 탠덤 리보자임-RNA 성분-리보자임 카세트를 포함하는 전사물을 발현하기 위한 DNA 폴리뉴클레오티드는 각각의 카세트 (예를 들어, 비암호화 스페이서 서열) 사이에 약 10, 20, 30, 40, 50, 60, 70, 80, 90, 100개 또는 그 이상의 뉴클레오티드가 존재하도록 설계될 수 있다. 각각의 카세트 사이의 거리는 동일하거나 상이할 수 있다.
상기 실시 형태 중 일부는 망치머리 리보자임 서열에 관하여 기술하고 있지만, 이러한 실시 형태는 또한 이에 따라 망치머리 리보자임 서열 대신에 본 명세서의 임의의 다른 리보자임 서열 (예를 들어, HDV 리보자임)의 관점에서 특징지어질 수 있다. 당업자는 이러한 다른 리보자임 서열을 특정 부위에서 절단하도록 배치하는 방법을 이해할 것이다.
소정 실시 형태에서, 효모는 (ii) Cas 단백질 (예를 들어, Cas9)을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 추가로 포함한다. 이러한 폴리뉴클레오티드 서열은 효모에 의해 RNA 성분과 복합체를 형성하여 RGEN을 형성하는 Cas 단백질을 발현하는 데 사용된다. 이러한 폴리뉴클레오티드 서열은, 예를 들어, 플라스미드, YAC, 코스미드, 파스미드, BAC, 바이러스 또는 선형 DNA (예를 들어, 선형 PCR 생성물), 또는 폴리뉴클레오티드 서열을 통상적이지 않은 효모 세포로 전달하는 데 유용한 임의의 다른 타입의 벡터 또는 구조물의 형태일 수 있다. 본 명세서에 개시된 임의의 Pol II 프로모터가, 예를 들어 사용될 수 있다. 따라서, RNA 성분을 발현하기 위한 DNA 폴리뉴클레오티드 서열에 관련하여 상기 개시된 임의의 특징은 Cas 단백질을 발현하기 위한 DNA 폴리뉴클레오티드 서열에 적용될 수 있다. 이러한 폴리뉴클레오티드 서열은 본 명세서의 효모 세포 에서 일시적으로 존재할 수 있거나 (즉, 게놈으로 통합되지 않음) 안정적으로 존재할 수 있다 (즉, 게놈으로 통합됨). 다른 측면에서, 효모는 Cas 단백질을 발현하기 위한 DNA 폴리뉴클레오티드에 더하여, RNA 성분을 발현하기 위한 DNA 폴리뉴클레오티드를 가질 수 있다 (예를 들어, 상기한 바와 같음). 이러한 DNA 폴리뉴클레오티드는 모두 효모에 안정적이거나 일시적일 수 있거나; Cas 단백질을 발현하기 위한 DNA 폴리뉴클레오티드는 안정적이고 RNA 성분을 발현하기 위한 DNA 폴리뉴클레오티드는 일시적일 수 있다 (또는 그 반대).
대안적으로, DNA 폴리뉴클레오티드 서열은 효모 세포에서 RGEN을 제공하기 위한 Cas 단백질과 적합한 RNA 성분 모두를 발현하는 것일 수 있다. 이러한 DNA 폴리뉴클레오티드는, 예를 들어, (i) (RGEN의) RNA 성분 (RNA 성분 카세트)을 암호화하는 뉴클레오티드 서열에 작동가능하게 연결된 프로모터 및 (ii) Cas 단백질 (예를 들어, Cas9) (Cas 카세트)을 암호화하는 뉴클레오티드 서열에 작동가능하게 연결된 프로모터를 포함할 수 있다. Cas 단백질 또는 RNA 성분을 발현하기 위한 DNA 폴리뉴클레오티드에 관련하여 상기 기재된 임의의 특징은, 예를 들어, 통상적이지 않은 효모 세포에서 Cas 단백질과 적합한 RNA 성분을 모두 발현하기 위한 DNA 폴리뉴클레오티드 서열에 적용될 수 있다. 또한, 본 명세서에 개시된 임의의 Cas 단백질과 RNA 성분 (예를 들어, crRNA 또는 gRNA)은 이러한 DNA 폴리뉴클레오티드 서열로부터 발현될 수 있다. 소정 실시 형태에서, 하나 이상의 RNA 성분 및/또는 Cas 카세트는 DNA 폴리뉴클레오티드 서열 내에 포함될 수 있다. 다른 측면에서, 하나 이상의 RNA 성분은 상기한 바와 같이 탠덤으로 발현될 수 있다. Cas 카세트 및 RNA 카세트에서 사용된 프로모터는 동일하거나 상이할 수 있다. 이러한 DNA 폴리뉴클레오티드 서열이 통상적이지 않은 효모와 통상적인 효모 모두에서 RGEN을 발현하는 데 유용할 것으로 생각된다.
개시된 발명은 또한 통상적이지 않은 효모 내의 염색체 또는 에피솜의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법에 관한 것이다. 이러한 방법은 효모의 핵에 5'-캡을 갖지 않는 적어도 하나의 RNA 성분을 포함하는 RGEN을 제공하는 단계를 포함하며, 여기서 RNA 성분은 표적 부위 서열에 상보적인 서열을 포함하고, RGEN은 표적 부위 서열의 전부 또는 일부에 결합하고, 임의로 절단한다.
이러한 표적화 방법은, 예를 들어, 방법의 특징 (예를 들어, 효모 타입, RGEN, RNA 성분, 등)각각에 관한 상기 개시된 실시 형태 또는 하기 실시예 중 어느 하나를 사용하여 실시될 수 있다. 따라서, 상기 또는 실시예에서 개시된 특징 중 어느 하나 또는 이들 특징의 임의의 조합이 적절하게 사용되어, 본 명세서의 표적화 방법의 실시 형태를 특징지을 수 있다. 다음의 표적화 방법 특징은 예이다.
본 명세서의 표적화 방법의 소정 실시 형태에서, 통상적이지 않은 효모는 다음과 같은 속의 임의의 구성원일 수 있다: 야로위아, 피키아, 쉬반니오마이세스, 클루이베로마이세스, 아르술라, 트리코스포론, 칸디다, 유스틸라고, 토룰롭시스, 자이고사카로마이세스, 트리고놉시스, 크립토코커스, 로도토룰라, 파피아, 스포로볼로마이세스 및 파치솔렌. 야로위아 리폴리티카는 본 명세서에 적합한 야로위아 효모이다. 표적화 방법에서 유용한 통상적이지 않은 효모의 다른 비제한적인 예가 본 명세서에 개시되어 있다.
본 명세서의 표적화 방법에 사용하기에 적합한 RGEN은 타입 I, II 또는 III 크리스퍼 시스템의 Cas 단백질을 포함할 수 있다. 소정 실시 형태에서, Cas9 단백질, 예컨대 스트렙토코커스 Cas9가 사용될 수 있다. 표적화 방법에 사용하기에 적합한 스트렙토코커스 Cas9 단백질의 예에는 S. 피오제네스, S. 써모필러스, S. 뉴모니애, S. 아갈락티아, S. 파라상귀니스, S. 오랄리스, S. 살리바리우스, S. 마카카, S. 디스갈락티애, S. 안지노서스, S. 콘스텔라투스, S. 슈도포르시누스 또는 S. 뮤탄스의 Cas9 단백질로부터 유래된 아미노산 서열을 포함하는 Cas9 단백질이 포함된다. 본 명세서의 표적화 방법에 유용한 RGEN 및 Cas9 단백질의 비제한적인 예가 본 명세서에 개시되어 있다. 예를 들어, DNA 표적 서열에서 하나 또는 두 가닥을 절단할 수 있는 RGEN이 사용될 수 있다.
본 명세서의 표적화 방법에 사용하기 위한 RGEN의 RNA 성분은, 예를 들어, tracrRNA에 작동가능하게 연결되거나 융합된 crRNA를 포함하는 gRNA를 포함할 수 있다. 본 명세서에 개시된 바와 같은 임의의 crRNA 및/또는 tracrRNA (및/또는 이들의 부분, 예컨대 tracrRNA 메이트 서열 또는 tracrRNA 5'-말단 서열)는, 예를 들어, gRNA에 포함될 수 있다. 또한, 본 명세서에 개시된 임의의 gRNA가, 예를 들어, 표적화 방법에서 사용될 수 있다.
PAM (프로토스페이서-인접 모티프) 서열은, 예를 들어, 표적 부위 서열에 인접할 수 있다. 본 명세서의 표적화 방법의 소정 실시 형태에서, PAM 서열은 결과적으로 RNA 성분 가이드 서열에 상보적인 표적 부위의 가닥에 상보적인 표적 부위 서열로부터 바로 하류에, 또는 이의 2 또는 3개의 뉴클레오티드 하류에 있다. RGEN이 RNA 성분과 복합체를 형성한 핵산 내부 분해 방식으로 활성인 Cas9 단백질인 본 명세서의 실시 형태에서, Cas9는 RNA 성분에 의해 유도된 바와 같이 표적 서열에 결합하고, PAM 서열의 상류 세 번째 뉴클레오티드 위치의 5' 부근에서 두 가닥을 절단한다. 적합한 PAM 서열의 예에는 S. 피오제네스 (NGG [서열 번호:47]) 및 S. 써모필러스 (NNAGAA [서열 번호:48]) PAM 서열이 포함되며, 이는 각각 각각의 종으로부터 유래된 Cas9 단백질로 표적으로 하는데 사용될 수 있다. 또한, 본 명세서에 개시된 임의의 PAM 서열이, 예를 들어, 표적화 방법에서 사용될 수 있다.
본 명세서의 표적화하는 방법의 소정 실시 형태에서, 효모는 (ii) RNA 성분을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 추가로 포함한다. RNA 성분이 DNA 폴리뉴클레오티드로부터 전사되기 때문에, 이러한 DNA 폴리뉴클레오티드를 사용하여 RGEN의 RNA 성분이 효모의 핵에 제공될 수 있다. 효모 핵에서 (RGEN의) RNA 성분을 발현하기 위한 적합한 DNA 폴리뉴클레오티드 서열의 예가 본 명세서에 개시되어 있다. 본 명세서에 개시된 바와 같은 임의의 프로모터가, 예를 들어, 강력한 프로모터 및/또는 Pol II 프로모터 서열을 포함하는 것과 같은 이러한 DNA 폴리뉴클레오티드 서열에서 사용될 수 있다. 소정 실시 형태에서, RNA 성분을 암호화하는 DNA 폴리뉴클레오티드가 Cas 단백질을 발현하도록 이미 조작된 (예를 들어, 안정한 Cas 발현) 효모에서 RNA 성분을 제공하는 데 사용될 수 있다.
본 명세서에서 RNA 성분을 암호화하는 뉴클레오티드 서열은, 예를 들어, RNA 성분을 암호화하는 서열의 상류에 있는 리보자임을 추가로 암호화할 수 있다. 따라서, 본 명세서의 표적화 방법의 소정 실시 형태에서, 효모는 (ii) 5'에서 3' 방향으로 리보자임과 RNA 성분을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 포함할 수 있다. RNA 성분이 DNA 폴리뉴클레오티드로부터 전사되기 때문에, 이러한 DNA 폴리뉴클레오티드를 사용하여 RGEN의 RNA 성분을 효모의 핵에 제공할 수 있다. 본 명세서에서, 리보자임은, 예를 들어, 망치머리 리보자임, 델타 간염 바이러스 (HDV) 리보자임, 그룹 I 인트론 리보자임, RnaseP 리보자임 또는 헤어핀 리보자임일 수 있다. 본 명세서에 개시된 바와 같은 임의의 리보자임 및 본 명세서에 개시된 바와 같은 임의의 RNA 성분에 연결된 리보자임을 암호화하는 폴리뉴클레오티드 서열이, 예를 들어, 표적화 방법에서 사용될 수 있다.
본 명세서의 표적화 방법의 소정 실시 형태에서, 효모는 (ii) Cas 단백질 (예를 들어, Cas9)을 암호화하는 뉴클레오티드 서열에 (i) 작동가능하게 연결된 프로모터를 포함하는 DNA 폴리뉴클레오티드 서열을 추가로 포함한다. 이러한 DNA 폴리뉴클레오티드을 사용하여 효모에서 RGEN의 Cas 단백질 성분을 제공할 수 있다. 효모에서 (RGEN의) Cas 단백질 성분을 발현하기 위한 적합한 DNA 폴리뉴클레오티드 서열의 예가 본 명세서에 개시되어 있다. 본 명세서에 개시된 바와 같은 임의의 프로모터가, 예를 들어, 강력한 프로모터을 포함하는 것과 같은 이러한 DNA 폴리뉴클레오티드 서열에서 사용될 수 있다.
DNA 표적 부위 서열에 또는 그 부근의 서열에 상동성인 적어도 하나의 서열을 포함하는 공여 폴리뉴클레오티드는 또한 표적화 방법의 소정 실시 형태에서 (표적 부위에 서열에서 닉을 형성하거나 절단하는 RGEN으르 제공하는 것과 함께) 효모에 제공될 수 있다. 적합한 예에는 상동성 암을 갖는 공여 폴리뉴클레오티드가 포함된다. 본 명세서에 개시된 바와 같은 임의의 공여 폴리뉴클레오티드가 표적화 방법에서 사용될 수 있다. 이러한 방법의 이러한 실시 형태는 전형적으로 공여 폴리뉴클레오티드와 표적 서열 (표적 서열의 RGEN-매개 닉킹 또는 절단 후) 사이의 HR을 포함하며; 따라서, 이러한 방법은 임의로 통상적이지 않은 효모에서 HR을 수행하는 방법으로도 지칭될 수 있다. 이러한 방법으로 수행될 수 있는 HR 전략의 예가 본 명세서에 개시되어 있다. 효모 세포에서 표적화하기 위한 공여 DNA 폴리뉴클레오티드의 적합한 양은 효모 세포 당 적어도 약 300, 400, 500, 600, 700 또는 800개의 공여 DNA 분자일 수 있다.
RGEN 성분을 발현하기 위해 본 명세서에 기재된 DNA 폴리뉴클레오티드를 포함하는 임의의 구조물 또는 벡터는 임의의 표준 기술에 의해 통상적이지 않은 효모 세포로 도입될 수 있다. 이러한 기술에는, 예를 들어 형질전환 (예를 들어, 아세트산리튬 형질전환 (문헌[Methods in Enzymology, 194:186-187]), 바이올리스틱 충격(biolistic impact), 전기천공 및 미세주입이 포함된다. 예로서, 본 명세서에 참고로 포함되는 미국 특허 제4880741호 및 제5071764호 및 문헌[Chen et aL. (AppL. MicrobioL. BiotechnoL. 48:232-235)]은 야로위아 리폴리티카에 대한 DNA 전달 기술을 기재한다.
본 명세서의 표적화 방법은 통상적이지 않은 효모에서 삽입-결실을 생성하기 위한 목적으로 수행될 수 있다. 이러한 방법은 상기 개시된 바와 같이 수행될 수 있지만, 표적 DNA 부위 또는 그 부근에서 HR을 행할 수 있는 공여 DNA 폴리뉴클레오티드를 추가로 제공하지 않는다. (즉, 이러한 방법에서는 NHEJ가 유도된다). 생성될 수 있는 삽입-결실의 예가 본 명세서에 개시되어 있다. 삽입-결실의 크기는, 예를 들어, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10개 또는 그 이상의 염기일 수 있다. 소정 실시 형태에서 삽입-결실은 적어도 약 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140 또는 150개의 염기와 같이 심지어 더 클 수 있다. 또 다른 실시 형태에서, 삽입 또는 결실은 적어도 약 500, 750, 1000 또는 1500개의 염기일 수 있다. 소정 실시 형태에서 삽입-결실을 생성하고자 시도하는 경우, 표적 부위 서열에 단일 염기 치환이 대신 형성될 수 있다. 따라서, 본 명세서의 표적화 방법은, 예를 들어, 단일 염기 치환을 생성하기 위한 목적으로 수행될 수 있다.
본 명세서의 삽입-결실 형성을 목적으로 하는 표적화 방법의 소정 실시 형태에서, 통상적이지 않은 효모 (예를 들어, 야로위아 리폴리티카)에서 삽입-결실의 형성의 빈도는 통상적인 효모, 예컨대 S. 세레비시아에서 동일하거나 유사한 표적화 전략을 사용하여 관찰되는 것보다 상당히 더 높다. 예를 들어, 삽입-결실의 형성의 빈도는 통상적인 효모에서 약 0.0001 내지 0.001일 수 있는 반면 (문헌[DiCarlo et al., Nucleic Acids Res. 41:4336-4343]), 본 명세서의 통상적이지 않은 효모에서의 빈도는 적어도 약 0.05, 0.10, 0.15, 0.20, 0.25, 0.30, 0.35, 0.40, 0.45, 0.50, 0.55, 0.60, 0.65, 0.70, 0.75 또는 0.80일 수 있다. 따라서, 본 명세서의 통상적이지 않은 효모에서 삽입-결실의 형성의 빈도는 통상적인 효모에서 동일하거나 유사한 Cas-매개 표적화 전략을 사용하여 발견된 것보다, 예를 들어, 적어도 약 50, 100, 250, 500, 750, 1000, 2000, 4000 또는 8000배 더 높을 수 있다. 이러한 실시 형태의 소정 측면은 공여 DNA를 포함하지 않고/않거나 RGEN 성분 (Cas 및 적합한 RNA 성분)이 동일한 벡터/구조물로부터 발현되는 표적화 방법에 관한 것일 수 있다.
본 명세서의 표적화 방법은, 예를 들어, 방법에서 둘 이상의 DNA 표적 부위가 표적화되는 방식으로 수행될 수 있다. 이러한 방법은 본 명세서에 개시된 바와 같은 탠덤 리보자임-RNA 성분 카세트 (예를 들어, 탠덤 리보자임-RNA 성분-리보자임 카세트)를 포함하는 전사물을 발현하는 DNA 폴리뉴클레오티드를 효모에 제공하는 단계를 포함할 수 있다. 이러한 방법은 동일 서열 (예를 들어, 프로모터 또는 오픈 리딩 프레임)에서 매우 가까운 DNA 부위 및/또는 서로 멀리 있는 (예를 들어, 상이한 유전자 및/또는 염색체) 부위를 표적화할 수 있다. 이러한 방법은 표적화의 원하는 결과에 따라 적합한 공여 DNA 폴리뉴클레오티드를 사용하거나 (HR의 경우) 사용하지 않고 (삽입-결실 및/또는 염기 치환으로 유도되는 NHEJ의 경우) 수행될 수 있다
소정 실시 형태에서 표적화 방법이 수행되어 단백질 또는 비암호화 RNA를 암호화하는 하나 이상의 DNA 폴리뉴클레오티드 서열을 파괴할 수 있다. 파괴의 표적이 될 수 있는 이러한 서열의 예는 마커 (즉, 마커 유전자)를 암호화하는 것이다. 본 명세서의 마커의 비제한적인 예에는 스크리닝가능한 마커와 선택가능한 마커가 포함된다. 본 명세서에서 스크리닝가능한 마커는 적절한 조건 하에서 효모를 시각적으로 상이하게 만드는 것일 수 있다. 스크리닝가능한 마커의 예에는 베타-글루쿠로니다제 (GUS), 베타-갈락토시다제 (lacZ) 및 형광 단백질 (예를 들어, GFP, RFP, YFP, BFP)을 암호화하는 폴리뉴클레오티드가 포함된다. 본 명세서에서 선택가능한 마커는 효모를 선택적 제제 또는 선택적 환경에 내성으로 만드는 것일 수 있다. 선택가능한 마커의 예는 효모가 각각 외인성으로 공급된 히스티딘, 류신, 트립토판, 메티오닌 또는 우라실의 부재 하에 생존할 수 있게 해주는 영양요구성 마커, 예컨대 HIS3, LEU2, TRP1, MET15 또는 URA3이다. 선택가능한 마커의 다른 예는 효모에 하이그로마이신 B, 누오르세오트리신(nourseothricin), 플레오마이신, 퓨로마인신 또는 네오마이신 (예를 들어, G418)에 대한 내성을 부여하는 것들과 같은 항생제 (항진균제)-저항 마커이다.
소정 실시 형태에서 마커를 파괴하는 적어도 하나의 목적은 마커 재활용(marker recycling)일 수 있다. 마커 재활용은, 예를 들어, (i) 효모를 마커 및 이종 DNA 서열로 형질전환시키는 단계, (ii) 마커 및 이종 DNA 서열으로 형질전환된 효모 (여기서, 마커-선택가능한 효모는 전형적으로 이종 DNA 서열을 함유할 확률이 더 높음)를 선택하는 단계, (iii) 마커를 파괴한 후 단계 (i) 내지 (iii)을 필요한 만큼 많이 반복하여 (동일한 마커를 사용하지만, 각각의 사이클은 상이한 이종 DNA 서열을 사용함) 효모를 다수의 이종 DNA 서열로 형질전환시키는 단계를 포함하는 공정이다. 이러한 공정에서 하나 이상의 이종 서열은 공여 폴리뉴클레오티드의 형태로 마커 그 자체를 포함할 수 있다 (예를 들어, 특정 유전자좌를 표적화기 위한 상동성 암에 의해 플랭킹된 마커). 본 명세서의 마커 재활용 공정의 예에는 통상적이지 않은 효모, 예컨대 야로위아 리폴리티카에서 마커로서 URA3을 사용하는 것들이 포함된다.
본 명세서에 개시된 조성물 및 방법의 비제한적인 예는 다음과 같다:
1.
5'-캡을 갖지 않는 적어도 하나의 RNA 성분을 포함하는 적어도 하나의 RNA-유도 엔도뉴클레아제 (RGEN)를 포함하는 통상적이지 않은 효모로서, 여기서 RNA 성분은 효모 내의 염색체 또는 에피솜 상에 표적 부위 서열에 상보적인 서열을 포함하고, RGEN은 표적 부위 서열의 전부 또는 일부에 결합할 수 있는, 통상적이지 않은 효모.
2.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 1의 통상적이지 않은 효모.
3.
상기 효모가 야로위아, 피키아, 쉬반니오마이세스, 클루이베로마이세스, 아르술라, 트리코스포론, 칸디다, 유스틸라고, 토룰롭시스, 자이고사카로마이세스, 트리고놉시스, 크립토코커스, 로도토룰라, 파피아, 스포로볼로마이세스 및 파치솔렌으로 이루어진 군으로부터 선택되는 속의 구성원인, 실시 형태 1의 효모.
4.
RGEN이 크리스퍼(CRISPR; 규칙적으로 사이 간격을 두고 분포하는 짧은 회문구조 반복 서열)-관련 (Cas) 단백질-9 (Cas9) 아미노산 서열을 포함하는, 실시 형태 1의 효모.
5.
Cas9 단백질이 스트렙토코커스 Cas9 단백질인, 실시 형태 4의 효모.
6.
RNA 성분이 트랜스-활성화 크리스퍼 RNA (tracrRNA)에 작동가능하게 연결된 크리스퍼 RNA (crRNA)를 포함하는 가이드 RNA (gRNA)를 포함하는, 실시 형태 4의 효모.
7.
PAM (프로토스페이서-인접 모티프) 서열이 표적 부위 서열에 인접한, 실시 형태 4의 효모.
8.
적어도 하나의 뉴클레오티드 서열에 작동가능하게 연결된 프로모터를 포함하는 폴리뉴클레오티드 서열을 포함하는 통상적이지 않은 효모로서, 여기서 상기 뉴클레오티드 서열은 RNA 성분을 암호화하는 DNA 서열의 상류에 리보자임을 암호화하는 DNA 서열을 포함하고, 상기 RNA 성분은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열에 상보적인 가변 표적화 도메인을 포함하며, RNA 성분은 표적 부위 서열의 전부 또는 일부에 결합할 수 있는 RNA-유도 엔도뉴클레아제 (RGEN)를 형성할 수 있는, 통상적이지 않은 효모.
9.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 8의 통상적이지 않은 효모.
10.
뉴클레오티드 서열로부터 전사된 RNA가 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하며, 상기 RNA 성분이 5' 캡을 포함하지 않는, 실시 형태 8의 통상적이지 않은 효모.
11.
리보자임이 망치머리 리보자임, 델타 간염 바이러스 리보자임, 그룹 I 인트론 리보자임, RnaseP 리보자임 또는 헤어핀 리보자임인, 실시 형태 10의 통상적이지 않은 효모.
12.
뉴클레오티드 서열로부터 전사된 RNA가 5' 캡이 없는 리보자임-RNA 성분 융합 분자를 생성하기 위해 자체 촉매 작용으로 리보자임을 제거하지 않는, 실시 형태 8의 통상적이지 않은 효모.
13.
리보자임이 HDV 리보자임인, 실시 형태 12의 통상적이지 않은 효모.
14.
프로모터가 강력한 프로모터인, 실시 형태 8의 통상적이지 않은 효모.
15.
프로모터가 Pol II 프로모터 서열을 포함하는, 실시 형태 8의 통상적이지 않은 효모.
16.
통상적이지 않은 효모 내의 염색체 또는 에피솜의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법으로서, 상기 방법은 상기 효모에 5'-캡을 갖지 않는 적어도 하나의 RNA 성분을 포함하는 RGEN을 제공하는 단계를 포함하며, 여기서 RNA 성분은 표적 부위 서열에 상보적인 서열을 포함하고, RGEN은 표적 부위 서열의 전부 또는 일부에 결합하는, 방법.
17.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 16의 방법.
18.
통상적이지 않은 효모 내의 염색체 또는 에피솜의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법으로서, 상기 방법은 상기 효모에 적어도 하나의 리보자임-RNA 성분 융합 분자를 포함하는 RGEN을 제공하는 단계를 포함하며, 여기서 RNA 성분은 표적 부위 서열에 상보적인 서열을 포함하고, RGEN은 표적 부위 서열의 전부 또는 일부에 결합하는, 방법.
19.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 18의 방법.
20.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법으로서, 상기 방법은 상기 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 RNA 성분의 상류에 리보자임을 암호화하는 DNA 서열을 포함하는 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 제2 재조합 DNA 구조물로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하고, RNA 성분과 Cas9 엔도뉴클레아제는 표적 부위 서열의 전부 또는 일부에 결합할 수 있는 RGEN을 형성할 수 있는, 방법.
21.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 20의 방법.
22.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 표적 부위 서열로 RNA-유도 엔도뉴클레아제 (RGEN)를 표적화하는 방법으로서, 상기 방법은 상기 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 리보자임-RNA 성분 융합 분자를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 리보자임-RNA 성분 융합 분자와 Cas 엔도뉴클레아제는 표적 부위 서열의 전부 또는 일부에 결합하고, 임의로 절단할 수 있는 RGEN을 형성할 수 있는, 방법.
23.
RGEN이 표적 부위 서열의 전부 또는 일부에 결합하여 절단할 수 있는, 실시 형태 22의 방법.
24.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 표적 부위를 변형시키는 방법으로서, 상기 방법은 통상적이지 않은 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 RNA 성분의 상류에 리보자임을 암호화하는 DNA 서열을 포함하는 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 제2 재조합 DNA 구조물로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 5' 캡을 포함하지 않는 상기 RNA 성분을 생성하고, Cas9 엔도뉴클레아제는 상기 표적 부위에서 단일 또는 이중 가닥 절단을 도입하는, 방법.
25.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 표적 부위를 변형시키는 방법으로서, 상기 방법은 통상적이지 않은 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 5' 캡을 포함하지 않는 리보자임-RNA 성분 융합 분자를 암호화하는 DNA 서열을 포함하는 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 리보자임-RNA 성분 융합 분자와 Cas9 엔도뉴클레아제는 상기 표적 부위에 단일 또는 이중 가닥 절단을 도입하는 RGEN을 형성할 수 있는, 방법.
26.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 다수의 표적 부위를 변형시키는 방법으로서, 상기 방법은 통상적이지 않은 효모에 Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제1 재조합 DNA 구조물 및 적어도 하나의 폴리뉴클레오티드에 작동가능하게 연결된 프로모터를 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 적어도 하나의 폴리뉴클레오티드는 RNA 성분의 상류에 리보자임을 포함하는 RNA 분자를 암호화하고, 상기 RNA 분자는 자체 촉매 작용으로 리보자임을 제거하여 상기 RNA 성분을 생성하며, Cas9 엔도뉴클레아제는 상기 표적 부위에서 단일 또는 이중 가닥 절단을 도입하는, 방법.
27.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 다수의 표적 부위를 변형시키는 방법으로서, 상기 방법은 통상적이지 않은 효모에 Cas9 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제1 재조합 DNA 구조물 및 적어도 하나의 폴리뉴클레오티드에 작동가능하게 연결된 프로모터를 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 적어도 하나의 폴리뉴클레오티드는 리보자임-RNA 성분 융합 분자를 암호화하고, 상기 리보자임-RNA 성분 융합 분자와 Cas9 엔도뉴클레아제는 상기 표적 부위에 단일 또는 이중 가닥 절단을 도입하는 RGEN을 형성할 수 있는, 방법.
28.
상기 표적에서 변형을 갖는 적어도 하나의 통상적이지 않은 효모 세포를 동정하는 단계를 추가로 포함하며, 여기서 변형은 상기 표적 부위에서 하나 이상의 뉴클레오티드의 적어도 하나의 결실, 첨가 또는 치환을 포함하는, 실시 형태 22 내지 실시 형태 25 중 어느 하나의 방법.
29.
상기 효모에 공여 DNA를 제공하는 단계를 추가로 포함하며, 여기서 상기 공여 DNA는 관심 대상의 폴리뉴클레오티드를 포함하는, 실시 형태 24 내지 실시 형태 28 중 어느 하나의 방법.
30.
상기 표적 부위에 통합된 관심 대상의 폴리뉴클레오티드를 이의 염색체 또는 에피솜에 포함하는 적어도 하나의 효모 세포를 동정하는 단계를 추가로 포함하는, 실시 형태 29의 방법.
31.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 뉴클레오티드 서열을 편집하는 방법으로서, 상기 방법은 통상적이지 않은 효모에 폴리뉴클레오티드 변형 주형 DNA, Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 RNA 성분의 상류에 리보자임을 암호화하는 DNA 서열을 포함하는 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 제2 재조합 DNA 구조물로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 5' 캡을 포함하지 않는 상기 RNA 성분을 생성하고, Cas9 엔도뉴클레아제는 상기 효모의 염색체 또는 에피솜의 표적 부위에 단일 또는 이중 가닥 절단을 도입하며, 상기 폴리뉴클레오티드 변형 주형 DNA는 상기 뉴클레오티드 서열의 적어도 하나의 뉴클레오티드 변형을 포함하는, 방법.
32.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 뉴클레오티드 서열을 편집하는 방법으로서, 상기 방법은 통상적이지 않은 효모에 폴리뉴클레오티드 변형 주형 DNA, Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 제1 재조합 DNA 구조물 및 5' 캡을 포함하지 않는 리보자임-RNA 성분 융합 분자를 암호화하는 DNA 서열을 포함하는 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 리보자임-RNA 성분 융합 분자와 Cas9 엔도뉴클레아제는 상기 효모의 염색체 또는 에피솜의 표적 부위에 단일 또는 이중 가닥 절단을 도입하는 RGEN을 형성할 수 있고, 상기 폴리뉴클레오티드 변형 주형 DNA는 상기 뉴클레오티드 서열의 적어도 하나의 뉴클레오티드 변형을 포함하는, 방법.
33.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 뉴클레오티드 서열을 편집하는 방법으로서, 상기 방법은 통상적이지 않은 효모에 Cas9 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제1 재조합 DNA 구조물 및 적어도 하나의 폴리뉴클레오티드에 작동가능하게 연결된 프로모터를 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 적어도 하나의 폴리뉴클레오티드는 RNA 성분의 상류에 리보자임을 포함하는 RNA 분자를 암호화하고, 상기 RNA 분자는 자체 촉매 작용으로 리보자임을 제거하여 5' 캡을 포함하지 않는 상기 RNA 성분을 생성하며, Cas9 엔도뉴클레아제는 상기 효모의 염색체 또는 에피솜의 표적 부위에 단일 또는 이중 가닥 절단을 도입하고, 상기 폴리뉴클레오티드 변형 주형 DNA는 상기 뉴클레오티드 서열의 적어도 하나의 뉴클레오티드 변형을 포함하는, 방법.
34.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 뉴클레오티드 서열을 편집하는 방법으로서, 상기 방법은 통상적이지 않은 효모에 적어도 하나의 폴리뉴클레오티드 변형 주형 DNA, Cas 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제1 재조합 DNA 구조물 및 적어도 하나의 폴리뉴클레오티드에 작동가능하게 연결된 프로모터를 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 적어도 하나의 폴리뉴클레오티드는 5' 캡을 포함하지 않는 리보자임-RNA 성분 융합 분자를 암호화하고, 상기 리보자임-RNA 성분 융합 분자와 Cas9 엔도뉴클레아제는 상기 효모의 염색체 또는 에피솜의 표적 부위에 단일 또는 이중 가닥 절단을 도입하는 RGEN을 형성할 수 있으며, 상기 폴리뉴클레오티드 변형 주형 DNA는 상기 뉴클레오티드 서열의 적어도 하나의 뉴클레오티드 변형을 포함하는, 방법.
35.
제1 재조합 DNA와 제2 재조합 DNA가 동일한 플라스미드 상에 위치하는, 실시 형태 24 내지 실시 형태 34 중 어느 하나의 방법.
36.
제1 재조합 DNA와 제2 재조합 DNA가 별도의 플라스미드 상에 위치하는, 실시 형태 24 내지 실시 형태 34 중 어느 하나의 방법.
37.
통상적이지 않은 효모 내의 염색체 또는 에피솜 상의 뉴클레오티드 서열을 침묵시키는 방법으로서, 상기 방법은 통상적이지 않은 효모에 불활성화된 Cas9 엔도뉴클레아제를 암호화하는 DNA 서열을 포함하는 적어도 하나의 제1 재조합 DNA 구조물 및 적어도 하나의 폴리뉴클레오티드에 작동가능하게 연결된 프로모터를 포함하는 적어도 하나의 제2 재조합 DNA 구조물을 제공하는 단계를 포함하며, 여기서 상기 적어도 하나의 폴리뉴클레오티드는 5' 캡을 포함하지 않는 리보자임-RNA 성분 융합 분자를 암호화하고, 상기 리보자임-RNA 성분 융합 분자와 불활성화된 Cas9 엔도뉴클레아제는 상기 효모의 염색체 또는 에피솜 내의 상기 뉴클레오티드 서열에 결합하는 RGEN을 형성함으로써 상기 뉴클레오티드 서열의 전사를 차단할 수 있는, 방법.
38.
통상적이지 않은 효모에서 유전자 변형을 위한 다수의 가이드 RNA를 고효율로 생성하는 방법으로서, 상기 방법은 하기 단계를 포함하는, 방법:
a)
5' 에서 3' 순서로 리보자임을 암호화하는 제1 DNA 서열, 역선별제를 암호화하는 제2 DNA 서열, 가이드 RNA의 CER 도메인을 암호화하는 제3 DNA 서열 및 종결 서열에 작동가능하게 연결된 프로모터를 포함하는 재조합 DNA 구조물을 제공하는 단계;
b) (a)의 재조합 DNA 구조물에 적어도 하나의 올리고뉴클레오티드 듀플렉스를 제공하는 단계로서, 상기 올리고뉴클레오티드 듀플렉스는 가이드 RNA 표적 서열의 가변 표적화 도메인 (VT)을 암호화할 수 있는 DNA 서열을 포함하는 제1 단일 가닥 올리고뉴클레오티드와 가변 표적화 도메인을 암호화하는 DNA 서열에 상보적인 서열을 포함하는 제2 단일 가닥 올리고뉴클레오티드를 조합하는 것으로부터 유래되는 단계;
c) (a)의 역선별제를 (b)의 적어도 하나의 올리고듀플렉스와 교환함으로써, 각각이 가이드 RNA의 가변 표적화 도메인을 암호화할 수 있는 DNA 서열을 포함하는 재조합 DNA 구조물의 라이브러리를 생성하는 단계; 및
d) (c)의 재조합 DNA 구조물의 라이브러리를 전사함으로써, 리보자임-가이드 RNA 분자의 라이브러리를 생성하는 단계.
39.
리보자임-가이드 RNA 분자가 리보자임과 리보자임 상류에 임의의 RNA 서열을 자체 촉매 작용으로 제거하도록 상기 분자의 라이브러리를 유도하여, 5' 캡을 함유하지 않는 가이드 RNA 분자의 라이브러리를 생성하는 단계를 추가로 포함하는, 실시 형태 38의 방법.
40.
리보자임-가이드 RNA 분자가 리보자임 상류에 임의의 RNA 서열을 절단하도록 상기 분자의 라이브러리를 유도하여, 5' 캡을 함유하지 않는 리보자임-gRNA 융합 분자를 생성하는 단계를 추가로 포함하는, 실시 형태 38의 방법.
41.
(ii) 리보자임과 가이드 RNA를 암호화하는 뉴클레오티드 서열에 작동가능하게 연결된 (i) 폴리머라제-II 프로모터를 포함하는 재조합 DNA 서열로서, 여기서 상기 리보자임은 상기 가이드 RNA의 상류이고, (ii)의 뉴클레오티드 서열로부터 전사된 RNA는 자체 촉매 작용으로 리보자임을 제거하여 상기 가이드 RNA를 생성하며, 상기 가이드 RNA는 통상적이지 않은 효모의 게놈 내의 표적 부위를 인식하고, 결합하며, 임의로 절단할 수 있는 RGEN을 형성할 수 있는, 재조합 DNA 서열.
42.
리보자임과 가이드 RNA를 포함하는 재조합 RNA 서열로서, 여기서 상기 리보자임은 상기 가이드 RNA의 상류이고, 자체 촉매 작용으로 상기 리보자임을 제거하여 상기 가이드 RNA를 생성하며, 상기 가이드 RNA는 통상적이지 않은 효모의 게놈 내의 표적 부위를 인식하고, 결합하며, 임의로 절단할 수 있는 RGEN을 형성할 수 있는, 재조합 RNA 서열.
43.
(ii) 리보자임과 가이드 RNA를 암호화하는 뉴클레오티드 서열에 작동가능하게 연결된 (i) 폴리머라제-II 프로모터를 포함하는 재조합 DNA 서열로서, 여기서 상기 리보자임은 상기 가이드 RNA의 상류이고, (ii)의 뉴클레오티드 서열로부터 전사된 RNA는 리보자임-가이드 RNA 융합 분자를 생성하며, 상기 리보자임-가이드 RNA 융합 분자는 통상적이지 않은 효모의 게놈 내의 표적 부위를 인식하고, 결합하며, 임의로 절단할 수 있는 RGEN을 형성할 수 있는, 재조합 DNA 서열.
44.
리보자임-가이드 RNA 융합 분자를 포함하는 재조합 RNA 서열로서, 여기서 상기 리보자임-가이드 RNA 융합 분자는 통상적이지 않은 효모의 게놈 내의 표적 부위를 인식하고, 결합하며, 임의로 절단할 수 있는 RGEN을 형성할 수 있는, 재조합 RNA 서열.
실시예
개시된 발명은 하기 실시예에서 추가로 정의된다. 이들 실시예는 본 발명의 소정의 바람직한 측면을 나타내지만 단지 예로서만 주어짐을 이해해야 한다. 상기 논의 및 이들 실시예로부터, 당업자는 본 발명의 본질적인 특징을 확인할 수 있으며, 본 발명의 사상 및 범주로부터 벗어나지 않고서, 본 발명을 다양하게 변경 및 변형하여 본 발명이 다양한 용도와 조건에 적합하도록 할 수 있다.
실시예 1
야로위아의
Pol
III 프로모터로부터 발현된
sgRNA는
Cas9를
표적 부위로 유도하지 않으며 DNA 절단을 매개하지 않음
이러한 실시예는 Leu2 유전자좌를 표적으로 하는 야로위아 리폴리티카에서 sgRNA와 Cas9 단백질을 발현하도록 설계된 벡터 및 카세트를 개시한다. 이러한 효모에서 생성된 sgRNA와 Cas9가 상호작용하고, 표적 부위를 찾아 절단할 수 있는 경우, 오류가 발생하기 쉬운 비상동 말단 결합 (NHEJ)을 통해 표적 부위에 돌연변이가 발생될 것이다.
도 1은 가변 표적화 도메인 (VT) (가이드 서열)과 Cas 엔도뉴클레아제 인식 도메인 (CER) 두 영역을 함유하는 단일 RNA 분자인 sgRNA 분자를 도시한다. VT 영역은 표적화된 핵산 분자와 동일성을 갖는 RNA 폴리뉴클레오티드의 20mer일 수 있다. VT 도메인은 PAM 모티프 (예를 들어, NGG, 서열 번호:47)의 5'에 위치하는 표적 부위에서 절단을 위한 표적 부위를 특정한다. CER 도메인은 Cas9 단백질과 상호작용하여, VT 도메인이 상호작용하도록 하여 Cas9 단백질 절단을 유도한다 (문헌[Jinek et al., Science 337:816-821]). VT 도메인과 CER 도메인 모두가 sgRNA의 기능을 위해 필요하다.
야로위아의 LEU2 유전자좌의 암호화 영역에서 3개의 개별 표적 부위 (Leu2-1, Leu2-2, Leu2-3)로 Cas9를 표적화하는 VT 도메인을 암호화하는 DNA 서열을 표 3에 열거한다. 표 3은 또한 야로위아 CAN1 유전자좌의 암호화 영역을 표적으로 하는 VT 도메인을 암호화하는 DNA 서열을 열거한다.
[표 3]
표 3의 LEU2-표적화 DNA 서열 각각을 스트렙토코커스 피오제네스 Cas9 단백질과 상호작용하는 CER 도메인 (서열 번호:1)을 암호화하는 DNA 서열에 개별적으로 융합시켜, CER 도메인과 VT 도메인을 모두 갖는 완전 sgRNA를 암호화하는 DNA 서열을 생성하였다 (서열 번호:1은 5'에서 3' 방향으로 서열 번호:56의 tracrRNA 메이트 서열, 서열 번호:43의 루프 형성 서열 (GAAA) 및 서열 번호:58의 tracrRNA 서열을 포함함에 유의한다). 세포의 핵에서 이러한 sgRNA을 발현시키고, 핵외수송과 5' 변형 시스템을 회피하기 위하여, sgRNA를 암호화하는 DNA 서열을 사카로마이세스 세레비시아 (Snr52 [서열 번호:5] 또는 Rpr1 [서열 번호:6]) 또는 야로위아 리폴리티카 (Snr52 [서열 번호:7])로부터의 RNA Pol III 프로모터의 제어 하에 두었다. 구체적으로, Sc Snr52를 Leu2-1에 융합시키고, Sc Rpr1을 Leu2-2에 융합시키며, Yl Snr52를 Leu2-3에 융합시켰다. 각각의 sgRNA를 암호화하는 DNA 서열의 3' 말단을 사카로마이세스 세레비시아 (서열 번호:8)의 Sup4 유전자로부터의 강력한 종결자에 융합시켰다. 따라서, 3개의 상이한 Pol III-유도된 sgRNA 카세트를 제조하였다.
표준 기술에 따라 스트렙토코커스 피오제네스 M1 GAS (SF370)로부터의 Cas9 유전자의 오픈 리딩 프레임을 야로위아에서의 발현을 위해 코돈-최적화하여, 서열 번호:9를 생성하였다. 유인원 바이러스 40 (SV40) 1부분 핵 국재화 신호 (NLS)와 짧은 링커 (4개의 아미노산)를 암호화하는 DNA 서열을 서열 번호:9의 마지막 센스 코돈 다음에 혼입하여 서열 번호:10을 만들었다. 서열 번호:10은 서열 번호:11에 나타낸 아미노산 서열을 암호화한다. 서열 번호:11의 마지막 7개의 아미노산은 첨가된 NLS를 암호화하면 반면, 서열 번호:11의 1369 내지 1372 위치의 잔기는 첨가된 링커를 암호화한다. 야로위아 코돈-최적화된 Cas9-NLS 서열 (서열 번호:10)을 표준 분자생물학 기술에 의해 야로위아 항시성 프로모터인 FBA1 (서열 번호:12)로 융합시켰다. 항시성 FBA1 프로모터, 야로위아 코돈-최적화된 Cas9 및 SV40 NLS를 함유하는 야로위아 코돈-최적화된 Cas9 발현 카세트 (서열 번호:13)의 예가 도 2a에 예시되어 있다. 이러한 Cas9 발현 카세트 (서열 번호:13)를 플라스미드 pZUF로 클로닝하여, 구조물 pZUFCas9 (도 3a, 서열 번호:14)를 만들었다.
(상기) 각각의 sgRNA 발현 카세트를 pZUFCas9 (서열 번호:14)의 PacI/ClaI 부위로 개별적으로 클로닝하여, 효모 세포를 야로위아 코돈-최적화된 Cas9 발현 카세트 및 Pol III-유도된 sgRNA 발현 카세트로 공동형질전환(co-transform)하는 데 사용될 수 있는 pZUFCas9/sgRNA 구조물을 만들었다. 이러한 구조물의 예는 pZUFCas9/PolIII-sgRNA (도 3b)이며, 이는 야로위아에서 Leu2-3을 표적화하기 위한 Yl Snr52 - sgRNA 발현 카세트를 함유한다.
우라실 영양요구성 야로위아 리폴리티카 세포를 200 ng의 플라스미드 pZUFCas9 (서열 번호:14) 또는 특정 pZUFCas9/sgRNA (예를 들어, pZUFCas9/PolIII-sgRNA, 도 3b)로 형질전환시키고, 우라실이 없는 완전 최소 플레이트 (CM-ura) 상에서 우라실 원영양성(prototroph)을 선별하였다. CM-ura 플레이트 상에 발생하는 콜로니를 류신이 없는 완전 최소 플레이트 (CM-leu) 상에서 류신 영양요구성에 대하여 스크리닝하였다. 우라실 원영양성 형질전환체 중 어느 것도 류신 영양요구성을 나타내지 않았다. 이러한 결과는 야로위아 코돈-최적화된 Cas9 및 Pol III 프로모터-유도된 sgRNA가 발현되지 않았고/거나, 생성되지 않았으며/거나, 상호작용하지 않았고/거나, DNA를 표적하지 않았으며/거나 DNA를 절단하지 않았음을 제시한다. 이러한 실험이 류신 영양요구성을 생성하는 경우, 이러한 결과는 Cas9/sgRNA 복합체가 Leu2 암호화 영역을 표적화하고 절단하여 오류가 발생하기 쉬운 NHEJ를 유도하고, 결과적으로 삽입-결실을 형성하여, 프레임시프트 돌연변이를 일으킨다는 것을 나타낼 가능성이 있다.
따라서, sgRNA의 Pol III-유도된 발현은 야로위아에서 기능적 Cas9-sgRNA 복합체를 제공하는 데 유용하지 않을 수 있다.
실시예 2
DNA 폴리머라제 II 프로모터에 의해 유도된 5'- 및 3'-리보자임을 포함하는 야로위아-최적화된 sgRNA 발현 카세트
이러한 실시예는 야로위아에서 발현 및 Cas9-매개 표적화를 위해 최적화된 sgRNA를 개시한다. 이러한 발현을 위해 사용된 각각의 카세트는 5'-리보자임 및 3'-리보자임 (리보자임-sgRNA-리보자임 또는 RGR)에 융합된 sgRNA의 전사를 유도하기 위한 Pol II 프로모터를 포함하였다. 5' 및 3' 리보자임을 제공하여 sgRNA로부터 Pol II 프로모터-관련 전사물 변형, 예컨대 5' 캡 구조를 제거하여, sgRNA 서열만을 남겼다. 이러한 발현 카세트는 sgRNA 발현을 위한 더 넓은 프로모터 선택을 허용한다. 또한, 이러한 카세트로부터 전사된 sgRNA는 5'-캡 구조가 없기 때문에 핵외수송의 대상이 아니다. 이러한 특징은 야로위아 세포에서 sgRNA의 강한 발현을 허용하여, Cas9 엔도뉴클레아제를 생체 내 게놈의 표적화 영역으로 유도할 수 있다.
sgRNA 서열에 5' 망치머리 (HH) 및 3' 델타 간염 바이러스 (HDV) 리보자임을 첨가함으로써 일부 RNA 폴리머라제 (예를 들어 Pol II)에 의해 전사된 프로모터에서 발생하는 전사후 변형을 고려하지 않고 임의의 프로모터로부터 sgRNA의 발현을 허용하고, sgRNA 발현을 위한 현재의 프로모터의 제한된 선택을 피한다. 이러한 sgRNA가 발현되는 경우, 예비-sgRNA 전사물에 존재하는 리보자임은 자가절단(autocleave)됨으로써, 전사물로부터 분리되어, 변형되지 않은 sgRNA를 남긴다.
시험된 각각의 sgRNA의 경우, sgRNA를 암호화하는 DNA 서열을 (i) 이의 5'-말단에서 5' HH 리보자임을 암호화하는 서열 (서열 번호:15)로 (ii) 이의 3'-말단에서 3' HDV 리보자임을 암호화하는 서열 (서열 번호:16)로 융합시켰다. HH 리보자임의 5'-결합은 HH 리보자임의 처음 6개의 뉴클레오티드가 sgRNA의 VT 영역 (가이드 서열)의 처음 6개의 뉴클레오티드의 역 상보서열이 되도록 하였다. 각각의 리보자임-플랭킹된 예비-sgRNA (RGR)를 표준 분자생물학 기술을 사용하여 FBA1 프로모터 (서열 번호:12)에 융합시켜, 야로위아-최적화된 sgRNA 발현 카세트 (도 2b에 도시된 최종 카세트)를 얻었다. 이러한 카세트의 예시적인 서열이 서열 번호:18에 도시되어 있으며, 이는 RGR을 암호화하는 서열 (HH-sgRNA-HDV)에 작동가능하게 연결된 FBA1 프로모터 (서열 번호:12)를 포함하고, 여기서 sgRNA는 이의 CER 도메인으로서 서열 번호:17 (Can1-1) 및 서열 번호:1에 의해 암호화되는 VT 도메인을 포함한다 (서열 번호:18, pRF38 (서열 번호:19) 및 pRF84 (서열 번호:41)의 CER 도메인-암호화 영역 각각이 서열 번호:1의 위치 73-74 (CER 도메인)에 상응하는 잔기 위치 사이에 있는, 첨가된 'TGG'를 가짐에 유의한다). 이러한 VT 도메인은 야로위아 CAN1 유전자 오픈 리딩 프레임의 암호화 영역에 있는 부위를 표적으로 한다 (진뱅크 수탁 번호 NC_006068, YALI0B19338g, 염색체 B의 ~bp 2557513-2559231). 암호화된 HH 리보자임의 처음 6개의 잔기는 sgRNA의 처음 6개의 잔기 (즉, VT 도메인의 처음 6개의 잔기)에 상보적이다. 서열 번호:18의 서열 번호:12 (FBA1 프로모터) 바로 다음에 3개의 잔기 (ATG)가 존재하며, 이는 예비-sgRNA의 발현과 리보자임-매개 자체촉매 작용에 영향을 주지 않는 것으로 여겨짐에 유의한다. 서열 번호:18을 pRF38 (도 3c, 서열 번호:19)로 명명된 구조물로 클로닝하였다.
따라서, 5' 및 3' pol II 프로모터-관련 전사물 변형없이 sgRNA를 발현시키기 위한 DNA 카세트를 제조하였다. 이러한 유형의 카세트를 야로위아에서의 Cas9 유전자 표적화를 위해 실시예 3에서 사용하였다.
실시예 3
야로위아-최적화된 sgRNA가 sgRNA/Cas9 엔도뉴클레아제 시스템에서 사용되어 염색체 DNA를 절단할 수 있음
이러한 실시예는 실시예 2에 기재된 바와 같은 야로위아-최적화된 sgRNA 발현 카세트를 사용하여 야로위아에서 염색체 DNA를 인식하여 절단하기 위해 Cas9와 함께 작용할 수 있는 sgRNA를 발현하는 것을 개시한다. 이러한 절단은 절단 부위에서 오류가 발생하기 쉬운 NHEJ DNA 수복으로 인해 예측되는 DNA 절단 부위의 영역에서 돌연변이가 발생한 것에 의해 나타났다.
야로위아 리폴리티카의 CAN1 유전자는 절단의 표적이었다. 야로위아 형질전환체에서의 CAN1의 성공적인 표적화를 각각 표현형 (카나바닌 저항) 및 돌연변이 빈도 및 스펙트럼에 대한 서열 분석으로 조사하였다.
Ura- 야로위아 리폴리티카 세포 (균주 ATCC 20362로부터 직접 유래된 우라실 영양요구성인 균주 Y2224가 본 명세서에 참고로 포함되는 미국 특허 출원 공개 제2010/0062502호에 개시되어 있음)를 리튬 이온-매개 형질전환 (문헌[Ito et al., J. Bacteriology 153:163-168])에 의해 pZUFCas9 (도 3a, 서열 번호:14) 및 CAN1 유전자좌를 표적화하기 위한 야로위아-최적화된 RGR 예비-sgRNA 카세트 (서열 번호:18에 포함됨)를 함유하는 pRF38 (도 3c, 서열 번호:19)로부터 증폭된 선형 PCR 생성물로 공동형질전환시켰다. 이러한 PCR 증폭에 사용된 프라이머는 서열 번호:20 (정방향) 및 서열 번호:21 (역방향)이었다. pZUFCas9 (서열 번호:14)로만 형질전환된 Ura- 야로위아 리폴리티카 세포 (Y2224)는 음성 대조군으로 역할을 하였다. pZUFCas9 (서열 번호:14) 및 RGR 예비-sgRNA 발현 카세트로 형질전환된 세포를 CM-ura 배지 상에서 우라실 원영양성으로서 선별하였다. CM-ura 플레이트를 우라실과 아르기닌이 없고, 60 ㎍/ml의 독성 아르기닌 유사체인 카나바닌 (Cm+can)이 보충된 완전 최소 배지 상으로 복제 평판(replica-plating)하여 CAN1 유전자에 기능 상실 돌연변이를 함유하는 세포를 스크리닝하였다. 기능적 CAN1 유전자를 가진 세포는 카나바닌을 세포로 수송하여 세포 사멸을 일으킬 수 있다. CAN1 유전자에 기능 상실 대립유전자를 가진 세포는 카나바닌을 수송하지 못하며 CM+can 플레이트 상에서 성장할 수 있다.
카나바닌 내성의 표현형 스크리닝에 의한 회복된 기능 상실 돌연변이체의 빈도는 Cas9 단독으로 형질전환된 세포에서는 0이었다 (도 4). 그러나, Cas9가 RGR 예비-sgRNA 발현 카세트로 공동형질전환된 경우, 카나바닌-내성 형질전환체의 빈도가 10%로 증가하였다 (도 4).
카나바닌-내성 콜로니의 CAN1 유전자좌를 정방향 (서열 번호:22) 및 역방향 (서열 번호:23) PCR 프라이머를 사용하여 증폭시켰다. PCR 생성물을 자이모클린(Zymoclean)TM 및 농축기 컬럼 (캘리포니아주 얼바인 소재의 자이모 리서치(Zymo Research))을 사용하여 정제하였다. PCR 생성물을 서열 분석 프라이머 서열 번호:24를 사용하여 서열 분석하였다 (생어법). 서열을 표적 부위를 함유하는 야생형 (WT) 야로위아 CAN1 암호화 서열과 정렬시켰다 (도 5). Cas9와 sgRNA를 모두 발현하는 세포의 CAN1 유전자좌에서 주된 기능 상실 돌연변이 (서열분석된 분리주의 73%)는 Cas9 절단 부위에서의 -1 프레임시프트 돌연변이었다 (도 5). 보다 적은 수의 다른 결실 및 삽입이 CAN1 유전자좌에서의 돌연변이의 나머지를 구성하였다. 총, 90%의 돌연변이가 작은 결실 또는 삽입이었다 (도 5). 드물게, 다른 염색체로부터의 소량의 서열의 삽입 (4%), 절단 부위에서 야로위아-최적화된 sgRNA 발현 카세트의 삽입 (1.5%) 또는 더 큰 결실 (1%)과 같은 다른 이벤트가 발생하였다. 스크리닝한 3.5%의 카나바닌-내성 콜로니가 CAN1 유전자좌에서 서열 분석으로 결정되지 않은 복잡한 재배열을 가졌다. 전체적으로, CAN1 표적 부위에서 관찰된 돌연변이는 Cas9/sgRNA 복합체에 의해 생성된 절단을 수복하기 위해 오류가 발생하기 쉬운 NHEJ가 세포에서 사용되었음을 나타낸다.
(i) CAN1-특이적 Cas9 엔도뉴클레아제가 발현되도록 형질전환된 세포에서 카나바닌-내성 콜로니의 증가된 빈도 및 (ii) 카나바닌-내성 돌연변이는 예측되는 Cas9 절단 부위에서의 오류가 발생하기 쉬운 NHEJ 이벤트에 의한 것임을 나타내는 서열 분석 데이터 모두에 의해 실시예 2에 기재된 야로위아-최적화된 Cas9 및 RGR 예비-sgRNA 발현 카세트가 야로위아 염색체 DNA를 절단하여 돌연변이를 생성함을 확인하였다.
따라서, RNA 성분의 5' 캡이 리보자임의 자체 촉매 작용으로 제거되는, 5'-캡이 없는 RGEN (예를 들어, Cas9)의 RNA 성분 (예를 들어, sgRNA)을 발현하는 것은 통상적이지 않은 효모에서 DNA 서열의 RGEN-매개 표적화를 허용한다.
실시예 4
(3' 리보자임없이) 5'-리보자임을 사용하여 발현된 야로위아-최적화된 sgRNA는 염색체 DNA를 절단하기 위한 sgRNA/Cas9 엔도뉴클레아제 시스템에 유용함
이러한 예에서, 3' 리보자임없이 5' HH 리보자임만을 함유하는 야로위아-최적화된 카세트로부터 생성된 sgRNA의 기능을 평가하여, sgRNA가 Cas9와 상호작용하고, DNA 표적 서열을 인식하며, Cas9에 의한 DNA 절단을 유도하고, 오류가 발생하기 쉬운 NHEJ에 의한 돌연변이를 유도할 수 있는지를 결정하였다.
Pol II 프로모터로부터 전사된 RNA는 이들의 5' 및 3' 말단 모두에서 심하게 가공되고 변형되어, Pol II 프로모터로부터 기능적 sgRNA를 생성하기 위해 5' 및 3' 말단을 모두 절단해야 한다는 것을 시사한다. 플랭킹 영역을 갖는 시험관 내에서 생성된 sgRNA가 (i) 5'-플랭킹 영역이 존재하는 경우 비기능적이고 (ii) 3' 플랭킹 영역이 존재하는 경우 기능적으로 상당히 손상된다는 것이 이전부터 밝혀져 왔다 (문헌[Gao et al., J. Integr. Plant BioL. 56:343-349]). 5' 리보자임과 3' 플랭킹 영역도 함유하는 예비-sgRNA가 Cas9와 함께 사카로마이세스 세레비시아로 발현되는 경우, sgRNA는 절단을 위한 표적 부위로 Cas9를 유도하도록 기능하지 않았다 (문헌[Gao et al., ibid]).
(3'-위치한 리보자임이 없는) 5' 리보자임-플랭킹된 sgRNA가 통상적이지 않은 효모에서 기능할 수 있는지 시험하기 위해, 5'에서 3' 방향으로, S. 세레비시아 Sup4 유전자 (서열 번호:8)로부터의 강력한 전사 종결자에 융합된 Can1-1 표적 부위 (서열 번호:17)를 표적화하는 sgRNA (서열 번호:70의 예)를 암호화하는 서열에 융합된 HH 리보자임 (서열 번호:15)에 융합된 FBA1 프로모터 (서열 번호:12)를 함유하는 야로위아-최적화된 sgRNA 발현 카세트 (서열 번호:25)를 구축하였다 (이러한 카세트는 RG [리보자임-sgRNA] RNA를 발현하는 것으로 특징지어질 수 있다). RG 발현 카세트에서 암호화된 sgRNA는 CER 도메인 (서열 번호:1)에 연결된 서열 번호:17에 상응하는 VT 도메인을 포함한다. 암호화된 HH 리보자임의 처음 6개의 잔기는 sgRNA의 처음 6개의 잔기 (즉, VT 도메인의 처음 6개의 잔기)에 상보적이다. 서열 번호:25의 서열 번호:12 (FBA1 프로모터) 바로 다음에 3개의 잔기 (ATG)가 존재하며, 이는 예비-sgRNA의 발현과 리보자임-매개 자체촉매 작용에 영향을 주지 않는 것으로 여겨짐에 유의한다. 이러한 야로위아-최적화된 RG 발현 카세트 (서열 번호:25)가 도 2c에 예시되어 있다.
Cas9와 상호작용하고, Cas9를 Cas9에 의한 절단을 위한 DNA 표적 서열로 유도할 수 있는 sgRNA를 발현하는 야로위아-최적화된 RG 카세트의 능력을 시험하기 위해, RG 구조물 (서열 번호:25) 또는 RGR 구조물 (서열 번호:18, 실시예 2)을 함유하는 PCR 생성물을 pZUFCas9 (서열 번호:14)과 함께 Ura- 야로위아 리폴리티카 세포 (Y2224)로 리튬 이온-매개 형질전환에 의해 공동형질전환시켰다 (문헌[Ito et al., ibid]). Ura+ 형질전환체를 CM+can 플레이트 상으로 복제 평판하여 카나바닌-내성 세포 (실시예 3에서와 같이)에 대해 스크리닝하였는데, 여기서 RG 또는 RGR 예비-sgRNA로부터 생성된 sgRNA는 Cas9를 유도하는 데 작용하여 CAN1 표적 서열을 절단하여 NHEJ를 통한 오류가 발생하기 쉬운 수복을 초래하였다. 야로위아-최적화된 RG 또는 RGR 카세트가 Cas9 매개 절단을 표적 부위로 유도하는 빈도는 동일하며 (도 6), 이는 S. 세레비시아를 사용한 문헌[J. Integr. Plant BioL. 56:343-349]의 결과와 대조적으로 야로위아에서 효율적인 Cas9/sgRNA 표적 절단 및 돌연변이를 위해 3' 리보자임이 필요하지 않음을 나타내었다.
이러한 예는 통상적이지 않은 효모, 예컨대 야로위아에서 리보자임 전략을 사용하는 경우, Pol II 프로모터로부터 기능적 sgRNA를 생성하기 위해 5'-플랭킹 리보자임만이 필요할 수 있음을 입증한다. 이러한 결과는 Cas9에 의한 표적 서열의 효율적인 절단 및 돌연변이를 위해 5' 및 3' 리보자임이 모두 필요한 통상적인 효모인 S. 세레비시아에서 관찰된 것과 대조적이다 (문헌[Gao et al., ibid]).
따라서, 이러한 예는 RNA 성분의 5' 캡이 리보자임의 자체 촉매 작용으로 제거되는, 5'-캡이 없는 RGEN (예를 들어, Cas9)의 RNA 성분 (예를 들어, sgRNA)을 발현하는 것은 통상적이지 않은 효모에서 DNA 서열의 RGEN-매개 표적화를 허용함을 추가로 입증한다.
실시예
5
Cas9
/
sgRNA
-유도된 DNA 이중 가닥 절단의 상동 재조합 (HR) 수복을 촉진하기 위한 선형 폴리뉴클레오티드 변형 주형의 사용
이러한 실시예는 야로위아에서 야로위아-최적화된 Cas9 및 예비-sgRNA 발현 카세트를 발현시킴으로써 생성된 이중 가닥 절단 (DSB)을 수복하기 위해 선형 폴리뉴클레오티드 변형 주형 DNA 서열을 사용하는 HR 시스템(machinery)의 능력을 시험하는 방법을 개시한다. 각각 염색체 DNA에서 Cas9/sgRNA 표적화 부위 외부의 영역에 상동성인 5'- 및 3'-암 서열을 갖는 3개의 상이한 선형 주형 서열을 제조하였다.
처음 2개의 유형의 폴리뉴클레오티드 변형 주형 서열을 상보적인 합성 올리고뉴클레오티드로부터 제조하였다. 상보적인 올리고뉴클레오티드를 어닐링한 후 에탄올 침전에 의해 정제하였다.
상보성 올리고뉴클레오티드 (서열 번호:28 및 29)를 사용하여 제1 폴리뉴클레오티드 변형 주형을 제조하였고, 20-뉴클레오티드 Can1-1 표적 부위 (서열 번호:17), 3-뉴클레오티드 PAM 도메인 및 Can1-1 표적 부위의 바로 상류에 있는 2개의 뉴클레오티드를 결실시킴으로써 CAN1 유전자에서 -1bp 프레임시프트를 일으키는 8개의 코돈 및 1개의 염기쌍을 결실시키도록 설계하였다. 서열 번호:28과 이의 역 상보서열인 서열 번호:29를 어닐링함으로써 제1 폴리뉴클레오티드 변형 주형을 조립하였다. 제1 공여 DNA의 상동성 암 (각각 약 50-bp)은 서로 바로 옆에 위치하며; 이들 사이에는 이종 서열이 없다.
상보성 올리고뉴클레오티드 (서열 번호:30 및 31)을 사용하여 제2 폴리뉴클레오티드 변형 주형을 제조하였고, CAN1 오픈 리딩 프레임에 2개의 인-프레임(in-frame) 번역 종결 코돈 (즉, 넌센스 돌연변이)을 생성하도록 설계하였다. 또한, Can1-1 표적 부위 하류의 PAM 서열 (CGG를 ATG로 대체함) 및 시드(seed) 서열의 제1 뉴클레오티드 (즉, 서열 번호:17의 Can1-1 표적 서열의 마지막 잔기) (C를 G로 대체함)를 파괴하도록 설계되었다. 서열 번호:30과 이의 역 상보서열인 서열 번호:31을 어닐링함으로써 이러한 폴리뉴클레오티드 변형 주형을 생성하였다. 상기로부터 얻을 수 있듯이, 제2 공여 DNA의 상동성 암 (각각 약 50-bp)은 이종 서열의 단지 수개의 염기쌍에 의해 이격된다.
2개의 PCR 생성물을 생성함으로써 부분적으로 제3 폴리뉴클레오티드 변형 주형을 제조하였다. PCR 생성물 (서열 번호:32, 서열 번호:33 [정방향] 및 서열 번호:34 [역방향]의 프라이머를 사용하여 야로위아 리폴리티카 ATCC 20362 게놈 DNA로부터 증폭됨) 중 하나에서, 서열 번호:32의 위치 638은 CAN1 오픈 리딩 프레임 시작 코돈 상류의 뉴클레오티드 3 bp에 상응한다. 역방향 프라이머 (서열 번호:34)는 CAN1 오픈 리딩 프레임 하류의 37 bp에 위치하는 서열에 상보적인 17개의 뉴클레오티드를 첨가한다. 제2 PCR 생성물 (서열 번호:35, 서열 번호:36 [정방향] 및 서열 번호:37 [역방향]의 프라이머를 사용하여 야로위아 리폴리티카 ATCC 20362 게놈 DNA로부터 증폭됨)은 CAN1 오픈 리딩 프레임의 종결 코돈 하류의 14번째 염기쌍에서 시작하는 637개의 염기쌍(637 base pairs starting 14 base pairs)을 포함한다. 정방향 프라이머 (서열 번호:36)는 CAN1 오픈 리딩 프레임 상류의 종결되는 2개의 염기쌍(ending 2 base pairs) 영역에 상보적인 20개의 뉴클레오티드를 첨가한다. 상류 (서열 번호:32) PCR 생성물 및 하류 PCR 생성물 (서열 번호:35) 모두를 자이모클린TM 및 농축기 컬럼을 사용하여 정제하였다. 이러한 PCR 생성물 (각각 10 ng)을 새로운 PCR 반응에서 혼합하였다. 생성물 상류의 3'-최외각(most) 37개의 뉴클레오티드는 생성물 하류의 5'-최외각 37개의 뉴클레오티드와 동일하다. 상류 및 하류 단편을 사용하여 서로를 프라이밍하여, 상류 및 하류 서열 모두를 함유하는 중첩 말단으로부터의 합성에 의해 단일 생성물 (서열 번호:38)을 생성하였다 (문헌[by Horton et al., Biotechniques 54:129-133]에 기재된 기술). 서열 번호:38 공여 DNA의 상동성 암 (각각 600-bp 이상)은 서로 바로 옆에 위치하며; 이들 사이에는 이종 서열이 없다. 이러한 폴리뉴클레오티드 변형 주형은 Can1-1 표적 부위의 Cas9/sgRNA-매개 이중 가닥 절단의 영역에서 전체 CAN1 오픈 리딩 프레임을 포함하는 큰 결실을 가능하게 할 수 있다.
Ura- 야로위아 리폴리티카 세포 (Y2224)를 상기 리튬 이온 형질전환 방법을 사용하여 (i) pZUFCas9 (서열 번호:14), (ii) 1 ㎍의 야로위아-최적화된 RGR 예비-sgRNA 발현 카세트 (서열 번호:18) 및 (iii) 1 nmol의 "프레임시프트 주형" DNA (서열 번호:28), 1 nmol의 "점 돌연변이 주형" DNA (서열 번호:30) 또는 1 ㎍의 "큰 결실 주형" DNA (서열 번호:38)로 형질전환하였다. 형질전환된 세포를 CM-ura 플레이트 상에서 우라실에 대한 원영양성으로서 회수하였다. 원영양성 콜로니를 CM+can에 복제 평판으로 스크리닝하여 CAN1 돌연변이를 가진 카나바닌-내성 세포를 동정하였다. 각각의 형질전환으로부터의 CanR 콜로니의 CAN1 유전자좌를 정방향 (서열 번호:22) 및 역방향 프라이머 (서열 번호:23)를 사용하는 PCR 증폭을 통해 스크리닝하였다. 각각의 PCR 생성물을 ExoSAP-IT? (캘리포니아주 산타 클라라 소재의 아피메트릭스(Affymetrix))를 사용하여 정제하고, 서열 분석 프라이머 서열 번호:24를 사용하여 서열분석 (생어법)하였다. CanR 콜로니의 총 개수에서 (특정 주형 DNA가 형질전환에 사용되었다는 관점에서) 예측되는 상동 재조합 이벤트를 나타내는 콜로니의 빈도는 약 15%이었다 (도 7).
3개의 상이한 폴리뉴클레오티드 변형 주형 DNA 서열의 HR 수복의 효율성이 약간 상이하다 (도 8). 특히, 각각의 이러한 주형에 대한 HR 빈도는 약 11% (큰 결실 및 프레임시프트 공여자) 내지 22% (점 돌연변이 주형) (도 8)이었으며, 이는 Can1-1 표적 부위에서의 Cas9/sgRNA-생성된 절단 이벤트의 일부가 폴리뉴클레오티드 변형 주형 DNA가 제공되는 경우 충실도가 높은(high-fidelity) 방식으로 HR 경로를 사용하여 수복되었음을 나타낸다.
DNA 수복의 2가지 주요 경로인 NHEJ 또는 HR의 사용은 야로위아에서 NHEJ에 대한 분명한 편향을 나타내며 (도 7), 이는 통상적인 효모의 Cas9/sgRNA-매개 절단 이벤트에서의 수복의 연구에서 관찰된 것과는 상이하다. 예를 들어, 문헌[DiCarlo et aL. (Nucleic Acids Res. 41:4336-4343)]은 Cas9/sgRNA-매개 DNA 절단의 수복을 위해 공여 DNA가 제공되는 경우 얻어지는 거의 모든 S. 세레비시아 돌연변이체가 HR을 통해 생성되었지만, 공여 DNA가 제공되지 않는 경우 빈도는 4 내지 5배만큼(4 to 5 orders of magnitude) 떨어졌고, 이는 HR에 대한 분명한 편향을 나타내었다. 대조적으로, Cas9/sgRNA (RGR 카세트로부터 발현된 sgRNA) 절단 부위에서 야로위아의 총 돌연변이 빈도는 폴리뉴클레오티드 변형 주형 DNA를 수용하거나 수용하지 않는 형질전환체 (도 9, 두 유형의 형질전환체에 대한 ~15% 돌연변이율을 나타냄) 사이에서 다르지 않았고, HR은 공여 DNA가 제공되는 경우 생성되는 돌연변이 형질전환체의 약 15%만을 차지한다 (도 7). 따라서, 상기 관찰된 바와 같은 야로위아의 폴리뉴클레오티드 변형 주형 DNA 서열을 갖는 HR의 빈도는 단지 약 2.25%이었고, 이는 통상적인 효모에서 공여 DNA를 사용하여 관찰된 100%에 가까운 HR-매개 돌연변이율과 현저한 대조를 이룬다 (문헌[DiCarlo et al., ibid]).
따라서, 이러한 예는 RNA 성분의 5' 캡이 리보자임의 자체 촉매 작용으로 제거되는, 5'-캡이 없는 RGEN (예를 들어, Cas9)의 RNA 성분 (예를 들어, sgRNA)을 발현하는 것은 통상적이지 않은 효모에서 DNA 서열의 RGEN-매개 표적화를 허용함을 추가로 입증한다. 이러한 예는 또한 통상적이지 않은 효모에서 RGEN-매개 절단이 적합한 공여 DNA (폴리뉴클레오티드 변형 주형)가 제공되는 경우 소정 비율로 HR에 의해 수복될 수 있음을 입증한다.
실시예
6
단일 안정한 벡터로부터의 Cas9 및 야로위아-최적화된 RGR 또는 RG 예비-sgRNA의 발현은 Cas9/sgRNA-매개 표적 DNA 절단을 제공함
이러한 예에서, 야로위아-최적화된 RGR 또는 RG 예비-sgRNA 발현 카세트를 각각 야로위아-최적화된 Cas9 발현 카세트로서 동일한 안정한 발현 플라스미드로 개별적으로 옮겼다. 특히, 서열 번호:18 (RGR 발현에 대한) 또는 서열 번호:25 (RG 발현에 대한)를 각각 개별적으로 pZUFCas9 (도 3a, 서열 번호:14)로 클로닝하였다. 이는 단일-성분 형질전환이 세포에서 Cas9 엔도뉴클레아제, 및 RG 또는 RGR 예비-sgRNA를 발현하도록 허용함으로써, Cas9/sgRNA-매개 표적 부위 절단, 이어서 오류가 발생하기 쉬운 NHEJ 수복을 제공하였다.
야로위아-최적화된 RGR (서열 번호:18) 또는 RG (서열 번호:25) sgRNA 발현 카세트를 정방향 (서열 번호:39) 및 역방향 (서열 번호:40) 프라이머를 사용하여 PCR에 의해 증폭시켰다. 각각의 산물을 PacI/ClaI 제한 부위에서 플라스미드 pZUFCas9 (서열 번호:14)로 개별적으로 클로닝하여, 각각 Cas9 발현 및 최적화된 RGR 예비-sgRNA (pRF84, 서열 번호:41, 도 10a) 또는 최적화된 RG 예비-sgRNA (pRF85, 서열 번호:42, 도 10b)의 발현을 위한 각각의 카세트를 갖는 2개의 새로운 플라스미드를 생성하였다.
Cas9/sgRNA-매개 표적 부위 (Can1-1) 절단을 제공하기 위해 각각 Cas9 및 sgRNA를 효율적으로 발현하는 pRF84 (서열 번호:41) 및 pRF85 (서열 번호:42) 플라스미드 구조물의 능력을 시험하기 위해, Ura- 야로위아 리폴리티카 세포 (Y2224)를 상기 리튬 이온 형질전환 방법을 사용하여 200 ng의 pRF84 (서열 번호:41), pRF85 (서열 번호:42) 또는 pZUFCas9 (서열 번호:14)로 형질전환하였다. 각각의 플라스미드로 형질전환된 세포를 CM-ura 배지 상에서 우라실 원영양성으로 선별하였다. 각각의 형질전환으로부터의 우라실 원영양성을 CM+can 상의 복제 평판에 의해 CAN1 돌연변이체에 대해 스크리닝하였다. CM+can 플레이트 상에서 성장한 콜로니의 수를 사용하여 pZUFCas9 (Cas9 단독 발현), pRF84 (Cas9 및 RGR 예비-sgRNA 발현) 또는 pRF85 (Cas9 및 RG 예비-sgRNA 발현)로 형질전환된 세포의 CAN1 돌연변이 빈도 (도 11)를 생성하였다. CAN1 유전자좌에서 pZUFCas9 (서열 번호:14)로 형질전환된 야로위아 세포의 Cas9/sgRNA-매개 돌연변이의 빈도가 0인 반면, (i) Cas9 및 (ii) RGR 예비-sgRNA (pRF84) 또는 RG sgRNA (pRF85)를 발현하는 세포의 CAN1 돌연변이 빈도 (~69%)는 카나바닌-내성 (도 11)에 의해 나타낸 바와 유사하다.
이러한 결과는 동일한 벡터로부터 Cas9 및 예비-sgRNA를 발현하는 것이 상당히 더 높은 비율의 Cas9/sgRNA-매개 절단을 유도하고 결과적으로 예측되는 절단 부위에서 NHEJ-매개 돌연변이를 유도함을 나타낸다. Cas9 및 예비-sgRNA (RGR 또는 RG 예비-sgRNA)를 암호화하는 별개의 서열로 형질전환된 야로위아 세포는 약 5%의 표적화된 돌연변이 빈도 (실시예 4, 도 6)를 나타내는 반면, Cas9 및 sgRNA 암호화 서열을 형질전환에 사용되는 동일한 벡터 상에 위치시키면 표적화된 돌연변이 빈도는 약 69%이었다 (도 11).
따라서, 통상적이지 않은 효모를 형질전환하는데 사용되는 동일한 구조물로부터 Cas 단백질 및 이의 상응하는 RNA 성분을 발현하면 RGEN 단백질 및 RNA 성분을 발현하기 위해 별개의 구조물을 사용하는 것과 비교하여 효모에서 Cas-매개 DNA 표적화의 비율이 더 높아진다.
실시예 7
야로위아 리폴리티카에서 HDV 리보자임-sgRNA 융합을 사용하는 고-효율 유전자 표적화
이러한 실시예는 단일 가이드 RNA (HDV 리보자임에 의해 5' 말단 상에 플랭킹된 sgRNA) (리보자임-단일 가이드 RNA 융합)의 사용을 논의한다. 발현되는 경우, HDV 리보자임은 그 자체의 서열의 5'을 절단하여, 임의의 선행되는 전사물을 제거하지만, sgRNA의 5' 말단에 융합된 HDV 서열을 남겨둔다.
플라스미드 pZuf-Cas9 (서열 번호: 14)를 애질런트(Agilent) 퀵체인지(QuickChange) 및 다음과 같은 프라이머 AarI-제거-1 (
, 서열 번호: 90) 및 AarI-제거-2 (
, 서열 번호:91)를 사용하여 돌연변이화하여, pZuf-Cas9 (서열 번호: 14) 상의 Cas9 유전자 (서열 번호: 10)에 존재하는 내인성 AarI 부위를 제거하고, pRF109 (서열 번호: 92)를 생성하였다. 변형된 Aar1- Cas9 유전자 (서열 번호: 93)를 pRF109로부터의 NcoI/NotI 단편으로서 pZufCas9의 NcoI/NotI 부위에 클로닝하여, 기존 Cas9 유전자 (서열 번호: 10)를 Aar1- Cas9 유전자로 대체하여 pRF141 (서열 번호: 94)을 생성하였다.
고효율 클로닝 카세트 (도 12a, 서열 번호: 95)는 yl52 프로모터 (서열 번호: 96), HDV 리보자임 (서열 번호: 16), 대장균 역선별 카세트 rpsL (서열 번호: 97), 가이드 RNA CER 도메인을 암호화하는 DNA (서열 번호: 1) 및 S. 세레비시아 Sup4 종결자 (서열 번호: 8)로 이루어진다. 고효율 클로닝 카세트 (서열 번호: 95)의 말단을 플랭킹하는 것은 PacI 및 ClaI 제한 효소 인식 부위이다. 고효율 클로닝 카세트를 pRF141 (서열 번호: 94)의 PacI/ClaI 부위로 클로닝하여 pRF291 (서열 번호: 98)을 생성하였다. rpsL 역선별 카세트 (서열 번호: 97)는 S12 리보솜 단백질 서브유닛(subunit)을 암호화하는 대장균 유전자 rpsL의 WT 카피를 함유한다 (문헌[Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology, 1987, First eD. American Society of Microbiology, Washington, DC]). S12 서브유닛의 일부 돌연변이는 열성 방식으로(recessive manner) (문헌[Lederberg J. 1951. Streptomycin Resistance; a genetically recessive mutatioN. Journal of bacteriology 61:549-550]) 항생제 스트렙토마이신에 대한 내성을 일으키므로 (문헌[Ozaki M, Mizushima S, Nomura M. 1969. Identification and functional characterization of the protein controlled by the streptomycin-Resistant locus in E. coli. Nature 222:333-339]), rpsL 유전자의 야생형 카피가 존재하는 경우 균주는 표현형적으로 스트렙토마이신에 민감하다. Top10 (라이프 테크놀로지스(Life technologies))과 같은 일반적인 클로닝 균주는 세포가 스트렙토마이신에 내성을 갖도록 그들의 염색체 상에 rpsL 카피를 가지고 있다.
가이드 RNA의 가변 표적화 도메인을 암호화하는 DNA 단편을 플라스미드 (예컨대 pRF291)로 클로닝하는 것은 어닐링되는 경우 가변 표적화 도메인을 암호화하는 DNA 단편 및 고효율 클로닝 카세트에 존재하는 2개의 AarI 부위로의 클로닝을 위한 정확한 돌출부를 함유하는 2개의 부분적으로 상보적인 올리고뉴클레오티드를 필요로 한다. 2개의 올리고뉴클레오티드 Can1-1F (
, 서열 번호: 99) 및 Can1-1R (
, 서열 번호: 100)를 100μM에서 듀플렉스 완충액 (30mM HEPES pH 7.5, 100mM 아세트산나트륨)에서 재현탁시켰다. Can1-1F (서열 번호 99) 및 Can1-1R (서열 번호 100)을 단일 튜브 내에서 각각 50μM의 최종 농도로 혼합하고, 5분 동안 95℃로 가열하여, 0.1℃/min으로 25℃로 냉각시켜, 2개의 올리고뉴클레오티드를 어닐링하여, Can1-1 표적 부위를 표적화할 수 있는 가이드 RNA의 가변 표적 도메인을 암호화하는 DNA 단편을 함유하는 작은 듀플렉스 DNA 분자 (도 12b)를 형성하였다 (PAM 서열 CGG를 포함하는 서열 번호: 101로 나타냄). 최종 부피 20μl 중에 50ng의 pRF291, Can1-1F 및 Can1-1R로 구성된 2.5μM의 소형 듀플렉스 DNA, 1x T4 리가제 완충액 (50mM Tris-HCl, 10mM MgCl
2, 1mM ATP, 10mM DTT pH 7.5), 0.5 μM의 AarI 올리고뉴클레오티드, 2 단위 AarI, 40 단위 T4 DNA 리가제를 함유하는 단일 튜브 분해(digestion)/라이게이션 반응을 생성하였다. 이중가닥 Can1-1F 및 Can1-1R 듀플렉스가 없는 제2 대조 반응물을 또한 조립하였다. 반응물을 30분 동안 37℃에서 인큐베이션하였다. 10μl의 각각의 반응물을 이전에 기재된 바와 같은 Top10 대장균 세포로 형질전환하였다 (문헌[Green MR, Sambrook J. 2012. Molecular Cloning: A Laboratory Manual, Fourth Edition eD. Cold Spring Harbor Laboratory PRess, Cold Spring Harbor, NY]). Can1-1F 및 Can1-1R의 듀플렉스가 AarI 제한 부위 (도 12a)에 의해 플랭킹된
rpsL 역선별 마커를 대체한 pRF291의 존재를 선별하기 위해, 세포를 100 ㎍/ml의 앰피실린 및 50 ㎍/ml의 스트렙토마이신을 함유하는 1.5% (w/v) 박토(Bacto) 한천으로 고형화된 용원성 브로쓰 상에 플레이팅하였다. 고효율 클로닝 카세트를 함유하는 pRF291의 존재는 플라스미드 상의 역선별 카세트의 존재로 인해 항생제 암피실린에 대해 표현형적으로 내성이지만 항생제 스트렙토마이신에 민감성인 콜로니를 생성하였다. 그러나, 역선별 카세트가 AarI 효소를 통해 제거되고 Can1-1 이중 DNA가 부위로 라이게이션되는 경우 (AarI에 대한 인식 서열을 제거함), 플라스미드로 형질전환된 세포는 암피실린 내성, 스트렙토마이신 내성 표현형을 가졌다 (도 12a). Can1-1 가변 표적화 도메인 표적화 (역선별 카세트를 대체함)를 암호화하는 DNA 단편을 함유하는 pRF291은 sup4 종결자 (서열 번호: 8)에 융합된 가이드 CER 도메인 (서열 번호: 1)을 암호화하는 DNA에 융합된 Can1-1 가변 표적화 도메인 (서열 번호: 17)을 암호화하는 DNA에 융합된 HDV 리보자임 (서열 번호: 16)을 암호화하는 DNA에 융합된 yl52 프로모터를 함유하는 재조합 HDV- sgRNA 발현 카세트 (서열 번호: 102)를 생성하였다. 이러한 구조물 pRF303 (서열 번호: 103)을 함유하는 플라스미드를 사용하여 (Cas9 엔도뉴클레아제와 복합체를 형성한 경우) HDV 리보자임-가이드 RNA (서열 번호: 104)를 암호화하여, 돌연변이 유발을 위해 야로위아 리폴리티카의 Can1 유전자 (서열 번호: 21)를 표적화하였다.
(문헌[Richard M, Quijano RR, Bezzate S, Bordon-Pallier F, Gaillardin C. 2001. Journal of bacteriology 183:3098-3107]에 기재된 바와 같이) 플라스미드가 없거나 sgRNA 발현 카세트 (pRF291, 서열 번호: 98)가 없는 100 ng의 플라스미드, RGR 발현 카세트 (서열 번호: 41)를 갖는 pRF84 플라스미드, 5' 리보자임이 sgRNA (서열 번호: 42)로부터 그 자체를 제거하는 RG 카세트를 갖는 pRF85 플라스미드 또는 야로위아의 Can1-1 표적 부위를 표적화하는 HDV-sgRNA 융합 발현 카세트 (서열 번호: 102)를 갖는 pRF303(서열 번호 103)로 야로위아 리폴리티카를 형질전환하였다. 형질전환체를 우라실 원영양성에 대해 선별하고, Can1 유전자에서 아르기닌 유사체 카나바닌에 대한 표현형 내성에 의해 돌연변이에 대해 기록하였다. HDV-sgRNA 융합을 발현하는 플라스미드는 리보자임으로부터 유리된 sgRNA 중 하나를 발현하는 플라스미드와 동일한 빈도로 Can1 유전자에서 기능 상실 돌연변이를 유발하였으며, 이는 HDV 리보자임과 sgRNA 표적화 Can1-1의 5' 융합이 sgRNA 기능에 영향을 주지 않음을 시사하였다 (표 4).
[표 4]
다수의 추가의 표적 부위 (표 5)를 표적화하는 가변 표적 도메인을 암호화하는 다수의 추가의 DNA 단편을 상기 기재한 바와 동일한 전략을 사용하여 도 12a에 예시된 바와 같이 pRF291 (서열 번호: 98) 플라스미드로 클로닝하였다. Can1 유전자 (서열 번호: 105) 내에서, can1-2 표적 부위 (서열 번호: 106) 및 다른 표적 부위, 예컨대 sou2-1 (서열 번호: 107), Sou2-2 (서열 번호: 108), Tgl1-1 (서열 번호: 112), Acos10-1 (서열 번호: 113), Fat1-1 (서열 번호: 114) 및 Ura3-1 (서열 번호: 116)을 표적화하는 제2 표적 부위를 표적화하는 가변 표적화 도메인을 암호화하는 DNA 단편을 포함함.
[표 5]
표적 부위의 돌연변이 빈도는 모든 HDV-sgRNA 융합체가 Cas9 엔도뉴클레아제와 복합체를 형성할 수 있음을 나타내었으며, 이는 NHEJ를 통한 돌연변이를 유도하는 각각의 표적 부위에서 절단을 일으켰다 (표 6).
[표 6]
실시예 8
불활성화된-Cas9 및 HDV-sgRNA 융합을 사용한 유전자 침묵.
HNH 및 RuvC 뉴클레아제 도메인 (서열 번호: 117)에 돌연변이를 함유하는 촉매적으로 불활성화된 Cas9 변이체는 sgRNA와 상호작용하고 생체 내 표적 부위에 결합할 수 있지만, 표적 DNA의 어느 한 가닥을 절단할 수는 없다. DNA에 결합하지만 절단하지 않는 이러한 작용 방식은 영구적인 유전적 변화를 일으키지 않으면서, 염색체의 특정 유전자좌의 발현을 일시적으로 감소시키는 데 사용될 수 있다.
야로위아 리폴리티카를 위한 촉매적으로 불활성화된 Cas9 발현 카세트를 생성하기 위해, 프라이머 D10AF (
, 서열 번호: 118) 및 D10AR (
, 서열 번호: 119)로 기재된 바와 같은 퀵체인지 부위-지정 돌연변이 유발 (스트라타진(Stratagene))을 사용하여 D10A 돌연변이를 플라스미드 pZufCas9 (서열 번호: 14)에 도입하였다. 이는 RuvC 뉴클레아제 (서열 번호 120) 및 구조물 pRF111 (서열 번호 121)을 함유하는 상응하는 플라스미드를 불활성화시키는 D10A 돌연변이를 갖는 야로위아 코돈 최적화된 Cas9 유전자를 생성시켰다. 제2 뉴클레아제 도메인 (HNH)을 불활성화시키기 위해, H840A 돌연변이를 야로위아 코돈 최적화된 D10A 유전자로 도입하여 야로위아 코돈 최적화된 Cas9 불활성화된 유전자 (서열 번호: 124) 및 야로위아에서의 발현을 위한 유전자를 포함하는 플라스미드인 pRF143 (서열 번호: 125)를 생성하는 프라이머 H840A1 (
, 서열 번호: 122) 및 H840A2 (
, 서열 번호: 123)를 사용하여 추가의 1회의 퀵체인지 돌연변이 유발 (스트라타진)을 수행하였다.
야로위아 리폴리티카에서 유전자 침묵을 평가하기 위해, 야로위아 코돈 최적화된 dsRED익스프레스 오픈 리딩 프레임 (서열 번호: 126)을 5' NcoI 제한 부위 및 3' NotI 제한 부위 (서열 번호: 127)를 갖는 클로닝 단편으로서 제조하였다. 클로닝 단편 (서열 번호: 127)을 pZufCas9의 NcoI/NotI 부위로 클로닝하여 플라스미드 pRF165 (서열 번호: 129) 상에 함유된 FAB1-dsRED 융합 카세트 (서열 번호: 128)를 생성하는 야로위아 최적화된 dsRED익스프레스 클로닝 단편 (서열 번호: 127)에 융합된 FBA1 프로모터 (서열 번호: 12)를 생성하였다. FBA1-dsRED익스프레스 카세트 (서열 번호: 128)를 염색체로 통합하기 위해, 카세트 (서열 번호: 130)를 함유하는 PmeI-NotI 단편을 통합 플라스미드 p2P069 (서열 번호: 131)의 PmeI/NotI 부위로 라이게이션하여 FBA1-dsRED익스프레스 발현 카세트인 pRF201 (서열 번호: 132)을 포함하는 통합 벡터를 생성하였다. FBA1-dsRED익스프레스 융합체 및 Leu2 유전자 (서열 번호: 133)의 카피를 포함하는 pRF201의 SphI/AscI 단편을 표준 기술을 사용하여 류신 원영양성에 대해 선별함으로써 야로위아의 염색체로 통합하였다 (문헌[Richard M, Quijano RR, Bezzate S, Bordon-Pallier F, Gaillardin C. 2001. Tagging morphogenetic genes by insertional mutagenesis in the yeast Yarrowia lipolyticA. Journal of bacteriology 183:3098-3107]). FBA1-dsRED익스프레스 발현 카세트의 존재를 표준 PCR 기술 및 프라이머 HY026 (
, 서열 번호: 134) 및 HY027 (
, 서열 번호: 135)을 사용하여 야로위아 게놈에서 확인하였다. FBA1-dsRED익스프레스 카세트 (서열 번호: 128)를 함유하는 2개의 독립 균주 YRF41 및 YRF42를 추가의 사용을 위해 선택하였다.
야로위아 최적화된 dsRED익스프레스 발현 카세트 (서열 번호: 128)를 표적화하기 위한 sgRNA를 생성하기 위해 실시예 12와 유사한 전략을 사용하였다. 도 13a에 도시된 바와 같이, 플라스미드 구조물인 pRF169 (서열 번호: 136)는 야로위아로부터의 GPD 프로모터 (서열 번호: 137), 역선별가능한 마커, 가이드 RNA CER 도메인 (서열 번호: 1)을 암호화하는 DNA 및 Sup4 종결자 (서열 번호: 8) 카세트 (서열 번호: 138)를 함유하였다. HH 리보자임을 암호화하는 DNA 단편에 연결된 야로위아의 표적 부위를 표적화하는 sgRNA의 가변 표적화 도메인을 암호화하는 DNA를, 도 13b에 도시된 바와 같이 망치머리 리보자임의 처음 6개의 뉴클레오티드가 가변 표적 도메인의 처음 6개의 뉴클레오티드의 역 상보서열이되도록 하는 HH 리보자임을 암호화하는 DNA 단편을 제외하고는 실시예 12에 기재된 바와 같이 pRF169 (서열 번호 136)로 클로닝하였다. 정확한 돌출부를 갖는 이중 가닥 올리고뉴클레오티드가 AarI 부위 사이의 역선별 카세트를 대체하는 경우, 리보자임-가이드 RNA (RG) 발현 카세트가 생성되었다 (도 13-a). 전사되는 경우, HH 리보자임은 리보자임-가이드 RNA 분자로부터 5' 전사물 및 그 자체를 제거하여, 세포에 온전한 sgRNA를 남긴다. dsRED익스프레스 오픈 리딩 프레임 (서열 번호: 126)을 표적으로 하는 3개의 가이드 RNA를 생성하였는데; 2개는 주형 가닥, ds-주형-1 (서열 번호: 139), ds-주형-2 (서열 번호: 140)를 표적화하고 하나는 비주형 가닥 ds-비주형-1 (서열 번호: 141)을 표적화하였다.
각각의 표적 부위에 대해, 표적 특이적인 망치머리 리보자임, 가변 표적 도메인 (VTD) 및 pRF169의 AarI 부위로 클로닝하기 위한 정확한 중첩 말단을 암호화하는 DNA 서열을 함유하는 2개의 올리고뉴클레오티드를 설계하였다. 각 부위의 올리고뉴클레오티드; ds-주형-1F (서열 번호 144) ds-주형-1R (서열 번호 145), ds-주형-2F (서열 번호 146), ds-주형-2R (서열 번호 147) ds-비주형-1F (서열 번호 148) 및 ds-비주형-1R (서열 번호 149)을 이중 가닥화하여(duplexed) pRF169의 고효율 카세트 (도 13a 및 도 13b)에 남아있는 AarI 돌출부로 클로닝하기 위한 정확한 돌출부를 갖는 이중 가닥 DNA 분자를 형성하였고, pRF291로 클로닝하기 위해 실시예 12에 기재된 바와 같이 수행하였다. 역선별 카세트를 대체하는 sgRNA의 가변 표적 도메인을 암호화하는 DNA 단편의 삽입은 Sup4 종결자에 융합된 가이드 RNA CER 도메인을 암호화하는 DNA에 융합된 망치머리 리보자임-표적 부위 듀플렉스 DNA에 융합된 GPD 프로모터를 갖는 각각의 표적 부위에 대한 새로운 플라스미드를 생성하였다 (도 13a). 이러한 듀플렉스를 함유하는 플라스미드는 pRF296 (ds-주형-1, 서열 번호: 150), pRF298 (ds-주형-2, 서열 번호: 151), pRF300 (ds-비주형-1, 서열 번호: 152)이다.
유전자 침묵을 위한 구조물을 생성하기 위해, pRF143으로부터의 불활성화된 Cas9 (서열 번호: 125)를 표준 기술을 사용하여 NcoI/NotI 단편으로서 pRF293, pRF298 및 pRF300에 클로닝하고, 이들 플라스미드의 NcoI/NotI 부위에 존재하는 기능적 Cas9 (서열 번호: 93)를 대체하여 각각 플라스미드 pRF339 (서열 번호: 153), pRF341 (서열 번호: 154) 및 pRF342 (서열 번호: 155)를 생성하였다.
균주 YRF41 및 YRF42는 표준 기술을 사용하여 pRF339, pRF341 및 pRF343으로 우라실 원영양성으로 형질전환되었다 (문헌[Richard M, Quijano RR, Bezzate S, Bordon-Pallier F, Gaillardin C. 2001. Tagging morphogenetic genes by insertional mutagenesis in the yeast Yarrowia lipolyticA. Journal of bacteriology 183:3098-3107]). 각각의 형질전환에 있어서, 12개의 형질전환체를 우라실이 없는 플레이트 상에 도말 정제하여, 플라스미드를 유지하였다. 각각의 분리주를 사용하여 2ml의 CM-ura 브로쓰 (테크노바(Teknova))를 접종하고, 30℃, 250 RPM에서 하룻밤 동안 성장시켰다. 각각 2 내지 5μl를 하룻밤 동안 200μl의 ddH20로 희석하고, 아큐리(Accuri) 유세포 분석기의 dsRED익스프레스 채널에서 분석하여, 각 세포 내의 dsRED익스프레스 단백질의 양을 평가하였다. 7,151 내지 10,000개의 세포를 각 배양물로부터 분석하였다. dsRED익스프레스 발현 카세트가 없는 야로위아 세포의 평균 형광도를 분석한 각 배양물의 평균 형광도에서 제하여, 이들을 평균낸 각 균주/플라스미드 조합 내에서 보정된 평균 형광도를 얻었고, 표준 편차를 측정하였다 (표 7). 목적 유전자를 표적화하는 발현 벡터를 통해 발현된 리보자임-sgRBA (RG)와 조합된 불활성화된 Cas9는 유전자의 발현을 2 내지 10배로 침묵시켰다. 배수 침묵화(fold silencing)는 표적 부위의 위치 및 쇄의 수(strandedness) 및/또는 야로위아 세포에서 기능적 형태의 DNA 폴리머라제 프로모터로부터 발현되는 리보자임 플랭킹된 sgRNA의 능력에 따라 달라졌다 (표 7).
[표 7]
실시예 9
단일 플라스미드로부터 발현된 Cas9 및 HDV 리보자임-sgRNA 융합체 (RG)를 사용한 정확한 유전자 편집.
이러한 예에서 우리는 동일한 안정한 벡터로부터 발현된 Cas9와 HDV-sgRNA 융합체의 안정한 발현이 상동성 재조합을 통한 정확한 유전자 편집을 위한 기질이될 수 있는 야로위아의 표적 부위에 DNA 이중 가닥 절단을 생성할 수 있음을 보여준다.
실시예 4에 기재된 Can1 결실 폴리뉴클레오티드 변형 주형 DNA (서열 번호: 38)를 HinDIII로 분해하고 표준 기법을 사용하여 pUC18의 HinDIII 부위로 클로닝하여 pRF80 (서열 번호: 156)을 생성하였다. 더 짧은 Can1 결실 편집 주형 (서열 번호: 157)을 표준 PCR 기술 및 프라이머 80F (
, 서열 번호: 158) 및 80R (
, 서열 번호: 159)을 사용하여 pRF80으로부터 증폭시켜, 더 많은 양의 편집 주형을 생성하였다.
Ura 영양요구성 야로위아 세포를 표준 기술을 사용하여 (문헌[Richard M, Quijano RR, Bezzate S, Bordon-Pallier F, Gaillardin C. 2001. Tagging morphogenetic genes by insertional mutagenesis in the yeast Yarrowia lipolyticA. Journal of bacteriology 183:3098-3107]) Cas9 유전자의 카피를 포함하지만 sgRNA가 없는 100ng의 플라스미드 pRF291, 및 Cas9 유전자의 카피 및 편집 주형 DNA가 없거나 1000 ng의 짧은 Can1 결실 편집 주형 (서열 번호: 157)을 갖는 Can1-1 표적 부위 HDV-sgRNA 발현 카세트를 함께 포함하는 pRF303으로 형질전환하였다. 형질전환체를 CM-ura 배지 (테크노바) 상에서 선별하였다. 각각의 형질전환에 대해, 20개의 개별 콜로니를 CM-ura 배지 (테크노바) 상에서 도말 정제하였다. 도말 정제된 각각의 콜로니로부터 4개의 개별 콜로니 (형질전환 당 총 80개)를 60 ㎍/ml의 L- 카나바닌을 함유하는 CM-arg 플레이트 상으로 패치하여, Can-1 유전자에서 기능적 대립유전자의 결실을 포함하는 콜로니에 대해 스크리닝하였다. 카나바닌에 대한 내성을 보이는 패치를 기록하고 유전자 불활성화의 빈도를 기록하였다 (표 8). 상동성 재조합으로 인해 Can1 기능을 상실한 콜로니 및 NHEJ로 인해 Can1 기능을 상실한 콜로니를 결정하기 위해, Can1 유전자좌 (서열 번호: 160)를 Can1-PCRF (
, 서열 번호: 22) 및 Can1-PCRR (
, 서열 번호: 23)을 사용하여 증폭시켰다. 이전 실시예에 기재된 바와 같이 작은 삽입-결실을 갖는 세포에서, PCR 생성물은 Can1 결실 편집 주형과 상동성 재조합에 의한 결실을 함유하는 균주에서 WT Can1 좌위 (서열 번호: 160)와 크기 (2125bp)가 매우 유사해야 하며, Can1-PCRF (서열 번호: 22) 및 Can1-PCRR (서열 번호: 23)을 갖는 PCR 단편 (서열 번호: 161)은 더 작을 것이다 (392bp). 2 μl의 PCR 생성물을 전기영동으로 분해하고, 표준 기술을 사용하여 영상을 얻었다 (그림 14). 편집 주형 (서열 번호: 161)과의 재조합에 해당하는 짧은 밴드를 갖는 도말 정제 시에, 1개 이상의 콜로니를 생성하는 원래의 20개의 도말된 콜로니의 백분율을 사용하여 HR의 빈도를 측정하였다 (표 8). pRF303 (서열 번호: 103)을 수용한 세포에서, 카나바닌 내성 콜로니의 빈도는 세포가 편집 주형을 받았는지 여부와 유사하였다 (표 8). 형질전환된 세포의 총 개체군 중 pRF303 (서열 번호: 103) 및 Can1 짧은 편집 형질 (서열 번호: 157) 모두를 수용한 세포에서, 약 1/10이 편집 주형(서열 번호: 157)으로부터의 Can1 유전자좌의 정확한 편집을 포함하였다 (표 8).
[표 8]
실시예 10
야로위아에서 URA3 유전자 불활성화
본 실시예는 야로위아에서 URA3 유전자 불활성화를 위해 단일 가이드 RNA (sgRNA)와 Cas9 엔도뉴클레아제를 개별적으로 또는 함께 발현하는 플라스미드의 구성 및 사용을 기술한다.
pYRH235 및 pYRH236은 각각 URA3.1 표적 서열 (5'-ctgttcagagacagtttcct-3; 서열 번호:165)을 표적화하는 리보자임 플랭킹된 예비-sgRNA (RGR-URA3.1; 서열 번호: 164) 및 URA3.2 표적 서열 (5'-taacatccagagaagcacac-3'; 서열 번호:167)을 표적화하는 리보자임 플랭킹된 예비-sgRNA (RGR-URA3.2; 서열 번호: 166)를 발현하였다. RGR-URA3.1의 DNA 단편을 암호화하는 NcoI-NotI 제한 분해 단편 및 RGR-URA3.2를 암호화하는 BspHI-NotI 제한 분해 단편을 FBA1L 프로모터 (서열 번호: 168)에 융합시켜 각각 pYRH235 및 pYRH236을 생성하였다. pYRH235 및 pYRH236 플라스미드는 설포닐 우레아 내성을 부여하는 단일 아미노산 변화 (W497L)를 갖는 천연 아세토하이드록시산 합성효소 (AHAS 또는 아세토락테이트 합성효소; E.C.4.1.3.18; 서열 번호: 169)의 마커 유전자를 함유하였다.
야로위아 균주 ATCC20362의 Ura- 유도체 (Y2224)를 먼저 SphI-BsiWI 제한 효소 분해에 의해 선형화된 pZufCas9 (서열 번호: 14)로 형질전환시키고, 형질전환체를 우라실이 없는 완전 최소 (CM) 플레이트에 상에서 선별하였다. 선형화된 Cas9 발현 카세트는 야로위아 게놈에 무작위로 통합되었으므로, 형질전환체는 URA3 유전자의 카피를 적어도 2개를 함유하였다. 이어서, sgRNA를 발현하는 pYRH235 또는 pYRH236을 Cas9를 발현하는 야로위아 균주로 형질전환시키고, 형질전환체를 600 mg/L의 설포닐 우레아를 함유하는 CM 플레이트 상에서 선별하였다. 50개의 형질전환체를 CM-ura 플레이트 및 5-FOA를 갖는 SC 플레이트에 패치하여, URA3에 대한 Cas9 및 sgRNA에 의한 URA3 유전자 불활성화의 빈도를 찾았다. 각각 94% 및 100%의 pYRH235 및 pYRH236 형질전환체가 우라실 영양요구성이 되었다.
표적 부위 URA3.1 또는 URA3.2에서의 돌연변이의 서열 분석 확인을 수행하였다. ZufCas9 및 pYRH235의 20개의 형질전환체를 서열 분석을 위해 무작위로 선택하고, 각각의 콜로니를 플라스미드 pZufCas9의 URA3 유전자의 돌연변이 및 천연 게놈 URA3에 대해 분석하였다. 플라스미드 pZufCas9로부터의 URA3 유전자를 서열 분석하기 위해, URA3에 대한 RHO705 (서열 번호: 170) 및 FBA1 프로모터 서열에 대한 RHO719 (서열 번호: 171)를 사용하여 영역을 PCR 증폭하였고, 프라이머 RHO733 (서열 번호: 172) 또는 RHO734 (서열 번호: 173)를 사용하여 주형으로서 PCR 증폭 산물을 서열 분석하였다. 천연 게놈 기원의 URA3 유전자를 서열 분석하기 위해, 프라이머 RHO705 (서열 번호: 170) 및 RHO707 (서열 번호: 174)을 사용하여 PCR 증폭을 수행하였고, 프라이머 RHO733 (서열 번호: 172) 및 RHO734 (서열 번호: 173)를 사용하여 주형으로서 PCR 증폭 산물을 서열 분석하였다. 20개의 콜로니 모두는 플라스미드 및 게놈 유래 URA3 유전자 모두에서 돌연변이를 함유하였다 (도 15). 5개의 대표적인 콜로니 (콜로니 1, 2, 3, 5 및 6, 각각 서열 번호: 176, 177, 178, 179 및 180, 및 서열 번호: 181, 182, 183, 184 및 185)의 플라스미드 및 게놈 유래 URA3 유전자, 및 야생형 URA3.1 (서열 번호: 175)에 대한 서열 분석 결과의 단편 정렬을 도 15에 나타낸다. 이러한 결과는 동일한 세포에서 다수의 유전자 카피가 야로위아의 sgRNA/Cas9 엔도뉴클레아제 시스템에 의해 표적화되고 돌연변이되었음을 보여준다.
실시예 11
야로위아에서의 URA3 유전자 돌연변이 또는 결실.
본 실시예는 마커 재활용에 사용하기 위해 야로위아에서의 URA3 유전자 돌연변이 또는 결실을 위한 동일한 벡터 시스템 상에 2개의 sgRNA 및 Cas9 엔도뉴클레아제를 발현하는 플라스미드의 구성 및 사용을 기술한다.
pYRH222는 FBA1 프로모터 (서열 번호: 12), 및 도 16a에 도시된 URA3.2 표적 서열 (서열 번호: 167)을 표적화하는 리보자임 플랭킹된 예비-sgRNA (RGR-URA3.2; 서열 번호: 166)를 암호화하는 FBA1L 프로모터 유도된 DNA 단편 하에 Cas9 엔도뉴클레아제 (서열 번호: 10)를 발현한다. YRH222 벡터는 플라스미드의 염색체외 복제를 수용하는 자가 복제 서열(autonomously replicating sequence) (ARS18, 서열 번호: 208)뿐만 아니라 TDH1 (GPD라고도 지칭됨) 프로모터 (서열 번호: 187) 하에서 발현된 하이그로마이신 항생제 내성 선택 마커 (서열 번호: 186)를 함유하였다 (문헌[PNAS, Fournier, P. et al., 1993, 90:4912-4916]). ARS18의 존재는 선별 압력이 없는 경우 세포가 플라스미드를 상실하게 만들었다.
pYRH282는 pYRH222로부터 유래되었다. pYRH235 유래의 RGR-URA3.1 (서열 번호: 164)을 암호화하는 DNA 단편에 융합된 FBA1L 프로모터 (서열 번호: 168)를 프라이머 RHO804 (서열 번호: 188) 및 RHO805 (서열 번호: 189)를 사용하여 PCR 증폭시켰다. 이어서, PCR 생성물을 BsiWI로 분해하고 pYRH222로 클로닝하였다. 클로닝된 유전자의 배향 및 서열 동일성은 서열 분석에 의해 확인되었고, 구조물은 pYRH282로 명명되었다.
pYRH283은 pYRH222로부터 유래되었다. RGR-URA3.3 (서열 번호: 191)을 암호화하는 DNA에 융합된 TDH1 프로모터 (서열 번호: 187)로 이루어진 BsiWI 부위 (서열 번호: 190)에 의해 플랭킹된 합성 DNA 단편을 IDT (아이오와주 코랄빌 소재)에 의해 합성하고 BsiWI 부위에서 pYRH222로 클로닝하였다. 클로닝된 유전자의 배향 및 서열 동일성은 서열 분석에 의해 확인되었고, 구조물은 pYRH283으로 명명되었다.
야로위아 균주 ATCC20362의 자손을 pYRH222, pYRH282 및 pYRH283으로 형질전환시키고, 형질전환체를 300 mg/L의 하이그로마이신을 함유하는 YPD 플레이트 상에서 선별하였다. 상대적으로 높은 배경 성장이 DNA 대조군이 없는 플레이트 상에서 관찰되었다 (표 9). 각 구조물의 30개의 형질전환체를 무작위로 선별하고, 5-FOA가 함유된 SC 플레이트 상으로 도말하여 우라실 영양요구성에 대해 역선별하였다. DNA 대조군이 없는 플레이트로부터의 콜로니에서는 성장이 관찰되지 않았다. 4 내지 11 개의 패치는 pYRH222, pYRH282 및 pYRH283 형질전환체로 성장을 보였다. 콜로니 PCR을 프라이머 RHO610 (서열 번호: 192) 및 RHO611 (서열 번호: 193)을 사용하여 수행하여, sgRNA 표적화 부위를 함유하는 DNA 영역을 증폭시켰고, PCR 증폭된 생성물은 아가로스 겔 상에서 상이한 이동을 나타내었다 (도 17). PCR 생성물을 주형으로 사용하여 서열 분석을 수행하고, 프라이머 RHO704 (서열 번호: 194)를 사용하여 서열 분석을 수행하였다.
pYRH222 형질전환체의 경우에, 11개의 중 6개의 서열 분석이 성공적으로 수행되었고, 이들 모두는 URA3.2 표적 부위에서 돌연변이되었다 (도 16b; 서열 번호: 195-201). pYRH282의 경우, 성공적으로 서열 분석된 모두가 표적 부위에서 돌연변이를 나타냈으며, 그 중 2개가 2개의 표적 부위 사이에서 결실을 보였다 (도 16c; 서열 번호: 202 내지 204). pYRH283의 경우, 8개 중 7개의 성공적인 서열 분석이 표적 부위에서 돌연변이를 나타냈으며, 그 중 2개가 2개의 표적 부위 사이에서 결실을 보였고 (도 16d; 서열 번호: 205 내지 207), URA3 유전자의 거의 완벽한 결실을 생성하였다.
본 실시예는 야로위아에서 sgRNA/Cas9 엔도뉴클레아제 시스템을 사용하여 2개의 표적 부위 사이에서 표적화된 결실을 만들기 위해 동일한 플라스미드 상에서 2개의 가이드 RNA가 발현되는 것을 보여주는데, 동정을 겔을 사용하거나 서열 분석에 의해 수행하였다. 이러한 플라스미드 상의 ARS18 (서열 번호: 208)의 존재는 선별 압력이 없을 경우 세포가 플라스미드를 상실하게 하여, URA3 마커 재활용을 위해 플라스미드를 반복적으로 사용할 수 있게 하였다.
[표 9]
실시예 12
유전자 불활성화를 위한 야로위아에서의 Csy4 (Cas6)의 사용
본 실시예는 통상적이지 않은 효모에서 DNA 서열 (예를 들어, CAN1이지만 이에 한정되지 않음)을 표적화할 수 있는 RGEN 복합체를 형성할 수 있는 5' 캡이 없는 가이드 RNA를 생성하기 위한 Csy4 (Cas6으로도 지칭됨)의 사용을 기술한다.
Csy4를 암호화하는 유전자 (Cas6으로도 알려짐)를 야로위아에서 CAN1 유전자 불활성화를 위해 28bp의 Csy4 인식 부위에 의해 플랭킹된 sgRNA를 표적화하는 CAN1를 암호화하는 DNA와 함께 Cas9 발현 플라스미드 상으로 도입하였다.
pYRH290은 FBA1 프로모터 (서열 번호: 12) 하에 Cas9 엔도뉴클레아제 (서열 번호: 10)를 FBA1 프로모터 (서열 번호: 210) 하에 Csy4 발현 (서열 번호: 209)을 위한 야로위아 리폴리티카 코돈-최적화된 유전자 하에 발현하였다. pYRH290은 또한 CAN1 표적 서열 (서열 번호: 214)을 표적화하는 28bp의 Csy4 엔도뉴클레아제 인식 서열 (서열 번호: 212) 플랭킹된 예비-sgRNA (서열 번호: 213)를 암호화하는 DNA 단편 (TDH1:28bp-gCAN1-28bp; 서열 번호: 211)을 함유하였다. Csy4에 의한 처리 후, 생성된 sgRNA (서열 번호: 222)는 8-뉴클레오티드 5'-플랭킹 서열 (서열 번호: 223) 및 20-뉴클레오티드 3'-플랭킹 서열 (서열 번호: 224)을 함유하였다.
야로위아 균주 ATCC20362의 Ura- 유도체 (Y2224)를 pYRH290으로 형질전환시키고, 형질전환체를 우라실이 없는 CM 플레이트 상에서 선별하였다. 86개의 형질전환체를 카나바닌을 함유하는 CM 플레이트로 복제 평판하여, can1 돌연변이체를 선별하였다. 86개 중 40개의 형질전환체가 카나바닌을 함유하는 CM 플레이트 상에서 성장시켰다. 40개 중 16개의 카나바닌 내성 콜로니를 서열 분석하여, CAN1 표적 부위 (서열 번호: 214)에서 돌연변이를 확인하였고, 14개의 콜로니를 CAN1 표적 부위에서 돌연변이를 확인하였다. 도 18은 CAN1 표적 부위 (서열 번호: 215) 및 콜로니 14, 16, 18, 19, 24 및 25 (각각 서열 번호: 216 내지 221)의 CAN1 표적 서열에서 돌연변이를 포함하는 야생형 CAN1 유전자의 단편의 정렬을 도시한다.