KR20170053636A - 내열성의 미스매치 엔도뉴클레아제의 이용방법 - Google Patents
내열성의 미스매치 엔도뉴클레아제의 이용방법 Download PDFInfo
- Publication number
- KR20170053636A KR20170053636A KR1020177007069A KR20177007069A KR20170053636A KR 20170053636 A KR20170053636 A KR 20170053636A KR 1020177007069 A KR1020177007069 A KR 1020177007069A KR 20177007069 A KR20177007069 A KR 20177007069A KR 20170053636 A KR20170053636 A KR 20170053636A
- Authority
- KR
- South Korea
- Prior art keywords
- amino acid
- mismatch
- acid sequence
- nucleic acid
- polypeptide
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Abandoned
Links
Images










Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6813—Hybridisation assays
- C12Q1/6827—Hybridisation assays for detection of mutation or polymorphism
- C12Q1/683—Hybridisation assays for detection of mutation or polymorphism involving restriction enzymes, e.g. restriction fragment length polymorphism [RFLP]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases [RNase]; Deoxyribonucleases [DNase]
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6844—Nucleic acid amplification reactions
- C12Q1/6848—Nucleic acid amplification reactions characterised by the means for preventing contamination or increasing the specificity or sensitivity of an amplification reaction
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q2521/00—Reaction characterised by the enzymatic activity
- C12Q2521/30—Phosphoric diester hydrolysing, i.e. nuclease
- C12Q2521/301—Endonuclease
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Genetics & Genomics (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Microbiology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Biochemistry (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Biomedical Technology (AREA)
- Analytical Chemistry (AREA)
- Immunology (AREA)
- Medicinal Chemistry (AREA)
- Chemical Kinetics & Catalysis (AREA)
- Plant Pathology (AREA)
- Enzymes And Modification Thereof (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
도 2는 본 발명의 미스매치 엔도뉴클레아제 활성측정에서 사용한 기질 DNA 일람이다.
도 3은 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단활성을 나타내는 Native-PAGE 결과와 그래프이다.
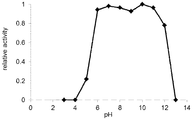
도 4는 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단반응에 대한pH의 영향을 나타내는 그래프이다.
도 5는 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단반응에 대한 염화나트륨, 염화칼륨, 및 글루타민산칼륨의 영향을 나타내는 그래프이다.
도 6은 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단반응에 대한 염화 마그네슘의 영향을 나타내는 그래프이다.
도 7은 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단반응에 대한 염화 망간의 영향을 나타내는 그래프이다.
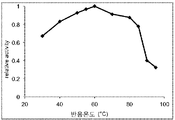
도 8은 본 발명의 미스매치 엔도뉴클레아제의 미스매치 DNA 절단반응 온도의 영향을 나타내는 그래프이다.
도 9는 본 발명의 미스매치 엔도뉴클레아제가 다양한 미스매치 DNA에 대한 절단활성을 나타내는 그래프이다.
도 10은 본 발명의 미스매치 엔도뉴클레아제의 DNA결합활성 측정에서 사용한 프로브 DNA의 일람이다.
도 11은 본 발명의 미스매치 엔도뉴클레아제의 DNA 결합 활성 측정을 나타내는 Native-PAGE의 결과이다.
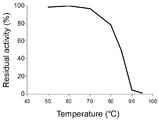
도 12는 본 발명의 미스매치 엔도뉴클레아제의 내열성을 나타내는 그래프이다.
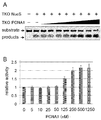
도 13은 본 발명의 미스매치 엔도뉴클레아제의 TKO 유래 PCNA1 존재 하 미스매치 DNA 절단활성을 나타내는 Native-PAGE 결과와 그래프이다.
도 14는 본 발명의 미스매치 엔도뉴클레아제의 A-A 미스매치 DNA에 대한 절단활성을 나타내는 Native-PAGE의 결과이다.
Claims (10)
- 더블-스트랜드 핵산의 절단방법으로서, 하기 (i)∼(iii)으로 이루어지는 그룹에서 선택된 적어도 1종의 폴리펩티드를 미스매치 염기쌍을 가지는 더블-스트랜드 핵산에 작용시키고, 당해 더블-스트랜드 핵산의 양쪽 스트랜드를 G-G, G-T 혹은 T-T 미스매치 염기쌍부위에서 인식해서 절단하는 것을 특징으로 하는 방법:
(i) 서열목록의 서열번호 1에 기재된 아미노산 서열을 가지는 폴리펩티드;
(ii) 서열목록의 서열번호 1에 기재된 아미노산 서열에 있어서 1 내지 수개의 아미노산 잔기가 치환, 결실, 삽입, 및/또는 부가되어 이루어지는 아미노산 서열을 가지며, 또 G-G, G-T 혹은 T-T 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드; 및
(iii) 서열목록의 서열번호 1에 기재된 아미노산 서열에 대해서 95% 이상의 아미노산 서열의 동일성을 가지는 아미노산 서열을 가지며, 또 G-G, G-T 혹은 T-T 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드. - 하기 (a)∼(c)를 포함하는 조성물:
(a) DNA 폴리머라아제;
(b) 적어도 한 쌍의 올리고뉴클레오티드 프라이머; 및
(c) 하기 (i)∼(iii)으로 이루어지는 그룹에서 선택된 적어도 일종의 폴리펩티드:
(i) 서열목록의 서열번호 1에 기재된 아미노산 서열을 가지는 폴리펩티드;
(ii) 서열목록의 서열번호 1에 기재된 아미노산 서열에 있어서 1 내지 수개의 아미노산 잔기가 치환, 결실, 삽입, 및/또는 부가되어 이루어지는 아미노산 서열을 가지며, 또 G-G, G-T 혹은 T-T 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드; 및
(iii) 서열목록의 서열번호 1에 기재된 아미노산 서열에 대해서 95% 이상의 아미노산 서열의 동일성을 가지는 아미노산 서열을 가지며, 또 G-G, G-T 혹은 T-T 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드. - 핵산의 증폭 방법으로서, 하기 (a)∼(b)의 공정을 포함하는 방법:
(a) 제2 항에 기재된 조성물과 주형이 되는 핵산분자를 포함하는 조성물을 조제하는 공정; 및
(b) 공정(a)에 의해 수득된 조성물을 적절한 조건하에서 반응시키고, 핵산 증폭을 실시하는 공정. - 하기 (a)∼(C)로 이루어지는 그룹에서 선택되는 폴리펩티드:
(A) 서열목록의 서열번호 3에 기재된 아미노산 서열을 가지는 폴리펩티드,
(B) 상기 (a)의 폴리펩티드의 아미노산 서열에 있어서, 47번째와 76번째의 아미노산 잔기를 제외하는 1 내지 수개의 아미노산 잔기가 치환, 결실, 삽입, 및/또는 부가되어 이루어지는 아미노산 서열을 가지며, 또 A-A, A-C 혹은 C-C 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드; 및
(C) 서열목록의 서열번호 3에 기재된 아미노산 서열에 대해서 90% 이상의 아미노산 서열의 동일성을 가지는 아미노산 서열에 있어서, 서열목록의 서열번호 1에 기재된 아미노산 서열에 있어서의 47번째의 세린과 76번째의 아스파라긴에 대응하는 아미노산 잔기가 다른 아미노산 잔기로 치환되어 이루어지는 아미노산 서열을 가지며, 또 A-A, A-C 혹은 C-C 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드. - 하기 (a)∼(c)를 포함하는 조성물:
(a) DNA 폴리머라아제;
(b) 적어도 한 쌍의 올리고뉴클레오티드 프라이머; 및
(c) 하기 (i)∼(iii)으로 이루어지는 그룹에서 선택된 적어도 1종의 폴리펩티드:
(i) 서열목록의 서열번호 3에 기재된 아미노산 서열을 가지는 폴리펩티드;
(ii) 상기 (i)의 폴리펩티드의 아미노산 서열에 있어서, 47번째와 76번째의 아미노산 잔기를 제외하는 1 내지 수개의 아미노산 잔기가 치환, 결실, 삽입, 및/또는 부가되어 이루어지는 아미노산 서열을 가지며, 또 A-A, A-C 혹은 C-C 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드; 및
(iii) 서열목록의 서열번호 3에 기재된 아미노산 서열에 대해서 90% 이상의 아미노산 서열의 동일성을 가지는 아미노산 서열에 있어서, 서열목록의 서열번호 1에 기재된 아미노산 서열에 있어서의 47번째의 세린과 76번째의 아스파라긴에 대응하는 아미노산 잔기가 다른 아미노산 잔기로 치환되어 이루어지는 아미노산 서열을 가지며, 또 A-A, A-C 혹은 C-C 미스매치를 인식해서 절단하는 미스매치 엔도뉴클레아제 활성을 가지는 폴리펩티드. - 핵산의 증폭 방법으로서, 하기 (a)∼(b)의 공정을 포함하는 방법:
(a) 제5 항에 기재된 조성물과 주형이 되는 핵산분자를 포함하는 조성물을 조제하는 공정; 및
(b) 공정(a)에 의해 수득된 조성물을 적절한 조건하에서 반응시키고, 핵산 증폭을 실시하는 공정. - 핵산 증폭 반응에 있어서 특정한 염기서열을 가지는 핵산의 증폭을 억제하는 방법으로서, 하기 (a)∼(d)의 존재 하에서 핵산 증폭 반응을 실시하는 공정을 포함하는 방법:
(a) 상기 특정의 염기서열을 가지는 핵산 또는 그 상보 스트랜드와 하이브리드화시켰을 때에 1개 혹은 수개의 미스매치를 발생하도록 설계된 올리고데옥시리보뉴클레오티드;
(b) DNA 폴리머라아제;
(c) 적어도 한 쌍의 올리고뉴클레오티드 프라이머; 및
(d) 제1 항에 기재된 방법에서 사용하는 폴리펩티드 및/또는 제4 항에 기재된 폴리펩티드. - 표적 핵산을 우선적으로 증폭하는 방법으로서, 당해 표적 핵산과 1개 혹은 수개의 염기가 다른 염기서열의 핵산 증폭을 제7 항에 기재된 방법에서 억제하는 것을 특징으로 하는 방법.
- 제8 항에 기재된 방법으로서, 추가로 내열성균 유래의 증식세포 핵항원(PCNA) 혹은 그 호모로그의 존재 하에서 표적 핵산의 증폭을 실시하는 것을 특징으로 하는 방법.
- 제1 항에 기재된 방법에서 사용하는 폴리펩티드 및/또는 제4 항에 기재된 폴리펩티드를 사용한 표적 핵산의 돌연변이 검출방법.
Applications Claiming Priority (3)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| JPJP-P-2014-184934 | 2014-09-11 | ||
| JP2014184934 | 2014-09-11 | ||
| PCT/JP2015/075603 WO2016039377A1 (ja) | 2014-09-11 | 2015-09-09 | 耐熱性のミスマッチエンドヌクレアーゼの利用方法 |
Publications (1)
| Publication Number | Publication Date |
|---|---|
| KR20170053636A true KR20170053636A (ko) | 2017-05-16 |
Family
ID=55459121
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| KR1020177007069A Abandoned KR20170053636A (ko) | 2014-09-11 | 2015-09-09 | 내열성의 미스매치 엔도뉴클레아제의 이용방법 |
Country Status (6)
| Country | Link |
|---|---|
| US (1) | US10975415B2 (ko) |
| EP (1) | EP3192868A4 (ko) |
| JP (1) | JP6550649B2 (ko) |
| KR (1) | KR20170053636A (ko) |
| CN (1) | CN107002067A (ko) |
| WO (1) | WO2016039377A1 (ko) |
Families Citing this family (13)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP6009649B2 (ja) | 2013-03-14 | 2016-10-19 | タカラバイオ株式会社 | 耐熱性のミスマッチエンドヌクレアーゼの利用方法 |
| CN106604994B (zh) | 2014-06-23 | 2021-12-14 | 通用医疗公司 | 通过测序评估的DSBs的全基因组无偏鉴定(GUIDE-Seq) |
| KR20170053636A (ko) | 2014-09-11 | 2017-05-16 | 다카라 바이오 가부시키가이샤 | 내열성의 미스매치 엔도뉴클레아제의 이용방법 |
| EP3272865A4 (en) * | 2015-03-20 | 2018-10-17 | Takara Bio Inc. | High-sensitivity method for detecting target nucleic acid |
| WO2019005463A1 (en) * | 2017-06-28 | 2019-01-03 | New England Biolabs, Inc. | METHOD FOR REMOVING AND / OR DETECTING NUCLEIC ACIDS HAVING MESAPPARATED NUCLEOTIDES |
| WO2019075197A1 (en) * | 2017-10-11 | 2019-04-18 | The General Hospital Corporation | METHODS OF DETECTION OF INDIVIDUAL SITE-SPECIFIC PARASITE GENOMIC DEAMINATION BY BASE EDITING TECHNOLOGIES |
| US11898203B2 (en) | 2018-04-17 | 2024-02-13 | The General Hospital Corporation | Highly sensitive in vitro assays to define substrate preferences and sites of nucleic-acid binding, modifying, and cleaving agents |
| EP4017994A1 (en) * | 2019-08-22 | 2022-06-29 | New England Biolabs, Inc. | Cleavage of single stranded dna having a modified nucleotide |
| EP4114972A1 (en) * | 2020-03-06 | 2023-01-11 | Life Technologies Corporation | High sequence fidelity nucleic acid synthesis and assembly |
| JP7511832B2 (ja) * | 2020-03-13 | 2024-07-08 | 国立大学法人九州大学 | 耐熱性ミスマッチエンドヌクレアーゼによる核酸の切断方法 |
| CN119242611A (zh) | 2020-03-19 | 2025-01-03 | 宝生物工程株式会社 | 耐热的错配核酸内切酶变体 |
| CN111748610A (zh) * | 2020-06-24 | 2020-10-09 | 西北农林科技大学 | T7EI-like核酸酶反应缓冲体系、DNA错配检测方法及应用 |
| CN119343466A (zh) * | 2022-06-16 | 2025-01-21 | 深圳华大智造科技股份有限公司 | 含甘氨酸-NaOH缓冲液的切除试剂及其在核酸测序中的应用 |
Family Cites Families (34)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| US3395021A (en) | 1964-10-14 | 1968-07-30 | Gen Foods Corp | Beverage powder producing pulpy mouth feel when dissolved |
| US4889818A (en) | 1986-08-22 | 1989-12-26 | Cetus Corporation | Purified thermostable enzyme |
| US6582908B2 (en) | 1990-12-06 | 2003-06-24 | Affymetrix, Inc. | Oligonucleotides |
| US5602011A (en) * | 1995-01-18 | 1997-02-11 | Pharmacia Biotech Inc. | Purified Thermococcus barossii DNA polymerase |
| US6428955B1 (en) | 1995-03-17 | 2002-08-06 | Sequenom, Inc. | DNA diagnostics based on mass spectrometry |
| AUPN245295A0 (en) | 1995-04-13 | 1995-05-11 | Johnson & Johnson Research Pty. Limited | Assay for genetic abnormalities |
| WO1997021837A1 (en) | 1995-12-15 | 1997-06-19 | Amersham Life Science, Inc. | Methods for the detection and removal of mutant sequences that arise during enzymatic amplification using mismatch repair systems |
| EP0964929B1 (en) | 1996-06-05 | 2004-10-20 | Fox Chase Cancer Center | Mismatch endonucleases and uses thereof in identifying mutations in targeted polynucleotide strands |
| WO1999042595A1 (en) | 1998-02-19 | 1999-08-26 | Trevigen, Inc. | Mismatch cleavage enzymes from extreme thermophiles and uses thereof |
| ATE342378T1 (de) * | 1999-03-19 | 2006-11-15 | Cornell Res Foundation Inc | Verfahren das auf den gekoppelten reactionen der polymerase-kettenreaktion (pcr)-verdauung mit restriktionsenzymen und ligasenachweis basiert |
| AUPQ495700A0 (en) * | 2000-01-05 | 2000-02-03 | Johnson & Johnson Research Pty. Limited | Method for concurrent amplification and real time detection of polymorphic nucleic acid sequences |
| US20020128215A1 (en) | 2000-02-02 | 2002-09-12 | Hans-Ulrich Thomann | Novel sequence variants of the human N-acetyltransferase -2 (NAT -2) gene and use thereof |
| AU2001247212B2 (en) * | 2000-02-22 | 2005-12-22 | Fox Chase Cancer Center | Nucleic acid molecule encoding a mismatch endonuclease and methods of use thereof |
| US6391557B1 (en) | 2000-02-22 | 2002-05-21 | Fox Chase Cancer Center | Nucleic acid molecule encoding a mismatch endonuclease and methods of use thereof |
| AU2002230526A1 (en) | 2000-12-01 | 2002-06-11 | Cornell Research Foundation, Inc. | Detection of nucleic acid differences using combined endonuclease cleavage and ligation reactions |
| EP1347060A4 (en) | 2000-12-26 | 2004-08-18 | Takara Bio Inc | METHOD FOR DETECTION OF A PATHOGENIC MICROORGANISM |
| EP1367136A4 (en) | 2001-02-15 | 2005-01-12 | Takara Bio Inc | METHOD FOR DETECTING NUCLEOTIDE POLYMORPHISM |
| JPWO2002101042A1 (ja) | 2001-06-12 | 2005-04-07 | タカラバイオ株式会社 | 核酸増幅又は検出反応用試薬の安定化方法ならびに保存方法 |
| US20030143605A1 (en) | 2001-12-03 | 2003-07-31 | Si Lok | Methods for the selection and cloning of nucleic acid molecules free of unwanted nucleotide sequence alterations |
| CA2497338A1 (en) | 2002-08-30 | 2004-03-18 | Japan Science And Technology Corporation | Method of targeted gene disruption, genome of hyperthermostable bacterium and genome chip using the same |
| JP2008520245A (ja) | 2004-11-23 | 2008-06-19 | ワン,シャオ・ビン | 切断−増幅法による核酸多様性の検出 |
| US20100291548A1 (en) | 2006-03-12 | 2010-11-18 | Applera Corporation | Methods of Detecting Target Nucleic Acids |
| JP2007295838A (ja) | 2006-04-28 | 2007-11-15 | Fujifilm Corp | 核酸増幅を利用した核酸ミスマッチ検出方法 |
| JP2007319096A (ja) | 2006-06-01 | 2007-12-13 | Institute Of Physical & Chemical Research | エンドヌクレアーゼのニッキング活性を利用した核酸増幅法 |
| JP4918409B2 (ja) | 2006-07-26 | 2012-04-18 | 西川ゴム工業株式会社 | 核酸配列の増幅方法 |
| WO2011102802A1 (en) | 2010-02-18 | 2011-08-25 | Agency For Science, Technology And Research | Method for reducing mismatches in double-stranded dna molecules |
| KR101312241B1 (ko) | 2010-04-27 | 2013-09-27 | 사회복지법인 삼성생명공익재단 | 증폭억제시발체를 이용하는 유전자 돌연변이 검출 방법 |
| WO2012064975A1 (en) | 2010-11-12 | 2012-05-18 | Gen9, Inc. | Protein arrays and methods of using and making the same |
| US10424394B2 (en) | 2011-10-06 | 2019-09-24 | Sequenom, Inc. | Methods and processes for non-invasive assessment of genetic variations |
| EP3597764A3 (en) | 2012-02-01 | 2020-05-06 | SGI-DNA, Inc. | Material and methods for the synthesis of error-minimized nucleic acid molecules |
| US20150353921A9 (en) | 2012-04-16 | 2015-12-10 | Jingdong Tian | Method of on-chip nucleic acid molecule synthesis |
| JPWO2013175815A1 (ja) | 2012-05-22 | 2016-01-12 | 国立研究開発法人理化学研究所 | 核酸増幅反応におけるエラーを抑制する方法 |
| JP6009649B2 (ja) | 2013-03-14 | 2016-10-19 | タカラバイオ株式会社 | 耐熱性のミスマッチエンドヌクレアーゼの利用方法 |
| KR20170053636A (ko) | 2014-09-11 | 2017-05-16 | 다카라 바이오 가부시키가이샤 | 내열성의 미스매치 엔도뉴클레아제의 이용방법 |
-
2015
- 2015-09-09 KR KR1020177007069A patent/KR20170053636A/ko not_active Abandoned
- 2015-09-09 WO PCT/JP2015/075603 patent/WO2016039377A1/ja not_active Ceased
- 2015-09-09 JP JP2016547473A patent/JP6550649B2/ja not_active Expired - Fee Related
- 2015-09-09 US US15/507,796 patent/US10975415B2/en not_active Expired - Fee Related
- 2015-09-09 CN CN201580048826.8A patent/CN107002067A/zh active Pending
- 2015-09-09 EP EP15839690.3A patent/EP3192868A4/en not_active Withdrawn
Also Published As
| Publication number | Publication date |
|---|---|
| JPWO2016039377A1 (ja) | 2017-06-22 |
| JP6550649B2 (ja) | 2019-07-31 |
| CN107002067A (zh) | 2017-08-01 |
| US20170253909A1 (en) | 2017-09-07 |
| US10975415B2 (en) | 2021-04-13 |
| WO2016039377A1 (ja) | 2016-03-17 |
| EP3192868A4 (en) | 2018-03-14 |
| EP3192868A1 (en) | 2017-07-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| JP6550649B2 (ja) | 耐熱性のミスマッチエンドヌクレアーゼの利用方法 | |
| US20250340914A1 (en) | Polynucleotide enrichment using crispr-cas system | |
| US10294465B2 (en) | Method for using heat-resistant mismatch endonuclease | |
| EP2825645B1 (en) | Methods and compositions for discrimination between cytosine and modifications thereof, and for methylome analysis | |
| KR20230116944A (ko) | 고온 내성 Cas 단백질의 용도, 표적 핵산 분자의 검출방법 및 시약 키트 | |
| CN110234770A (zh) | 通过dna单链断裂识别碱基编辑脱靶位点的方法 | |
| US20180163263A1 (en) | High-sensitivity method for detecting target nucleic acid | |
| CN115210380B (zh) | 耐热的错配核酸内切酶变体 | |
| US20250146059A1 (en) | Method and Reagent Kit for Targeted Genomic Enrichment |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PA0105 | International application |
Patent event date: 20170314 Patent event code: PA01051R01D Comment text: International Patent Application |
|
| PG1501 | Laying open of application | ||
| A201 | Request for examination | ||
| A302 | Request for accelerated examination | ||
| PA0201 | Request for examination |
Patent event code: PA02012R01D Patent event date: 20200903 Comment text: Request for Examination of Application |
|
| PA0302 | Request for accelerated examination |
Patent event date: 20200903 Patent event code: PA03022R01D Comment text: Request for Accelerated Examination |
|
| E902 | Notification of reason for refusal | ||
| PE0902 | Notice of grounds for rejection |
Comment text: Notification of reason for refusal Patent event date: 20201111 Patent event code: PE09021S01D |
|
| E701 | Decision to grant or registration of patent right | ||
| PE0701 | Decision of registration |
Patent event code: PE07011S01D Comment text: Decision to Grant Registration Patent event date: 20210427 |
|
| NORF | Unpaid initial registration fee | ||
| PC1904 | Unpaid initial registration fee |