[0001] A presente invenção diz respeito a um método para medir a polimerização de DNA e suas aplicações. Mais precisamente, a invenção diz respeito a um método para medir a polimerização de DNA dependente do DNA.
FUNDAMENTOS
[0002] As últimas décadas têm experimentado um desenvolvimento rápido de novos métodos para a medição da transcrição reversa, polimerização de DNA dependente do RNA. O ponto mais complexo em relação aos métodos para a quantificação da polimerização de DNA dependente do DNA tem até agora recebido muito menos atenção.
[0003] Os ensaios da atividade da DNA polimerase clássica envolvem o uso de DNA tratado com DNAse (“DNA ativado”) como iniciador / padrão e a incorporação de nucleotídeos radiorrotulados no DNA (Aposhian and Kornberg 1962). A medição de radioatividade precipitável por ácido permite o cálculo da quantidade de nucleotídeos incorporada e o número de unidades de enzima presentes. No entanto, o uso de radioatividade é correntemente restrito e desaconselhado em muitos laboratórios e existe devido a isto uma tendência geral longe das técnicas com base em radioatividade.
[0004] Para as DNA polimerases um ensaio comercial com base na detecção ELISA de nucleotídeos rotulados por digoxigenina incorporados no DNA recentemente produzido está disponível (Roche Molecular Biochemicals Cat. No. 1468120, US5635350). Este ensaio é dificultado pelo uso de dois análogos de substrato de nucleotídeo diferentes com grupos volumosos, digoxigenina como rótulo e biotina para a imobilização do produto. Como um resultado a velocidade da reação de polimerização e subsequente sensibilidade de detecção são reduzidas. A utilização de análogos de substrato com propriedades cinéticas altamente divergentes toma este sistema menos relevante para estudos da suscetibilidade ao medicamento de diferentes polimerases.
[0005] Outro método alternativo mais atrativo é um ensaio com base na fluorescência para holoenzima DNA polimerase, baseado na reação específica da tintura PicoGreen com DNA trançado em filamento duplo (Seville et al. 1996). O último processo foi recentemente modificado para torná-lo adequado para uma faixa mais ampla de diferentes enzimas de polimerização do DNA (Tveit and Kristensen 2001). Este ensaio é tecnicamente simples e se baseia na utilização de nucleotídeos naturais. A sensibilidade de detecção está, no entanto, ainda na mesma faixa como o ensaio de DNA polimerase radioativo clássico e os pedidos descritos demonstram uma faixa de 0,05 a 0,5 U de DNA polimerase / amostra.
[0006] A terapia para HIV hoje se baseia na terapia de múltiplos medicamentos. Os regimes baseiam-se nas combinações de todos os três tipos de medicamentos disponíveis: análogos nucleosídeos, análogos não nucleosídeos e inibidores da protease. A estratégia é minimizar a probabilidade de um vírus mutante sobreviver.
[0007] Os inibidores da transcriptase reversa (RT) são ou análogos nucleosídeos ou análogos não nucleosídeos. Os inibidores não nucleosídeos se ligam a uma bolsa hidrofóbica na enzima RT intimamente ligada, mas não contígua, ao sítio ativo. A replicação do HIV-1 é inibida alostericamente mediante o deslocamento dos resíduos de aspartato catalíticos em relação ao sítio de ligação da polimerase.
[0008] Os inibidores de nucleosídeo usados hoje finalizam o alongamento da cadeia de DNA quando eles carecem de um grupo de 3’-hidroxila. A terapia prolongada com inibidores de nucleosídeo comumente leva ao desenvolvimento de vírus resistente. Este processo é associado com o aparecimento gradual de mutações no gene de ligação do vírus, cada uma levando a substituições definidas de aminoácido (para uma recapitulação ver Vandamme et al. 1998). Os efeitos destas substituições nos níveis enzimáticos são complicados e incluem a intensificação de uma função de correção do DNA primitiva. Esta reação é dependente do nucleotídeo e produz polifosfato de dinucleosídeo e uma extremidade 3’ no DNA ampliável (Arion et al. 1998, Meyer et al. 1999).
[0009] A RT do HIV-1 assim como outras transcriptases reversas executam três reações enzimáticas diferentes: polimerização de DNA dependente do RNA, polimerização de DNA dependente do DNA e degradação do RNA no híbrido DNA- RNA (RNase H). A transcriptase reversa do HIV, codificada pelo gene de ligação, é um heterodímero consistindo de uma subunidade p66 e p51. Tanto a polimerização de DNA dependente do RNA quanto a polimerização de DNA dependente do DNA são executadas pelo mesmo sítio ativo localizado na subunidade p66 (para uma recapitulação ver Goff 1990). O mecanismo de reação destes medicamentos foi principalmente definido de acordo com a sua ação sobre a reação de polimerização de DNA dependente do RNA. O efeito sobre a reação de polimerização de DNA dependente do DNA é comparativamente menos estudada.
[0010] Contanto que o mecanismo de reação e o medicamento metabolizado ativo sejam conhecidos e disponíveis, a suscetibilidade do medicamento ao vírus fenotípico pode ser determinada no nível da enzima. Dependendo da capacidade dos ensaios de enzima e das técnicas de isolamento do vírus usadas, o teste de sensibilidade ao medicamento pode teoricamente ser executado ou sobre os sobrenadantes da propagação na cultura do vírus, o isolamento do vírus primário ou sobre as preparações de vírus recuperadas diretamente dos pacientes. O ensaio de atividade da RT convencional é executado mediante a utilização de uma construção padrão- iniciador artificial e trifosfato de deoxinucleosídeo rotulado como substrato de nucleotídeo. O par de padrão / iniciador poli(rA) / oligo(dT) é o mais eficiente e a combinação mais usada para determinação do HIV, assim como para outras RTs retrovirais. Uma desvantagem deste tipo de ensaio quando o teste de sensibilidade ao medicamento está envolvido, é que somente os análogos não nucleosídeos ou análogos que podem ter pareamento de base com rA podem ser testados. Os análogos com as outras bases de nucleotídeo necessitarão de um ensaio com base em um padrão de polímero variável. Os polímeros de RNA contendo bases de pirimidina são notoriamente sensíveis às RNases e na prática não compatíveis com as amostras biológicas. Deve portanto ser vantajoso estabelecer um ensaio de polimerase destinado para teste de sensibilidade ao medicamento sobre um padrão de DNA variável, contanto que o sistema de ensaio ofereça resultados que se correlacionam com aqueles da inibição de transcrição reversa e testes de resistência ao medicamento fenotípicos.
[0011] A terapia do HIV usada hoje é apenas um exemplo da potência de inibidores da DNA polimerase. A situação corrente com relação ao desenvolvimento da resistência entre as bactérias e outros microorganismos motiva a procura por novas classes de medicamentos antimicrobianos. As DNA polimerases são uns dos alvos principais durante esta realização. Como tal existe uma grande demanda para ensaios de polimerase tecnicamente simples, que não causam riscos ambientais potenciais e podem ser aplicados para a triagem de medicamentos sobre uma ampla faixa de isoenzimas DNA polimerases microbianas. A toxicidade do medicamento mostra que a descoberta deve ainda ser avaliada com respeito às DNA polimerases de mamífero correspondentes.
[0012] A quantificação das polimerases associadas com a proliferação tais como oc e δ polimerase pode ser usadas para monitorar a proliferação celular. Pode ser mencionado neste contexto que os níveis de soro de timidina cinase, uma outra enzima associada com a proliferação celular, são correntemente usados para prognóstico e classificação de doenças virulentas (US4637977). A fosforilação de timidina é exatamente aquela dos dois caminhos sintéticos intracelulares, que fornece trifosfato de timidina para a síntese do DNA. A medição da própria DNA polimerase tem o potencial de dar uma estimativa mais correta da síntese total do DNA comparada com a atividade da timidina cinase ou incorporação de timidina. DESCRIÇÃO DA INVENÇÃO
[0013] A presente invenção fornece um ensaio de DNA polimerase não radioativa em formato de placa de microtítulo que possibilita a detecção do produto calorimétrico e fluorimétrico.
[0014] Em uma modalidade preferida ela utiliza, como substrato de trifosfato de nucleosídeo, 5-bromodeoxiuridina 5’-trifosfato (BrdUTP). A diferença no raio de Van der Waals entre o bromo em BrdUTP na posição 5’ e o grupo de metila em trifosfato de timidina na posição 5’ é mínima (1,95 A comparado com 2,0 A) e as propriedades cinéticas da enzima destes dois nucleotídeos são completamente similares. O método pode ser aromatizado e possui uma faixa de detecção abaixo da atividade 3nU polimerase / amostra.
[0015] Uma das aplicações da presente invenção é o teste da suscetibilidade ao medicamento. Todos os medicamentos anti-retrovirais aprovados até agora interferem com a reação enzimática da protease viral ou da RT. Existem além disso medicamentos candidatos na linha de serviço que afetam a função da integrase retroviral.
[0016] Em particular, a presente invenção fornece um procedimento para medir uma ampla faixa de diferentes DNA polimerases dependentes do DNA. Tem se demonstrado adequado até para estudos de sistemas de DNA polimerase altamente processáveis apesar do padrão comparativamente curto usado. A utilidade para a determinação da atividade de polimerase bacteriana I e III, DNA polimerase de mamífero oc, β e y, uma atividade de polimerase associada com a proliferação no soro humano e polimerização de DNA dependente do DNA por RT do HIV é demonstrada, mas o método pode ser usado para estudos de DNA polimerases virais e celulares virtualmente completas. Um dos aspectos que distinguem o presente ensaio de DNA polimerase da técnica anterior é sua sensibilidade destacada, que torna a detecção de até 3nU de atividade DNA polimerase I do E. coli possível.
[0017] Assim, um aspecto da invenção é direcionado a um método para medira a polimerização de DNA dependente do DNA em uma amostra biológica, compreendendo as etapas de a) fornecer um iniciador com uma sequência específica curta trançada em filamentos única, o qual é incapaz de pareamento interno de bases, ligadas a uma fase sólida, b) colocar em contato a construção do iniciador com uma mistura de reação contendo um padrão de deoxinucleotídeo trançado em filamentos único com uma parte da sequência complementar no iniciador e os quatro trifosfatos de deoxinucleosídeo, um dos quais é modificado de modo que seja especificamente reconhecido por um anticorpo rotulado, c) adicionar uma amostra biológica que compreende a DNA polimerase na mistura de b), d) deixar a reação de polimerase prosseguir, e) incubar o produto da reação imobilizado resultante de d) com o anticorpo rotulado, f) detectar a quantidade de anticorpo rotulado ligado, e g) medir a quantidade de deoxinucleosídeo modificado incorporado, como uma medida da polimerização de DNA, com o auxílio do rótulo do anticorpo ligado.
[0018] Em uma modalidade a polimerização de DNA é produzida por uma transcriptase reversa (RT) de retrovirus, tal como a RT do vírus da imunodeficiência humana.
[0019] Em outra modalidade o trifosfato de deoxinucleosídeo modificado é 5- bromodeoxiuridina 5’-trifosfato (BrdllTP) e o anticorpo rotulado é um anticorpo monoclonal anti-BrdU conjugado porfosfatase alcalina (Ap).
[0020] Em uma modalidade preferida do método de acordo com a invenção, a polimerização do DNA medida é usada para o teste de suscetibilidade ao medicamento.
[0021] O teste de suscetibilidade ao medicamento é executado para avaliar se um certo medicamento é eficaz em um indivíduo mamífero, e o resultado pode ser usado para a seleção da terapia de tratamento de medicamento para este indivíduo. Na prática, o indivíduo será submetido ao teste em vários pontos de tempo para monitorar o desenvolvimento do tratamento de medicamento em dito indivíduo.
[0022] A invenção é também direcionada a uma embalagem comercial que compreende instruções escritas ou portadoras de dados para medira a polimerização de DNA dependente do DNA de acordo com a invenção. A embalagem compreenderá pelo menos os seguintes itens: a) um iniciador com uma sequência específica curta trançada em filamentos única, o qual é incapaz de pareamento interno de bases, ligadas a uma fase sólida, b) um padrão de deoxinucleotídeo trançado em filamentos único com uma parte da sequência complementar no iniciador em a), c) os quatro trifosfatos de deoxinucleosideo, um dos quais é modificado de modo que seja especificamente reconhecido por um anticorpo rotulado, e d) o anticorpo rotulado que reconhece o trifosfato de deoxinucleosideo modificado em c).
[0023] A invenção agora será ilustrada pela descrição não limitativa que segue das modalidades e desenhos da invenção.
[0024] Os ensinamentos da literatura citada é aqui incorporada por referência. BREVE DESCRIÇÃO DOS DESENHOS
[0025] A Figura 1 demonstra os efeitos da variação na sequência do padrão sobre a inibição da DNA polimerase III com TMAU.
[0026] A Figura 2. exemplifica a sensibilidade de detecção do ensaio da DNA polimerase. RT tipo silvestre do HIV-1 (♦), DNA polimerase β de mamífero (•) e DNA polimerase I de E. coli (A).
[0027] A Figura 3. demonstra a capacidade do ensaio da DNA polimerase em medir a inibição pelos análogos de dideóxi em todas as quatro bases de DNA. Símbolos ddATP (□), ddGTT (♦), ddCTP (•) e ddTP (A).
[0028] A Figura 4. mostra que a resistência fundamental do mecanismo bioquímico ao tenofovir do medicamento antiviral baseia-se na intensificação de uma reação de fosforolise dependente do ATP. Símbolos: RT tipo silvestre do HIV-1 em solução de reação padrão (O). RT tipo silvestre do HIV-1 em solução de reação com ATP (•), RT mutante do HIV-1 em solução de reação padrão (0), RT mutante do HIV-1 em solução de reação com ATP (♦).
[0029] A Figura 5. exemplifica a determinação de suscetibilidade ao medicamento antiviral Nevirapine usando RTs isoladas do plasma de indivíduos infectados por HIV. Símbolos: (| [), (□) e (A) RTs de indivíduos infectados (•) um controle consistindo de RT tipo silvestre de HIV-1 recombinante (♦) um controle consistindo de RT do HIV-1 recombinante modificado (L100I) com resistência a Nevirapine intermediária.
DESCRIÇÃO DAS MODALIDADES
[0030] Produção de placas de microtítulos revestidas por iniciador
[0031] Cloridreto de 1-etil-3-(3-dimetilamino-propila)carboimida (concentração final 10 mg/ml) foi adicionado a um tamponante de 1 -metilimidazol 100 mM (pH 7,0) e a mistura foi usada para diluir a construção do iniciador em uma concentração final de 1 jig/ml. 100 |1I da solução iniciadora foi fracionada para cada reservatório de uma placa de microtítulo que consiste de tiras transparentes Nalge Nunc NucleoLink® (Cat no 248259). As placas foram incubadas de 6 a 8 horas em 37°C, lavadas completamente em 2M NaOH com ácido elilenodiaminatetraacético (EDTA) 2 mM e infiltradas em três frascos de 5 litros com água. O fluido residual nos reservatórios foi removido mediante o esvaziamento por punção das placas de cabeça para baixo sobre tecido ou papel absorvente. As placas foram deixadas secar por 30 min em temperatura ambiente e finalmente congeladas para armazenagem em -20°C.
[0032] Protocolo para o ensaio de DNA polimerase
[0033] O ensaio de DNA polimerase baseia-se em um iniciador concentrado com uma sequência específica que é covalentemente ligada aos reservatórios de uma placa de microtítulo de 96 reservatórios. A mistura de reação contém um padrão de deoxinucleotídeo trançado em filamentos único com uma parte da sequência complementar no iniciador e os quatro trifosfatos de deoxinucleosídeo. O trifosfato de timidina é, no entanto, substituído por 5-bromodeoxiuridina 5’-trifosfato (BrdllTP). A quantidade de monofosfato de bromodeoxiuridina (BrdUMP) incorporada dentro do DNA durante a reação da polimerase, é detectada com um anticorpo monoclonal anti- BrdU conjugado com fosfatase alcalina (Ap). Um substrato de Ap, fosfato de 4- metilumbeliferila, é usado para a detecção do produto fluorimétrico.
[0034] 100 |1I da mistura de reação da DNA polimerase foi adicionado a cada reservatório das placas de microtítulo revestido com iniciador. As amostras foram diluídas em tamponante de base DNA polimerase e a reação de polimerase foi iniciada mediante a transferência de 50 jil de diluição da amostra para cada reservatório na placa. A placa de microtítulo foi incubada em 33°C e a reação foi finalizada após os tempos indicados por lavagem da placa com tamponante de borato 3 mM (pH 8,9) com 1,5 % (v/v) de octofenoxipolietoxietanol (Triton X-100). Geralmente dois tempos de incubação, 4 horas e durante a noite (16 horas), foram usados para verificar a linearidade da reação de polimerização. As placas foram lavadas completamente em NaOH 2 M com EDTA 2 mM e infiltradas em três frascos de 5 litros com água.
[0035] Logo depois as placas foram incubadas por 90 minutos em 33°C com 100 pl de anticorpo monoclonal anti-Brdll conjugado por fosfatase alcalina (Ap) diluído com 4,8 |ig/ml em tampão de (bis[2-hidroxietila]iminotris[hidroximetila]metano;2-bis[2- hidroxietila]amino-2-[hidroximetila]-1,3-propanodiol) (Bis Tris) 25 mM (pH 7,2) com NaCI 50 mM, (NhU^SCM 37,5 mM, 1 mg/ml de sulfato de dextrano, TritonX-100 a 1% e 25 mg/ml de leite em pó desnatado da Sigma.
[0036] As placas foram depois disso lavadas novamente com tampão de borato 3 mM (pH 8,9) com 1,5 % (v/v) de TritonX-100 para remover o anticorpo rotulado não ligado. A atividade da fosfatase alcalina foi determinada usando substrato de fosfato de 4-metilumbeliferila dissolvido em Tris-tamponante (pH 8,9). A fluorescência foi interpretada em 460 nm com uma leitora Wallac Victor 2 em intervalos definidos (excitação 355 nm).
[0037] Protocolo para a determinação de inibição da segunda síntese do filamento sobre o padrão de DNA variável.
[0038] Os estudos de inibição foram executados em um ensaio de DNA polimerase modificado. Os medicamentos foram diluídos em série em cinco etapas e alíquotas de 25 |1I foram transferidas para cada reservatório na placa de microtítulo, misturadas com 100 |1I da mistura de reação da DNA polimerase e a reação da enzima foi iniciada pela adição de 25 |1I de diluição da enzima. Os análogos não nucleosídeos foram estudados nas condições de reação padrão enquanto a concentração de todos os quatro trifosfatos de deoxinucleosídeo (dNTP) foi reduzida para 1 |1M nos estudos de inibidores competitivos dNTP. A reação de polimerase foi deixada prosseguir durante a noite (16 a 24 horas em 33°C). Depois disso, a reação foi finalizada por uma lavagem da placa. O valor ICso foi definido como a concentração de medicamento dando 50 % de inibição da atividade polimerase estudada.
[0039] Protocolo para a determinação da atividade de RT.
[0040] Uma modificação do ensaio de RT colorimétrico (kit da atividade de RT Cavidi® Lenti), disponível da Cavidi Tech, Uppsala, Sweden foi usada para a determinação do nível de atividade RT nas preparações de vírus estudadas. O método foi descrito (Ekstrand et al. 1996). Em resumo, poli(rA) covalentemente ligado aos reservatórios de uma placa de microtítulo de 96 reservatórios serve como padrão para a incorporação de 5-bromodeoxiuridina 5’-trifosfato (BrdUTP) durante a etapa de transcrição reversa em 33°C. A quantidade de monofosfato de bromodeoxiuridina (BrdUMP) incorporada dentro do DNA, é detectada com um anticorpo monoclonal anti- BrdU conjugado por fosfatase alcalina (Ap). Um substrato de Ap, fosfatase de 4- metilumbeliferila é finalmente usada para a detecção fluorimétrica.
[0041] Protocolo para a determinação de inibição da transcrição reversa.
[0042] Os estudos de inibição foram em um ensaio de RT Cavidi HS-kit Lenti. Os inibidores foram diluídos em série em cinco etapas e alíquotas de 25 pl foram transferidas para cada reservatório na placa de microtítulo, misturadas com 100 pl da mistura de reação da RT e a reação da enzima foi iniciada pela adição de 50 pl de diluição da enzima. A concentração de substrato de trifosfato de nucleosídeo (BrdUTP) final foi de 16 pM e a quantidade de iniciador (odT22) 12 ng per reservatório. A reação de RT foi deixada prosseguir durante a noite (16 a 24 horas em 33°C). Depois disso, a reação foi finalizada por uma lavagem da placa. O valor ICso foi definido como a concentração de medicamento dando 50 % de inibição da atividade RT estudada.
[0043] Protocolo para o isolamento da RT virai a partir do material que contém anticorpos bloqueadores da RT, com base na destruição das enzimas celulares solúveis seguido pelo isolamento da RT viral das mini colunas. 1) Rotular os tubos de plástico de 4,5 ml a serem usados. Colocá-los em uma caixa Nalgene. Adicionar 1 ml de amostra (por exemplo, plasma de EDTA de indivíduos infectados com o HIV) a cada tubo rotulado. Adicionar 100 pl de uma solução a 66 mM de 5,5’-ditiobis-(ácido 2- nitrobenzóico) em água tamponada, vórtice e incubar as amostras por uma hora em temperatura ambiente. A atividade das enzimas de plasma livres é destruída durante este procedimento enquanto as enzimas contidas dentro dos virions permanecem intactas. Os virions podem depois ser purificados a partir de 5,5’-ditiobis-(ácido 2-nitrobenzóico), anticorpos bloqueadores da atividade da enzima e outras substâncias que interferem com a quantificação da RT virai por vários procedimentos de separação. O protocolo abaixo baseia-se no uso de gel Fractogel® EMD TMAE Hicap. 2) Colocar em suspensão o gel de separação cuidadosamente e transferir 1500 pl da pasta de gel para cada tubo de pré-tratamento da amostra. 3) incubar as amostras com a pasta de gel por 90 minutos em temperatura ambiente com os tubos deitados horizontalmente sobre um agitador orbital. 4) Rotular a quantidade desejada de mini colunas plásticas e 10 ml para identificar as amostras sendo analisadas. Montar as colunas em um dispositivo de lavagem de colunas isto é, um tubo de distribuição a vácuo com extração da fase sólida Supelco Visiprep. Transferir os conteúdos dos tubos de ligação para suas colunas correspondentes. Antes de transferir turbilhonar o tubo rapidamente para de forma uniforme distribuir o gel. 5) Quando todas as colunas estiverem enchidas, aplicar o vácuo e absorver os géis secos. Desligar o vácuo e iniciar a lavagem mediante o enchimento de cada coluna com 9 ml de tamponante A. Quando todas as colunas tiverem sido enchidas, aplicar o vácuo e absorver os géis secos. 6) Repetir a etapa 5 três vezes mais, dando um total de quatro lavagens. Absorver os géis secos após cada lavagem. Após a absorção dos géis secos após as quatro lavagens, desligar o vácuo e prosseguir para a etapa 7. A etapa de lavagem remove os anticorpos bloqueadores da RT não ligada e o 5,5’-ditiobis-(ácido 2-nitrobenzóico) do sistema. 7) Adicionar a todos os géis secos 9 ml de tampão condicionador (B). Após um minuto aplicar vácuo e absorver os géis secos. 8) Repetir a etapa 7. Antes de desligar o vácuo verificar que todo o tampão condicionador (B) foi removido dos géis totais. 9) suspender verticalmente a parte superior do dispositivo de lavagem de coluna. Montar o recipiente de tubos com os tubos rotulados dentro de um recipiente limpo. Reequipar a parte superior do dispositivo. Controlar para que as tubagens pequenas de cada coluna desçam em seus tubos correspondentes. 10)Adicionar 600 |1I de tamponante de lise (C) a cada coluna. Deixar o tamponente repousar na coluna por cinco minutos. Depois aplicar o vácuo lentamente e absorver os géis secos. Isto dará em cada tubo aproximadamente 600 |1I de lisato de vírus do gel conectado.
[0044] A atividade de RT recuperada nos lisatos da etapa 10 é essencialmente livre de anticorpos bloqueadores de RT, medicamentos e atividade de polimerase celular, e pode ser quantificada com um ensaio da atividade de RT sensível, isto é, a RT Cavidi HS-kit Lenti, que se baseia no método descrito por Ekstrand et al. [7], 25 |1I de lisato obtido de acordo com o protocolo corrente é suficiente para a determinação da atividade de RT na amostra. A amostra remanescente de 575 |1I deve ser congelada em -70°C ou abaixo para uso posterior no teste de sensibilidade do medicamento.
[0045] Nota: As enzimas RT que não são sensíveis aos agentes modificadores de cisteína, por exemplo, com RT do HIV 1 tipo silvestre podem opcionalmente ser avaliadas na presença de até 5 mM de 5,5’-ditiobis-(ácido 2-nitrobenzóico). As enzimas sensíveis tais como MULV RT e RT de certas cepas de HIV 1 resistentes a terapia (contendo por exemplo a mutação Y181C) por outro lado requerem a adição de um agente redutor de sulfidrila, isto é, cisteína ou cisteamina, no tampão de lise. MATERIAIS
[0046] Iniciador / Padrão para o ensaio de DNA polimerase.
[0047] A sequência iniciadora é 18 bases 5’-GTC-CCT-GTT-CCG-GCG-CCA-3’ (SEQ ID NO: 12) e ligada na extremidade 5’ a uma amina primária por um braço separador C6.
[0048] A construção do padrão contém três partes com funções diferentes. A partir da extremidade 5’: Um polímero (A)n usado para amplificar o sinal BrdU, uma parte variável (CTGA)m para obter uma reação de polimerase que é dependente dos quatro trifosfatos de deoxinucleosideo e uma sequência complementar ao iniciador.
[0049] Nas experiências incluídas nos exemplos n = 12 e m = 5, senão for de outra maneira mencionada.
[0050] Nucleosídeos, inibidores da enzima e medicamentos antivirais.
[0051] DdATP, trifosfato de dideoxiadenosina; ddGTP, trifosfato de 2’3’ dideoxiguanosina; ddCTP, trifosfato de 2’3’ dideóxi citidina; ddTTP trifosfato de 2’3’ dideóxi timidina.
[0052] TMAU, 6-([3,4-trimetileno]anilino)uracila.
[0053] Tenofovir, (R)-9-(2-fosfonilmetoxipropil)adenina; Nevirapine, (11- ciclopropiI-5,11 -diidro-4-metil-6H-dipirido[3,2-b:2’,3’-f][1,4]diazepin-6-ona) (NVP); e Efavirenz, (-)6-cloro-4-ciclopropiletinil-4-trifluorometi 1-1,4-diidro-2H-3,1 -benzoxazin-2- ona) (EFV). Enzimas
[0054] DNA polimerase I (E. coli) foi adquirido da Amersham Bioscience. DNA polimerase recombinante III de Staphylococus aureus foi produzida como descrito (Brown et al. 1998). DNA polimerase de mamífero oc (glândula vascular de bezerro) e β (humano) foram adquiridos da CHIMERx (Milwaukee). DNA polimerase y foi purificada do coração de boi como descrito (Pileur et al. 2000).
[0055] As formas mutantes resistentes NNRTI de RT do HIV-1 foram produzidas (L100I, K103N, L100I/K103N, Y181C). Como padrão para as mutações foi usado o vetor de expressão pETRT, que foi construído a partir do BH10 isolado. As mutações foram geradas usando kits de mutagênese direcionados ao sítio comercial, QuikChange (Stratagene). As mutações foram verificadas pela análise da sequência de DNA. As formas modificadas e nativas de RT foram isoladas como anteriormente descritas (Lindberg et al. 2002).
[0056] O procedimento para a produção de RTs do HIV-1 recombinantes com mutações específicas de AZT foi similar, mas as mutações foram introduzidas na região de codificação da RT a partir do HXB2-D isolado.
[0057] Amostras de plasma de indivíduos infectados com o HIV.
[0058] As amostras de plasma de pacientes simples tratados com a terapia de combinação costumeira foram selecionadas retrospectivamente. A quantidade de RNA do HIV-1 e cada amostra foi medida por HIV 1 RNA PCR padrão (Cobas, Roche Diagnostica).
[0059] Amostras de soro de pacientes com distúrbios proliferativos da linfa foram obtidas do Department of Internal Medicine, Uppsala Universitym Akademiska sjukhuset, Uppsala.
[0060] Gel de separação: por exemplo, Fractogel® EMD TMAE ou Fractogel® EMD TMAE Hicap em 314 mM de (2-(N-Morfolino)ácido etanossulfônico) MÊS) pH 5,1,413 mM de lodeto de potássio e Heparin 0,5 mg I ml.
[0061] Mini colunas, por exemplo, Biorad Poly-Prep® (7311553)
[0062] Dispositivo de lavagem da mini coluna, isto é, tubo de distribuição a vácuo de extração da fase sólida Supelco Visiprep.
[0063] Tubos plásticos, por exemplo, tubos criogênicos de 4,5 ml Nunc.
[0064] Placas de microtítulo com prA imobilizado, isto é, Nalge Nunc NucleoLinck®.
[0065] Agente modificador de cisteína, por exemplo, 5,5’-ditiobis-(ácido 2- nitrobenzóico) 66 mM em água tamponada com tris(hidroximetila)aminometano 0,87 M (pH 8,3).
[0066] Agente redutor de sulfidrila suave, por exemplo, cisteamina 33 mM em água.
[0067] Tamponantes usados: A) Tamponante de lavagem: 20 mM MES pH 5,4, 500 mM Acetato de potássio (KAc). B) Tamponante condicionador. Um tamponante compatível como o ensaio de RT, por exemplo, (N-(2-hidroxietilpiperazina-N’-(ácido 2- etanossulfônico) 50 mM (Hepes) pH 7,6, KAc 25 mM, cloreto de magnésio (MgCte) 20 mM. Etileno glicol-bis(éter β-aminoetilico) N,N,N’,N’-ácido tetraacético (EGTA) 0,2 mM, espermina 2 mM e albumina de soro bovino inativada por calor (BSA) 0,5 mg / ml. C) Tamponante de lise: Um tamponante compatível com o ensaio de RT incluindo um detergente, por exemplo, Éter Laurílico de Polioxietileno 4 1,25 % (Brij 30), 13 ng / ml de odT22 e os mesmos componentes como no tamponante condicionador (B). Um agente redutor de sulfidrila, isto é, cisteamina 0,2 mM é opcionalmente adicionado quando se processa o vírus com a RT que são sensíveis à oxidação SH / modificação.
[0068] Mistura da reação de RT: (N-(2-hidroxietilpiperazina-N’-(ácido 2- etanossulfônico) (Hepes) 11,7 mM pH 7,6, BrdUTP 28,3 piM, odT22 120 ng/ml, MgCl2 4 mM, sulfato de dextrano 0,05 g/l, espermina 2 mM, Triton-X 100 0,5 % (v/v), Etileno glicol-bis(éter β-aminoetilico) N,N,N’,N’-ácido tetraacético (EGTA) 0,2 mM, e albumina de soro bovino (BSA) 0,5 mg / ml.
[0069] DNA polimerase ye tamponante de base de DNA polimerase retro. Hepes 50 mM pH 8,0, MgCl2 8 mM, sulfato de dextrano 1,5 pig/l, espermina 1 mM, Triton-X 100 0,5 % (v/v), EGTA 0,2 mM, ditiotreitol (DTT) 1,5 mM e albumina de soro bovino (BSA) 0,5 mg/ml.
[0070] Tamponante de base de DNA polimerase III. (2-(N-morfolino)ácido etanossulfônico) (MES) 40 mM pH 6,8, Acetato de potássio (KAc) 40 mM, MgCl2 10 mM, espermina 2 mM, monolaureato de polioxietilenossorbitano (Tween 20) 0,5 % (v/v), EDTA 0,1 mM, ditiotreitol 1 mM e albumina de soro bovino (BSA) 50 pg / ml.
[0071] Tamponante de base de DNA polimerase β. Ácido 3-[(1,1-dimetil-2- hidroxietila)amino]-2-hidróxi-1-propanossulfônico (AMPSO) 20 mM pH 8,3, MgCl2 1 mM, Espermidina 3 mM, BSA1 pg/ml, EDTA 10 pM, DTT 0,1 mM, Tween 20 0,01 %.
[0072] Mistura de reação de DNA polimerase. Tampão de base de DNA polimerase reforçado com BrdUTP 24 ptM, dGTP 49,5 |1M, dATP 49,5 |1M, dCTP 49,5 |1M e 500 padrão ng / ml.
[0073] Mistura da reação de DNA polimerase retro com ATP. A mistura de reação de DNA polimerase ye DNA polimerase retro foi reforçada com ATP 3,2 mM e o pH foi ajustado para 7,1.
EXEMPLOS
[0074] Exemplo 1. Utilização de padrões diferentes para a segunda síntese do filamento por RT do HIV.
[0075] Séries de diluição de duas etapas de formações indicadas de padrão partindo de 200 ng / ml foram incluídas em cada reservatório de placas de microtítulo com iniciador imobilizado de acordo com a “Produção de placas de microtítulo revestidas com iniciador”. 100 fg de RT do HIV 1 recombinante foram adicionados a cada reservatório e a duração da reação de RT foi de 18 horas. A atividade polimerase em cada padrão foi determinada de acordo com o “Protocolo para o ensaio de DNA polimerase” usando tamponante de base DNA polimerase ye DNA polimerase retro. NaCI 50 mM ou alternativamente 100 mM foi usado durante a ligação de anticorpo monoclonal anti BrdU. As atividades observadas foram marcadas em gráfico com respeito à concentração do padrão e o sinal máximo obtido para cada tipo de padrão foi calculado a partir do valor plateau do respectivo gráfico.
[0076] Os resultados são resumidos na Tabela 1. Uma correlação clara foi observada entre o comprimento da cauda A e o sinal máximo obtenível no ensaio de polimerase. O comprimento da cauda A também afetou a capacidade do anticorpo usado para a detecção do produto de se ligar na força iônica aumentada.
[0077] Exemplo 2. Efeitos da sequência padrão na inibição da DNA polimerase III com TMAU
[0078] As 6-anilinouracilas são inibidores seletivos da DNA polimerase III das bactérias gram-positivas. A molécula de anilinouracila inibe seu alvo polimerase III por sequestrá-la dentro de um complexo de DNA-medicamento-proteína inativo (Tarantino et al. 1990). O medicamento TMAU pode ser considerado como um análogo ao GTP. A capacidade inibidora das concentrações indicadas de TMAU para 1,25 ng / reservatório de DNA polimerase III recombinante foi determinada de acordo com o “Protocolo para a determinação de inibição da segunda síntese do filamento no padrão de DNA variável” utilizando ou (CTGA)6-A12 (SEQ ID NO: 10) (■) ou (CTG)6-A3 (SEQ ID NO: 11) (A) como padrão. O tempo de reação da polimerase foi de uma hora e a concentração de GTP na mistura de reação de DNA polimerase III foi reduzida para 2,5 piM. As atividades de polimerase obtidas no padrão indicado foram recalculadas em % das atividades observadas com a mesma polimerase incubada na ausência de inibidor.
[0079] Os resultados são representados na Figura 1. Este inibidor altamente específico apresentou capacidade inibidora variável dependente da sequência da padrão usado. A invenção corrente fornece um sistema onde o padrão de DNA usado pode facilmente ser mudado por considerar as condições específicas requeridas pela enzima ou inibidor estudado.
[0080] Exemplo 3. Sensibilidade de detecção do ensaio de DNA polimerase.
[0081] A atividade de diluições em série de: RT tipo silvestre do HIV 1 (♦), DNA polimerase β (•) e DNA polimerase I de E. coli (A) forma medidas de acordo com o “Protocolo para o ensaio de DNA polimerase” utilizando tamponantes de base otimizados para a enzima indicada. O período de reação da polimerase usado foi durante a noite (18 horas). Os resultados são apresentados na Figura 2. Cada uma das três preparações de enzima mostraram uma ligação entre a quantidade de enzima usada e a quantidade de produto recuperado. A base do ensaio foi 3800 rfu/hora. Utilizando o dobro da base como valor de corte para a detecção de sinal significativa foi possível detectar abaixo de 10 nU de RT tipo silvestre do HIV 1, 6 nU da DNA polimerase β de mamífero e 3 nU da DNA polimerase I de E. coli.
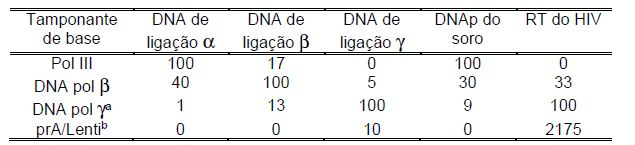
[0082] Exemplo 4. A atividade de cinco DNA polimerases em diferentes sistemas de ensaio.
[0083] A atividade das diluições em série de DNA polimerase oc, DNA polimerase β, DNA polimerase y, soro de um paciente que sofre de linfoma não-Hodgkins e RT do HIV-1 foi medida de acordo com o “Protocolo para o ensaio de DNA polimerase” e “Protocolo para a determinação da atividade de RT” usando soluções de reação com base em tamponantes de base polimerase indicados. 0 tempo de reação de polimerase usado foi 2 horas. As atividades observadas foram recalculadas em % da atividade no padrão de DNA variável nas condições de reação ideais para cada enzima. Os resultados são apresentados na Tabela 2. Cada uma das cinco preparações de enzima investigadas tinha suas preferências individuais a respeito das condições de reação ideais. A DNA polimerase e a polimerase de soro humano, no entanto, mostraram um modelo similar. Somente as RT do HIV-1 e DNA polimerase y deram uma atividade significativa no ensaio da transcriptase reversa.
[0084] Exemplo 5. Demonstração da capacidade do ensaio de DNA polimerase para medir a inibição pelos análogos de dideóxi para todas as quatro bases de DNA.
[0085] A capacidade inibidora de concentrações indicadas de ddATP (□), ddGTT (♦), ddCTP (•) eddTP (A) com respeito à atividade de 80 fg RT do HIV 1 tipo silvestre recombinante foi determinada de acordo com “Protocolo para a determinação de inibição da segunda síntese do filamento sobre o padrão de DNA variável”. A atividade polimerase em cada concentração de inibidor foi recalculada em % da atividade de um controle na ausência de inibidor. Os resultados são representados na Figura 3.
[0086] A reação de polimerase que utiliza o padrão (CTGA)e-Ai2 (SEQ ID NO: 10) foi observada sensível para a inibição pelos análogos de dideóxi para todas as quatro bases de DNA. O valor IC50 observados variaram de 20 nM para ddCTP a 80 nM para ddATP.
[0087] Exemplo 6. Comparação do efeito de inibidores não nucleosídeos na primeira e segunda síntese de DNA do filamento por RT do HIV 1.
[0088] Os efeitos de três inibidores não nucleosídeos sobre a RT do HIV 1 recombinante indicada foram determinados de acordo com “Protocolo para a determinação de inibição da segunda síntese do filamento sobre 0 padrão de DNA variável” usando tamponante de base DNA polimerase y e DNA polimerase retro e “Protocolo para a determinação de inibição da transcrição reversa” respectivamente. A duração das reações de polimerização foram 19 horas para a segunda síntese do filamento em (CTGA)e-Ai2 (SEQ ID NO: 10) e 2 horas para a primeira síntese do filamento em prA respectivamente. As atividade de RT obtidas foram recalculadas em % das atividades observadas com a mesma RT incubada na ausência de inibidor.
[0089] Os resultados são resumidos na Tabela 3. Ambos os sistemas de ensaio tinham a capacidade para distinguir entre as enzimas RT resistentes (Y181C, V179D) e sensíveis. Os valores IC50 para qualquer um dos inibidores não foram significativamente afetados de uma diferença de cinco vezes na quantidade de enzima usada em qualquer um dos dois sistemas de ensaio. Além disso não existiu nenhuma diferença significativa entre os valores IC50 obtidos mediante a medição da inibição da primeira ou segunda síntese de DNA do filamento.
[0090] Exemplo 7. Demonstração da resistência fundamental do mecanismo bioquímico no medicamento antiviral tenofovir.
[0091] A terapia prolongada de indivíduos infectados com o HIV com análogos de nucleosídeo leva ao desenvolvimento de vírus resistente. Este processo é associado com o aparecimento gradual de mutações no gene de ligação virai. Os efeitos destas substituições nos níveis enzimáticos são complicados e incluem a intensificação de uma função de montagem do DNA primitivo. Esta reação é dependente do nucleotídeo e produz polifosfato de dinucleosídeo e uma extremidade 3 ‘ do DNA prolongável.
[0092] Os efeitos de diluições seriais de trifosfato tenofovir em 4 pg/reservatório de RT do HIV 1 recombinante indicada foram determinados de acordo com o “Protocolo para a determinação de inibição da segunda síntese do filamento sobre o padrão de DNA variável” utilizando solução de reação padrão com base em tamponante de base DNA polimerase y e DNA polimerase retro e a mesma solução de reação suplementada com ATP. A duração das reações de polimerização foi de 19 horas. As atividades de RT obtidas foram recalculadas em % da atividade da mesma RT incubada na ausência de inibidor. A Figura 4 apresenta os resultados.
[0093] Símbolos: RT tipo silvestre do HIV-1 em solução de reação de DNA polimerase padrão (O). RT tipo silvestre do HIV-1 em solução de reação de DNA polimerase com ATP (•), RT mutante do HIV-1 T69S-^SS/L210W/T215Y em solução de reação de DNA polimerase padrão (0), RT mutante do HIV-1 T69S^SS/L210W/T215Y em solução de reação de DNA polimerase com ATP (♦).
[0094] Os resultados representados na figura 4 demonstram que a diferença na suscetibilidade do medicamento entre a RT tipo silvestre e mutante aumentou aproximadamente 10 vezes quando uma solução de reação com capacidade de suportar uma reação de fosforólise dependente do ATP foi usada.
[0095] Exemplo 8. Determinação da suscetibilidade a Nevirapine usando RT derivada do plasma.
[0096] Amostras de um ml de plasma de 3 indivíduos infectados por HIV da Stockholm, Sweden foram processadas de acordo com o “Protocolo para o isolamento da RT virai a partir do material que contém anticorpos bloqueadores da RT, com base na destruição das enzimas celulares solúveis seguido pelo isolamento da RT virai das mini colunas”. Cada RT do plasma e duas enzimas de controle foram tituladas com respeito a um conjunto de diluições em série de Nevirapine de acordo com o “Protocolo para a determinação de inibição da segunda síntese do filamento sobre o padrão de DNA variável” usando “tamponante de base DNA polimerase y e DNA polimerase retro”. Ver a Figura 5.
[0097] Símbolos: (□) RT do paciente 1 tendo 140.000 cópias de genoma /ml, (□) RT do paciente 2 tendo 180.000 cópias de genoma /ml, (A) RT do paciente 3 tendo 390.000 cópias de genoma /ml, (•) um controle consistindo de RT tipo silvestre do HIV-1 recombinante, (♦) e um controle consistindo do RT mutante do HIV-1 recombinante (L100I), com resistência a Nevirapine intermediária.
[0098] Os valores de IC50 com respeito à Nevirapine observados para as RTs do paciente variaram de 0,7 a 1,2 piM. Para ser comparado com 0,5 |1M para a RT tipo silvestre de controle e > 10 |1M para a RT mutante com resistência a Nevirapine intermediária (L100I).
[0099] Exemplo 9. Detecção de uma atividade de DNA polimerases no soro de pacientes com distúrbios linfoproliferativos.
[0100] O soro de quatro pacientes que sofrem de linfoma não-Hodgkins e de seis doadores de sangue saudáveis foram diluídos em série com tampão de base DNA polimerase III. A atividade de polimerase em cada etapa de diluição foi medida de acordo com o “Protocolo para o ensaio de DNA polimerase” usando duas a seis horas de tempo de reação da polimerase.
[0101] A atividade de DNA polimerase / |1I de amostra de soro e o tempo do ensaio de polimerase foram calculados na faixa de diluição onde existiu uma ligação linear entre a quantidade de produto formado e a quantidade de plasma adicionado ao ensaio (ver a tabela 4).
[0102] Cada amostra de soro dos pacientes com linfoma não-Hodgkins continha quantidades individuais de atividade de DNA polimerase, com propriedades similares como DNA polimerase oc (ver a tabela 3). A quantidade de atividade variou de aproximadamente 2 a 190 vezes o valor médio observado entre os doadores de sangue saudáveis. TABELA 1. Utilização de padrões diferentes para a segunda síntese do filamento por RT do HIV.
Séries de diluição de duas etapas de padrão indicada iniciada de 200 ng / ml foram incluídas em cada reservatório de placas de microtítulo com iniciador imobilizado. 100 fg de RT do HIV 1 recombinante foram adicionados a cada reservatório e a duração da reação de RT foi de 18 horas. 50 mM foi usado durante a ligação de anticorpo monoclonal anti BrdU. As atividades observadas foram marcadas em gráfico com respeito à concentração do padrão e o sinal máximo obtido para cada padrão foi calculado.
[0103] a Condições e cálculos de reação de RT foram idênticos como no “sinal máximo”, mas NaC1100 mN foi usado durante a ligação de anticorpo. TABELA 2. Atividade aparente de quatro DNA polimerases de mamífero e RT do HIV em diferentes sistemas de ensaio. Atividade da isoenzima DNA polimerase no ensaio indicado (% *)
• % de atividade no padrão de DNA variável com tamponante de base ideal para a isoenzima indicada
[0104] a Tamponante de base DNA Polimerase ye DNA polimerase retro.
[0105] b Atividade no ensaio de RT com prA / odT como padrão iniciador. TABELA 3. Comparação do efeito de inibidores não nucleosídeos na primeira e segunda síntese de DNA do filamento por RT do HIV 1.
• Os efeitos de três inibidores não nucleosídeos sobre a RT do HIV 1 recombinante indicada foram determinados de acordo com “Protocolo para a determinação de inibição da segunda síntese do filamento sobre o padrão de DNA variável” e “Protocolo para a determinação de inibição da transcrição reversa” respectivamente. A duração das reações de polimerização foram 19 horas para a segunda síntese do filamento em (CTGA)e-Ai2 (SEQ ID NO: 10) e 2 horas para a primeira síntese do filamento em prA respectivamente. As atividade de RT obtidas foram recalculadas em % das atividades observadas com a mesma RT incubada na ausência de inibidor. TABELA 4. Detecção de uma atividade de DNA polimerase no soro de pacientes com distúrbios linfoproliferativos.
A Paciente sofrendo de linfoma não-Hodgkins • NS, não significativo.
[0106] O soro de quatro pacientes que sofrem de linfoma não-Hodgkins e de seis doadores de sangue saudáveis foram diluídos em série com tampão de base DNA polimerase III. A atividade de polimerase em cada etapa de diluição foi medida de acordo com o “Protocolo para o ensaio de DNA polimerase”. As atividades de DNA polimerase classificadas foram calculadas a partir da faixa de diluição onde existiu uma ligação linear entre a quantidade de produto formado e a quantidade de plasma adicionado. A base do ensaio de 854 flu/h foi subtraída de todos os valores.
REFERÊNCIAS
[0107] Aposhian HV, Kornberg A. J. Biol. Chem. 1962, 237, 519.
[0108] Arion D, Kaushik N, McCormick S, Borlow G, Parniak MA. Phenotypic mechanism of HIV-1 resistance to 3’-azido-3’-deoxythymidine (AZT): increased polimerization processivity and enhanced sensitivity to pyrophosphate of the mutant viral reverse transcriptase. Biochemistry 10 de novembro de 1998; 37(45): 15908-17.
[0109] Barnes MH, Leo CJ, Brown NC. DNA polymerase III of Gram-positive eubacteria is a zinc metalloprotein conserving an essential finger-like domain. Biochemistry. 03 de novembro de 1998; 37 (44): 15254-60.
[0110] Eberle J, Seibl R, Kessler, C, Konig, Bernhard B. Reagents and kits for determining polymerase activity. US5635350 1997.
[0111] Ekstrand DH, Awad RJ, Kallander CF, Gronowitz JS. A sensitive assau for the quantification of reverse transcriptase activity based on the use of carrier-bound template and non-radioactive-product detection, with special reference to human- immunodeficiency-virus isolation. Biotechnol Appl Biochem. Abril de 1996; 23 (Pt 2): 95-105.
[0112] Goff, S.P. (1990) Retrovirus reverse transcriptase: Synthesis, Structure and Function. Review. J Acquir Imm Defic Syndr 3: 817-831.
[0113] Gronowitz; J S, Kallander, C F R Method of determining dTk isoenzyme activity and the use thereof. US4637977 1987.
[0114] Meyer PR, Matsuura SE, Mian AM, So AG, Scott WA. Related Articles. A mechanism of AZT resistance: an increase in nucleotide-dependent primer unblocking by mutant HIV-1 reverse transcriptase. Mol Cell. Julho de 1999; 4(1): 35-43.
[0115] Pileur F, Toulme JJ, Cazenave C. Eukaryotic ribonucleases HI and Hll generate characteristic Hydrolytic patterns on DNA-RNA hybrids: further evidence that mitochondrial RNase H is an RNase Hll. Nucleic Acids Res. 15 de setembro de 2000; 28(18: 3674-83).
[0116] Seville M, West AB, Cull MG, McHenry CS. Fluorometric assay for DNA polymerases and reverse transcriptase. Biotechniques. Outubro de 1996; 21 (4):, 664, 666, 668, 670, 672.
[0117] Tarantino PM Jr, Zhi C, Wright GE, Brown NC. Related Inhibitors of DNA polymerase III as novel antimicrobial agents against gram-positive eubacteria. Antimicrob Agents Chemother. Agosto de 1999; 43(8): 1982-7.
[0118] Tveit H, Kristensen T. Fluorescence-based DNA polymerase assay. Anal Biochem. 01 de fevereiro de 2001; 289 (1): 96-8.
[0119] Vamdamme AM, Van Vaereπbergh K, De Clercq E. Aπti-humaπ immunodeficiency virus drug combination strategies. Antivir Chem Chemother. Maio de 1998; 9 (3): 187-203.