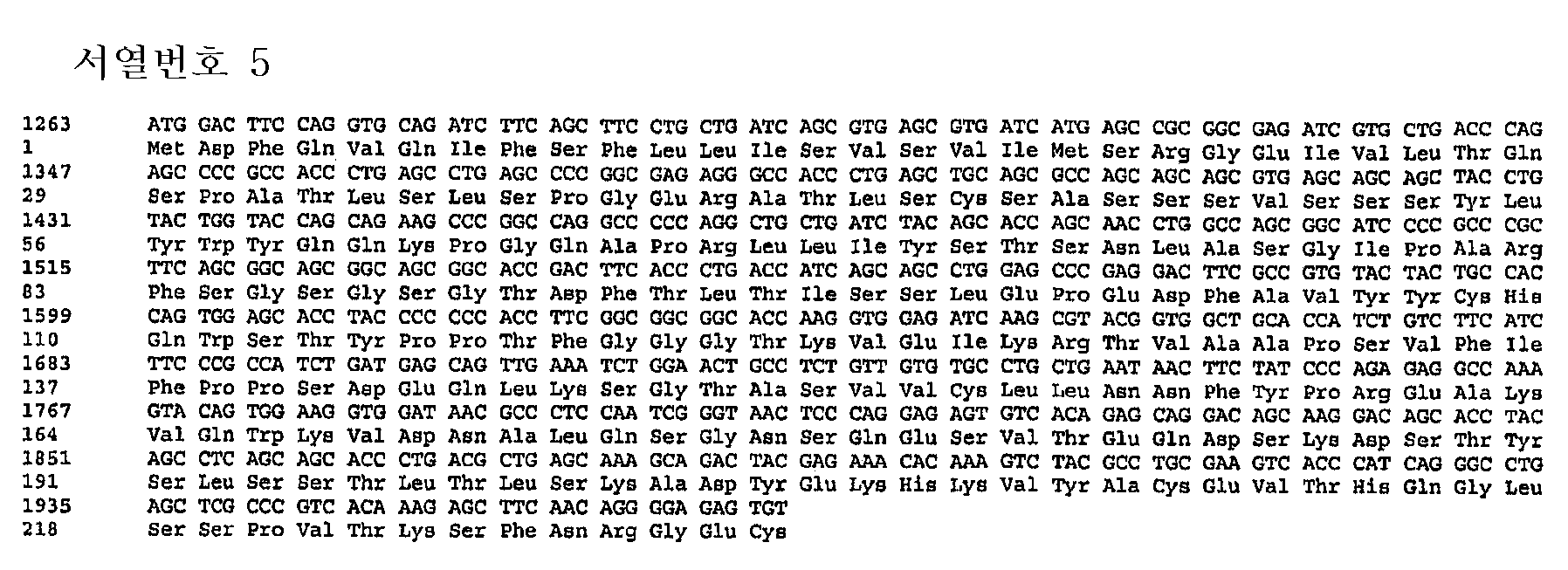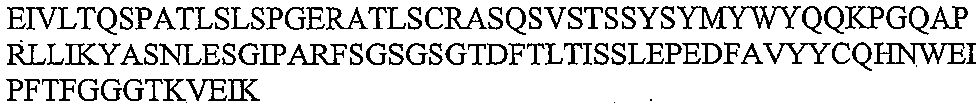본 명세서에서 달리 정의하지 않는 한, 본 명세서에서 사용된 모든 기술적 용어 및 과학적 용어는 본 발명이 속하는 분야에서 통상의 기술을 가진 자에 의해 통상적으로 이해되는 바와 동일한 의미를 가진다. 본 명세서에 기재된 방법 및 재료와 유사한 또는 동등한 임의의 방법 및 재료가 본 발명의 실시 또는 시험에 사용될 수 있지만, 바람직한 방법 및 재료는 이하에 기재될 것이다. 본 명세서에 기재된 방법 및 재료와 유사한 또는 동등한 방법 및 재료가 본 발명의 실시에 사용될 수도 있지만, 예시적 방법 및 재료가 하기에 기재된다. 본 명세서에서 언급된 모든 공개문헌 및 다른 참고문헌은 참고로 완전히 도입된다. 모순이 있는 경우, 정의를 포함하는 본 명세서가 조절할 것이다. 재료, 방법 및 실시예는 단지 예시적인 것이지 제한하기 위한 것이 아니다. 본 명세서 전체를 통해, 용어 "포함한다" 또는 이의 변형 용어 예컨대, "포함하며" 또는 "포함하는"은 명시된 정수 또는 정수 군의 포함을 내포하지만 임의의 다른 정수 또는 정수 군을 배제하지 않는 것으로 이해될 것이다.
정의
약: 임의의 수치적 값을 언급할 때 본 명세서에서 사용된 바와 같이, 용어 "약"은 명시된 값의 ±10%의 값을 의미한다(예를 들면, "약 50℃"는 45℃ 내지 55℃의 온도 범위를 포함하며; 유사하게, "약 100 mM"은 90 mM 내지 110 mM의 농도 범위를 포함함).
길항제: 본 명세서에서 사용된 바와 같이, 용어 "길항제"는 세포, 조직 또는 유기체 내의 ανβ6 인테그린의 생물학적 및/또는 생리학적 효과를 감소시키거나, 실질적으로 감소시키거나 완전히 억제하는 화합물, 분자, 잔기(moiety) 또는 복합체를 지칭한다. ανβ6에 대한 리간드일 수 있는 길항제는 하기 방법을 포함하나 이들로 한정되지 않는 다양한 방법으로 이러한 효과를 나타낼 수 있다: 세포 표면 상의 ανβ6에의 결합에 대해 또 다른 리간드와 경쟁하는 방법; 인테그린이 다른 리간드와 결합하는 능력을 감소시키거나, 실질적으로 감소시키거나 억제하는 방식으로 ανβ6과 상호작용하는 방법; 인테그린이 다른 리간드가 더 이상 결합할 수 없는 구조 (또는 감소된 또는 실질적으로 감소된 친화성 및/또는 효율로만 결합할 수 있는 구조)를 취하도록 세포 표면 ανβ6에 결합하여 ανβ6의 구조적 변화를 유도하는 방법; 다른 리간드의 결합, 또는 세포 상의 ανβ6에 결합할 때 상기 리간드에 의해 유도된 생리학적 신호가 감소되거나, 실질적으로 감소되거나 완전히 억제되도록 세포, 조직 또는 유기체 내에서의 생리학적 변화(예컨대, 세포내 신호전달 복합체에서의 증가; 전사 억제제에서의 증가; 세포 표면 ανβ6 발현에서의 감소 등)를 유도하는 방법; 및 길항제가 그의 활성을 수행할 수 있는 당업자에게 익숙한 다른 기작. 당업자가 이해하고 있는 바와 같이, 길항제는 그가 길항할 수 있거나(예컨대, 길항제가 작용제의 뮤테인(mutein), 변이체, 단편 또는 유도체일 수 있음), 전체적으로 관련없는 구조를 가질 수 있는 또 다른 ανβ6-결합 잔기(예컨대, ανβ6-결합 리간드)와 유사한 구조를 가질 수 있다.
결합된: 본 명세서에서 사용된 바와 같이, 용어 "결합된"은 예를 들면, 화학적 커플링에 의한 공유결합, 또는 비-공유결합 예컨대, 이온성 상호작용, 소수성 상호작용, 수소결합 등일 수 있는 결합 또는 부착을 지칭한다. 공유결합은 예를 들면, 에스테르, 에테르, 포스포에스테르, 티오에스테르, 티오에테르, 우레탄, 아미드, 아민, 펩티드, 이미드, 히드라존, 히드라지드, 탄소-황 결합, 탄소-인 결합 등일 수 있다. 용어 "결합된"은 "커플링된", "접합된" 및 "부착된"과 같은 용어들보다 더 넓고 상기 용어들을 포함한다.
접합체/접합: 본 명세서에서 사용된 바와 같이, "접합체"는 부분 예컨대, 화학적 잔기 또는 방사성 동위원소가 ανβ6에 결합하는 리간드 예컨대, ανβ6-결합 항체 또는 이의 단편에 공유부착되어 형성된 생성물을 지칭한다. "접합"은 상기 문장에서 정의된 바와 같은 접합체의 형성을 지칭한다. 화학물질 또는 방사성 동위원소와 생물학적 활성 물질 예컨대, 단백질 또는 폴리펩티드(항체를 포함함)의 접합 방법으로서 당업자에 의해 통상적으로 이용되는 임의의 방법이 본 발명에서 사용될 수 있다.
질환, 장애, 병태(condition): 본 명세서에서 사용된 바와 같이, 용어 "질환" 또는 "장애"는 종양, 암, 알레르기, 마약중독, 자가면역, 감염, 중독 또는 최적의 정신적 또는 신체적 기능의 손상을 포함하는 인간 또는 동물의 임의의 불리한 병태를 지칭한다. 본 명세서에서 사용된 바와 같이 "병태"는 질환 및 장애를 포함할 뿐만 아니라 생리학적 상태를 지칭한다. 예를 들면, 생식능력은 생리학적 상태이지만 질환 또는 장애가 아니다. 그러므로, 생식능력을 감소시켜 임신을 방지하기에 적합한 본 발명의 조성물은 병태(생식능력)의 치료로서 기재되나, 장애 또는 질환의 치료로서 기재되지 않는다. 당업자라면 다른 병태를 인지할 것이다.
유효량: 본 명세서에서 사용된 바와 같이, 용어 "유효량"은 원하는 생물학적 효과를 달성하기 위해 필요한 또는 충분한 소정의 화합물, 접합체 또는 조성물의 양을 지칭한다. 본 발명의 방법에 따른 소정의 화합물, 접합체 또는 조성물의 유효량은 이러한 선택된 결과를 달성하는 양일 것이며, 이러한 양은 과도한 실험을 필요로 하지 않고 당분야에 공지되고/되거나 본 명세서에 기재된 분석법을 이용하여 당업자에 의해 통상적으로 결정될 수 있다. 예를 들면, 암 전이를 치료하거나 예방하기 위한 유효량은 생체 내에서 기저막(basement membrane) 또는 상피층을 통과하는 종양 세포의 전이 및 침입을 방지하기 위해 필요한 양일 수 있다. 또한, 상기 용어는 "필요량"과 동의어이다. 임의의 구체적인 적용을 위한 유효량은 치료될 질환, 장애 또는 병태, 투여될 구체적인 조성물, 투여 경로, 대상체의 크기, 및/또는 질환 또는 병태의 심각도와 같은 인자에 따라 다를 수 있다. 당업자는 과도한 실험을 필요로 하지 않고 본 명세서에 기재된 지침에 따라 본 발명의 구체적인 화합물, 접합체 또는 조성물의 유효량을 실험적으로 결정할 수 있다.
하나: 용어 "하나"가 본 개시내용에서 사용될 때, 이는 달리 명시하지 않는 한 "적어도 하나" 또는 "하나 이상"을 의미한다. 따라서, 용어 "하나", "하나 이상" 및 "적어도 하나"는 본 명세서에서 상호교환적으로 사용될 수 있다.
펩티드, 폴리펩티드, 단백질: 본 명세서에서 사용된 바와 같이, 용어 "폴리펩티드"는 단수 형태의 "폴리펩티드" 뿐만 아니라 복수 형태의 "폴리펩티드들"을 포함하는 것이고, 아미드 결합(펩티드 결합으로서도 공지되어 있음)에 의해 선형으로 연결된 단량체들(아미노산들)로 구성된 분자를 지칭한다. 용어 "폴리펩티드"는 2개 이상의 아미노산으로 구성된 임의의 쇄 또는 쇄들을 지칭하며, 생성물의 특정한 길이를 지칭하는 것이 아니다. 따라서, 펩티드, 디펩티드, 트리펩티드, 올리고펩티드, "단백질", "아미노산 쇄", 또는 2개 이상의 아미노산들로 구성된 쇄 또는 쇄들을 지칭하기 위해 사용된 임의의 다른 용어는 "폴리펩티드"의 정의 내에 포함되며, 용어 "폴리펩티드"는 상기 용어들 중 어느 한 용어 대신에 사용될 수 있거나 상기 용어들 중 어느 한 용어와 상호교환적으로 사용될 수 있다. 또한, 용어 "폴리펩티드"는 글리코실화, 아세틸화, 인산화, 아미드화, 공지된 보호/차단 기의 유도체화, 단백질분해적 절단, 또는 비-천연 발생 아미노산에 의한 변형을 포함하나 이들로 한정되지 않는 폴리펩티드의 발현 후 변형의 생성물을 지칭하는 것이다. 폴리펩티드는 천연 생물학적 공급원으로부터 유래될 수 있거나 재조합 기술에 의해 생성될 수 있지만, 반드시 지정된 핵산 서열로부터 번역될 필요는 없다. 폴리펩티드는 화학적 합성을 비롯한 임의의 방법으로 생성될 수 있다. 상기 정의에 따라, 본 발명에서 사용되는 폴리펩티드의 크기는 약 3개 이상, 5개 이상, 10개 이상, 20개 이상, 25개 이상, 50개 이상, 75개 이상, 100개 이상, 200개 이상, 500개 이상, 1,000개 이상 또는 2,000개 이상의 아미노산일 수 있다. 폴리펩티드는 정해진 3-차원적 구조를 가질 수 있지만 반드시 이러한 구조를 가질 필요는 없다. 정해진 3-차원적 구조를 가진 폴리펩티드는 폴딩된 폴리펩티드로서 지칭되며, 정해진 3-차원적 구조를 가지는 것이 아니라 상당한 수의 다양한 구조를 채용할 수 있는 폴리펩티드는 폴딩되지 않은 폴리펩티드로서 지칭된다. 본 명세서에서 사용된 바와 같이, 용어 당단백질은 아미노산 잔기 예컨대, 세린 잔기 또는 아스파라진 잔기의 산소-함유 또는 질소-함유 측쇄를 통해 단백질에 부착되어 있는 하나 이상의 탄수화물 잔기에 커플링된 단백질을 지칭한다. 본 발명에 따라 사용되는 바람직한 폴리펩티드는 ανβ6 상의 하나 이상의 에피토프를 인식하여 이 에피토프에 결합하는 항체(특히, 단일클론 항체)를 포함하나 이로 한정되지 않는, 리간드인 폴리펩티드 또는 세포의 표면 상의 ανβ6 인테그린에 결합하는 폴리펩티드를 포함한다.
"단리된" 폴리펩티드, 또는 이의 단편, 변이체 또는 유도체는 그의 천연 환경에서 존재하지 않는 폴리펩티드를 의미한다. 구체적인 정제 수준이 전혀 요구되지 않는다. 예를 들면, 단리된 폴리펩티드는 그의 천연 또는 자연 환경으로부터 제거될 수 있다. 재조합 생성된 폴리펩티드 및 숙주 세포에서 발현된 단백질은 임의의 적절한 기법에 의해 분리되거나, 분획화되거나, 부분적으로 또는 실질적으로 정제된 천연 또는 재조합 폴리펩티드와 마찬가지로 본 발명의 목적을 위해 단리된 것으로 간주된다.
또한, 본 발명의 폴리펩티드에는 상기 폴리펩티드들의 단편, 유도체, 유사체 또는 변이체 및 이들의 임의의 조합물이 포함된다. 항-ανβ6 항체 또는 항체 폴리펩티드를 지칭할 때 용어 "단편", "변이체", "유도체" 및 "유사체"는 상응하는 천연 항체 또는 폴리펩티드의 항원-결합 성질의 적어도 일부를 보유하는 임의의 폴리펩티드, 즉 ανβ6 인테그린 상의 하나 이상의 에피토프에 결합하는 능력을 보유하는 폴리펩티드를 포함한다. 본 발명의 폴리펩티드의 단편은 본 명세서의 다른 곳에 기재된 특정한 항체 단편 이외에 단백질분해에 의해 생성된 단편 뿐만 아니라 결실 단편을 포함한다. 본 발명에 따라 사용할 수 있는 항-ανβ6 항체 및 항체 폴리펩티드의 변이체는 전술한 단편 뿐만 아니라, 아미노산 치환, 결실 또는 삽입으로 인해 변경된 아미노산 서열을 가진 폴리펩티드도 포함한다. 변이체는 천연적으로 발생할 수 있거나 비-천연적으로 발생할 수 있다. 비-천연 발생 변이체는 당분야에 공지된 돌연변이유발 기법을 이용하여 생성할 수 있다. 변이체 폴리펩티드는 보존적 또는 비-보존적 아미노산 치환, 결실 또는 부가를 포함할 수 있다. 본 발명에 따라 사용할 수 있는 항-ανβ6 항체 또는 항체 폴리펩티드의 유도체는 천연 폴리펩티드에서는 관찰되지 않는 추가 특징을 나타내도록 변경된 폴리펩티드이다. 그 예에는 융합 단백질이 포함된다. 변이체 폴리펩티드는 본 명세서에서 "폴리펩티드 유사체"로도 지칭될 수 있다. 본 명세서에서 사용된 바와 같이, 항-ανβ6 항체 또는 항체 폴리펩티드의 "유도체"는 기능적 측 기(side group)의 반응에 의해 화학적으로 유도된 하나 이상의 잔기를 가진 대상 폴리펩티드를 지칭한다. 또한, "유도체"에는 20종의 표준 아미노산의 하나 이상의 천연 발생 아미노산 유도체를 함유하는 펩티드가 포함된다. 예를 들면, 4-히드록시프롤린은 프롤린을 치환시킬 수 있고; 5-히드록시라이신은 라이신을 치환시킬 수 있으며; 3-메틸히스티딘은 히스티딘을 치환시킬 수 있고; 호모세린은 세린을 치환시킬 수 있으며; 오르니틴은 라이신을 치환시킬 수 있다.
실질적으로, 실질적인: 본 명세서에서 사용된 바와 같이, 단백질의 접합은 접합된 단백질과 수용체의 결합 속도 및/또는 양이 접합되지 않은 상응하는 사이토카인, 케모카인, 성장 인자 또는 폴리펩티드 호르몬의 결합 속도 및/또는 양의 약 40%, 약 50%, 약 60%, 약 65%, 약 70%, 약 75%, 약 80%, 약 85%, 약 90%, 약 91%, 약 92%, 약 93%, 약 94%, 약 95%, 약 96%, 약 97%, 약 98%, 약 99% 또는 약 100% 이상이면 상기 단백질이 그의 수용체(들)에 결합하는 능력을 "실질적으로" 방해하지 않는다고 말한다.
치료: 본 명세서에서 사용된 바와 같이, 용어 "치료", "치료하다", "치료된" 또는 "치료하는"은 특히, 원하는 않는 생리학적 변화 또는 장애 예컨대, 다발성 경화증의 진행을 예방하거나 늦추는(경감시키는) 것이 목적인 예방 및/또는 치료를 지칭한다. 유익한 또는 원하는 임상 결과에는 증상의 완화, 질환 정도의 경감, 질환의 안정화된(즉, 악화되지 않은) 상태, 질환 진행의 지연 또는 늦춰짐, 질환 상태의 개선 또는 완화, 및 검출가능한 또는 검출불가능한 (부분적 또는 전체적) 경감이 포함되나 이들로 한정되지 않는다. "치료"는 치료를 받지 않은 경우 기대되는 생존에 비해 연장된 생존을 의미할 수도 있다. 치료가 필요한 대상체에는 병태 또는 장애를 이미 가지고 있는 대상체뿐만 아니라 병태 또는 장애를 가지기 쉬운 대상체 또는 병태 또는 장애가 예방되어야 하는 대상체가 포함된다. "대상체" 또는 "개체" 또는 "동물" 또는 "환자" 또는 "포유동물"은 진단, 예후 또는 치료가 필요한 임의의 대상체, 특히 포유동물 대상체를 의미한다. 포유동물 대상체에는 인간 및 다른 영장류, 가축, 농장 동물, 동물원 동물, 스포츠용 동물 또는 애완 동물, 예컨대, 개, 고양이, 기니아 피그, 토끼, 래트, 마우스, 말, 소, 젖소 등이 포함된다.
개요
본 발명은 인테그린 ανβ6에 대해 특이적인 인간화된 항체를 특징으로 한다. 본 발명의 항체를 제조하는 다양한 방법이 본 명세서에 기재되어 있다. 당분야에서 공지되어 있으나 본 명세서에서 구체적으로 기재되어 있지 않은 방법도 본 발명의 범위 내에 있다.
적어도 부분적으로, 본 발명은 인테그린 ανβ6이 비-전이성 종양 세포 또는 낮은 전이력을 가진 종양 세포 상에서 관찰된 발현 수준에 비해 전이성 또는 더 높은 전이력을 가지는 종양 세포 상에서 증가된 양으로 발현된다는 점에서 상기 인테그린 ανβ6이 종양 세포의 표면 상에 차등적으로 발현된다는 발견에 기초한 것이다. 이러한 차등적 발현을 분석하기 위해, 본 발명은 인테그린 ανβ6에 결합하는 리간드, 특히 항체(보다 구체적으로, 본 발명에 의해 제공되는 인간화 항체)를 사용한다. 다른 실시양태에서, 본 발명은 종양 세포의 침입력 및/또는 전이력을 측정하고 침입성 또는 전이성 암종으로 진행할 가능성이 보다 더 높은 특정한 선암종 및 상피내 암종(DCIS 및 LCIS를 포함함)과 같은 암종을 확인하는 데 있어서 이러한 차등적 발현의 확인을 이용하는 방법도 제공한다. 또한, 본 발명은 인테그린 ανβ6에 결합하는 하나 이상의 리간드를 사용한 치료에 반응할 가능성이 보다 더 높은 종양을 구성하는 세포가 있는 종양을 확인하는 방법을 제공한다. 또한, 본 발명은 종양 전이의 진단 및 치료/예방 방법, 및 종양의 수술적 제거 후 잔류 전이성 종양 세포의 제거 방법을 제공한다.
인간화 항체
한 실시양태에서, 본 발명에 의해 제공되는 항체는 단일클론 항체이며, 바람직한 실시양태에서, 다른 종으로부터 유래된 동족 항-ανβ6 항체의 인간화 버젼이다. 인간화 항체는 항원 결합에 필요하지 않은 인간 면역글로불린 중쇄 또는 경쇄의 아미노산들의 일부 또는 전부(예를 들면, 가변 도메인의 불변 영역 및 골격 영역)를 사용하여 동족 비-인간 항체의 경쇄 또는 중쇄의 상응하는 아미노산을 치환시키는 재조합 DNA 기술에 의해 생성된 항체이다. 예를 들면, 소정의 항원에 대한 뮤린 항체의 인간화 버젼은 그의 중쇄 및 경쇄 둘다에서 (1) 인간 항체의 불변 영역; (2) 인간 항체의 가변 도메인의 골격 영역; 및 (3) 뮤린 항체로부터의 CDR을 가진다. 필요한 경우, 인간 골격 영역 내의 하나 이상의 잔기가 뮤린 항체 내의 상응하는 위치에 있는 잔기로 변경되어 항원에 대한 인간화 항체의 결합 친화성을 보존할 수 있다. 이 변화는 종종 "복귀(back) 돌연변이"로 불린다. 인간화 항체는 일반적으로 키메라 인간 항체에 비해 인간에서 면역 반응을 일으킬 가능성이 더 낮은데, 이는 인간화 항체가 상당히 적은 비-인간 구성요소들을 함유하기 때문이다.
본 발명의 인간화 항체의 제조에 적합한 방법은 예를 들면, 윈터(Winter)의유럽 특허 제0 239 400호; 문헌[Jones et al., Nature 321: 522-525 (1986); Riechmann et al., Nature 332: 323-327 (1988); Verhoeyen et al., Science 239: 1534-1536 (1988); Queen et al., Proc. Nat. Acad. Sci.. USA 86: 10029 (1989)]; 미국 특허 제6,180,370호; 및 문헌[Orlandi et al., Proc. Natl. Acad. Sci. USA 86: 3833 (1989)]에 기재되어 있고, 이들의 개시내용은 본 명세서에 참고로 완전히 도입된다. 일반적으로, 뮤린 (또는 다른 비-인간) CDR을 인간 항체로 이식하는 것은 다음과 같이 달성된다. 중쇄 및 경쇄 가변 도메인을 코딩하는 cDNA를 하이브리도마로부터 단리한다. CDR을 비롯한 가변 도메인의 DNA 서열을 시퀀싱으로 결정한다. CDR을 코딩하는 DNA를 부위 지정 돌연변이유발을 이용하여 인간 항체 중쇄 또는 경쇄 가변 도메인 코딩 서열의 상응하는 영역으로 옮긴다. 원하는 이소타입(예컨대, CH에 대해 γ1 및 CL에 대해 κ)의 인간 불변 영역 유전자 절편을 부가한다. 인간화 중쇄 및 경쇄 유전자를 포유동물 숙주 세포(예컨대, CHO 또는 NSO 세포)에서 동시에 발현시켜 가용성 인간화 항체를 생성한다. 항체의 대규모 생성을 용이하게 하기 위해, 항체-발현 세포를 함유하는 생물반응기에서 상기 인간화 항체를 제조하거나, 젖 중에 항체를 발현하는 형질전환 동물(예컨대, 염소, 소 또는 양)을 생성하는 것이 종종 바람직하다(예컨대, 미국 특허 제5,827,690호를 참조함).
종종, 인간 골격으로의 CDR의 직접적인 전달은 생성된 항체의 항원-결합 친화성을 상실시킨다. 이는 일부 동족 항체에서 골격 영역 내의 특정 아미노산이 CDR과 상호작용하여 항체의 전체 항원-결합 친화성에 영향을 주기 때문이다. 이 경우, 동족 항체의 항원-결합 활성을 보유하도록 수용 항체의 골격 영역 내에 "복귀 돌연변이"(상기)를 도입시키는 것이 중요하다.
복귀 돌연변이를 유발하는 일반적인 방법은 당분야에 공지되어 있다. 예를 들면, 문헌[Queen et al., (supra), Co et al., Proc. Nat. Acad. Sci. USA 88: 2869-2873 (1991)] 및 국제특허출원 공개 제WO 90/07861호(Protein Design Labs Inc.)에는 2개의 핵심 단계를 포함하는 방법이 기재되어 있다. 첫째, 동족 뮤린 항체의 가변 영역 골격과 상동성을 가진 최적 단백질 서열에 대한 컴퓨터 분석으로 인간 가변 골격 영역을 선택한다. 그 후, 뮤린 CDR과 상호작용할 가능성이 있는 골격 아미노산 잔기들을 가시화하기 위해 뮤린 가변 영역의 3차 구조를 컴퓨터로 모델링한 후, 이 뮤린 아미노산 잔기들을 상동성을 가진 인간 골격 위에 겹쳐 놓는다.
이 2-단계 방법에서, 인간화 항체를 디자인하기 위한 여러 기준이 있다. 제1 기준은 비-인간 공여 면역글로불린과 일반적인 상동성을 가진 특정한 인간 면역글로불린으로부터의 골격을 인간 수용 골격으로서 사용하거나, 많은 인간 항체로부터 유래된 컨센서스(consensus) 골격을 사용하는 것이다. 제2 기준은 인간 수용 잔기가 일반적인 잔기가 아니고 공여 잔기가 골격의 특정한 잔기에서 인간 서열에 대해 전형적일 경우 수용 아미노산보다 오히려 공여 아미노산을 사용하는 것이다. 제3 기준은 CDR에 바로 인접한 위치에서 수용 골격 아미노산 잔기보다 오히려 공여 골격 아미노산 잔기를 사용하는 것이다.
예를 들면, 문헌[Tempest, Biotechnology 9: 266-271 (1991)]에 기재된 다른 방법을 이용할 수도 있다. 이 방법에서, NEWM 및 REI 중쇄 및 경쇄 각각으로부터 유래된 가변 영역 골격을 마우스 잔기의 근치적 도입 없이 CDR-이식에 사용한다. 이 방법을 사용함으로써 얻는 이점은 NEWM 및 REI 가변 영역의 3-차원적 구조가 X-선 결정술로부터 공지되어 있으므로, CDR과 가변 영역 골격 잔기 사이의 특이적 상호작용이 용이하게 모델링될 수 있다는 점이다.
본 발명자들은 국제특허출원 공개 제WO 03/100033호에 기재된 바와 같이 하이브리도마 6.3G9 및 6.8G6으로부터 단리된 mRNA로부터 항체 중쇄 가변 영역 cDNA 및 경쇄 가변 영역 cDNA를 제조하였다. 이들 하이브리도마들은 ανβ6 인테그린에 결합하는 IgG1 클래스 마우스 단일클론 항체를 생성한다. 키메라 인간 항체 발현 벡터는 인간 항체 중쇄 불변 영역 또는 인간 항체 경쇄 불변 영역 코딩 서열을 포함하는 발현 벡터 내로 cDNA를 삽입함으로써 제작하였다. 그 다음, 상기 벡터를 동물 세포 내로 도입하여 항-ανβ6 키메라 인간 항체를 생성하였다. 생성된 키메라 항체들 중에서, 항-ανβ6 키메라 인간 항체인 3G9 및 8G6은 ανβ6 인테그린과 반응하며 차단 활성을 나타내는 것으로 밝혀졌다.
전술한 방법들을 이용하여, 키메라 항체 3G9 및 8G6의 인간화 버젼을 생성시켰다. 3G9 항체의 경우, 이는 본 명세서의 실시예에 기재된 뮤린 3G9 가변 중쇄 및 경쇄 영역의 클로닝을 수반한다. 그 다음, 경쇄 및 중쇄의 뮤린 3G9 가변 영역을 코딩하는 cDNA를 사용하여, 본 명세서의 실시예에 기재된 바와 같이 뮤린 3G9 가변 영역이 인간 IgG1(중쇄의 경우) 및 인간 카파(경쇄의 경우) 불변 영역에 연결되어 있는 뮤린-인간 키메라의 발현을 위한 벡터를 제작하였다. 경쇄 및 중쇄 3G9 발현 벡터를 293-EBNA 세포 내로 형질감염시킨 후 상기 벡터를 발현시키면 키메라 3G9 형질감염 세포가 중쇄 및 경쇄를 합성하고 효율적으로 조립하고 항체를 분비한다는 것이 확인되었다(실시예 2 참조). 또한, 키메라 3G9 항체의 비글리코실화 돌연변이체 형태도 생성되었다. 3G9의 경쇄의 첫 번째 CDR 내의 N-결합 글리코실화 부위 내에서 아스파라진(N)을 세린(S)으로 아미노산 치환시키면 결합 친화성을 변경시키지 않으면서 단백질 발현 및 정제가 크게 개선되는 것으로 밝혀졌다(도 1).
인간화 3G9 항체를 생성하기 위해, 인간 수용 골격 도메인을 인간 생식세포 서열과의 상동성 일치를 통해 선택하였다. 실시예 3에 기재된 바와 같이, 경쇄의 경우, 인간 L6 수용 골격이 가장 높은 상동성을 가진 것으로 밝혀졌고, 중쇄의 경우, 인간 3 내지 7 수용 골격 영역이 가장 높은 상동성을 가진 것으로 밝혀졌다. 선택된 인간 수용 골격을 사용하여, 경쇄 및 중쇄 가변 도메인을 디자인하고 각각의 많은 변이체/버젼을 발생시키고 발현시켰다(실시예 4).
본 발명은 하기 서열번호 1의 중쇄 가변 도메인 및 서열번호 2의 경쇄 가변 도메인을 포함하는 인간화 3G9 항체를 개시한다.
상이한 정도의 복귀 돌연변이를 가진 3G9 중쇄 및 경쇄의 다양한 변이체/버젼을 발생시켜 어떤 조합이 ανβ6에 대한 우수한 결합 친화성 및 블로킹 활성을 가진 가장 우수한 인간화 항체를 생성하는지를 결정하였다. 경쇄의 5가지 상이한 버젼 및 중쇄의 3가지 상이한 버젼들 중에서, 3G9 중쇄 버젼 3(HV3)과 3G9 경쇄 버젼 5(LV5)의 조합이 가장 우수한 인간화 항체를 발생시켰다(실시예 4). 이 인간화 3G9 버젼 5(H3/L5) 항체는 플라스미드 pKJS195(서열번호 5)를 포함하는 경쇄 버젼 5(LV5)에 대한 재조합 벡터와 함께 플라스미드 pKJS189(서열번호 6)를 포함하는 중쇄 버젼 3(HV3)에 대한 재조합 벡터를 발현시켜 생성한다.
일반적인 Fc 수용체 결합에 필요한 것으로 밝혀진 불변 영역 내의 글리코실화 부위가 제거되도록 중쇄가 돌연변이되어 있는 인간화 3G9 버젼 5(H3/L5) 항체의 또 다른 버젼도 발생시켰다(실시예 5). 인간화 3G9 항체의 이러한 비글리코실화 형태(a-H3/L5)는 중쇄 버젼 3(H3)의 불변 영역에서 아미노산 잔기 아스파라진(N)을 글루타민(Q)으로 치환시킴으로써 생성한다. 비글리코실화 인간화 3G9(a-H3/L5) 항체는 플라스미드 pKJS195(서열번호 5, 상기 참조)를 포함하는 경쇄 버젼 5(L5)에 대한 재조합 벡터와 함께 플라스미드 pKJS196(서열번호 7)을 포함하는 비글리코실화 중쇄 버젼 3(a-H3)에 대한 재조합 벡터를 발현시켜 생성한다.
유사한 방법을 인간화 8G6 항체의 디자인에 사용하였다(실시예 7). 8G6 가변 경쇄 및 가변 중쇄의 3가지 버젼을 디자인하였는데, 제1 버젼은 가장 많은 복귀 돌연변이를 함유하고, 제3 버젼은 가장 적은 복귀 돌연변이를 함유한다(가장 "인간화됨")(실시예 5).
서열번호 75(hu8G6 버젼 1 경쇄)
서열번호 76(hu8G6 버젼 2 경쇄)
서열번호 77(hu8G6 버젼 3 경쇄)
서열번호 78(hu8G6 버젼 1 중쇄)
서열번호 79(hu8G6 버젼 2 중쇄)
서열번호 80(hu8G6 버젼 3 중쇄)
다른 잔기(moiety)
이하에 더 상세히 기재된 바와 같이, 본 발명의 인간화 단일클론 항체는 원하는 기능을 수행하기 위해 다른 잔기들을 추가로 포함할 수 있다. 예를 들면, 상기 인간화 항체는 항체의 표적이 되는 세포의 사멸을 위한 독소 잔기(예컨대, 파상풍 독소 또는 리신) 또는 방사성핵종(예컨대, 111In 또는 90Y)을 포함할 수 있다(예를 들면, 미국 특허 제6,307,026호 참조). 상기 인간화 항체는 용이한 단리 및 검출을 위한 잔기(예를 들면, 바이오틴, 형광 잔기, 방사성 잔기, 히스티딘 태그 또는 다른 펩티드 태그)를 포함할 수 있다. 상기 인간화 항체는 그의 혈청 반감기를 연장시킬 수 있는 잔기, 예컨대, 폴리에틸렌 글리콜 (PEG) 잔기를 포함할 수도 있다.
다양한 화학요법제가 표적 인간화 항체에 커플링될 수 있다. 바람직하게는, 결합 시 내부로 이입되는 인간화 항체가 가장 좋지만, 내부로 이입되지 않는 인간화 항체의 사용도 배제되지 않는다. 예를 들면, 종양 세포 표면에 결합하여 종양 내에서 또는 종양 세포 근처에서 약물을 방출시켜 세포 내로 확산 또는 수송하는 항체-약물 접합체의 사용이 사용된 약물에 따라 항-종양 활성을 부여할 수 있다. 당업자가 접합체를 제조하기 위해 사용할 수 있는 약물의 목록은 광범위하며, 당업자는 소정의 화합물의 반응이 본 발명의 접합체의 제조에 더 적합하게 하기 위해 상기 화합물에 화학적 변형을 가하는 방법을 알 것이다. 예를 들면, 약물은 "혈청 내에서 차등적으로 보다 더 안정하지만 종양 세포 내에서 활성 약물을 방출하는 방출가능한 링커"를 통해 커플링될 것이다. 구체적인 약물에 따라 여러 방출 기작이 이용될 수 있다. 이 방출 기작의 예에는 산-감수성 히드라존, 산화환원 감수성 링커, 예를 들면, 디설파이드, 단백질분해에 의해 절단되는 펩티드 링커의 사용이 포함된다. 하기 물질들은 여러 상이한 부류로부터의 일부 대표적인 약물이다:
(A) 알킬화제. 이 약물들의 일부 구체적인 예로는 사이클로포스파미드, 클로르암부실, 부설판, 멜팔란 및 니트로소우레아가 있다.
(B) 항대사물질 및 항-증식제 예컨대, 안트라사이클린(anthracycline), 빈카(vinca) 약물, 미토마이신, 블레오마이신, 뉴클레오시드, 프테리딘(pteridine), 엔디이네스(endiynes). 예로는 안드리아마이신(adriamycin), 다우노루비신(daunorubicin), 독소루비신(doxorubicin), 아미노프테린(aminopterin), 메쏘트렉세이트(methotrexate), 미토마이신 C, 액티노마이신-D, 5-플루오로우라실, 6-머캡토퓨린, 사이토신 아라비노사이드, 탁솔, 탁산(taxane), 사이토칼라신(cytochalasin) B, 콜치신(colchicin), 퓨로마이신, 에토포사이드, 멜팔란, 빈블라스틴, 빈크리스틴, 칼리케아미신, 메이타나신(maytanasine) 유도체 및 돌리스타틴(dolistatin) 유도체가 있다.
(C) 호르몬 및 호르몬 길항제 예컨대, 코르티코스테로이드, 프로게스틴 및 에스트로겐.
프로드러그는 항체에 부착될 때 "덜 강력한" 화학적 형태로 존재하지만, 내부로 이입되면 효소에 의해 절단되어 보다 강력한 약물 형태를 생성하는 약물로서 정의된다. 이것은 내부로 이입되지 않는, 예컨대, 효소에 의한 절단이 종양 세포 표면 상에서 일어나 약물이 즉시 종양 환경 내로 방출되고 종양 세포에 의해 동화되는 항체 접합체에도 적용될 수 있다. 이것의 일부 예로는 포스페이트, 설페이트 및 펩타이드를 함유하는 약물이다.
생물학적 활성을 가진 단백질 독소 예컨대, 리신 A 쇄, 디프테리아 독소, 시가독소, 파상풍 또는 독성 효소의 부착은 본 발명에 포함되는 항체-접합체의 또 다른 형태이다. 이러한 접합체는 당업자에게 잘 공지되어 있는 화학적 접합 방법, 또는 항체-독소 컨스트럭트의 직접적 발현을 허용하는 유전적 재조합 기법을 이용하여 제조할 수 있다.
본 발명의 인간화 단일클론 항체는 방사성핵종과 같은 다른 잔기를 포함할 수도 있다. 방사선면역요법을 위해, 암을 치료하기 위한 치료적 방사성 동위원소를 특이적으로 표적화하는 인간화 ανβ6 항체의 사용이 본 발명에 포함된다. 관련 동위원소의 목록은 90Y, 125I, 131I, 123I, 111In, 105Rh, 153Sm, 67Cu, 67Ga, 166Ho, 177Lu, 186Re 및 188Re를 포함하나 이들로 한정되지 않는다. 211At, 212Bi와 같은 알파 방출자 동위원소도 포함된다. 동위원소를 부착시키는 방법은 다양하고 사용된 특정 동위원소에 의존한다. 당업자는 임의의 특정한 동위원소 부착을 위한 접합 화학적 방법을 잘 알고 이를 결정할 수 있을 것이다.
방사선면역진단을 위해, 인간화 ανβ6 항체는 임의의 특정한 질환의 표적화된 암 및/또는 병든 기관/조직을 영상화하고 방사선량측정(dosimetry)을 실시할 기회를 제공할 수 있다. 이는 공지된 종양 부위로의 편재화(localization)를 확인하는 데 유용할 뿐만 아니라 치료적 투여의 최적화된 투여를 가능하게 하는 데 유용할 것이다. 특히, 순수한 감마 동위원소 99mTc 이외에 양전자 방사성 동위원소(예컨대, 86Y)가 치료적 투여 기간 동안 제공될 수 있다.
상기 방사선면역요법/방사선면역진단 적용은 내부로 이입되지 않는 항체의 사용으로 한정되지 않는다. 예로는 방사성 동위원소, 특히 이화(catabolism) 후 킬레이트로서 세포 내에 보유되는 동위원소를 사용한 표적화를 위한 내부이입 항체의 효과적인 사용이 있다. 예를 들면, 90Y-표지 항체를 MX-DTPA 또는 CHX-DTPA와 같은 고 친화성 킬레이터를 사용하여 제조한다.
상기 항체 접합체 중 임의의 항체 접합체는 단편 Fab, F(ab')2 scFvs, 미니바디(minibodies), CH2 도메인-결실 항체 컨스트럭트 및 FcRn-돌연변이체의 사용 또한 포함한다. 이들 Ab 단편 또는 일반적으로-변형된 컨스트럭트는 특정한 적용에서 이점을 제공할 수 있는 온전한 IgG와는 상이한 약동학적 성질, 종양 침투 성질 및 종양 편재화 성질을 가진다. 예를 들면, 보다 빨리-제거되는 Fab는 방사선면역진단 분야에서 진단용으로 유용할 수 있다. 다른 한편으로, 방사선면역요법 또는 약물 표적화의 경우, 보다 긴 혈청 t1/2를 가진 표적화 비히클을 선택하는 것이 보다 효과적이다.
병태 및 동물 모델
본 발명의 인간화 항체는 ανβ6-매개 질환의 예방을 비롯한 진단 및 치료에서 유용하다. 예를 들면, 이 인간화 항체는 TGF-β의 활성화를 차단하거나 ανβ6과 임의의 다른 리간드 예컨대, 피브로넥틴, 비트로넥틴 및 테나신의 결합을 차단함으로써 섬유화증(예컨대, 폐 섬유화증, 급성 폐 손상, 신장 섬유화증, 간 섬유화증, 알포츠 증후군 및 피부경화증), 및 본 명세서의 다른 곳에 기재된 다른 질환 및 장애의 치료를 위해 사용될 수 있다. 구체적으로, 본 발명의 인간화 항체는 특발성 폐 섬유화증, 방사선조사-유도 섬유화증, 만성 폐색성 폐 질환(COPD), 피부경화증, 블레오마이신-유도 섬유화증, 만성 천식, 실리코시스(silicosis), 석면-유도 섬유화증, 급성 폐 손상 및 급성 호흡곤란증(박테리아 뉴모니아-유도, 외상-유도, 바이러스 뉴모니아-유도, 통풍기-유도, 비-폐혈증-유도 및 흡입-유도 급성 호흡곤란증을 포함함)을 포함하나 이들로 한정되지 않는 손상/섬유화증과 관련된 폐 질환의 치료를 위해 사용될 수 있다. 본 발명의 인간화 항체는 루푸스, 당뇨병, 피부경화증, 사구체 신염, 국소성 및 분절성 사구체 경화증, IgA 신병증, 고혈압, 동종이식 및 알포츠병과 같은 손상/섬유화증과 관련된 만성 신병증의 치료를 위해 사용될 수도 있다. 인간화 항체는 장 섬유화증, 피부경화증, 방사선-유도 섬유화증의 치료를 위해 사용될 수도 있다. 본 발명의 인간화 항체는 담관 손상-유도 섬유화증을 포함하나 이로 한정되지 않는 간 섬유화증의 치료를 위해 사용될 수도 있다. 본 발명의 인간화 항체를 사용하여 치료할 수 있는 다른 증상에는 두경부 섬유화증, 방사선-유도 섬유화증, 각막 흉터형성, LASIX, 각막 이식, 섬유주절제술, 비대 흉터형성(hypertrophic scarring), 화상-유도 섬유화증, 수술적 섬유화증, 사르코이드증, 건선 및 척수 손상/섬유화증이 포함된다.
이하에 상세히 기재된 바와 같이, 섬유화증성 질환 또는 병태 이외에, 본 발명의 인간화 항체는 암 또는 암 전이(종양 성장 및 침입을 포함함), 특히 상피 암의 치료에 유용하다. 상피 암의 하위세트는 편평세포 암종, 예컨대, 두경부 암종(구강 암종, 후두 암종, 인두 암종, 식도 암종을 포함함), 유방 암종, 폐 암종, 전립선 암종, 자궁경부 암종, 결장 암종, 췌장 암종, 피부 암종(기저 세포 암종) 및 난소 암종이다. 신규한 ανβ6 단일클론 항체를 사용하는 본 연구는 ανβ6이 많은 상피 암, 특히 종양의 선도(leading) 가장자리에서 높은 수준으로 발현된다는 것을 입증하였다. 상기 신규 항체는 건선을 비롯한, ανβ6에 의해 매개되는 임의의 다른 질환에도 사용될 수 있다.
본 발명의 항체의 효능은 다양한 동물 모델에서 시험될 수 있고, 상기 모델들 중 일부는 이하의 비-제한적인 실시예에 기재되어 있다. 폐 섬유화증에 대한 마우스 모델에는 블레오마이신-유도가능성 폐 섬유화증(Pittet et al., J. Clin. Invest. 107(12): 1537-1544 (2001); and Munger et al., supra) 및 방사선-유도가능성 폐 섬유화증(Franko et al., Rad. Res. 140: 347-355 (1994))이 포함된다. 블레오마이신-처리 마우스에서, ανβ6의 발현은 폐의 상피 폐포 세포에서 증가한다. 그러나, β6 넉아웃(knockout) 마우스는 블레오마이신-유도 손상 및 섬유화증으로부터 보호된다.
신장 섬유화증에 대한 마우스 모델에는 COL4A3-/- 마우스[예컨대, 문헌(Cosgrove et al., Amer. J. Path. 157: 1649-1659 (2000))을 참조함], 아드리아마이신-유도 손상을 가진 마우스[Wang et al., Kidney International 58: 1797-1804 (2000); Deman et al., Nephrol. Dial. Transplant 16: 147-150 (2001)], db/db 마우스[Ziyadeh et al., PNAS USA 97: 8015-8020 (2000)], 및 편측 요관 폐쇄(unilateral ureteral obstruction)를 가진 마우스[Fogo et al., Lab Investigation 81: 189A (2001); and Fogo et al., Journal of the American Society of Nephrology 12: 819A (2001)]가 포함된다. 이 모델들 모두에서, 마우스는 신부전으로 진행할 수 있는 신장 손상 및 섬유화증을 발달시킨다. ανβ6은 COL4A3-/- 마우스, 아드리아마이신-처리 마우스 및 편측 요관 폐쇄를 앓는 마우스 신장의 상승 세뇨관과 하강 세뇨관의 경계를 형성하는의 상피에서 상향조절된다. ανβ6 발현도 다양한 신장 손상 모델에서 증가하는 것으로 보인다.
이하에 상세히 기재되는 바와 같이, 항-ανβ6 단일클론 항체는 표준 생체내 종양 성장 및 전이 모델과 같은 동물 모델에서 종양 성장, 진행 및 전이를 억제하는 그의 능력에 대해 시험될 수도 있다. 예를 들면, 문헌[Rockwell et al., J. Natl. Cancer Inst. 49: 735 (1972); Guy et al., Mol. Cell Biol. 12: 954 (1992); Wyckoff et al., Cancer Res. 60: 2504 (2000); and Oft et al., Curr. Biol. 8: 1243 (1998)]을 참조한다. 암에서 중요한 ανβ6 리간드에는 전이에 관여하는 TGF-β[검토를 위해서는 문헌(Akhurst et al., Trends in Cell Biology 11: S44-S51 (2001))을 참조함], 피브로넥틴 및 비트로넥틴이 포함될 수 있다.
본 발명의 치료 효능은 신체 검사, 혈액 시험, 단백뇨 측정, 크레아티닌 수준 및 크레아티닌 제거율, 폐 기능 시험, 혈장 혈액 우레아 질소(BUN) 수준, 흉터형성 또는 섬유화증성 병소의 관찰 및 스코어링, 세포외 매트릭스 예컨대, 콜라겐, 평활근 액틴 및 피브로넥틴의 침착, 신장 기능 시험, 초음파, 자기 공명 영상화(MRI) 및 CT 스캔을 비롯한 다수의 이용가능한 진단 수단에 의해 측정될 수 있다.
약학 조성물
본 발명은 경우에 따라 임의의 약학적으로 허용가능한 담체와 함께 본 발명의 하나 이상의 인간화 항체 또는 이의 약학적으로 허용가능한 유도체를 포함하는 약학 조성물을 제공한다. 본 명세서에서 사용된 용어 "담체"는 공지된 허용가능한 보조제 및 비히클을 포함한다.
본 발명에 따르면, 상기 약학 조성물은 주사가능한 멸균 제제, 예를 들면, 주사가능한 멸균 수성 또는 유성 현탁액의 형태일 수 있다. 이 현탁액은 적당한 분산제, 습윤제 및 현탁화제를 사용하여 당분야에 공지된 기법에 따라 제제화될 수 있다.
본 발명의 약학적 조성물은 소망에 따라 경구, 국소, 정맥내, 피하, 복강내, 근육내, 골수내, 동맥내, 활막내, 줄기내, 수막공간내, 간내 또는 두개내로 투여될 수 있거나, 염증 또는 종양 성장 부위에만 국소 투여될 수 있다. 본 발명의 약학 조성물은 예를 들면, 분무기, 건조 분말 흡입기 또는 계량 투여 흡입기의 사용을 통해 흡입 투여될 수도 있다.
원하는 효과를 제공하기에 효과적인 본 발명의 항체의 투여량 및 투여 속도는 다양한 인자, 예컨대, 치료될 질환의 성질, 투여받을 대상체의 크기, 치료 목적, 사용된 구체적인 약학 조성물 및 치료하는 의사의 판단에 달려 있을 것이다. 활성 성분 화합물의 유용한 투여 수준은 1일 당 약 0.001 내지 약 100 mg/체중 1 kg, 예컨대, 1일당 약 0.1 내지 약 50 mg/체중 1 kg이다. 예를 들면, 본 발명의 항체는 1일 내지 14일 간격으로 1일 당 약 0.01 내지 약 20 mg/체중 1 kg, 예컨대, 약 0.1 내지 약 10 mg/체중 1 kg의 투여량으로 투여될 것이다. 또 다른 실시양태에서, 본 발명의 항체는 복강내로 투여될 경우 약 0.3 내지 1 mg/체중 1 kg의 투여량으로 투여된다. 또 다른 실시양태에서, 본 발명의 항체는 정맥내로 투여될 경우 약 5 내지 12.5 mg/체중 1 kg의 투여량으로 투여된다. 한 실시양태에서, 항체 조성물은 1 mg/㎖ 이상의 항체 혈장 수준을 제공하기에 효과적인 양으로 투여된다.
다른 적합한 투여량 및 투여 섭생법 및 방식은 당업자에게 공지되어 있으며 이하에 더 상세히 기재되어 있다.
인테그린 α
ν
β
6
에 결합하는 리간드
추가 실시양태에서, 본 발명은 전이성 암세포를 확인하는 방법, 또는 종양 내의 세포에 의한 인테그린 ανβ6의 발현 수준을 측정함으로써 상기 세포의 전이력(생체 내에서 종양 내의 세포가 원발성 종양 부위에서 2차 전이 부위로 전이될 가능성)을 예측하는 방법에 관한 것으로서, 이때 세포 표면에서의 ανβ6 발현의 증가는 암세포가 더 높은 전이력을 가진다는 것을 의미한다. 관련 실시양태에서, 본 발명은 종양을 제거하기 위한 의학적 처치(예컨대, 종양의 수술적 제거, 또는 종양의 화학요법적 또는 방사선요법적 감소 또는 제거) 후 ανβ6을 발현하는 잔류 종양 세포, 특히 전이성 종양 세포를 제거하는 방법에 관한 것이다. 추가 관련 실시양태에서, 본 발명은 침입성 또는 전이성 형태로 진행될 가능성이 보다 더 높은 비침입성 형태의 암종, 특히 선암종 또는 상피내 암종(예컨대, 관 상내 유방 암종(DCIS) 또는 소엽 상피내 유방 암종(LCIS))을 확인하는 방법을 제공한다. 이러한 일부 실시양태는 상기 암종의 세포, 상기 암종을 둘러싸는 근육상피, 또는 상기 암종을 앓는 환자로부터 얻은 조직 절편에서의 인테그린 ανβ6의 발현 수준을 측정하는 단계를 포함하며, 이때 (이상적으로는, 동일한 환자의 동일한 기관으로부터 얻은) 비-종양 조직 샘플에 비해 인테그린 ανβ6의 증가된 발현 수준은 상기 암종이 가까운 장래에 언제가는 침입성 또는 전이성 형태의 암으로 진행할 가능성이 보다 더 높다는 것을 의미한다. 이러한 각각의 실시양태에서, 본 발명은 종양 세포 내의 ανβ6의 증가된 발현의 확인 또는 이용을 기초로 하며, 이 확인은 조직, 종양 또는 종양 세포를 조직, 종양 또는 종양 세포 내의 인테그린 ανβ6에 결합하는 하나 이상의 리간드와 접촉시킴으로써 달성한다. 일부 실시양태에서, 조직, 종양 또는 종양 세포는 암종 예컨대, 선암종으로부터 유래된 조직, 종양 또는 종양 세포를 비롯한 암종 조직, 종양 또는 종양 세포이다. 보다 구체적인 실시양태에서, 암종은 유방 암종, 자궁내막 암종, 췌장 암종, 결장직장 암종, 폐 암종, 난소 암종, 자궁경부 암종, 전립선 암종, 간 암종, 식도 암종, 두경부 암종, 위 암종 또는 비장 암종이다. 보다 구체적으로, 암종은 (관 상피내 암종(DCIS) 또는 소엽 상피내 암종(LCIS)과 같은 상피내 유방 암종을 포함하나 이들로 한정되지 않는) 유방 암종, 자궁내막 암종, 췌장 암종, 결장직장 암종, 자궁경부 암종 또는 폐 암종이다.
본 발명의 일부 실시양태에서, ανβ6에 결합하는 리간드는 ανβ6의 길항제이다. 이러한 길항제에는 ανβ6에 특이적으로 결합하는 항체; β6에 특이적으로 결합하는 항체; αν에 결합하는 항체; ανβ6에 대한 리간드에 결합하는 항체; ανβ6에 대한 리간드; 안티센스 핵산; 및 이러한 리간드의 펩티드, 비-펩티드 및 펩티드모사성 유사체가 포함되나, 이들로 한정되지 않는다.
본 발명의 이러한 일부 실시양태에서, 인테그린 ανβ6에 결합하는 리간드는 인테그린 ανβ6에 결합하는 항체, 또는 이의 인테그린 ανβ6-결합 단편, 변이체 또는 유도체이다. 이러한 항체는 상기 인테그린의 한 서브유니트에 결합할 수 있거나(예를 들면, αν 서브유니트 상에 위치된 에피토프 또는 β6 서브유니트 상에 위치된 에피토프에 결합하는 항체), 이들 서브유니트 둘다에 결합할 수 있다(예를 들면, αν 및 β6 서브유니트 둘다를 가교시키는 인테그린 이종이량체의 영역 내에 위치된 에피토프에 결합하는 항체). 천연 발생 항체와 같은 전장 길이의 항체를 구체적으로 언급하지 않는 한, 용어 "ανβ6 항체"는 전장 길이의 항체 뿐만 아니라 이 항체의 ανβ6-결합 단편, 변이체, 유사체 또는 유도체, 예를 들면, 천연 발생 항체, 또는 항체 분자와 유사한 방식으로 항원에 결합하는 면역글로불린 분자 또는 개조된 항체 분자 또는 단편도 포함한다. 항체는 합성 항체, 단일클론 항체 또는 다중클론 항체일 수 있고 당분야에 잘 공지되어 있는 기법에 의해 제조될 수 있다. 치료적 적용에서, 항체에 대한 환자의 면역 반응을 최소화하기 위해 인간 불변 및 가변 영역을 가진 "인간" 단일클론 항체가 종종 바람직하다. 이러한 항체는 인간 면역글로불린 유전자를 함유하는 형질전환 동물을 면역화시킴으로써 발생시킬 수 있다[예를 들면, 문헌(Jakobovits et al., Ann. N.Y. Acad. Sci. 764: 525-535 (1995))을 참조함]. 합성 항체 및 반-합성 항체의 경우, 상기 용어들은 항체 단편, 이소타입 전환 항체, 인간화 항체(예컨대, 마우스-인간, 인간-마우스 등), 하이브리드, 여러 특이성을 가진 항체, 완전히 합성된 항체-유사 분자 등을 포함하나, 이들로 한정되지 않는다.
용어 "항체" 및 "면역글로불린"은 본 명세서에서 상호교환적으로 사용된다. 항체 또는 면역글로불린은 중쇄의 하나 이상의 가변 도메인을 포함하고, 통상 중쇄 및 경쇄의 하나 이상의 가변 도메인을 포함한다. 척추동물 시스템 내의 기본 면역글로불린 구조는 비교적 잘 알려져 있다. 예를 들면, 문헌[Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988)]을 참조한다. 당업자가 이해하고 있는 바와 같이, 용어 "항체" 및 "면역글로불린"은 생화학적으로 구별될 수 있는 다양한 넓은 부류의 폴리펩티드를 포함한다. 당업자는 중쇄가 감마, 뮤, 알파, 델타 또는 엡실론(γ, μ, α, δ, ε)으로서 분류되며, 이들 중 일부는 하위분류로 더 분류된다(예를 들면, γ1 내지 γ4). 항체의 "부류"를 IgG, IgM, IgA, IgG 또는 IgE 각각으로서 결정하는 것은 항체 쇄의 성질이다. 면역글로불린 하위분류(이소타입) 예컨대, IgG1, IgG2, IgG3, IgG4, IgA1 등은 그 특징이 잘 규명되었고 기능적 전문화를 부여하는 것으로 공지되어 있다. 이들 부류 및 이소타입 각각의 변형된 버젼은 본 명세서의 개시내용에 비추어 당업자에게 용이하게 도출될 수 있으므로 본 발명의 범위 내에 있다.
본 발명에서 사용하기에 적합한, ανβ6에 결합하는 항체 또는 이의 ανβ6-결합 단편, 변이체 또는 유도체에는 다중클론 항체, 단일클론 항체, 다중특이적 항체, 인간 항체, 인간화 항체, 영장류화 항체, 키메라 항체, 단일쇄 항체, 에피토프-결합 단편, 예컨대, Fab, Fab' 및 F(ab')2, Fd, Fvs, 단일쇄 Fvs(scFv), 단일쇄 항체, 이황화-결합 Fvs(sdFv), VL 또는 VH 도메인을 포함하는 단편, Fab 발현 라이브러리에 의해 생성된 단편, 및 항-이디오타입 (항-Id) 항체(예컨대, 본 명세서에서 개시된 항-ανβ6 항체 대한 항-Id 항체를 포함함)가 포함되나, 이들로 한정되지 않는다. ScFv 분자는 당분야에 공지되어 있고, 예컨대, 미국 특허 제5,892,019호에 기재되어 있다. 본 발명의 면역글로불린 또는 항체 분자는 임의의 종류(예컨대, IgG, IgE, IgM, IgD, IgA, 및 IgY), 부류(예컨대, IgG1, IgG2, IgG3, IgG4, IgAl 및 IgA2) 또는 하위부류의 면역글로불린 분자일 수 있다.
단일쇄 항체를 비롯한 항체 단편은 가변 영역(들)을 단독으로 포함할 수 있거나 가변 영역(들)을 힌지 영역, CH1, CH2 및 CH3 도메인의 전부 또는 일부와 함께 포함할 수 있다. 또한, 본 발명에는 힌지 영역, CH1, CH2 및 CH3 도메인과 가변 영역(들)의 임의의 조합물을 포함하는 항체-결합 단편도 포함된다. 본 명세서에 개시된 진단 방법 및 치료 방법에 사용하기 위한 항체 또는 이의 면역특이적 단편은 새 및 포유동물을 포함하는 임의의 동물로부터 유래할 수 있다. 바람직하게는, 상기 항체는 인간, 뮤린, 래트, 당나귀, 토끼, 염소, 기니아 피그, 낙타, 라마, 말, 소 또는 닭 항체이다. 가장 바람직하게는, 상기 항체는 인간, 인간화 또는 영장류화 항체, 또는 키메라 항체, 특히 단일클론 항체이다. 본 명세서에 사용된 "인간" 항체에는 인간 면역글로불린의 아미노산 서열을 가진 항체가 포함되며, 인간 면역글로불린 라이브러리로부터 단리된 항체, 또는 전술된 바와 같이, 그리고 예컨대, 미국 특허 제5,939,598호(Kucherlapati et al.)에 기재된 바와 같이 내인성 면역글로불린을 발현하지 않는, 하나 이상의 인간 면역글로불린에 대한 형질전환 동물로부터 단리된 항체도 포함된다. 본 명세서에서 사용된 용어 "키메라 항체"는 면역반응성 영역 또는 부위가 제1 종으로부터 얻어지거나 유래되고 (본 발명에 따른 온전한 불변 영역, 부분 불변 영역 또는 변형된 불변 영역일 수 있는) 불변 영역이 제2 종으로부터 얻어진 임의의 항체를 의미할 것이다. 바람직한 실시양태에서, 표적 결합 영역 또는 부위는 비-인간 공급원(예컨대, 마우스 또는 영장류)으로부터 유래될 것이고, 불변 영역은 인간으로부터 유래된다.
본 발명에 따라 사용하기에 특히 바람직한 항체는 항-ανβ6 단일클론 항체 예컨대, 문헌[Weinreb et al., J. Biol. Chem. 279(17): 17875-17877 (2004)](이의 개시내용은 본 명세서에 참고로 완전히 도입됨)에 개시된 단일클론 항체 6.8G6("8G6") 및 6.3G9("3G9")를 포함하는, 상기 문헌에 개시된 항체와 같은 항-ανβ6 단일클론 항체이다. ανβ6에 결합하여 본 발명에 따라 사용하기에 적합한 추가 항체에는 인테그린 ανβ6의 β6 서브유니트에 결합하는 항체 (또는 이의 단편, 변이체 또는 유도체)(따라서, 이 항체는 "항-β6 항체"로 간주됨), 예컨대, 문헌[Weinacker et al., J. Cell Biol. 269: 1-9 (1994); 이의 개시내용은 본 명세서에 참고로 완전히 도입됨] 및 미국 특허 제6,692,741호 B2(이의 개시내용은 본 명세서에 참고로 완전히 도입됨)에 개시된 항체, 특히 상기 미국 특허의 컬럼 2 및 3과 7 및 8에 개시된 항체[10D5로 명명된 단일클론 항체(1997년 8월 6일 아메리칸 타입 컬쳐 콜렉션(버지니아주 20108 마나사스 피. 오. 박스 1549 소재)에 기탁된 ATCC 기탁번호 HB12382)(미국 특허 제6,692,741호의 컬럼 3, 7행 내지 13행 및 컬럼 7 및 8 참조) 및 CSβ6으로 명명된 단일클론 항체(미국 특허 제6,692,741호의 컬럼 7 및 8 참조)를 포함함]가 포함된다. 본 발명의 이 양태에 따른 적합한 실시양태는 ανβ6-결합 항체 또는 이의 ανβ6 에피토프-결합 단편인 ανβ6 인테그린-결합 리간드를 사용한다. 본 발명의 이 양태에 따라 사용하기에 적합한 추가 항체에는 미국 특허 출원 공보 제2005/0255102호 A1(이의 개시내용은 본 명세서에 참고로 완전히 도입됨)에 개시된 ανβ6-결합 단일클론 항체(상기 공보에서 3G9, 8G6, 1A8, 2B1, 2B10, 2A1, 2E5, IG1O, 7G5 및 1C5로 명명된 항체를 포함함) 뿐만 아니라 이의 단편, 키메라 및 하이브리드가 포함되나 이들로 한정되지 않는다. 본 발명에 따라 사용하기에 특히 적합한 항체는 단일클론 항체 2B1, 3G9 및 8G6이다.
일부 실시양태에서, 상기 항체는 하이브리도마 6.1A8, 6.3G9, 6.8G6, 6.2B1, 6.2B10, 6.2Al, 6.2E5, 7.1G10, 7.7G5 또는 7.1C5에 의해 생성된 항체와 동일한 중쇄 및 경쇄 폴리펩티드 서열을 포함한다. 본 발명에 따라 사용하기에 특히 적합한 항체는 하이브리도마 6.2B1(2001년 8월 16일 아메리칸 타입 컬쳐 콜렉션(버지니아주 20108 마나사스 피. 오. 박스 1549 소재)에 기탁된 ATCC 기탁번호 PTA-3646)에 의해 생성된 2B1 항체, 하이브리도마 6.8G6(2001년 8월 16일 아메리칸 타입 컬쳐 콜렉션(버지니아주 20108 마나사스 피. 오. 박스 1549 소재)에 기탁된 ATCC 기탁번호 PTA-3645)에 의해 생성된 8G6 항체 또는 하이브리도마 6.3G9(2001년 8월 16일 아메리칸 타입 컬쳐 콜렉션(버지니아주 20108 마나사스 피. 오. 박스 1549 소재)에 기탁된 ATCC 기탁번호 PTA-3649)에 의해 생성된 3G9 항체와 동일한 중쇄 및 경쇄 폴리펩티드 서열을 포함하는 단일클론 항체(미국 특허 출원 공보 제2005/0255102 Al호, 특히 제1면, 단락 0008; 제2면 단락 0032 및 0036; 및 제6면 내지 제14면의 실시예를 참조함, 상기 공보의 개시내용은 본 명세서에 참고로 완전히 도입됨) 및 10D5로 명명된 항체(이 항체를 분비하는 하이브리도마는 1997년 8월 6일 아메리칸 타입 컬쳐 콜렉션(버지니아주 20108 마나사스 피. 오. 박스 1549 소재)에 ATCC 기탁번호 HB12382로서 기탁됨)(그 개시내용이 본 명세서에 참고로 완전히 도입되는 미국 특허 제6,692,741호, 특히 컬럼 3, 제7행 내지 제13행 및 컬럼 7 및 8 참조)이다.
일부 실시양태에서, 상기 항체는 하기 표 1에 기재된 서열을 본질적으로 포함하는 (즉, 일부 보존적 변이가 제외된) 상보성 결정 영역(CDR) 1, 2 및 3을 가진 중쇄를 포함한다. 이러한 일부 실시양태에서, 항체는 서열번호 101 내지 105의 서열 중 어느 하나를 본질적으로 포함하는 CDR1, 서열번호 106 내지 111의 서열 중 어느 하나를 본질적으로 포함하는 CDR2, 및 서열번호 112 내지 117의 서열 중 어느 하나를 본질적으로 포함하는 CDR3을 가진 중쇄; 및/또는 서열번호 118 내지 123의 서열 중 어느 하나를 본질적으로 포함하는 CDR1, 서열번호 124 내지 127의 서열 중 어느 하나를 본질적으로 포함하는 CDR2, 및 서열번호 128 내지 133의 서열 중 어느 하나를 본질적으로 포함하는 CDR3을 가진 경쇄를 포함한다.
[표 1]
다른 관련 실시양태에서, 본 발명에 따라 사용되는 단일클론 항체는 키메라 항체, 즉 한 종(예컨대, 뮤린, 래트 또는 토끼)으로부터 유래된 동족 항체의 중쇄 및/또는 경쇄의 힌지 및/또는 불변 영역의 일부 또는 전부가 또 다른 종(예컨대, 인간)으로부터 유래된 항체의 상응하는 구성요소로 치환되도록 상기 동족 항체를 재조합 DNA 기법으로 변경시킨 항체이다. 일반적으로, 개조된 항체의 가변 도메인은 동족 항체의 가변 도메인과 동일하거나 실질적으로 동일하게 유지된다. 이러한 개조된 항체는 키메라 항체로 지칭되며, 힌지 및/또는 불변 영역이 유래된 종(예컨대, 인간)의 개체에게 투여될 때 상기 동족 항체보다 더 낮은 항원성을 가진다. 키메라 항체의 제조 방법은 당분야에 잘 공지되어 있다.
다른 관련 실시양태에서, 본 발명에 따라 사용되는 단일클론 항체는 완전한 인간 항체이다. 이러한 완전한 인간 단일클론 항체의 제조 방법은 당분야에 잘 공지되어 있다(예컨대, 본 명세서에 참고로 도입되는 미국 특허 출원 공보 제2005/0255102호 A1의 제4면, 단락 0069 내지 0070 참조).
다른 관련 실시양태에서, 본 발명에 따라 사용되는 단일클론 항체는 다른 종으로부터 유래된 항-ανβ6 항체의 인간화 버젼이다. 인간화 항체는 항원 결합에 필요하지 않은 인간 면역글로불린 경쇄 또는 중쇄 아미노산의 일부 또는 전부(예컨대, 불변 영역 및 가변 도메인의 골격 영역)가 동족 비-인간 항체의 경쇄 또는 중쇄로부터 유래된 상응하는 아미노산을 치환시키는 데 사용되는 재조합 DNA 기법에 의해 생성된 항체이다. 예를 들면, 소정의 항원에 대한 뮤린 항체의 인간화 버젼은 그의 중쇄 및 경쇄 둘다에서 (a) 인간 항체의 불변 영역; (b) 인간 항체의 가변 도메인으로부터 유래된 골격 영역; 및 (c) 뮤린 항체로부터 유래된 CDR을 가진다. 필요한 경우, 인간 골격 영역 내의 하나 이상의 잔기는 항원에 대한 인간화 항체의 결합 친화성을 보존하기 위해 뮤린 항체 내의 상응하는 위치에 있는 잔기로 변경될 수 있다. 이 변경은 종종 "복귀 돌연변이"로 지칭된다. 일반적으로, 인간화 항체는 키메라 인간 항체에 비해 인간에서 면역 반응을 덜 이끌어 내는 것으로 보이는데, 이는 인간화 항체가 상당히 더 적은 비-인간 구성요소들을 함유하기 때문이다. 이러한 인간화 단일클론 항체의 제조 방법은 당분야에 잘 공지되어 있다(예컨대, 본 명세서에 참고로 도입되는 미국 특허 출원 공보 제2005/0255102호 A1의 제4면 및 제5면, 단락 0072 내지 0077 참조).
이러한 추가 실시양태에서, 인간화 항체는 상이한 항체의 중쇄 및/또는 경쇄 내의 상응하는 CDR로부터 유래된 중쇄 및/또는 경쇄 내의 하나 이상의 CDR을 포함한다. 이러한 항체의 한 적합한 비-제한적 예는 기탁된 3G9 항체에 대한 경쇄 CDR1의 서열(서열번호 121) 대신에 2B1 항체로부터 유래된 경쇄 CDR1의 서열(서열번호 120)을 가진 경쇄 CDR1을 포함하는 인간화 3G9 항체이다. 서열번호 120에 기재된 경쇄 CDR1 서열을 가진 이러한 인간화 3G9 항체는 본 명세서에서 hu3G9(또는 BGOOO11)로서 지칭된다. 이러한 항체의 또 다른 적합한 비-제한적 예는 기탁된 8G6 항체에 대한 경쇄 CDR1의 서열(서열번호 118) 대신에 2B1 항체로부터 유래된 경쇄 CDR1의 서열(서열번호 120)을 가진 경쇄 CDR1을 포함하는 인간화 8G6 항체이다. 서열번호 120에 기재된 경쇄 CDR1 서열을 가진 이러한 인간화 86G 항체는 본 명세서에서 hu8G9로서 지칭된다. 하나 이상의 중쇄 및/또는 경쇄 CDR이 또 다른 항체로부터 유래된 하나 이상의 상응하는 중쇄 및/또는 경쇄 CDR로 치환되어 있으며 본 발명에 따라 사용하기에 적합한 상기 유도체성 항체들의 추가 예는 표 1에 기재된 서열 및 본 명세서에 제공된 지침에 비추어 당업자에게 용이하게 자명하다. 이러한 유도체성 인간화 항체들을 비롯한 상기 인간화 항체의 제조에 적합한 방법들은 당업자에게 잘 알려져 있으며, 예를 들면, 그 개시내용이 본 명세서에 참고로 완전히 도입되는 미국 특허 출원 공보 제2005/0255102호 A1에 기재되어 있다.
ανβ6-결합 리간드의 접합체 및 다른 변형물
일부 실시양태에서, ανβ6에 결합하는 리간드, 예를 들면, 항체는 접합되지 않은 형태로 사용될 수 있다. 다른 실시양태에서, ανβ6에 결합하는 리간드, 예컨대, 항체는 예를 들면, 검출가능한 표지, 약물, 프로드러그 또는 동위원소에 접합될 수 있다.
이하에 보다 상세히 기재될 본 발명의 일부 방법, 예컨대, 종양 세포의 전이력의 척도로서 또는 조직 내의 상피내 암종(예컨대, DCIS 또는 LCIS)을 확인하는 방법으로서 세포 또는 조직에서의 ανβ6 발현을 검출하는 방법에 있어서, ανβ6-결합 리간드(예컨대, 항체)는 하나 이상의 검출가능한 표지에 접합된다. 이러한 용도의 경우, ανβ6-결합 리간드, 예컨대, ανβ6-결합 항체는 발색성 표지, 효소성 표지, 방사성 동위원소 표지, 동위원소 표지, 형광성 표지, 독성 표지, 화학발광성 표지, 핵 자기 공명 조영제 또는 다른 표지의 공유결합 또는 비-공유결합에 의해 검출가능하게 표지될 수 있다.
적합한 발색성 표지의 예에는 디아미노벤지딘 및 4-히드록시아조-벤젠-2-카르복실산이 포함된다.
적절한 효소성 표지의 예로는 말레이트 데히드로게나제, 스타필로코칼 뉴클레아제, Δ-5-스테로이드 이소머라제, 효모-알코올 데히드로게나제, α-글리세롤 포스페이트 데히드로게나제, 트리오스 포스페이트 이소머라제, 퍼록시다제, 알칼리성 포스파타제, 아스파라기나제, 글루코스 옥시다제, β-갈락토시다제, 리보뉴클레아제, 우레아제, 카탈라제, 글루코스-6-포스페이트 데히드로게나제, 글루코아밀라제 및 아세틸콜린 에스터라제가 있다.
적합한 방사성 동위원소 표지의 예로는 3H, 111In, 125I, 131I, 32P, 35S, 14C, 51Cr, 57To, 58Co, 59Fe, 75Se, 152Eu, 90Y, 67Cu, 217Ci, 211At, 212Pb, 47Sc, 109Pd 등이 있다. 111In은 생체 내 영상화에서 사용되는 경우 바람직한 동위원소이데, 이는 111In이 간에 의한 125I 또는 131I-표지 ανβ6-결합 리간드의 탈할로겐화(dehalogenation)라는 문제점을 피하기 때문이다. 추가로, 이 방사성 뉴클레오티드는 화상 형성에 보다 유리한 감마 방출 에너지를 가진다(Perkins et al., Eur. J. Nucl. Med. 10: 296-301 (1985); Carasquillo et al., J. Nucl. Med. 25: 281-287 (1987)). 예를 들면, 1-(P-이소티오시아네이토벤질)-DPTA와 함께 단일클론 항체에 커플링된 111In은 비-종양성 조직, 특히 간에서 거의 흡수되지 않으므로, 종양 위치 확인의 특이성을 증강시킨다(Esteban et al., J. Nucl. Med. 28: 861-870 (1987)).
적합한 비-방사성 동위원소 표지의 예로는 157Gd, 55Mn, 162Dy, 52Tr 및 56Fe가 있다.
적합한 형광성 표지의 예로는 152Eu 표지, 플루오레세인 표지, 이소티오시아네이트 표지, 로다민 표지, 파이코에리쓰린 표지, 파이코시아닌 표지, 알로파이코시아닌 표지, 녹색 형광 단백질(GFP) 표지, o-프탈데히드 표지 및 플루오레스카민 표지가 있다.
적합한 독소 표지의 예로는 디프테리아 독소, 리신 및 콜레라 독소가 있다.
화학발광성 표지의 예로는 루미놀 표지, 이소루미놀 표지, 방향족 아크리디늄 에스테르 표지, 이미다졸 표지, 아크리디늄 염 표지, 옥살레이트 에스테르 표지, 루시페린 표지, 루시퍼라제 표지 및 애쿠오린 표지가 있다.
핵 자기 공명 조영제의 예로는 중금속 핵 예컨대, Gd, Mn 및 철이 있다.
전술한 표지와 ανβ6-결합 리간드, 예컨대, ανβ6-결합 항체의 전형적인 결합 기법은 문헌[Kennedy et al., Clin. Chim. Acta, 70: 1-31 (1976)] 및 문헌[Schurs et al., Clin. Chim. Acta. 81: 1-40 (1977)]에 의해 제공된다. 후자 문헌에서 언급된 커플링 기법은 글루타르알데히드 방법, 페리오데이트 방법, 디말레이미드 방법 및 m-말레이미도벤질-N-히드록시-석신이미드 에스테르 방법이고, 이들 방법 모두가 본 명세서에 참고로 도입된다.
수술 후 잔류 종양 세포의 제거 또는 전이 예방과 같은 본 발명의 일부 치료 방법에서 사용하기 위해, ανβ6-결합 리간드를 하나 이상의 약물, 프로드러그 또는 동위원소에 접합될 수 있다. 이러한 바람직한 접합체는 하나 이상의 세포독성 제에 접합된, ανβ6에 결합하는 하나 이상의 리간드, 예컨대, 하나 이상의 항체, 또는 그의 단편, 유도체 또는 변이체를 포함하며; 이러한 접합체는 본 발명에 의해 제공되는 종양 전이의 치료 및 예방 방법에서 유용하다. 본 발명의 이러한 일부 실시양태에 따르면, ανβ6-결합 리간드, 예컨대, 항체는 세포독성제에 접합된다. ανβ6-결합 리간드-세포독성제 접합체의 발생에 유용한 세포독성 물질, 예컨대, 화학요법제는 당분야에 잘 공지되어 있고, 그 예로는 시스플라틴, 카르보플라틴, 옥살리플라틴, 팩클리탁셀, 멜팔란, 독소루비신, 메쏘트렉세이트, 5-플루오로우라실, 에토포사이드, 메클로르에타민, 시클로포스파미드 및 블레오마이신이 있으나, 이들로 한정되지 않는다. 본 발명의 이 양태에 따라 사용하기에 적합한 다른 화학요법제도 잘 공지되어 있으며 당업자에게 친숙할 것이다.
하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체와 하나 이상의 소분자 독소, 예컨대, 칼리케아미신, 메이탄신(미국 특허 제5,208,020호), 트리코텐 및 CC1065로 구성된 접합체의 용도 또한 본원에 포함된다. 본 발명의 한 실시양태에서, ανβ6-결합 리간드는 하나 이상의 메이탄신 분자(예컨대, ανβ6-결합 리간드 당 약 1 내지 약 10개의 메이탄신 분자)에 접합된다. 예를 들면, 메이탄신은 May-SH3으로 환원될 수 있는 May-SS-Me로 전환될 수 있고 변형된 ανβ6-결합 리간드와 반응하여(Chari et al., Cancer Research 52: 127-131 (1992)) 메이탄시노이드-ανβ6-결합 리간드 접합체를 발생시킬 수 있다.
별법으로, ανβ6-결합 리간드는 하나 이상의 칼리케아미신 분자에 접합될 수 있다. 칼리케아미신 족의 항생제는 피코몰 농도 미만의 농도에서 이중-가닥 DNA 절단(break)을 발생시킬 수 있다. 사용될 수 있는 칼리케아미신의 구조적 유사체로는 γ1 I, α2 I, α3 I, N-아세틸-γ1 I, PSAG 및 Φ1 I가 있으나 이들로 한정되지 않는다(Hinman et al., Cancer Research 53: 3336-3342 (1993) and Lode et al., Cancer Research 58: 2925-2928 (1998)).
하나 이상의 ανβ6-결합 리간드, 예를 들면, 하나 이상의 ανβ6-결합 항체와의 접합체를 생성하는 데 사용될 수 있는 효소적 활성을 가진 독소 및 이의 단편으로는 디프테리아 A 쇄, 디프테리아 독소의 비-결합 활성 단편, 외독소 A 쇄(슈도모나스 애루기노사로부터 유래됨), 리신 A 쇄, 애브린 A 쇄, 모덱신 A 쇄, 알파-사르신, 알루리테스포르디이 단백질, 디안틴 단백질, 파이토라카 아메리카나 단백질(PAPI, PAPII, 및 PAP-S), 모모르디카 카란티아 억제제, 쿠르신, 크로틴, 사포나리아 오피시날리스 억제제, 겔로닌, 미토겔린, 레스트릭토신, 페노마이신, 에노마이신 및 트리코테센이 있다. 예를 들면, 1993년 10월 28일자로 영어로 공개된 국제특허출원 공개 제WO 93/21232호(이의 개시내용은 본 명세서에 참고로 완전히 도입됨)를 참조한다. 메이탄시노이드는 하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체에 접합될 수 있다.
추가로, 본 발명은 핵산분해 활성을 가진 화합물(예컨대, 리보뉴클레아제 또는 DNA 엔도뉴클레아제 예컨대, 데옥시리보뉴클레아제; DNase)과 접합된 ανβ6-결합 리간드를 포함한다.
다양한 방사성 동위원소 또한 본 발명의 치료 방법에서 사용될 수 있는, 방사성 동위원소-접합 ανβ6-결합 리간드의 생성에 유용하다. 예로는 211At, 131I, 125I, 90Y,186Re, 188Re, 153Sm, 212Bi, 32P, Lu의 방사성 동위원소들이 있다.
ανβ6-결합 리간드와 세포독성제의 접합체는 다양한 이기능성 단백질 커플링제 예컨대, N-석신이미딜-3-(2-피리딜디티올) 프로피오네이트(SPDP), 석신이미딜-4-(N-말레이미도메틸) 사이클로헥산-I-카르복실레이트, 이미노티올란(IT), 이미도에스테르의 이기능성 유도체(예컨대, 디메틸 아디프이미데이트 HCl), 활성 에스테르(예컨대, 디석신이미딜 수베레이트), 알데히드(예컨대, 글루타르알데히드), 히스-아지도 화합물(예컨대, 비스(p-아지도벤조일)헥산디아민), 비스-디아조늄 유도체(예컨대, 비스-(p-디아조늄벤조일)-에틸렌디아민), 디이소시아네이트(예컨대, 톨리엔 2,6-디이소시아네이트) 및 비스-활성 불소 화합물(예컨대, 1,5-디플루오로-2,4-디니트로벤젠)을 사용하여 제조할 수 있다. 예를 들면, 리신 면역독소는 문헌[Vitetta et al., Science 238: 1098 (1987)]에 기재된 바와 같이 제조될 수 있다. 14탄소-표지 1-이소티오시아네이토벤질-3-메틸디에틸렌 트리아민펜타아세트산(MX-DTPA)은 방사성 뉴클레오티드와 ανβ6-결합 리간드의 접합을 위한 예시적 킬레이팅제이다. 국제특허출원 제WO 94/11026호를 참조한다. 링커는 세포 내의 세포독성 약물의 방출을 용이하게 하는 "절단될 수 있는 링커"일 수 있다. 예를 들면, 산-불안정성 링커, 펩티다제-감수성 링커, 디메틸 링커 또는 디설피드-함유 링커(Chari et al., Cancer Research 52: 127-131 (1992))를 사용할 수 있다.
별법으로, ανβ6-결합 리간드 및 세포독성제를 포함하는 융합 단백질을 예컨대, 재조합 기법 또는 펩티드 합성으로 제조할 수 있다.
또 다른 실시양태에서, ανβ6-결합 리간드를 "예비표적화"에 사용하기 위해 "수용체"(예컨대, 스트렙타비딘)에 접합시킬 수 있고, 이때 ανβ6-결합 리간드-수용체 접합체를 환자에게 투여한 후 정화제(clearing agent)를 사용하여 순환계로부터 결합되지 않은 접합체를 제거한 다음, 세포독성제(예컨대, 방사성 뉴클레오티드)에 접합된 "리간드"(예컨대, 아비딘)를 투여한다.
본 발명의 ανβ6-결합 리간드는 프로드러그(예컨대, 펩티딜 화학요법제, 국제특허출원 공개 제WO 81/01145호 참조)를 활성 약물로 전환시키는 프로드러그-활성화 효소와 접합될 수도 있다. 예를 들면, 국제특허출원 공개 제WO 88/07378호 및 미국 특허 제4,975,278호를 참조한다. 이러한 접합체의 효소 성분은 프로드러그가 보다 높은 활성을 가진 그의 세포독성 형태로 전환될 수 있는 방식으로 프로드러그에 작용할 수 있는 임의의 효소를 포함한다.
본 발명의 방법에서 사용될 수 있는 효소에는 포스페이트-함유 프로드러그를 유리 약물로 전환시키는 데 유용한 알칼리성 포스파타제; 설페이트-함유 프로드러그를 유리 약물로 전환시키는 데 유용한 아릴설파타제; 비-독성 5-플루오로사이토신을 항암제, 즉 5-플루오로우라실로 전환시키는 데 유용한 사이토신 데아미나제; 펩티드-함유 프로드러그를 유리 약물로 전환시키는 데 유용한 프로테아제, 예컨대, 세라티아(serratia) 프로테아제, 써모라이신 서브틸리신, 카르복시펩티다제 및 카셉신(예컨대, 카셉신 B 및 L); D-아미노산 치환을 함유하는 프로드러그를 전환시키는 데 유용한 D-알라닐카르복시펩티다제; 글리코실화 프로드러그를 유리 약물로 전환시키는 데 유용한 탄수화물-절단 효소 예컨대, O-갈락토시다제 및 뉴라미니다제; P-락탐으로 유도체화된 약물을 유리 약물로 전환시키는 데 유용한 P-락타마제; 및 아민 질소에서 페녹시아세틸기 또는 페닐아세틸기로 유도체화된 약물을 유리 약물로 전환시키는 데 유용한 페니실린 아미다제 예컨대, 페니실린 V 아미다제 또는 페니실린 G 아미다제가 포함되나, 이들로 한정되지 않는다.
효소는 이종이기능성 가교제의 사용과 같은, 당분야에 잘 공지되어 있는 기법에 의해 ανβ6-결합 리간드에 공유결합될 수 있다. 별법으로, 효소의 하나 이상의 기능적 활성 부위에 결합된 본 발명의 ανβ6-결합 리간드의 하나 이상의 항원 결합 영역을 포함하는 융합 단백질을 당분야에 잘 공지되어 있는 재조합 기법을 이용하여 제작할 수 있다[예컨대, 문헌(Neuberger et al., Nature 312: 604-608 (1984)) 참조].
질환 진단 및 예후
본 발명자들은 특정한 전이성 종양으로부터 유래된 세포가 전이성이 보다 더 낮거나 전이성이 없는 세포에 비해 유의하게 증가된 수준의 인테그린 ανβ6을 발현한다는 사실을 발견하였다. 또한, 본 발명자들은 상피내 암종의 특정한 형태, 예컨대, 관 상피내 유방 암종(DCIS) 또는 소엽 상피내 유방 암종(LCIS)에서 종양을 둘러싸는 근상피가 상기 암종의 종양 세포 및 정상 종양 조직에 비해 유의하게 증가된 수준의 인테그린 ανβ6을 발현한다는 사실을 발견하였다. 그러므로, 본 발명은 암종 예컨대, 선암종으로부터의 종양을 비롯한 종양 세포의 전이력을 진단하는 데 있어서 유용한 방법을 제공한다. 보다 구체적인 실시양태에서, 암종은 유방 암종, 자궁내막 암종, 췌장 암종, 결장직장 암종, 폐 암종, 난소 암종, 자궁경부 암종, 전립선 암종, 간 암종, 식도 암종, 두경부 암종, 위 암종 또는 비장 암종이다. 보다 구체적으로, 암종은 유방 암종(예컨대, 관 상피내 유방 암종(DCIS) 또는 소엽 상피내 유방 암종(LCIS)과 같은 상피내 유방 암종을 포함하나 이들로 한정되지 않음), 자궁내막 암종, 췌장 암종, 결장직장 암종, 자궁경부 암종 또는 폐 암종이다.
본 발명의 이 양태에 따른 방법은 조직 샘플 내의 종양 세포 또는 근상피에서의 ανβ6의 발현 수준을 분석하는 단계, 및 이 발현 수준을 표준 ανβ6 발현 수준(예컨대, 바람직하게는 동일한 동물 예컨대, 인간 환자로부터 얻은 정상 세포, 비-전이성 세포 또는 정상 조직에서의 ανβ6 발현 수준)과 비교하는 단계를 포함하며, 이때 종양 또는 이의 세포 내에서의 ανβ6의 발현 증가는 그 종양 또는 그의 세포가 높은 침입력 및/또는 전이력을 가지고 있다는 것을 표시하고, 종양을 둘러싸는 근상피에서의 ανβ6의 발현 증가 또는 조직 절편 내의 상피 세포 덩어리에서의 ανβ6의 발현 증가는 침입성을 가지게 되어 잠재적으로 전이를 형성할 가능성이 높은 상피내 암종 예컨대, DCIS 또는 LCIS의 존재를 표시한다.
통상적인 방법에 따라 이미 암으로 진단받은 경우, 본 발명은 예후 지표로서 유용하므로, ανβ6의 증가된 발현 수준을 나타내는 종양 세포는 침입성을 가지게 되어 원발성 종양 부위에서 멀리 떨어진 전이 부위로 전이될 가능성이 있는 것으로 예측될 것이다. 유사하게, 통상적인 방법(예컨대, 유방 내의 석회화된 결절의 유방촬영 검출)에 따라 상피내 암종으로 의심된다는 진단을 받은 경우, 본 발명은 확증 지표로서 유용함으로, 근상피에서의 ανβ6의 증가된 발현 수준을 나타내는 석회화 영역으로부터 얻은 생검 조직은 침입성을 가지게 되어 ανβ6 mAb 치료에 반응할 수 있는 상피내 암종, 예컨대, DCIS 또는 LCIS의 존재를 표시한다. 이러한 예후 및 진단 결과를 기초로 하여, 치료 임상의는 치료요법을 그에 알맞게 조정하여, 전이-전 또는 암-전 상태의 초기 검출을 제공함으로써 환자에게 보다 유리한 임상적 결과를 제공할 수 있다.
"ανβ6의 발현 수준의 분석"은 제1 생물학적 샘플(예컨대, 종양 샘플, 조직 생검 또는 흡입물 등)에서의 ανβ6의 수준을 직접적으로(예컨대, 샘플에서의 ανβ6의 절대량을 측정하거나 평가함으로써) 또는 간접적으로(예컨대, 제2 생물학적 샘플 중의 ανβ6의 발현 수준과 제1 생물학적 샘플 중의 ανβ6의 발현 수준을 비교함으로써) 정성적으로 또는 정량적으로 측정하거나 평가하는 것을 의미한다. 바람직하게는, 제1 생물학적 샘플 중의 ανβ6의 수준을 측정하거나 평가하고, 암 또는 암-전 병소를 가지지 않은 개체로부터 수득된 제2 생물학적 샘플로부터 얻은 표준물 중의 ανβ6의 수준과 비교한다. 당업자에게 인식되는 바와 같이, 일단 표준 ανβ6 발현 수준이 소정의 비-암성 조직에 대해 공지되면, 이는 비교를 위한 표준으로서 반복적으로 사용될 수 있다.
"생물학적 샘플"은 개체(예컨대, 환자), 세포주, 조직 배양물, 또는 세포외 매트릭스와 같은 세포 생성물 또는 세포를 함유할 수 있는 다른 공급원으로부터 수득된 임의의 생물학적 샘플이다. 이러한 생물학적 샘플에는 백혈구, 난소, 전립선, 심장, 태반, 췌장, 간, 비장, 폐, 유방, 두경부 조직(예컨대, 구강, 인두, 설하 및 후두 조직), 자궁내막, 결장(또는 결장직장), 자궁, 위 및 ανβ6을 발현할 수 있는 배꼽 조직을 비롯한 포유동물 신체 조직 및 세포가 포함된다. 조직 생검 및 체액을 포유동물로부터 수득하는 방법은 당분야에 잘 공지되어 있다. 바람직한 포유동물에는 원숭이, 유인원, 고양이, 개, 소, 돼지, 말, 토끼 및 인간이 포함된다. 특히, 인간이 바람직하다.
생물학적 샘플 중의 ανβ6 발현 수준은 당분야에 공지되어 있는 임의의 방법을 이용하여 분석할 수 있다. 생물학적 샘플 중의 ανβ6 발현 수준의 분석에는 면역학적 기법이 바람직하다. 예를 들면, 조직에서의 ανβ6 발현은 고전적인 면역조직학적 방법으로 연구될 수 있다. 여기서, 특이적 인식은 1차 리간드, 예컨대, ανβ6에 결합하는 항체(다중클론 또는 단일클론)에 의해 제공된다. 이 1차 리간드는 예를 들면, 형광성 표지, 화학발광성 표지, 인광성 표지, 효소성 표지 또는 방사성 동위원소 표지로 표지될 수 있다. 별법으로, 본 발명의 이 방법들은 ανβ6-결합 리간드를 인식하여 이에 결합하는 2차 리간드, 예를 들면, 1차 ανβ6-결합 항체를 인식하여 이에 결합하는 소위 "2차" 항체가 전술한 바와 같이 검출가능하게 표지되어 있는 2차 검출 시스템을 이용할 수 있다. 그 결과, 병리학적 검사에 대한 조직 절편의 면역조직학적 염색이 얻어진다. 별법으로, ανβ6의 발현 수준이 보다 더 낮은 것으로 공지되어 있는 표준 조직 또는 세포 샘플을 기준으로 한 직접적 정량화를 위한 웨스턴-블롯 또는 도트/블롯 분석(Jalkanen, M., et al., J. Cell. Biol. 101: 916-985 (1985); Jalkanen, M., et al., J. Cell . Biol. 105: 3087-3096 (1987))을 위해 ανβ6 단백질을 유리시키기 위해, 조직 및 세포 샘플을 예컨대, 우레아 및 중성 세제로 추출할 수도 있다.
전술한 바와 같이, 본 발명의 방법은 포유동물에서의 전이성 암의 검출, 종양 세포의 전이력의 측정(즉, 소정의 종양 세포가 원발성 종양 부위에서 멀리 떨어진 전이 부위까지 전이할 가능성의 예측), 비-침입성 또는 상피내 암종이 침입성 또는 전이성 암종으로 진행될 가능성의 결정에 유용하다. 구체적으로, 본 발명의 방법은 유방, 난소, 전립선, 간, 폐, 췌장, 결장(또는 결장직장), 두경부 조직(예컨대, 구강, 인두, 설하 및 후두 조직), 자궁내막, 자궁, 위 및 비장을 비롯한 상피 조직(즉, 침입성 및/또는 전이성 암종)의 침입성 및/또는 전이성 암을 검출하는 데 유용하다. 본 발명의 방법은 침입성 및/또는 전이성 표현형으로 진행될 가능성이 증가된 유방 암종, 췌장 암종, 결장직장 암종, 자궁경부 암종, 폐 암종, 및 상피내 암종 예컨대, 특정한 관 상피내 유방 암종(DCIS) 또는 소엽 상피내 유방 암종(LCIS)을 비롯한 침입성 및/또는 전이성 선암종을 검출하는 데 특히 적합하다. 이러한 암종의 초기 확인 및 치료는 환자에 대한 우수한 장기 예후와 관련되어 있다. 예를 들면, 치료받지 않으면 DCIS 종양의 상당한 비율이 침입성을 가지게 되고 훨씬 더 낮은 예후를 가지는 전이성 암을 초래할 수 있다고 보고되어 있다[문헌(Sakorafas, G.H., and Tsiotou, A.G.H., Cancer Treatment Rev. 2(6): 103-125 (2000)) 참조].
따라서, 본 발명은 환자에서 침입-전 병소 또는 암종을 확인함으로써 전이성 암을 치료하거나 예방하는 방법, 및 침입-전 병소가 침입성 형태로 진화할 기회를 가지기 전에 침입-전 병소를 제거하여 환자를 치료하는 방법을 포함한다. 이러한 방법은 예를 들면, 암 또는 침입-전 병소를 함유하는 것으로 의심되는 조직 샘플, 및 암 또는 침입-전 병소를 함유하지 않는 조직 샘플(바람직하게는 암 또는 침입-전 병소를 함유하는 것으로 의심되는 조직 또는 기관과 동일한 조직 또는 기관으로부터 얻은 조직 샘플)을 얻는 단계; (b) 존재하는 경우 언제나 조직 내에서 하나 이상의 ανβ6-결합 리간드와 ανβ66-인테그린의 결함에 유리한 조건 하에서 상기 조직 샘플을 하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체 또는 이의 단편과 접촉시키는 단계; (c) ανβ6-결합 리간드(들)와 상기 조직의 결합 수준 또는 패턴을 검출하는 단계를 포함하며, 이때 과다증식물 자체(또는 이의 세포)에서의 ανβ6-결합 리간드의 결합에 비해 과다증식물(예컨대, 종양)을 둘러싸는 근상피에서의 ανβ6-결합 리간드의 편재화된 결합의 증가, 또는 비-암성 조직 샘플(또는 이의 세포)에서의 ανβ6-결합 리간드의 결합에 비해 암성 또는 침입-전 병소를 함유하는 조직 샘플에서의 ανβ6-결합 리간드의 결합 수준의 증가는 침입성을 가지게 되어 잠재적으로 전이될 가능성이 보다 더 높은 암종을 표시한다. 다른 관련 실시양태에서, 본 발명은 전이-전 또는 침-전 종양 내의 하나 이상의 세포 상의 인테그린 ανβ6의 하나 이상의 서브유니트에 결합하는 치료 유효량의 하나 이상의 리간드를 환자에게 투여하는 단계를 포함하는, 환자에서 전이 전 또는 침입 전 종양이 전이성 또는 침입성 종양으로 진행하는 것을 감소시키거나 예방하는 방법을 포함하며, 이때 상기 인테그린과 상기 리간드의 결합은 원발성 종양을 둘러싸는 조직 영역 내로 전이-전 또는 침입-전 암 세포가 침입되는 것을 감소시키거나 예방한다.
본 발명의 이 방법들에 따라 사용하기 위해 적합한, 샘플로부터 얻어질 수 있는 조직 및 기관에는 본 명세서의 다른 곳에 기재된 상피 조직이 포함되나, 이로 한정되지 않는다. 본 발명의 이러한 방법들에 따라 유리하게 치료되거나 예방될 수 있는 암 및 종양에는 본 명세서에서 상세히 기재된 암종 및 선암종을 비롯한 암종, 특히 선암종이 포함되나 반드시 이들로 한정되지 않는다. 이러한 암종이 일단 본 발명의 방법에 따라 검출되면, 암종은 당분야에 잘 공지되어 있어 당업자에게 친숙한 수술, 화학요법, 방사선요법 또는 다른 암 치료요법을 통해 환자로부터 제거될 수 있다. 별법으로, 이러한 암종은 하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체 또는 이의 단편을 환자에게 또는 환자의 기관 또는 조직에 투여함으로써 본 발명의 치료 방법을 이용하여 제거할 수 있다. 이러한 실시양태의 일부 비-제한적 예에서, 하나 이상의 ανβ6-결합 리간드는 앞서 상세히 기재된 바와 같은 하나 이상의 세포독성 화합물 또는 물질과 접합되어 있다. 이러한 실시양태의 추가 비-제한적 예에서, 하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체 또는 이의 단편은 앞서 상세히 기재된 바와 같은 하나 이상의 세포독성 화합물 또는 물질과 함께 대상체, 예컨대, 환자에게 투여된다.
관련 실시양태에서, 본 발명은 종양 또는 암세포에 의한 ανβ6의 발현을 측정함으로써 종양 또는 암세포의 전이력을 결정하는 것을 포함한다. 이러한 실시양태에서, 종양 또는 세포 샘플은 전술한 바와 같이 환자로부터 얻으며, 그 종양 또는 암세포 상의 ανβ6의 발현 수준에 대해 본원에 기재된 방법에 따라 분석한다. 바람직한 이러한 방법에는 ανβ6-결합 항체(또는 이의 단편, 변이체 또는 유도체), 예컨대, 본원에 기재된 항체를 사용하는 면역조직화학적 기법이 포함된다. 본 발명의 이 방법들에 따라, 종양 또는 암세포에 의한 ανβ6의 발현 수준과 그 종양 또는 암세포의 전이력 사이에 직접적인 상관관계가 있다: 종양 또는 암세포에 의한 ανβ6의 발현 증가는 그 종양 또는 암세포가 원발성 종양 부위로부터 떨어진 2차 위치로 전이될 가능성이 보다 더 높다는 것을 의미한다. 따라서, 종양 또는 암세포에 의한 ανβ6의 발현 수준은 종양 또는 암세포의 전이력의 예후 지표로서 사용될 수 있고, 이 예후 지표는 암 환자 및 그의 임상의가 현재의 또는 예측된 장래의 암의 공격성 또는 침입성을 기초로 한 적절한 치료 결정을 하는 데 있어서 도움이 될 수 있다.
개체로부터 얻은 생물학적 샘플, 예컨대, 조직 또는 종양 세포 샘플 중의 ανβ6 발현 수준을 분석하는 것 외에, ανβ6의 발현 수준 및 패턴도 영상화에 의해 생체 내에서 검출될 수 있다. 본 발명의 이러한 방법에서, 하나 이상의 ανβ6-결합 리간드, 예컨대, 하나 이상의 ανβ6-결합 항체는 생체 내 영상화에 적합한 하나 이상의 표지로 검출가능하게 표지된다. 생체 내 영상화에 적합한 표지 또는 마커에는 X-방사선촬영, NMR 또는 ESR에 의해 검출가능한 것들이 포함된다. X-방사선촬영의 경우, 적합한 표지에는 검출가능한 방사선을 방출하나 대상체에게 명백히 해롭지 않은 바륨 또는 세슘과 같은 방사성 동위원소가 포함된다. NMR 및 ESR에 적합한 표지에는 검출가능한 특징적인 스핀을 가진 표지 예컨대, 중수소가 포함된다.
적절한 검출가능한 영상화 잔기, 예컨대, 방사성 동위원소(예컨대, 131I, 112In, 99mTc), 방사선-불투과성 물질, 또는 핵 자기 공명에 의해 검출가능한 물질로 표지된, ανβ6에 결합하는 리간드, 예를 들면, ανβ6-결합 항체 또는 항체 단편을 포유동물 내로 도입하여 암 또는 상피내 암종을 검사한다. 대상체의 크기 및 사용된 영상화 시스템이 진단 영상을 생성하는 데 필요한 영상화 잔기의 양을 결정할 것이라는 것은 당업자에게 인식되어 있을 것이다. 방사성 동위원소 잔기의 경우, 인간 대상체에게 주입되는 방사성의 양은 통상 약 5 내지 20 밀리큐리의 99mTc일 것이다. 그 후, 표지된 ανβ6 리간드, 예를 들면, ανβ6-결합 항체 또는 항체 단편은 ανβ6 인테그린을 함유하거나 발현하는 세포 또는 조직의 위치에서 우세하게 축적될 것이다. 그 다음, 생체내 종양 영상화는 문헌[S. W. Burchiel et al., "Immunopharmacokinetics of Radiolabeled Antibodies and Their Fragments" (Chapter 13 in Tumor Imaging: The Radiochemical Detection of Cancer, S.W. Burchiel and B.A. Rhodes, eds., Masson Publishing Inc. (1982))]에 기재된 바와 같이 달성된다.
α
ν
β
6
-결합 리간드의 치료적 용도
본 발명의 추가 실시양태에서, ανβ6-결합 리간드, 예컨대, ανβ6-결합 항체 또는 이의 단편은 특정 질환, 특히 본 명세서의 다른 곳에 기재된 것과 같은 특정 암종을 앓는 포유동물을 치료하기 위한 치료요법에 사용될 수 있다. 본 발명의 이러한 방법은 암, 및 (종양 성장, 전이 및 혈관신생을 포함하는) 관련 과정을 치료하는 데 유용하다. 이러한 방법으로 치료하기에 특히 적합한 것은 질환을 앓는 포유동물의 조직 또는 세포에서의 증가된 ανβ6 발현 수준을 특징으로 하며 ανβ6의 증가된 수준을 발현하는 조직 또는 세포를 표적화하여 이 조직 또는 세포를 제거하는 치료에 반응하는 질환 또는 암이다. 이 방법으로 특히 치료가능한 질환에는 유방, 난소, 전립선, 간, 폐, 췌장, 결장, 두경부 조직(예컨대, 구강, 인두, 설하 및 후두 조직), 자궁내막, 자궁, 위 및 비장을 비롯한 상피 조직의 전이성 암(즉, 전이성 암종 및/또는 선암종)이 포함된다. 본 발명의 방법에 의해 치료하기에 특히 적합한 것은 자궁내막 암종, 췌장 암종, 결장 암종(예컨대, 결장직장 암종), 자궁경부 암종, 폐 암종 및 유방 암종(관 상피내 유방 암종(DCIS) 및 소엽 상피내 암종(LCIS)을 포함함)이다. 치료하기에 바람직한 포유동물에는 원숭이, 유인원, 고양이, 개, 소, 돼지, 말, 토끼 및 인간이 포함된다. 특히 인간이 바람직하다.
이러한 특정한 치료요법에서, 본 발명의 방법은 상이한 방법에 의해 종양의 제거, 치료 또는 박멸 후 잔류하는 종양 세포, 예컨대, 잔류 전이 세포를 제거하기에 적합하다. 예를 들면, 본 발명의 이러한 방법은 종양의 수술적 절제, 또는 방사선요법, 화학요법 등과 같은 방법에 의한 종양 박멸 후 환자에게 잔존할 수 있는 잔류 종양 세포 또는 전이 세포를 제거하는 데 이용될 수 있다. 이러한 치료요법에서, 본 발명의 방법은 종양의 수술적, 방사선적 및/또는 화학요법적 제거 전, 제거 동안 및/또는 제거 후 ανβ6-결합 리간드, 예를 들면, ανβ6-결합 항체 또는 이의 단편을 환자에게 투여하는 단계를 포함할 수 있다.
관련 실시양태에서, 전술한 바와 같이, 본 발명은 전이-전 종양에 있는 하나 이상의 세포 상의 인테그린 ανβ6의 하나 이상의 서브유니트에 결합하는 치료 유효량의 하나 이상의 리간드를 환자에게 투여하는 단계를 포함하는, 환자에서 전이-전 종양이 전이성 종양으로 진행하는 것을 감소시키거나 예방하는 방법을 제공하며, 이때 상기 리간드와 상기 인테그린의 결합은 전이-전 암 세포가 원발성 종양을 둘러싸는 조직 영역 내로 침입하는 것을 감소시키거나 예방한다.
본 발명의 이 치료 방법을 수행함에 있어서, ανβ6-결합 리간드, 예컨대, ανβ6-결합 항체 또는 이의 단편은 치료 제제(본원에서 약학 조성물과 상호교환적으로 또는 동등하게 지칭됨)의 형태로 환자에게 투여될 수 있다. 본 발명에 따라 사용되는 ανβ6-결합 리간드의 치료 제제는 원하는 정도의 순도를 가진 ανβ6-결합 리간드를 경우에 따라 약학적으로 허용가능한 담체, 부형제 또는 안정화제(Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980))와 함께 예를 들면 동결건조 제제 또는 수용액의 형태로 혼합하여 저장용으로 제조된다. ανβ6-결합 리간드와 같은 약리학적 활성 화합물 이외에, 본 발명의 치료 방법에서 사용되는 조성물은 활성 화합물이 약학적으로 사용될 수 있는 제제로 용이하게 가공될 수 있도록 부형제 및 보조제를 포함하는 하나 이상의 약학적으로 허용가능한 적합한 담체를 함유할 수 있다. 본 발명의 약학 제제는 그 자체가 공지되어 있는 방식, 예컨대, 통상적인 혼합, 과립화, 당의정-제조, 용해 또는 동결건조 공정에 의해 제조된다. 따라서, 경구용 약학 제제는 활성 화합물을 고체 부형제와 혼합하고, 경우에 따라 생성된 혼합물을 분쇄하고, 원하거나 필요한 경우 적절한 보조제를 첨가한 후 과립 혼합물을 가공하여 정제 또는 당의정 코어를 수득함으로써 얻을 수 있다.
적합한 부형제는 특히, 사카라이드와 같은 충진제, 예를 들면, 락토스 또는 수크로스, 만니톨 또는 소르비톨, 셀룰로스 제제 및/또는 인산칼슘, 예를 들면, 제3 인산칼슘 또는 인산수소칼슘 뿐만 아니라, 결합제, 예컨대, 옥수수 전분, 밀 전분, 쌀 전분, 감자 전분을 사용하는 전분 페이스트, 젤라틴, 트라가칸쓰, 메틸 셀룰로스, 히드록시프로필메틸셀룰로스, 나트륨 카르복시-메틸셀룰로스 및/또는 폴리비닐 피롤리돈이다. 원하는 경우, 전술한 바와 같은 전분 및 또한 카르복시메틸-전분, 가교 폴리비닐 피롤리돈, 아가 또는 알긴산 또는 이의 염, 예컨대, 알긴산나트륨과 같은 붕해제를 첨가할 수 있다. 특히, 보조제는 유동-조절제 및 윤활제, 예를 들면, 실리카, 탈크, 스테아르산 또는 이의 염, 예컨대, 스테아르산마그네슘 또는 스테아르산칼슘, 및/또는 폴리에틸렌 글리콜이다. 원하는, 경우, 위액에 대한 내성을 가진 적당한 코팅물을 당의정 코어에 제공할 수 있다. 이를 위해, 경우에 따라 아라비아검, 탈크, 폴리비닐 피롤리돈, 폴리에틸렌 글리콜, 및/또는 이산화티탄, 락커용액 및 적절한 유기 용매 또는 용매 혼합물을 함유할 수 있는 농축된 사카라이드 용액을 사용할 수 있다. 위액에 대한 내성을 가진 코팅물을 제조하기 위해, 적절한 셀룰로스 제제 예컨대, 아세틸셀룰로스 프탈레이트 또는 히드록시프로필메틸셀룰로스 프탈레이트를 사용한다. 활성 화합물의 투여량의 조합을 확인하거나 그 특징을 규명하기 위해 염료 물질 또는 색소를 정제 또는 당의정 코팅물에 첨가할 수 있다.
경구 투여될 수 있는 다른 약학 제제에는 젤라틴으로 만들어진 푸쉬-피트(push-fit) 캡슐 뿐만 아니라, 젤라틴과 가소제 예컨대, 글리세롤 또는 소르비톨로 만들어진 연질 밀봉 캡슐이 포함된다. 푸쉬-피트 캡슐은 락토스와 같은 충진제, 전분과 같은 결합제, 및/또는 탈크 또는 스테아르산마그네슘과 같은 윤활제, 및 경우에 따라 안정화제와 혼합될 수 있는 과립의 형태로 활성 화합물을 함유할 수 있다. 연질 캡슐에서, 활성 화합물은 바람직하게는 지방 오일 또는 액상 파라핀과 같은 적절한 액체 중에 용해되거나 또는 현탁된다.
비경구 투여에 적합한 제제에는 수용형 형태 예컨대, 수용성 염 및 알칼리성 용액 중에 활성 화합물을 함유하는 수용액이 포함된다. 알칼리성 염에는 예를 들면, 트리스, 콜린 히드록사이드, 비스-트리스 프로판, N-메틸글루카민 또는 아르기닌으로 제조된 암모늄 염이 포함될 수 있다. 추가로, 적절한 유성 주사 현탁액으로서의 활성 화합물의 현탁액이 투여될 수 있다. 적절한 동결건조 용매 또는 비히클에는 지방 오일, 예컨대, 참께유, 또는 합성 지방 에스테르, 예컨대, 에틸 올레에이트 또는 트리글리세라이드 또는 폴리에틸렌 글리콜-400(화합물은 PEG-400 중에서 가용성을 나타냄)이 포함된다. 수성 주사 현탁액은 현탁액의 점도를 증가시키는 물질, 예컨대, 나트륨 카르복시메틸 셀룰로스, 소르비톨 및/또는 덱스트란을 함유할 수 있다. 경우에 따라, 상기 현탁액은 안정화제도 함유할 수 있다.
본 발명의 화합물은 동물 및 인간의 눈에 소적으로서 투여될 수 있거나, 연고, 겔, 리포좀, 생체적합성 중합체 디스크 또는 펠렛 내에 함유되어 투여될 수 있거나, 콘택트 렌즈 내에 보유될 수 있다. 안구내 투여용 조성물은 당업자가 통상적인 기준을 이용하여 선택할 수 있는 것과 같은 생리학적으로 상용가능한 안포(ophthalmic vehicle)를 함유할 수도 있다. 상기 안포는 물, 폴리에틸렌 글리콜 400과 같은 폴리에테르, 폴리비닐 알코올과 같은 폴리비닐, 포비돈, 카르복시메틸셀룰로스, 메틸셀룰로스 및 히드록시프로필메틸셀룰로스와 같은 셀룰로스 유도체, 광유 및 백색 광유와 같은 석유 유도체, 라놀린과 같은 동물 지방, 땅콩유와 같은 식물성 지방, 카르복시폴리메틸렌 겔과 같은 아크릴산의 중합체, 덱스트란 및 글리코스아미노글리칸과 같은 폴리사카라이드, 및 염화나트륨, 염화칼륨, 염화아연, 중탄산나트륨 또는 락트산나트륨과 같은 완충제를 포함하나 이들로 한정되지 않는 공지된 안포로부터 선택될 수 있다. 고 분자량 물질도 사용될 수 있다. 조성물 중에서 본 발명의 화합물을 불활성화시키지 않는 생리학적으로 상용가능한 보존제에는 클로로부탄올, 벤즈알코늄 클로라이드 및 EDTA와 같은 알코올, 또는 당업자에게 공지된 임의의 다른 적절한 보존제가 포함된다.
피하 투여에 적합한 항체의 동결건조 제제는 그 개시내용이 본 명세서에 참고로 완전히 도입되는 미국 특허 제6,267,958호에 기재되어 있다. 이러한 동결건조 제제는 적절한 희석제로 높은 단백질 농도까지 재구성될 수 있고 재구성된 제제는 본원에서 치료하고자 하는 환자에게 피하 투여될 수 있다.
또한, ανβ6-결합 리간드는 예를 들면, 코아세르베이션(coacervation) 기법 또는 계면 중합에 의해 제조된 마이크로캡슐, 예를 들면, 콜로이드성 약물 전달 시스템(예컨대, 리포좀, 알부민 미소구, 마이크로에멀젼, 나노-입자 및 나노캡슐) 또는 마크로에멀젼 중의 히드록시메틸셀룰로스 또는 젤라틴-마이크로캡슐 및 폴리-(메틸메타크릴레이트) 마이크로캡슐 내에 포획될 수 있다. 이러한 기법은 문헌[Remington's Pharmaceutical Sciences 16th edition, Osol, A. Ed. (1980)]에 개시되어 있다.
ανβ6-결합 리간드의 서방 제제를 제조할 수 있다. 서방 제제의 적합한 예에는 ανβ6-결합 리간드를 함유하는 고체 소수성 중합체로 구성되며 성형 제품 예컨대, 필름 또는 마이크로캡슐의 형태인 반투과성 매트릭스가 포함된다. 서방성 매트릭스의 예에는 폴리에스테르, 히드로겔(예컨대, 폴리(2-히드록시에틸-메타크릴레이트), 또는 폴리(비닐알코올)), 폴리락티드(미국 특허 제3,773,919호), L-글루탐산과 γ-에틸-L-글루타메이트의 공중합체, 비-분해성 에틸렌-비닐 아세테이트, 분해성 락트산-글리콜산 공중합체 예컨대, LUPRON DEPOT™(락트산-글리콜산 공중합체 및 류프롤리드 아세테이트로 구성된 주사가능한 미소구), 및 폴리-D-(-)-3-히드록시부티르산이 포함된다.
생체 내 투여에 사용될 제제는 멸균되어야 한다. 이는 멸균 여과 막을 통한 여과에 의해 용이하게 달성된다.
ανβ6-결합 리간드는 비경구, 폐내, 두개내, 경피 및 비강내를 비롯한 임의의 적합한 경로를 통해 대상체 또는 환자에게 투여될 수 있다. 비경구 관주에는 근육내, 정맥내, 동맥내, 복강내 또는 피하 투여가 포함된다. 또한, ανβ6-결합 리간드는 예컨대, 점차 감소하는 투여량의 ανβ6-결합 리간드를 사용한 펄스 관주에 의해 적절하게 투여될 수 있다. 바람직하게는, 주사, 가장 바람직하게는 정맥내 또는 피하 주사로 투여되고, 이는 부분적으로는 투여가 단기 투여인지 아니면 장기 투여인지에 달려 있다.
본 발명의 일부 예시적 실시양태에서, ανβ6-결합 리간드는 약 1 mg/m2 내지 약 500 mg/m2의 투여량으로 환자에게 (예컨대, 정맥내로) 투여된다. 예를 들면, ανβ6-결합 리간드는 약 1 mg/m2, 2 mg/m2, 3 mg/m2, 4 mg/m2, 5 mg/m2, 10 mg/m2, 15 mg/m2, 20 mg/m2, 25 mg/m2, 30 mg/m2, 35 mg/m2, 40 mg/m2, 45 mg/m2, 50 mg/m2, 55 mg/m2, 60 mg/m2, 65 mg/m2, 70 mg/m2, 75 mg/m2, 80 mg/m2, 85 mg/m2, 90 mg/m2, 95 mg/m2, 100 mg/m2, 105 mg/m2, 110 mg/m2, 115 mg/m2, 120 mg/m2, 125 mg/m2, 130 mg/m2, 135 mg/m2, 140 mg/m2, 145 mg/m2, 150 mg/m2, 155 mg/m2, 160 mg/m2, 165 mg/m2, 170 mg/m2, 175 mg/m2, 180 mg/m2, 185 mg/m2, 190 mg/m2, 195 mg/m2, 200 mg/m2, 205 mg/m2, 210 mg/m2, 215 mg/m2, 220 mg/m2, 225 mg/m2, 230 mg/m2, 235 mg/m2, 240 mg/m2, 245 mg/m2, 250 mg/m2, 255 mg/m2, 260 mg/m2, 265 mg/m2, 270 mg/m2, 275 mg/m2, 280 mg/m2, 285 mg/m2, 290 mg/m2, 295 mg/m2, 300 mg/m2, 305 mg/m2, 310 mg/m2, 315 mg/m2, 320 mg/m2, 325 mg/m2, 330 mg/m2, 335 mg/m2, 340 mg/m2, 345 mg/m2, 350 mg/m2, 355 mg/m2, 360 mg/m2, 365 mg/m2, 370 mg/m2, 375 mg/m2, 380 mg/m2, 385 mg/m2, 390 mg/m2, 395 mg/m2 또는 400 mg/m2의 투여량으로 투여될 수 있다 .
ανβ6-결합 리간드는 매우 다양한 투여 스케쥴에 따라 투여될 수 있다. 예를 들면, ανβ6-결합 리간드는 정해진 기간 동안(예컨대, 4 내지 8주, 또는 그 이상의 기간 동안) 매일 1회씩 투여될 수 있거나, 정해진 기간 동안(예컨대, 4 내지 8주, 또는 그 이상의 기간 동안) 주 단위 스케쥴(예를 들면, 1주당 1일, 1주당 2일, 1주당 3일, 1주당 4일, 1주당 5일, 1주당 6일 또는 1주당 7일)에 따라 투여될 수 있다. "1주당 1회" 투여 스케쥴의 구체적인 예는 치료 기간의 1일째 날, 8일째 날 및 22일째 날에 ανβ6-결합 리간드를 투여하는 것이다. 대안적 실시양태에서, ανβ6-결합 리간드는 수개월의 기간에 걸쳐 간헐적으로 투여될 수 있다. 예를 들면, ανβ6-결합 리간드는 2년마다 연속 3주 동안 주마다 투여될 수 있다(즉, 6개월마다 주 단위 투여 스케쥴을 반복함). 이러한 투여 섭생법이 연장된 기간 동안(대략 수년 동안) 계속 수행되어 초기 치료에 의해 제공되는 유익한 치료 효과를 유지할 수 있다. 다른 실시양태에서, 이러한 유지 요법은 암성, 전이성 또는 상피내 암종 병태의 즉각적인 증상을 감소시키도록 디자인된 긴급 투약 섭생법 후에 수행될 수 있다.
치료기간 전체에 걸쳐 각 시점에서 투여되는 ανβ6-결합 리간드의 양은 동일할 있거나, 치료기간 동안 각 시점에서 투여되는 양은 다를 수 있다(예를 들면, 소정의 시점에서 투여되는 양은 앞서 투여된 양보다 많거나 적을 수 있음). 예를 들면, 유지 요법 동안에 투여되는 투여량은 치료의 긴급 주기 동안에 투여된 양보다 적을 수 있다. 특정한 환경에 달려 있는 적절한 투여 스케쥴은 당업자에게 자명할 것이다.
본 발명의 일부 실시양태에서, 다양한 유형 또는 종의 ανβ6-결합 리간드는 서로 조합되고 환자에게 투여되어 하나 이상의 암성, 전이성 또는 상피내 암종 병태를 치료한다. 예를 들면, 본 발명은 둘 이상의 상이한 ανβ6-결합 항체 예컨대, 본 명세서에 개시된 항체를 환자에게 투여하는 것을 포함한다. 다수의 ανβ6-결합 리간드가 환자에게 투여되는 경우, 상이한 ανβ6-결합 리간드는 단일 약학 조성물로 함께 투여될 수 있거나, 보다 바람직하게는, 별도의 제제로 순차적으로 투여될 수 있다. 이러한 다른 약제의 유효량은 제제에 존재하는 ανβ6-결합 리간드의 양, 질환 또는 장애 또는 치료의 종류 및 다른 인자에 달려 있다.
또한, 본 발명은 제2 약제와 함께 제1 약제를 환자에게 투여하는 단계를 포함하는, 암성, 전이성 또는 상피내 암종 병태의 치료 방법을 포함하며, 이때 상기 제1 약제는 ανβ6-결합 리간드이고, 상기 제2 약제는 하나 이상의 암성, 전이성 또는 상피내 암종 병태를 치료하는 데 유용하나 반드시 ανβ6-결합 리간드인 것은 아닌 약제이다. 제2 약제"와 함께" 제1 약제를 투여한다는 것은 상기 두 약제가 치료 섭생법 동안 환자에게 투여되도록 제2 약제를 환자에게 투여하기 전, 제2 약제를 환자에게 투여함과 동시에, 또는 제2 약제를 환자에게 투여한 후에 제1 약제를 환자에게 투여할 수 있다는 것을 의미한다. 예를 들면, 본 발명의 이러한 일부 실시양태에 따르면, ανβ6-결합 리간드는 당분야에 공지되어 있는 하나 이상의 인테그린 수용체(예컨대, α1β1, α4β1, ανβ8, ανβ5, α5β1 등)에 특이적인 항체, 폴리펩티드 길항제 및/또는 소분자 길항제를 비롯한 하나 이상의 다른 인테그린 수용체(예컨대, α1β1, α4β1, ανβ8, ανβ5, α5β1 등)의 길항제의 투여와 함께(즉, 투여 전, 투여와 동시 또는 투여 후) 환자에게 투여된다.
본 발명의 이 양태의 일부 실시양태에서, ανβ6-결합 리간드와 함께 투여되는 제2 약제는 예컨대, 스테로이드, 세포독성 화합물(본 명세서의 다른 곳에 기재된 것들을 포함함), 방사성 동위원소(본 명세서의 다른 곳에 기재된 것들을 포함함), 프로드러그-활성화 효소(본 명세서의 다른 곳에 기재된 것들을 포함함), 콜치신, 산소, 항산화제(예컨대, N-아세틸시스테인), 금속 킬레이터(예컨대, 테트라티오몰리브데이트), IFN-β, IFN-γ, 알파-안티트립신 등이다. 본 발명의 이 양태에 따른 치료 목적을 위해 하나 이상의 제1 약제, 예컨대, 하나 이상의 ανβ6-결합 리간드와 함께 환자에게 투여될 수 있는 추가 제2 약제 또는 화합물은 당업자에게 친숙할 것이므로, 이러한 추가 제2 약제 또는 화합물의 사용은 본 발명에 포함되는 것으로 간주된다.
당업자는 본 발명의 범위 또는 본 발명의 임의의 실시양태를 벗어나지 않으면서 본 명세서에 기재된 방법 및 용도에 대한 다른 적절한 변형 및 개조를 행할 수 있다는 것을 용이하게 인식할 것이다. 본 발명은 지금부터 상세히 기재되며, 하기 실시예를 참고함으로써 보다 명백히 이해될 것이고, 하기 실시예는 본 발명을 설명하기 위한 목적으로 본 명세서에 포함되는 것이지 본 발명을 제한하기 위한 것이 아니다.
[실시예]
실시예 1: mu3G9 가변 영역의 클로닝
3G9 뮤린 하이브리도마 세포로부터 얻은 총 세포내 RNA를 제조자의 권장 프로토콜에 따라 퀴아젠 RNeasy 미니 키트를 사용하여 준비하였다. 중쇄 및 경쇄의 가변 영역을 코딩하는 상보적 DNA는 제조자의 권장 프로토콜에 따라 아마샴/파마샤(Amersham/Pharmacia) 제1 가닥 cDNA 합성 키트 및 프라이밍용 무작위 헥사머를 사용하여 총 세포내 RNA로부터 RT-PCR로 클로닝하였다.
뮤린 3G9 면역글로불린 중쇄 가변 도메인의 PCR 증폭을 위해 사용된 프라이머는 다음과 같았다:
5' AGGTCTAGAAYCTCCACACACAGGRRCCAGTGGATAGAC 3' (서열번호 8);
5' GGGGATATCCACCATGRACTTCGGGYTGAGCTKGGTTTT 3' (서열번호 9); (S=C/G, M=A/C, R=A/G, K=G/T, W=A/T, 및 Y=C/T).
반응은 95℃에서 2.5분 동안 초기 용융 후, 94℃에서 30초 동안 용융, 사이클 당 45초 동안 60℃ - 1℃에서 어닐링, 및 68℃에서 1분 동안 클론텍 어드벤티지 택(Clontech's Advantage Taq) DNA 중합효소를 사용한 연장으로 이루어진 10 사이클로 구성되었다. 이 반응은 94℃에서 30초 동안 용융, 55℃에서 45초 동안 어닐링, 68℃에서 1분 동안 연장 및 68℃에서 9분 동안 최종 연장으로 이루어진 추가 10 사이클 동안 계속 수행되었다. 반응 생성물을 제조자의 프로토콜에 따라 퀴아젠 퀴아퀵 PCR 정제 키트를 사용하여 정제하였다. 어드밴티지 택에 의해 증폭된 DNA의 말단을 과량의 dNTP의 존재 하에 T7 DNA 중합효소로 중합하여 블런트 말단을 생성하였다. 블런트 말단을 가진 정제된 3G9 중쇄 가변 영역 유전자 PCR 생성물을 제조자의 권장 프로토콜에 따라 인비트로겐(Invitrogen) TOPO 클로닝 키트를 사용하여 인비트로겐 pCR4Blunt-TOPO 클로닝 벡터 내로 서브클로닝하였다. 중쇄 RT-PCR 서브클론은 pKJS062로 명명되었다.
3G9 경쇄 가변 도메인 유전자를 하기 프라이머로 증폭하였다:
5' GCGTCTAGAACTGGATGGTGGGAGATGGA 3' (서열번호 10);
5' GGGGATATCCACCATGGATTTTCAGGTGCAGATTTTCAG 3' (서열번호 11).
반응은 95℃에서 2.5분 동안 초기 용융 후, 94℃에서 30초 동안 용융, 사이클 당 45초 동안 60℃ - 1℃에서 어닐링, 및 68℃에서 2분 동안 클론텍 어드벤티지 택 DNA 중합효소를 사용한 연장으로 이루어진 6 사이클로 구성되었다. 이 반응은 94℃에서 30초 동안 용융, 54℃에서 45초 동안 어닐링, 68℃에서 2분 동안 연장 및 68℃에서 10분 동안 최종 연장으로 이루어진 추가 24 사이클 동안 계속 수행되었다. 반응액의 1/10은 Pfu DNA 중합효소(Stratagene)를 사용한 제2 라운드의 증폭을 위한 주형으로서 사용되었다. 이 반응은 95℃에서 2.5분 동안 초기 용융 후, 94℃에서 30초 동안 용융, 45초 동안 55℃에서 어닐링, 및 72℃에서 1분 동안 연장으로 이루어진 20 사이클로 구성되었다. 반응 생성물은 제조자의 권장 프로토콜에 따라 퀴아젠 퀴아퀵 겔 추출 키트를 사용하여 겔 정제되었다. 정제된 3G9 경쇄 가변 영역 유전자 PCR 생성물을 인비트로겐 TOPO 클로닝 키트를 사용하여 인비트로겐 pCR4Blunt-TOPO 클로닝 벡터 내로 서브클로닝하였다. 경쇄 RT-PCR 서브클론은 pKJS054로 명명되었다.
pKJS054 및 pKJS062 둘다의 다수의 독립적인 서브클론들로부터 얻은 인서트를 시퀀싱하였다. 두 경우에서, 다수의 독립적인 서브클론의 인서트 서열은 동일하였다. 가변 도메인 서열의 블라스트(Blast) 분석은 이 서열이 면역글로불린 서열임을 확인시켜주었다. 3G9 중쇄 가변 도메인은 뮤린 하위군 IIID의 구성원이다. 3G9 경쇄 가변 도메인은 뮤린 카파 하위군 IV의 구성원이다.
실시예 2: ch3G9의 제작 및 발현
중쇄 및 경쇄의 뮤린 3G9 가변 영역을 코딩하는 cDNA를 사용하여 mu3G9 가변 도메인 영역이 인간 IgG1 및 카파 불변 영역에 연결되어 있는 뮤린-인간 키메라(ch3G9)의 발현용 벡터를 제작하였다.
중쇄 키메라를 제작하기 위해, 3G9 중쇄 가변 도메인 플라스미드 pKJS062로부터 얻은 508 bp의 EcoRI 단편을 선형화된 탈인산화 pUC-유래 클로닝 벡터 pNNO9의 EcoRI 부위 내로 서브클로닝하였다. 이 단계는 생성된 플라스미드 pKJS093 내에 플랭킹(flanking) Not1 부위를 부가하였다. 플라스미드 pKJS093 내의 중쇄 서열은 DNA 시퀀싱에 의해 확인되었다. 제조자의 권장 프로토콜에 따라 스트라타진(Stratagene) 퀵체인지 돌연변이유발 키트를 사용하여 하기 돌연변이유발 올리고뉴클레오티드를 사용한 부위 지정 돌연변이유발을 통해 스플라이스(splice) 공여 부위에 이어서 즉시 HindIII 제한 부위를 플라스미드 pKJS093의 가변 영역 코딩 서열의 바로 다운스트림에 부가시켰다: 5' CTGTCTCTGCAGGTAAGCTTACACCCCCATCTG 3' (서열번호 12), 및 5' CAGATGGGGGTGTAAGCTTACCTGCAGAGACAG 3' (서열번호 13). 이 단계는 플라스미드 pKJS116을 생성시켰다. pKJS116으로부터 얻은 0.48 kb NotI-HindIII 중쇄 가변 도메인 단편, 및 플라스미드 pEAG964로부터 얻은, 인간 IgG1 불변 영역을 함유하는 1.22 kb HindIII-Not1 단편을 pCEP4(인비트로겐) EBV 발현 벡터로부터 유도된 플라스미드 pCH269의 Not1 부위 내로 서브클로닝하여 플라스미드 pKJS136을 생성하였다.
경쇄 키메라를 제작하기 위해, 3G9 경쇄 가변 도메인 플라스미드 pKJS054로부터 얻은 474개 염기쌍 EcoRI 단편을 선형화된 탈인산화 클로닝 벡터 pNN09의 EcoRI 부위 내로 서브클로닝하고, 생성된 플라스미드 pKJS112 내에 플래킹 Not1 부위를 부가하였다. 플라스미드 pKJS112 내의 경쇄 서열은 DNA 시퀀싱에 의해 확인되었다. 스트라타진 퀵체인지 돌연변이유발 키트를 사용하여 하기 돌연변이유발 올리고뉴클레오티드를 사용한 부위 지정 돌연변이유발을 통해 BgIII 제한 부위를 플라스미드 pKJS112의 가변 영역 코딩 서열의 바로 다운스트림에 부가하였다: 5' GGCACCAAGCTGGAGATCTAACGGGCTGATGCTGC 3' (서열번호 14), 및 5' GCAGCATCAGCCCGTTAGATCTCCAGCTTGGTGCC 3' (서열번호 15). pKJS132로부터 얻은 453 bp NotI-BgIII 경쇄 가변 도메인 단편, 및 인간 카파 경쇄 불변 도메인을 함유하는 플라스미드 pEAG963으로부터 얻은 678 bp BcII-Not1 단편을 pCEP4(인비트로겐) EBV 발현 벡터로부터 유도된 플라스미드 pCH269의 Not1 부위 내로 서브클로닝하여, 플라스미드 pKJS141을 생성하였다. mu3G9의 클로닝 과정 도중에 경쇄의 첫 번째 CDR이 글리코실화 신호 서열(NXT/S)을 함유한다는 것이 발견되었다. 올리고뉴클레오티드를 사용한 퀵체인지 부위 지정 돌연변이유발의 단일 라운드, NSS 모티프의 SSS로의 전환 및 발현 벡터 pKJS157의 생성은 이 글리코실화 신호 서열을 제거하였다: 5' GGAACTTACACTTGAGCTGGCACTGCATGTCAAGG 3' (서열번호 16), 및 5' CCTTGACATGCAGTGCCAGCTCAAGTGTAAGTTCC 3' (서열번호 17). pKJS157의 경쇄 가변 영역 서열은 DNA 시퀀싱에 의해 확인되었다.
발현 벡터들(경쇄 pKJS141 또는 pKJS157 및 중쇄 pKJS136)을 293-EBNA 세포 내로 동시-형질감염시키고 형질감염된 세포를 항체 분비 및 특이성에 대해 시험하였다. 빈 벡터로 형질감염된 세포 및 chM92(분자적으로 클로닝된 CD154-특이적 mAb)에 대한 EBV 발현 벡터들로 동시-형질감염된 세포를 대조군으로서 사용하였다. 제조자의 권장 프로토콜에 따라 피어스 이지 타이터 키트(Pierce Easy Titer kit)를 사용한 항체 역가 분석, 및 컨디셔닝된(conditioned) 배지로부터의 웨스턴 블롯 분석(항-인간 중쇄 및 경쇄 항체로 현상함)은 ch3G9-형질감염 세포가 중쇄 및 경쇄를 합성하고 효율적으로 조립하여 항체를 분비한다는 것을 보여주었다. ανβ6에 대한 ELISA 분석은 ch3G9가 mu3G9와 유사하게 ανβ6에 결합하지만 chM92는 ανβ6에 결합하지 않는다는 것을 입증하였다.
도 1에 나타낸 바와 같이, 대규모 일시적 형질감염으로부터 생성된 키메라 3G9(ch3G9; 삼각형 기호로 표시됨), 및 경쇄의 첫 번째 CDR 내의 N-결합 글리코실화 부위 내에 N에서 S로의 치환을 함유하는 키메라 3G9의 비글리코실화 돌연변이체 형태가 정제되었고 ELISA 분석에서 ανβ6과의 유사한 결합을 보였다. 경쇄 가변 도메인의 CDR1 내의 글리코실화 부위의 제거는 항체의 결합 친화성을 변경시키거나 영향을 주지 않고 단백질 발현 및 정제를 개선하는 것으로 밝혀졌다.
실시예 3: hu3G9 버젼 1, 2 & 3의 제작
인간화 3G9(hu3G9)를 생성하기 위한 재구성된 가변 도메인의 디자인은 다음과 같이 수행하였다. 3G9 경쇄 가변 도메인은 인간 카파 3에 상응하고, 중쇄 가변 도메인은 인간 중쇄 하위군 3에 상응한다. 인간 수용 골격은 프로그램 IgBLAST를 이용하여 인간 생식세포 서열과의 상동성 일치에 의해 선택되었다: 경쇄에 대해 인간 L6(인간 JK4로부터 유래된 J 영역을 가짐), 및 중쇄에 대해 인간 3 내지 7(인간 JH4로부터 유래된 J 영역을 가짐). 재구성된 가변 경쇄 및 중쇄 각각의 3가지 버젼을 표 1에 나타낸 바와 같이 디자인하였다(뮤린 중쇄 = 서열번호 81; 3G9HV1 = 서열번호 82; 3G9HV2 = 서열번호 83; 3G9HV3 = 서열번호 84; VH3-7 = 서열번호 85; 뮤린 경쇄 = 서열번호 86; 3G9LV1 = 서열번호 87; 3G9LV2 = 서열번호 88; 3G9LV3 = 서열번호 89; 3G9LV4 = 서열번호 90; 3G9LV5 = 서열번호 91 및 L6 = 서열번호 92). 제1 버젼은 뮤린 공여 서열로의 복귀 돌연변이를 가장 많이 함유하지만, 제3 버젼은 가장 적은 복귀 돌연변이를 함유한다(즉, 가장 "인간화됨"). 하기 표 1에 나타낸 바와 같은 중쇄 및 경쇄 가변 도메인의 CDR 영역은 보편적인 카바트 넘버링 분류 시스템에 의해 한정되어 있다. 그러나, 상기 서열의 넘버링은 서로에 대해 상이한 서열의 상대적인 선형 위치결정(linear positioning)을 기초로 하여 아래에 표시하였다.
[표 1]
hu3G9 중쇄의 버젼 1 및 2, 및 경쇄의 버젼 1, 2 및 3은 짧은 바닥-가닥 올리고뉴클레오티드와 병렬로 배치되어 있는 인산화된 상부-가닥 올리고뉴클레오티드의 조합물을 Taq DNA 리가제(New England Biolabs)로 연결시킴으로써 합성적으로 제조하였다. 반응액을 94℃에서 1분, 사이클 당 55℃-1℃에서 1분, 65℃에서 4분으로 구성된 15 사이클을 통해 항온처리하여, 5' 제한효소 부위(Not1 및 BamH1), 신호 서열 및 가변 도메인을 포함하며 제1 잠재적 유니크 제한 부위(경쇄에 대해 BsiWI 및 중쇄에 대해 AgeI)까지 (경쇄) 또는 (중쇄) 불변 도메인 내로 연장되어 있는 단일 가닥 주형 DNA를 생성하였다. 합성 유전자에 대한 프라이머는 아래에 기재되어 있다. 유전자 주형은 하기 올리고뉴클레오티드를 사용하여 Pfu DNA 중합효소(Stratagene)를 사용한 PCR로 증폭하여 이중 가닥 DNA를 생성하였다:
중쇄에 대해 5' GCTGACAGCGGCCGCGGGATCCAC 3' (서열번호 18) 및 5' GCTCACGGTCACCGGTTCGGGG 3' (서열번호 19); 및 경쇄에 대해 5' GCTGACAGCGGCCGCGGGATCCAC 3' (서열번호 20) 및 5' GGAAGATGAACACACTGGGTGCGG 3 ' (서열번호 21). 반응은 95℃에서 2.5분 동안 초기 용융 후, 94℃에서 30초 동안 용융, 30초 동안 64℃에서 어닐링, 및 72℃에서 1분 동안 연장으로 이루어진 16 사이클로 구성되었다. 반응 생성물을 제조자의 권장 프로토콜에 따라 퀴아젠 퀴아퀵 겔 추출 키트를 사용하여 겔로부터 정제하였다.
중쇄의 버젼 1은 하기 상부 가닥 5' 인산화 올리고뉴클레오티드로부터 합성적으로 생성되었다:
5'GCTGACAGCGGCCGCGGGATCCACCATGGACTTCGGCCTGAGCTGGGTGTTCCTGGTGCTGGTGCTGAAGGGCGTGCAGTGCGAGGTGATGCTGGTGGAGAGCGGCGGC 3' (서열번호 22),
5'GGCCTGGTGCAGCCCGGCGGCAGCCTGAGGCTGAGCTGCGCCGCCAGCGGCTTCACCTTCAGCCGCTACGTGATGAGCTGGGTGCGCCAGGCCCCCGGCAAGGGCCTGGAGTGGGTGGCCAG 3' (서열번호 23),
5'CATCAGCAGCGGAGGCCGCATGTACTACCCCGACACCGTGAAGGGCCGCTTCACCATCAGCCGCGACAGCGCCAAGAACAGCCTGTACCTGCAGATGAACAGCCTGCGCGCCGAGGAC 3' (서열번호 24),
5'ACCGCCGTGTACTACTGCGCCCGCGGCAGCATCTACGACGGCTACTACGTGTTCCCCTACTGGGGCCAGGGCACCCTGGTGACCGTGAGCTCCGCCAGCACC 3' (서열번호 25),
5'AAGGGCCCCAGCGTGTTCCCCCTGGCCCCCAGCAGCAAGAGCACCAGCGGCGGCACCGCCGCCCTGGGCTGCCTGGTGAAGGACTACTTCCCCGAACCGGTGACCGTGAGC 3' (서열번호 26).
이들 올리고뉴클레오티드는 병렬된 상부 가닥 올리고뉴클레오티드와 대략 15 bp만큼 중첩된 하기 바닥 가닥 비-인산화 올리고뉴클레오티드와 병렬로 배치되었다. 바닥 가닥 올리고뉴클레오티드는 다음과 같다:
5' GCTGCACCAGGCCGCCGCCGCTCTCC 3" (서열번호 27),
5' CCGCTGCTGATGCTGGCCACCCAC 3" (서열번호 28),
5' GCAGTAGTACACGGCGGTGTCCTCGGCGCG 3" (서열번호 29),
5' GCTGGGGCCCTTGGTGCTGGCGG 3" (서열번호 30).
중쇄의 버젼 2는 하기 상부 가닥 5' 인산화 올리고뉴클레오티드로부터 합성적으로 생성되었다:
5'GCTGACAGCGGCCGCGGGATCCACCATGGACTTCGGCCTGAGCTGGGTGTTCCTGGTGCTGGTGCTGAAGGGCGTGCAGTGCGAGGTGCAGCTGGTGGAGAGCGGCGGC 3' (서열번호 31),
5'GGCCTGGTGCAGCCCGGCGGCAGCCTGAGGCTGAGCTGCGCCGCCAGCGGCTTCACCTTCAGCCGCTACGTGATGAGCTGGGTGCGCCAGGCCCCCGGCAAGGGCCTGGAGTGGGTGGCCAG 3' (서열번호 32),
5'CATCAGCAGCGGAGGCCGCATGTACTACCCCGACACCGTGAAGGGCCGCTTCACCATCAGCCGCGACAGCGCCAAGAACAGCCTGTACCTGCAGATGAACAGCCTGCGCGCCGAGGAC 3' (서열번호 33),
5'ACCGCCGTGTACTACTGCGCCCGCGGCAGCATCTACGACGGCTACTACGTGTTCCCCTACTGGGGCCAGGGCACCCTGGTGACCGTGAGCTCCGCCAGCACC 3' (서열번호 34),
5'AAGGGCCCCAGCGTGTTCCCCCTGGCCCCCAGCAGCAAGAGCACCAGCGGCGGCACCGCCGCCCTGGGCTGCCTGGTGAAGGACTACTTCCCCGAACCGGTGACCGTGAGC 3' (서열번호 35).
이들 올리고뉴클레오티드는 병렬된 상부 가닥 올리고뉴클레오티드와 대략 15 bp만큼 중첩된 하기 바닥 가닥 비-인산화 올리고뉴클레오티드와 병렬로 배치되었다. 바닥 가닥 올리고뉴클레오티드는 다음과 같다:
5' GCTGCACCAGGCCGCCGCCGCTCTCC 3" (서열번호 36),
5' CCGCTGCTGATGCTGGCCACCCAC 3" (서열번호 37),
5" GCAGTAGTACACGGCGGTGTCCTCGGCGCG 3" (서열번호 38),
5' GCTGGGGCCCTTGGTGCTGGCGG 3" (서열번호 39).
hu3G9 중쇄의 버젼 1 및 2에 대한 발현 벡터는 합성적으로 발생된 인간화 변이체로부터 얻은 538 bp NotI-AgeI 중쇄 가변 도메인 단편으로서, 인간 IgG1-불변 영역의 처음 105개 bp를 포함하는 단편, 및 플라스미드 pKJS160으로부터 얻은 919 bp AgeI/BamH1 단편으로서, 인간 IgG1 불변 영역의 나머지 부분을 함유하는 단편을, NotI/BamH1으로 절단된 pKJS160(pCEP4(인비트로겐)로부터 유도된 EBV 발현 벡터 pCH269와 동일함) 내로 서브클로닝하여 중쇄 발현 벡터 pKJS166(버젼 1) 및 pKJS167(버젼 2)을 생성함으로써 제조하였다.
중쇄의 버젼 3은 하기 올리고뉴클레오티드를 사용하여 플라스미드 pKJS167에 대한 단일 라운드의 퀵체인지 부위 지정 돌연변이유발을 수행함으로써 생성하였다:
5'CCATCAGCCGCGACAACGCCAAGAACAGCCTG 3' (서열번호 40), 및
5'CAGGCTGTTCTTGGCGTTGTCGCGGCTGATGG 3' (서열번호 41).
생성된 버젼 3 중쇄 플라스미드를 pKJS168로 명명하였다. 생성된 플라스미드 내의 가변 영역 cDNA 서열은 DNA 시퀀싱으로 확인하였다.
경쇄의 버젼 1은 하기 상부 가닥 5' 인산화 올리고뉴클레오티드로부터 합성적으로 생성되었다:
5'GCTGACAGCGGCCGCGGGATCCACCATGGACTTCCAGGTGCAGATCTTCAGCTTCCTGCTGATCAGCGTGAGCGTGATCATGAGCCGCGGCGAGATCGTGCTGACC 3' (서열번호 42),
5'CAGAGCCCCGCCACCCTGAGCCTGAGCCCCGGCGAGAGGGCCACCCTGAGCTGCAGCGCCAGCAGCAGCGTGAGCAGCAGCTACCTGTACTGGTACCAGCAGAAGCC CGGCCAGGCC 3' (서열번호 43),
5'CCCAGGCTGTGGATCTACAGCACCAGCAACCTGGCCAGCGGCGTGCCCGTGCGCTTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCAGCAGCCTGGAGCCCGAGGAC 3' (서열번호 44),
5'TTCGCCGTGTACTTCTGCCACCAGTGGAGCACCTACCCCCCCACCTTCGGCGGCGGCACCAAGGTGGAGATCAAGCGTACGGTGGCCGCACCCAGTGTGTTCATCTTCC 3' (서열번호 45).
이들 올리고뉴클레오티드는 병렬된 상부 가닥 올리고뉴클레오티드와 대략 15 bp만큼 중첩된 하기 바닥 가닥 비-인산화 올리고뉴클레오티드와 병렬로 배치되었다. 바닥 가닥 올리고뉴클레오티드는 다음과 같다:
5' GCGGGGCTCTGGGTCAGCACGATC 3" (서열번호 46),
5' CCACAGCCTGGGGGCCTGGCCG 3" (서열번호 47),
5' GTACACGGCGAAGTCCTCGGGCTC 3" (서열번호 48).
경쇄의 버젼 2는 하기 상부 가닥 5' 인산화 올리고뉴클레오티드로부터 합성적으로 생성되었다:
5'GCTGACAGCGGCCGCGGGATCCACCATGGACTTCCAGGTGCAGATCTTCAGCTTCCTGCTGATCAGCGTGAGCGTGATCATGAGCCGCGGCGAGATCGTGCTGACC 3' (서열번호 49),
5'CAGAGCCCCGCCACCCTGAGCCTGAGCCCCGGCGAGAGGGCCACCCTGAGCTGCAGCGCCAGCAGCAGCGTGAGCAGCAGCTACCTGTACTGGTACCAGCAGAAGCCCGGCCAGGCC 3' (서열번호 50),
5'CCCAGGCTGTGGATCTACAGCACCAGCAACCTGGCCAGCGGCGTGCCCGCCCGCTTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCAGCAGCCTGGAGCCCGAGGAC 3' (서열번호 51),
5'TTCGCCGTGTACTACTGCCACCAGTGGAGCACCTACCCCCCCACCTTCGGCGGCGGCACCAAGGTGGAGATCAAGCGTACGGTGGCCGCACCCAGTGTGTTCATCTTCC 3' (서열번호 52).
이들 올리고뉴클레오티드는 병렬된 상부 가닥 올리고뉴클레오티드와 대략 15 bp만큼 중첩된 하기 바닥 가닥 비-인산화 올리고뉴클레오티드와 병렬로 배치되었다. 바닥 가닥 올리고뉴클레오티드는 다음과 같다:
5' GCGGGGCTCTGGGTCAGCACGATC 3" (서열번호 53),
5' CCACAGCCTGGGGGCCTGGCCG 3" (서열번호 54),
5' GTACACGGCGAAGTCCTCGGGCTC 3" (서열번호 55).
경쇄의 버젼 3은 하기 상부 가닥 5' 인산화 올리고뉴클레오티드로부터 합성적으로 생성되었다:
5'GCTGACAGCGGCCGCGGGATCCACCATGGACTTCCAGGTGCAGATCTTCAGCTTCCTGCTGATCAGCGTGAGCGTGATCATGAGCCGCGGCGAGATCGTGCTGACC 3' (서열번호 56),
5'CAGAGCCCCGCCACCCTGAGCCTGAGCCCCGGCGAGAGGGCCACCCTGAGCTGCAGCGCCAGCAGCAGCGTGAGCAGCAGCTACCTGTACTGGTACCAGCAGAAGCCCGGCCAGGCC 3' (서열번호 57),
5'CCCAGGCTGTGGATCTACAGCACCAGCAACCTGGCCAGCGGCATCCCCGCCCGCTTCAGCGGCAGCGGCAGCGGCACCGACTTCACCCTGACCATCAGCAGCCTGGAGCCCGAGGAC 3' (서열번호 58),
5'TTCGCCGTGTACTACTGCCACCAGTGGAGCACCTACCCCCCCACCTTCGGCGGCGGCACCAAGGTGGAGATCAAGCGTACGGTGGCCGCACCCAGTGTGTTCATCTTCC 3' (서열번호 59).
이들 올리고뉴클레오티드는 병렬된 상부 가닥 올리고뉴클레오티드와 대략 15 bp만큼 중첩된 하기 바닥 가닥 비-인산화 올리고뉴클레오티드와 병렬로 배치되었다. 바닥 가닥 올리고뉴클레오티드는 다음과 같다:
5' GCGGGGCTCTGGGTCAGCACGATC 3" (서열번호 60),
5' CCACAGCCTGGGGGCCTGGCCG 3" (서열번호 61),
5' GTACACGGCGAAGTCCTCGGGCTC 3" (서열번호 62).
hu3G9 경쇄의 버젼 1, 2 및 3에 대한 발현 벡터는 합성적으로 발생된 인간화 변이체로부터 얻은 400 bp NotI/BsiWI 경쇄 가변 도메인 단편, 및 pKJS162로부터 얻은 324 bp BsiWI/BamH1 단편으로서, 인간 면역글로불린 카파 불변 영역을 함유하는 단편을, NotI/BamH1으로 절단된 pKJS161(pCEP4(인비트로겐)로부터 유도된 EBV 발현 벡터 pCH269와 동일함) 내로 서브클로닝하여 만들었다. 생성된 플라스미드는 pKJS172(버젼 1), pKJS173(버젼 2) 및 pKJS174(버젼 3)로 명명되었다.
실시예 4: hu3G9 버젼 1, 2 & 3의 발현 및 버젼 4 및 5의 제작
인간화 및 키메라 중쇄 및 경쇄 발현 벡터의 모든 가능한 조합물을 293-EBNA 세포(16개의 조합물) 내로 동시-형질감염시켰다. 형질감염된 세포를 항체 분비 및 특이성에 대해 시험하였다. 웨스턴 블롯 분석(항-인간 중쇄 및 경쇄 항체를 사용한 검출) 및 컨디셔닝된 배지의 이지 타이터(Pierce) 분석은 hu3G9-형질감염된 세포가 중쇄 및 경쇄를 합성하고 효율적으로 분비하며 발현 수준이 항체 인간화의 증가에 따라 증가하는 것으로 보인다는 것을 보여주었다. 도 2에 나타낸 바와 같이, 경쇄의 버젼 1을 함유하는 변이체는 잘 발현되지 않았지만, 경쇄의 버젼 2를 함유하는 변이체는 더 높은 수준으로 발현되어, 인간화의 증가에 따라 발현이 더 높아지는 경향이 있다는 것을 시사한다.
형질감염된 세포로부터의 컨디셔닝된 배지로 염색된 ανβ6 발현 SW480 세포의 FACS 분석은 3G9 중쇄의 인간화가 ανβ6 발현 세포에 결합하는 항체의 능력에 부정적인 영향을 전혀 주지 않았고, 중쇄의 버젼 3(CDR 이식 버젼)이 버젼 1 및 2에 비해 증가하는 결합 활성을 가진다는 것을 보여주었다(도 3). 또한, FACS 분석은 경쇄의 버젼 3을 함유하는 hu3G9 mAb 변이체가 hu3G9 경쇄 버젼 2를 함유하는 변이체보다 다소 덜 결합하였지만, 상기 두 버젼이 적어도 키메라 3G9만큼 결합한다는 것을 입증하였다(도 3). 경쇄 버젼 2와 CDR 이식 3G9 사이에는 2개의 아미노산 차이만 존재하였고 경쇄 버젼 3과 CDR 이식 3G9 사이에는 1개의 아미노산 차이만 존재하였다. 이들 경쇄 버젼이 키메라 3G9와 ανβ6의 결합 활성과 유사한 결합 활성을 가지기 때문에, 2개의 추가 버젼은 결합 활성이 개선되는지 아니면 CDR 이식 버젼이 기능성을 가지는지를 확인하기 위해 만들었다.
버젼 4는 경쇄의 위치 1에 존재하는 글루타민의 글루탐산으로의 치환 효과를 조사하였고, 버젼 5는 완전한 CDR 이식 3G9 경쇄이다(표 1). 이 변화들 각각의 개별적인 기여를 조사하기 위해, 새로운 경쇄 발현 벡터를 제작하였다. 플라스미드 pKJS186, 즉 버젼 3 경쇄의 E1Q 변이체는 스트라타진의 퀵체인지 돌연변이유발키트를 사용하여 하기 올리고뉴클레오티드를 사용한 플라스미드 pKJS174의 부위 지정 돌연변이유발을 통해 제조하였고 경쇄 버젼 4로 명명된다: 5' GTCAGCACGATCTGGCCGCGGCTCATGATC 3' (서열번호 63), 및 5' GATCATGAGCCGCGGCCAGATCGTGCTGAC 3' (서열번호 64). 플라스미드 pKJS188, 즉 CDR 이식 3G9 경쇄는 하기 올리고뉴클레오티드를 사용한 플라스미드 pKJS174의 부위 지정 돌연변이유발을 통해 제조하였고 경쇄 버젼 5로 명명된다: 5' CCCAGGCTGCTGATCTACAGCACC 3' (서열번호 65), 및 5' GGTGCTGTAGATCAGCAGCCTGGG 3' (서열번호 66). 이 경쇄 버젼들은 시퀀싱하여 확인한 후 중쇄 버젼 2 또는 3과 함께 293-EBNA 세포 내로 동시-형질감염시켰다. FACS 분석은 중쇄 버젼 3 및 경쇄 버젼 5, 즉 완전한 CDR 이식 쌍이 임의의 다른 인간화 변이체 쌍과 동등하거나 더 잘 ανβ6 발현 세포에 결합한다는 것을 보여주었다(도 4). pKJS168과 pKJS188의 조합은 hu3G9 버젼 5(H3/L5)로 명명되었다.
ch3G9 및 hu3G9 버젼 2 내지 5를 사용한 293-EBNA의 동시-형질감염의 스케일을 높이고 컨디셔닝된 배지를 회수하였다. 항체를 단백질 A-세파로스 상에서 정제하고 정제된 mAb를 활성에 대해 분석하였다. ανβ6에의 결합은 세포주 FDCP1-β6에 대한 FACS 분석(도 5), ELISA(도 6) 및 LAP에 대한 바이오티닐화 ανβ6의 차단(도 7)에 의해 결정되었다. 결합 활성의 순위 순서는 버젼 5(H3/L5) > 버젼 3(H3/L3) = ch3G9 = 버젼 2(H2/L2)이었다. LAP에의 ανβ6-매개 FDCP1-β6 세포-부착의 차단은 도 8에 나타나 있다. 생체활성의 순위 순서는 ch3G9 = 버젼 5(H3/L5) = 버젼 2(H2/L2) > 버젼 3(H3/L3)이었다. 버젼 5가 버젼 2보다 더 인간화되었기 때문에, 안정한 CHO 세포주의 생성을 위해 버젼 5를 선택하였다.
hu3G9 중쇄 가변 도메인의 다양한 버젼(버젼 1, 2, 3 및 5) 및 hu3G9 경쇄 가변 도메인의 다양한 버젼(버젼 1 내지 5)의 DNA 및 상응하는 단백질 서열을 표 2에 나타낸다. 중쇄 가변 도메인의 경우, 서열은
(a) VH3-7의 FR1로부터 유래된 인간 FR1;
(b) 뮤린 3G9 CDR1 중쇄 서열;
(c) VH3-7의 FR2로부터 유래된 인간 FR2;
(d) 뮤린 3G9 CDR2 중쇄 서열;
(e) VH3-7의 FR3으로부터 유래된 인간 FR3;
(f) 뮤린 3G9 CDR3 중쇄 서열; 및
(g) 서열 WGQGTLVTVSS(서열번호 151)를 가진 인간 항체들의 대다수에 존재하는 컨센서스 골격 서열로부터 유래된 인간 FR4를 포함한다.
경쇄 가변 도메인의 경우, 서열은
(a) L6의 FR1로부터 유래된 인간 FR1;
(b) 아스파라진(N)에서 세린(S)으로의 아미노산 치환을 가진 뮤린 3G9 CDR1 경쇄 서열;
(c) L6의 FR2로부터 유래된 인간 FR2;
(d) 뮤린 3G9 CDR2 경쇄 서열;
(e) L6의 FR3으로부터 유래된 인간 FR3;
(f) 뮤린 3G9 CDR3 경쇄 서열; 및
(g) 서열 FGGGTKVEIK(서열번호 152)를 가진 인간 항체들의 대다수에 존재하는 컨센서스 골격 서열로부터 유래된 인간 FR4를 포함한다.
[표 2]
hu3G9 가변 도메인의 중쇄 및 경쇄 서열
실시예 5: 야생형 및 비글리코실화-hu3G9 버젼 5에 대한 안정한 CHO 발현 벡터의 제작
전술한 EBV 벡터는 바람직하지 않은 외래 5' 및 3' UTR을 함유한다. 외래 서열이 제거된 hu-3G9 중쇄 버젼 3 및 경쇄 버젼 5 (H3/L5)(버젼 5로 총칭됨)에 대한 안정한 CHO 발현 벡터를 만들었다. 추가로, 발현된 3G9 항체와 Fc 감마 수용체 사이의 잠재적인 상호작용을 제거하기 위해 비글리코실화 중쇄 벡터를 만들었다.
도 9에 나타낸 바와 같이, pKJS188의 723 bp BamH1 단편을 pKJS077의 6188 bp neo-함유 BamH1 절단 벡터 단편과 연결시켜 플라스미드 pKJS195를 발생시킴으로써 경쇄에 대한 안정한 CHO 발현 벡터를 만들었다.
KJS168로부터 얻은 1449 bp BamH1 단편을 pKJS078(PDM-64-02-13)의 6051 bp BamH1 절단 벡터 단편과 먼저 연결시켜 중쇄 코딩 서열을 플랭킹하는 Not1 제한효소 부위를 제거하고 pKJS171을 생성함으로써 중쇄에 대한 안정한 CHO 발현 벡터를 만들었다. pKJS171에 의해 코딩된 중쇄로부터 C-말단 라이신 잔기를 유전적으로 제거하기 위해, pKJS171의 2190 bp BsrG1/Xba1 단편을 pKJS078(PDM-64-02-13)의 2187 bp BsrG1/Xba1 단편으로 치환하여 pKJS189를 생성하였다. 플라스미드 pKJS189는 도 10 나타낸 바와 같이, dhfr-함유 야생형 hu-3G9 안정한 CHO 발현 벡터를 나타낸다. 중쇄의 비글리코실화 형태를 발생시키기 위해, pKJS189의 587 bp Age1/BsrG1 단편을 pCR076의 587 bp Age1/BsrG1 단편으로 치환하여 pKJS196을 생성하였다. 이 벡터는 도 11에 나타낸 바와 같이, 정상 Fc 수용체 결합에 필요한 글리코실화 신호를 제거하는 N319Q 치환을 함유하는 dhfr-함유 비글리코실화 hu-3G9 안정한 CHO 발현 벡터를 나타낸다.
pKJS189, pKJS196 및 pKJS195 내의 BamH1 cDNA 인서트의 DNA 서열을 확인하였다. 발현 벡터들을 CHO 세포 내로 동시-형질감염시키고, 형질감염된 세포를 항체 분비에 대해 시험하였다. 컨디셔닝된 배지의 이지-타이터 (Pierce) 인간 항체 검출 분석은 형질감염된 세포가 도 12에 나타낸 바와 같이 CHO 발현 벡터로부터 중쇄 및 경쇄를 합성하고 효율적으로 분비한다는 것을 보여주었다.
따라서, 항체의 결합 친화성에 영향을 주지 않으면서 hu-3G9 항체에서 변형될 수 있는 2개의 잠재적인 글리코실화 부위가 존재한다: (1) 서열번호 2의 아미노산 잔기 26에 있는 CDR1 영역 내의 hu-3G9 경쇄 가변 도메인 내에서, 아스파라진(N)에서 세린(S)로의 치환이 단백질 발현 및 정제를 개선시키는 글리코실화 부위(이 부위는 경쇄 가변 도메인 서열의 모든 5가지 버젼에서 변형됨)를 제거하며; (2) hu-3G9 중쇄 버젼 3 불변 영역에서, 아스파라진(N)에서 글루타민(Q)으로의 치환이 Fc 수용체 결합에 필요한 글리코실화 부위를 제거한다.
실시예 6: hu3G9 버젼 5를 발현하는 CHO 세포주
hu3G9 버젼 5에 대한 발현 플라스미드 pKJS189, pKJS196 및 pKJS195를 CHO 세포 내로 형질감염시켰다. hu3G9 버젼 5(H3/L5)의 발현 및 비글리코실화화 hu3G9 버젼 5(a-H3/L5)의 발현은 ανβ6에 대한 결합 특이성을 나타내는 항체를 분비하는 형질감염된 세포로부터 관찰되었다.
실시예 7: 8G6 항-α
ν
β
6
항체의 인간화 디자인
인간화 항체의 디자인에서, 상보성 결정 영역(CDR)은 항원에 가장 잘 결합될 것으로 보이는 잔기들을 함유하며 재구성된 항체 내에 보유되어야 한다. CDR은 문헌[Kabat et al., Sequences of Proteins of Immunological Interest. 5th Edition, U.S. Dept. Health and Human Services, U.S. Govt. Printing Office (1991)]에 따른 서열에 의해 정의된다. CDR은 정규 부류에 속하며(Chothia et al., Nature, 342: 877-883 (1989)), 이때 핵심 잔기는 CDR 루프의 구조적 구성을 상당한 정도까지 결정한다. 이 잔기들은 재구성된 항체 내에 거의 항상 보유되어 있다. 중쇄 및 경쇄의 CDR은 하기와 같이 정규 부류로 분류된다:
경쇄
중쇄
L1: 15개의 잔기 부류 4 H1: 5개의 잔기 부류 1
L2: 7개의 잔기 부류 1 H2: 17개의 잔기 부류 2
L3: 9개의 잔기 부류 1 H3: 17개의 잔기 정규 부류 없음
이들 부류에 중요한 정규 잔기들은 하기 표 3에 나타내었다. 모든 정규 잔기들은 룰(rules)에 의해 기재된 바와 같다. 루프 H3에 대한 정규 부류는 없다.
[표 3]
가변 경쇄 및 중쇄는 프로그램 FASTA를 사용하여 마우스 및 인간 하위군(Kabat et al., 1991)에 대한 컨센서스 서열과 비교하였다.
8G6 가변 경쇄는 112개 아미노산 중첩에 있어서 81.250%의 동일성을 가진 마우스 하위군 카파 3의 구성원이고, 8G6 가변 중쇄는 129개 아미노산 중첩에 있어서 71.318%의 동일성을 가진 마우스 하위군 2a의 구성원이다. 8G6 가변 경쇄는 113개 아미노산 중첩에 있어서 65.487%의 동일성을 가진 인간 하위군 카파 4에 상응한다. 8G6 가변 중쇄는 134개 아미노산 중첩에 있어서 58.955%의 동일성을 가진 인간 하위군 1에 상응한다. VH/VL 팩킹 계면 잔기는 (서열번호 4의) 경쇄 내의 아미노산 위치 50에 있는 희귀 F 및 (서열번호 3의) 중쇄 내의 아미노산 위치 39에 있는 희귀 L을 제외하고 보존되어 있다.
가변 영역의 구조의 모델링은 다음과 같이 수행하였다. 경쇄 및 중쇄를 가장 최근의 PDB 데이타베이스의 국소 카피에 대해 정렬하여 경쇄 및 중쇄의 3차원적 모델을 제작하는 데 사용되는 구조적 틀(frame)을 결정하였다. FASTA를 이용하여, 8G6 경쇄가 111개 아미노산 중첩에 있어서 뮤린 N1O Fab와 90.991%의 서열 동일성을 가진 것으로 밝혀졌다(INSN; 2.9Å 해상도). 8G6 중쇄는 126개 아미노산 중첩에 있어서 뮤린 JEL42 Fab와 80.952%의 서열 동일성을 가진 것으로 밝혀졌다(2JEL; 2.5Å 해상도). 완전한 구조적 주형은 2JEL의 중쇄와 INSN의 경쇄를 조합하여 수득하였다. 분자적 모델링 팩키지 모델러(Modeler) 5.0(Accelrys Inc.)을 사용하여, 주형 구조를 이용하여 경쇄 및 중쇄의 3차원적 구조를 구축하였다. 10개의 상동성 모델을 만들었고, 모델러 에너지의 관점에서 가장 좋은 모델을 선택하였다. 사전점검(procheck) 분석은 phi/psi 지도의 허용되지 않는 영역 내에서 잔기가 전혀 없다는 것을 보여주었다.
재구성된 가변 영역을 디자인하기 위해, 항체 골격으로서 사용하기 위한 가장 유사한 인간 발현 항체 서열을 찾기 위한 노력을 하였다. 가장 근접한 발현 서열을 찾기 위해, NCBI NR 데이타베이스, TrEMBL 데이타베이스 및 카바트 데이타베이스에서 가장 상동성이 높은 발현된 인간 골격에 대한 검색을 수행하였다. 중쇄 및 경쇄 서열의 경우, (가려진 CDR 및 가려지지 않은 CDR을 사용한) 두 가지 검색을 수행하였다. 가장 적합한 발현된 서열의 선택은 정규 및 경계 잔기의 서열 확인의 조사 뿐만 아니라 CDR 루프 길이의 유사함에 대한 조사를 포함한다. 항체의 공급원도 결정 요인이다. 종래의 인간화 항체는 배제된다. NCBI NR 및 TrEMBL 데이타베이스 검색의 경우, BLAST를 사용하였고, 카바트 데이타베이스 검색의 경우, FASTA를 사용하였다.
가장 유사한 발현 경쇄는 카바트 데이타베이스에서 발견되었다(Kabat id 026520 AC21B'CL; Ohlin et al., Mol. Immunol., 33 : 47-56 (1996)). 이는 파지-디스플레이로부터의 PCR 증폭된 scFv이지만 골격 영역에서 L6 생식세포주와 100% 동일하다. 중쇄의 경우, NCBI의 NR 데이타베이스로부터의 인간 골격 gi|392715를 선택하였다. 이것은 골격 영역에서 생식세포주 VH1-2와 100% 동일하다. 생식세포주 서열의 데이타베이스(http://www.ncbi.nlm.nih.gov/igblast/)로부터 두 서열을 검색하여 하기 생식세포주를 선택하였다: 경쇄에 대해 L6, 및 중쇄에 대해 1-2.
단일클론 항체의 인간화에 있어서 가장 중요한 절차는 인간 골격 잔기가 마우스 것으로 복귀 돌연변이된 것을 확인하는 것이다. 경험상, 정규 잔기, 경계 팩킹 잔기, 및 결합 부위에 가까운 희귀 뮤린 잔기를 보유하는 것이 특히 중요하다는 것을 알 수 있었다. 또한, CDR 잔기들 중 임의의 잔기의 6Å 내에 위치된 잔기들이 CDR의 구조에 대한 잠재적 효과에 관해 철저히 분석될 필요가 있다.
8G6 가변 경쇄 재구성 쇄의 3가지 버젼 및 8G6 가변 중쇄 재구성 쇄의 3가지 버젼을 디자인하였다. 제1 버젼은 가장 많은 복귀 돌연변이를 함유하며, 제3 버젼은 가장 적은 복귀 돌연변이를 함유한다(즉, 가장 "인간화되어" 있음). 하기 표 4는 인간화 8G6(hu8G6) 항체에 대한 중쇄 및 경쇄 가변 도메인 서열을 보여준다(뮤린 중쇄 = 서열번호 93; 8G6HV1 = 서열번호 94; 8G6HV2 = 서열번호 95; 8G6HV3 = 서열번호 96; VH1-2 = 서열번호 97; 뮤린 경쇄 = 서열번호 98; 8G6LV1 = 서열번호 99; 8G6LV2 = 서열번호 100; 8G6LV3 = 서열번호 134; L6 = 서열번호 135).
[표 4]
hu8G6 중쇄(버젼 1, 2 및 3) 및 경쇄(버젼 1, 2 및 3) 가변 도메인의 다양한 버젼들의 단백질 서열은 하기 표 5에 나타내었다. 중쇄 가변 도메인의 경우, 서열은
(a) VH1-2의 FR1로부터 유래된 인간 FR1;
(b) 뮤린 8G6 CDR1 중쇄 서열;
(c) VH1-2의 FR2로부터 유래된 인간 FR2;
(d) 뮤린 8G6 CDR2 중쇄 서열;
(e) VH1-2의 FR3으로부터 유래된 인간 FR3;
(f) 뮤린 8G6 CDR3 중쇄 서열; 및
(g) NR 데이타베이스로부터의 인간 골격 gi|392715와 100% 동일하며 서열 WGQGTLVTVSS(서열번호 151)를 가진 인간 항체의 대부분에 존재하는 컨센서스 골격 서열로부터 유래된 인간 FR4를 포함한다.
경쇄 가변 도메인의 경우, 서열은
(a) L6의 FR1로부터 유래된 인간 FR1;
(b) 뮤린 8G6 CDR1 경쇄 서열;
(c) L6의 FR2로부터 유래된 인간 FR2;
(d) 뮤린 8G6 CDR2 경쇄 서열;
(e) L6의 FR3으로부터 유래된 인간 FR3;
(f) 뮤린 8G6 CDR3 경쇄 서열; 및
(g) 서열 FGGGTKVEIK(서열번호 152)를 가진 인간 항체의 대부분에 존재하는 컨센서스 골격 서열로부터 유래된 인간 FR4를 포함한다.
다음은 재구성된 가변 경쇄에서의 복귀 돌연변이를 기술하는 것이다:
E1D - 이것은 CDR 구조/항원 결합에 영향을 주는 것으로 밝혀졌다(Kolbinger et al., Protein Eng., 8: 971-980 (1993)). 모델에서, 이것은 CDR L1 및 L3에서 S26, Q27 및/또는 E93의 골격 또는 측쇄와 상호작용할 것이다. 이것은 치환이 보존적 치환이기 때문에 버젼 2 및 3에서 제거된다.
L46F - 이것은 VH/VL 팩킹 경계 잔기이다. 또한, 이것은 CDR-L2 잔기 E55 아래의 우측에 있는 것으로 보인다. 이것은 버젼 3에서 제거된다.
Y49K - 이것은 CDR-L2에 인접하고 이 모델에서 잔기 E55와 상호작용하는 것으로 보인다. 이것은 매우 중요한 복귀 돌연변이일 가능성이 있으므로, 제거되지 않는다.
다음은 재구성된 가변 중쇄에서의 복구 돌연변이를 기술하는 것이다:
A24G - 이는 CDR-H1에 대한 정규 잔기이다. 보존 치환. 버젼 2에서 제거된다.
G26S - 이것은 CDR-H1에 대한 정규 잔기이다. 보존 치환. 버젼 2에서 제거된다.
Q39L - 이것은 팩킹 경계 잔기이다. 이것은 경쇄와 매우 제한된 상호작용을 하므로 버젼 2에서 제거된다. M48I - 이것은 공통된 복귀 돌연변이이다. 이 모델에서, 이것은 CDR-H2에서 Y59 및 F63과 상호작용할 수 있다. 이것은 버젼 3에서 상실되어 있다. V68A - 이 잔기는 CDR-H2 아래에 위치하며 Y59 및 F63과 상호작용할 수 있다.
R72V - 이것은 CDR-H2에 대한 정규 잔기이다. T74K - 이 잔기는 CDR-H2 아래에 위치하며 Y53과 상호작용할 수 있거나 항원과 직접 접촉할 수 있다.
실시예 8: α
ν
β
6
항체 내부이입
세포에 의해 내부이입되는 항체는 암과 같은 특정한 임상적 증상에 대한 이점을 제공하는데, 이는 상기 항체가 독소, 방사성 화합물, 또는 암세포를 선택적으로 표적화하여 암세포의 성장을 억제하는 다른 항암제와 접합될 수 있기 때문이다. 내부이입될 항-ανβ6 항체의 능력은 본 명세서에 참고로 완전히 도입되는 국제특허출원 공개 제WO 03/100033호에 이미 기재되어 있다. 국제특허출원 공개 제WO 03/100033호는 양이온-의존적 단일클론 항체(RGD-함유 리간드 모사체) 예컨대, 6.8G6 및 6.1A8에 대해 내부이입이 관찰된다고 개시하였다. 그러나, 양이온-독립적 mAb 예컨대, 6.3G9, 7.1C5 및 6.4B4에 대해서는 내부이입이 관찰되지 않았다. 세포에 의해 내부이입되는 항체 예컨대, 8G6의 능력은 내부이입 항체를 치료 잔기/물질과 커플링시켜 상기 치료 잔기/물질이 세포 내로 전달되게 할 수 있다는 이점을 제공한다. 예를 들면, 약물 또는 독소 잔기를 8G6 내부이입 항체와 접합할 수 있다. 그러나, 이 동일한 적용이 비-내부이입 항체 예컨대, 3G9에 적용될 수도 있는데, 이때 화학적 잔기는 상기 항체에 접합되어 표적의 세포 표면(예를 들면, 종양 세포 표면)에 전달될 수 있다.
실시예 9: α
ν
β
6
은 원발성 종양에 비해 전이에서 많이 발현된다.
본 실험에서, 본 발명자들은 상피 유래의 다양한 암 및 전이성 병소에서 ανβ6의 발현을 연구하고, ανβ6 mAb를 차단하는 기능이 생체 내에서 ανβ6을 발현하는 종양의 성장을 억제할 수 있다는 것을 확인하고자 하였다. 본 발명자들은 인간 인두 암종, 즉 Detroit62에 대한 본 발명자들의 항-인간 ανβ6 mAb의 시험관 내 및 생체 내 항-종양 활성을 평가하고 이를 TβRII:Fc의 생체 내 항-종양 활성과 비교하였다. 본 발명자들의 데이타는 ανβ6에 대한 인간 암에서의 역할 및 기능-차단 ανβ6 mAb를 사용한 치료적 중재가능성을 지지한다.
A. 재료 및 방법:
면역조직화학적 분석을 위해, 크실렌 및 에탄올 중에서 조직 절편으로부터 파라핀을 제거하고, 증류수 중에서 재수화시킨 후, 0.45% H2O를 함유하는 메탄올 중에 함침시켰다. 조직을 펩신(00-3009, Zymed, San Francisco, CA)과 함께 항온처리하고 아비딘 및 바이오틴(SP-2001; Vector Laboratories, Burlingame, CA)으로 차단하였다. 1차 항체를 0.1% 소 혈청 알부민(BSA)이 함유된 포스페이트-완충 식염수(PBS) 중에서 희석시키고 조직을 4℃에서 밤새 항온처리하였다. 마우스 이종이식 조직 상의 β6을 면역염색하기 위해, 절편을 항-ανβ6 mAb의 인간/마우스 키메라 형태, 즉 2A1(Weinreb, P.H. et al., J. Biol, Chem. 279(17): 17875-17887 (2004)), 및 항-인간 바이오티닐화 2차 항체(PK-6103, Vector Laboratories, Burlingame, CA)와 함께 항온처리하였다. 인간 조직 상의 β6을 면역염색하기 위해, 절편을 뮤린 2A1 및 항-마우스-바이오티닐화 2차 항체(PK-6102, Vector Laboratories)와 함께 항온처리하였다. 아비딘-바이오틴 복합체-호스라디쉬 퍼록시다제(벡터 키트, PK-6102)를 절편에 가하고, 실온에서 30분 동안 항온처리하고, 3,3'-디아미노벤지딘(DAB) 기질을 지시대로 준비하고(SK-4100, Vector Laboratories), 실온에서 5분 동안 절편에 가하였다. 조직 절편을 메이어스 헤마톡실린(Mayer's Hematoxylin)으로 1분 동안 염색하고 물 및 PBS 중에서 린싱하였다.
B. 결과:
1. 전이에서 α
ν
β
6
발현
ανβ6 면역염색을 다양한 종양 전이에 대해 평가하였다. 전이의 78%(43/55)가 양성적으로 면역염색되어 전이의 대부분에 대한 강한 염색을 보였다(도 13A 내지 13F; 도 14A 내지 14I). 이 결과는 두경부, 자궁 및 췌장 종양이 동등한 수준의 발현을 나타내는 것으로 발견되었다는 점에서 양성 면역염색률(%)의 증가가 있는 것으로 밝혀졌다(표 1):
[표 1]
인간 종양에서의 ανβ6 발현(면역조직화학)
2. 자궁내막 종양 및 환자-일치 전이에서의 α
ν
β
6
발현
상기 표 1에 나타낸 바와 같이, ανβ6 면역염색은 검사된 자궁내막 종양의 53%에 대해 양성이었다. 몇몇을 제외하고, 염색은 보다 높은 등급의 종양의 보다 높은 침입성을 가진 영역에서 보다 더 현저하였다. 3가지 원발성 종양 샘플은 림프절 전이와 일치하였다. 이 경우들 중 2가지 경우에서, 면역염색은 일치된 원발성 종양에 비해 림프절 전이에 대해 훨씬 더 높았다(도 15A와 도 15B를 비고하고 도 15C와 도 15D를 비교함). 세 번째 경우, 면역염색은 림프절 전이 및 매칭된 원발성 종양 둘다에 대해 높았다(데이타는 나타내지 않음). 3가지 원발성 종양에서의 ανβ6 양성 종양 상피 염색률(%)은 각각 10%, 20% 및 90%이었지만, 매칭된 전이성 림프절에서의 ανβ6 양성 종양 상피 비율(%)은 각각 80%, 100% 및 100%이었다. 정상 자궁내막에서, 염색은 간헐적인 표면 층 상의 세포 뿐만 아니라 낭(cyst)에 한정되어 있었다.
3. 침입성 인간 유방 종양 샘플에서의 α
ν
β
6
발현
100개 초과의 인간 유방암 샘플 중의 ανβ6의 발현 수준을 재료 및 방법에서 전술한 절차에 따라 면역조직화학적 기법을 이용하여 평가하였다. 관 상피내 암종(DCIS)의 여러 경우에서, ανβ6은 종양을 둘러싸는 근상피에 한정되었고 종양 그 자체에서는 관찰되지 않았다(예컨대, BrCa19, 도 16A 참조). 그러나, 침입성 유방 암종의 여러 경우에서, ανβ6은 종양에서도 발현되었다(예컨대, BrCa23, 도 16B 참조).
유방 조직의 세포 내의 특정 유전자들의 발현이 정상 유방 조직을 DCIS와 같은 비-침입성 종양을 함유하는 유방 조직, 및 침입성 유방 암종을 함유하는 유방 조직과 비교할 때 변경되었다는 증거가 있다(Alinen, M. et al., Cancer Cell 6: 17-32 (2004); Burstein, H.J. et al., N. Engl. J. Med. 350: 1430-1441 (2004)). 이 유전자 발현 변화들 중 다수가 근상피 세포 내에서 검출되었다. (정상 조직 및 DCIS 둘다에서) 근상피 상의 ανβ6 인테그린의 발현은 아마도 국소화된 TGF-β 활성화를 통해 종양 생존을 지지하면서 침입성 종양의 진행을 촉진하는 미세환경을 제공할 가능성이 있다. 침입성 암종으로 진행할 수 있는 DCIS의 생체 내 모델 예컨대, MCF10DCIS.com(Miller, F.R. et al., J. Natl. Canc. Inst. 92: 1185-1186 (2000))은 진행성 유방 종양의 내용물 내에서 ανβ6의 발현을 평가하는 방법을 제공할 것이다. 당업자는 종양의 초기 비-침입성 단계에서 근상피 내의 ανβ6의 발현을 평가할 수 있고, 생체 내에서 침입성 표현형으로 진행할 때의 ανβ6의 발현도 평가할 수 있다. 이 모델은 당업자가 차단 ανβ6 mAb를 사용하여 ανβ6의 기능적 역할을 시험할 수 있게 하며 접합된 항-ανβ6 mAb의 효능도 시험할 수 있게 한다.
4. 인간 췌장 종양 샘플, 환자-일치 전이 및 침입성 췌장 종양의 마우스 이 종이식 모델에서의 ανβ6 발현
상기 표 1에서 나타낸 바와 같이, ανβ6 면역염색은 검사된 췌장 종양의 80%에 대해 양성이었다. 8명의 상이한 환자로부터 얻은 원발성 췌장 종양으로부터의 샘플을 면역조직화학적 기법으로 검사하였을 때, 염색은 고 등급 종양의 침입성 영역에서 현저하였다(도 17A 내지 17C; 18A 내지 18E). 원발성 종양 샘플은 강한 ανβ6 염색을 나타내는 림프절 전이와도 일치하였고(도 17D 내지 17F; 18F 내지 18J), 이는 ανβ6-양성 세포가 원발성 종양 부위로부터 퍼진다는 견해를 지지한다. 정상 췌장(도 17G 내지 17H; 18K 내지 18L)에서, 염색은 간헐적인 표면 층 상의 세포에 한정되어 있었다.
종양 세포 침입에 대한 ανβ6의 영향을 더 조사하기 위해, 본 발명자들은 침입성 인간 췌장 선암종의 모델로서 BxPC-3 마우스 종양 이종이식물을 사용하였다. 멸균 식염수 중에 현탁된 5 x 106 세포/마우스를 0.1 ㎖/마우스의 주사를 이용하여 동물의 측면에 피하 이식하였다(0일). 30일째 날, 확립된 대략 60 내지 100 ㎣의 종양을 가진 마우스를 모든 연구를 위한 3개의 처리군의 각 군에 쌍별로 일치시켰다(PBS; mAb 3G9; 가용성 TGF-β 수용체 II-Ig 융합 단백질(solTGFβRII-Fc)). 시약을 주당 3회 처리 스케쥴에 따라 마우스의 복강내로 투여하였다. 마우스에게 lO mg/kg의 3G9, 2 mg/kg의 solTGFβRII-Fc 또는 PBS(음성 대조군)를 주사하였다. 종양 성장을 주당 2회씩 측정하였고, 종양 부피를 식 [(너비)2 x 길이]/2에 따라 평가하였다. 처리군들로부터 종양을 절제하고, 10% 파라포름알데히드 중에서 고정시키고, 파라핀-포매하고, 비-차단 v6 키메라 mAb 6.2A1을 사용한 면역조직화학적 분석을 위해 절개하였다.
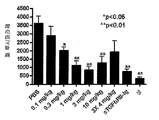
항-ανβ6 mAb 3G9를 사용한 처리는 상기 항체를 사용한 약 48일간의 처리 후 관찰된 종양 성장을 상당히 감소시키면서 종양 성장에 대해 직접적인 효과를 나타내었다(도 19B 및 19C). solTGFβRII-Fc에서 관찰된 성장 억제 수준은 3G9에 대해 관찰된 성장 억제 수준보다 다소 더 낮았다. 이 결과는 항-ανβ6 mAb 3G9는 인간 췌장 암의 이종이식 모델에서 종양 성장을 억제한다는 것을 보여주었고, 이러한 차단 항체가 종양 성장의 억제 및 원발성 인간 췌장 선암종에서의 종양 침입의 확장 억제에 유용할 수 있음을 암시한다.
실시예 10: α ν β 6 기능-차단 mAb는 종양 세포 이동, 침입, 및 α ν β 6 -발현 종양 세포의 MMP 생성을 억제한다.
침입하고, 이동하고, 시험관 내에서 매트릭스 메탈로프로테이나제 9(metalloproteinase-9; MMP-9)를 생성하는 β6-형질감염 세포의 능력을 차단하는 ανβ6 차단 단일클론 항체(mAb) 3G9(Weinreb, P.H. et al., J. Biol. Chem. 279(17): 17875-17887 (2004)) 및 가용성 재조합 TGF-βRII-Ig(Cosgrove, D. et al., Am. J. Pathol. 157(5): 1649-1659 (2000))의 능력을 평가하였다. 종양 세포 침입 및 이동과 독립적으로 관련되어 있는 상기 활성들을 모니터링하였다. 3G9 및 TGF-βRII-Ig의 효과는 형질감염되지 않은 모세포(C1) 및 C1 세포의 β6-형질감염 유도체(VB6)를 사용하여 평가하였다.
이동 및 침입 분석. C1 및 VB6 인간 구강 편평 암종 세포를 전술한 바와 같이 KGM 배지 중에서 증식시켰다(Thomas, G.J. et al., J. Invest. Derm. 117(l): 67-78 (2001). 이동을 측정하기 위해, 본 발명자들은 제조자의 지시에 따라 FLUOROBLOK™ 플레이트 및 인서트(BD Biosciences; Bedford, MA)를 사용하였다. 요약하면, 빈 웰을 음성 대조군으로서 KGM 배지 또는 혈청 무함유 KGM으로 채웠다. 세포를 수거하고 항체와 함께 혈청 무함유 배지 중에서 예비항온처리하였다. 50,000개의 세포를 인서트에 첨가한 후, 이를 웰의 내부에 넣고 조직 배양 항온처리기 중에서 37℃에서 24시간 동안 항온처리하였다. 항온처리 후, 세포 및 배지를 인서트의 상부로부터 제거하였다. 1시간 동안 2 μg/㎖의 칼세인(Calcein)(Invitrogen Corpn., Carlsbad, CA)으로 표지하고 바닥-판독 모드에서 형광도를 측정함으로써 필터 밑면으로 이동한 세포를 정량하였다. 억제율(%)은 배지 단독과 비교할 때 항체의 존재 하에 이동한 세포의 수의 감소로서 계산하였다. 침입은 MATRIGEL®-코팅된 FLUROBLOK 인서트를 사용하여 48시간 동안 항온처리하는 유사한 방식으로 측정하였다.
MMP 생성의 정량화. 1% FBS를 함유하는 배지 중의 세포를 MATRIGEL®-코팅된 웰(BD Biosciences) 중에서 표시된 시간 동안 배양하였다. 상청액을 수거하고 원심분리하여 세포 데브리스를 제거하고 분석할 때까지 동결시켰다. MMP 수준을 ELISA(R&D Systems, Minneapolis, MN)로 정량하였다.
결과:
도 20A 내지 20C에 나타낸 바와 같이, ανβ6-차단 mAb 3G9는 시험관 내에서 이동, 침입, 및 VB6 세포에 의한 MMP-9의 생성을 상당히 억제하였다. 또한, 재조합 가용성 TGF-βRII-Ig를 사용한 TGF-β 활성의 차단은 침입 및 VB6 세포에 의한 MMP-9 생성을 억제하였지만(도 20B 및 20C), 상기 세포의 이동에는 영향을 주지 않았다(도 21A). 이 데이타는 ανβ6 기능의 차단을 TGF-β 활성의 차단과 비교할 때 상이한 기능적 차이를 보여준다. 이 결론은 피브로넥틴과의 결합을 통한 세포 부착 및 이동뿐만 아니라 잠복 전구체 TGF-β의 활성화를 매개하는 ανβ6의 능력과 일치한다(Sheppard, D., Cancer Metast. Rev. 24: 395-402 (2005)).
실시예 11: α ν β 6 mAb는 인간 결장직장 암(LIM1863)의 이종이식 모델에서 간질(stromal) 침입을 억제한다 .
1. 배경
새로운 결장 암종 모델인 LIM1863의 특징은 최근에 규명되었다(Bates, R.C. and Mercurio, A.M., Mol. Biol. Cell 74: 1790-1800 (2003); Bates, R.C. et al., J. Clin. Invest. 115(2): 339-347 (2005)). 시험관 내에서, LIM1863 세포는 잘-분화된 3D 타원체 형태(유선조직절편)로 현탁 배양물 중에서 증식하였다. 그러나, 이 세포주는 TGF-β 및 TNF-α에 노출된 후 E-캐드린의 상실을 전형적인 특징으로 하는, 상피-중간엽 전이(EMT)의 특징적인 형태적 변화를 가지면서 이동 단일층 표현형으로 전환된다. 이 전이는 ανβ6 발현의 상당한 증가에 동반된다. 생체 내에서, LIM1863 세포는 누드 마우스의 측면에 피하 주사될 때 종양을 유발한다(Bates, R.C. et al., J. Clin. Invest. 115(2): 339-347 (2005)). 도 6에 나타낸 바와 같이, LIM1863 세포는 ανβ6 발현의 놀라운 패턴을 나타낸다(상기 문헌). 구체적으로, ανβ6의 발현은 원발성 종양 덩어리로부터 종양 간질 내로 침입하는 것으로 보이는 세포 상에서 특히 현저하다. 이 발견은 이들 세포가 전술한 바와 같은 상피-중간엽 전이(EMT)를 겪는다는 견해와 일치한다(상기 문헌 참조).
그러므로, 본 발명자들은 이 모델에서 간질 침입에 있어서의 ανβ6의 관여 가능성을 조사하기 위해 인간 결장직장 암의 이종이식 모델로서 LIM1863 세포를 사용하는 것을 선택하였다.
2. 재료 및 방법
LIM1863 세포는 5% 태아 소 혈청(FCS)으로 보충된 RPMI-1640(GIBCO; Invitrogen Corp., La Jolla, CA)에서 유선조직절편으로서 성장하였다. LIM1863 유선조직절편(대략 8 x lO6 세포)을 암컷 누드 마우스의 측면 내로 피하 접종하였다. 모든 동물 연구는 바이오겐 아이덱(Biogen Idec)의 동물 보호 및 사용 조직 위원회(IACUC)의 승인을 받았다. 10 mg/kg의 항-ανβ6 특이적 뮤린 단일클론 항체 3G9(Weinreb, P.H. et al., J. Biol. Chem. 279(17): 17875-17887 (2004)), 2 mg/kg의 재조합 가용성 TGF-βRII-Ig 융합 단백질(Cosgrove, D. et al., Am. J. Pathol. 157(5): 1649-1659 (2000)), 또는 비히클 대조군(PBS)을 주당 3회씩 복강내로 주사함으로써 마우스를 처리하였다. 종양 부피를 칼리퍼(caliper)를 사용하여 상응하는 시점에서 측정하였고, 종양 부피를 식 (L x W2)/2을 이용하여 계산하였다. 이종이식물을 7주 후에 수거하여 포르말린으로 고정하였고, 실시예 9에서 전술된 바와 같이 수행된 면역조직화학적 분석을 위해 파라핀-포매 절편을 사용하였다(도 21).
결과
항-ανβ6 mAb 3G9 및/또는 재조합 가용성 TGF-βRII-Ig는 종양 성장에 대해 직접적인 영향을 주지 못하였다(도 22A 및 22B). 그러나, 이 리간드들은 처리에 대해 알지 못하는 병리학자에 의해 수행된 간질 내의 ανβ6 양성 종양 세포 영역의 정량화(도 22D 내지 22F)에 의해 평가될 때 LIM1863 세포의 간질 침입을 대략 80%(도 22C)까지 유의하게 억제하였다.
실시예 12: 비-인간 영장류 세포 상에 발현된 α
ν
β
6
인테그린에 대한 뮤린 6.3G9 (m3G9)의 친화성 및 생체활성
요약. 비-인간 영장류(NHP) ανβ6 인테그린에 대한 항-ανβ6 차단 단일클론 항체 6.3G9(m3G9)의 친화성 및 생체활성을 다양한 시험관 내 방법을 통해 측정하였다. 형광 활성화 세포 분류(FACS)를 이용하여, 높은 수준의 ανβ6을 발현하는 NHP 세포주를 2종의 상이한 종으로부터 확인하였다(아프리칸 녹색 원숭이로부터의 12MBr6, 및 레서스(rhesus) 원숭이로부터의 4MBr5). m3G9는 12MBr6 및 4MBr5 상에 발현된 ανβ6에 각각 0.30 μg/㎖ 및 0.38 μg/㎖의 ED50으로 결합하였다. 또한, m3G9는 12MBr6 및 4MBr5와 TGFβ1 잠복 관련 펩티드(LAP_의 결합을 각각 0.22 μg/㎖ 및 0.29 μg/㎖의 IC50 값으로 억제하였다. 최종적으로, m3G9는 플라스미노겐 활성화제 억제제 1 프로모터(TMLC)의 일부를 안정하게 발현하는 TGFβ 반응성 밍크 폐 상피 레포터 세포를 사용한 동시배양 분석을 이용하여 측정한 때 잠복 TGFβ를 활성화하는 4MBr5 세포주의 능력을 0.31 μg/㎖의 IC50 값으로 차단하였다.
도입. 본 연구의 목적은 비-인간 영장류(NHP) 세포 상에서 발현된 ανβ6 인테그린에 대한 항-ανβ6 단일클론 항체 6.3G9(m3G9)의 친화성 및 생체활성을 측정하기 위한 것이다. m3G9는 인간화 단일클론 항체 hu3G9(BGOOO11)의 뮤린 전구체이다. 이 뮤린 항체는 친화성이 높은 특이적 ανβ6 인테그린-표적화 시약이다1. m3G9는 그의 표적인 ανβ6 인테그린에 높은 친화성(KD = 16 pM)으로 결합하고, 세포에 의해 발현된 ανβ6과 리간드(TGFβ1 잠복 관련 펩티드(LAP) 및 피브로넥틴)의 결합을 억제하고, 잠복 TGFβ1을 활성화는 ανβ6의 능력을 차단한다1. 상기 항체는 결합을 위해 αν 서브유니트 및 β6 서브유니트 둘다의 존재를 필요로 하며, 다른 관련 인테그린(αvβ3, αvβ3, αvβ5, αvβ3, 및 αIIbβ3)과의 교차-반응성은 관찰되지 않았는데, 이는 m3G9가 ανβ6에 대해 높은 특이성을 가짐을 의미한다1.
뮤린 β6 인테그린과 인간 β6 인테그린은 뮤린 αν 인테그린과 인간 αν 인테그린이 고도의 서열 유사성을 가지는 것과 마찬가지로(92.8% 동일성)3 고도의 서열 유사성을 가진다(89.5% 동일성)2. NHP로부터의 ανβ6의 서열은 결정되어 있지 않지만, 고도의 서열 동일성을 공유할 것으로 예측된다.
NHP ανβ6에 대한 m3G9의 친화성을 측정하기 위해, 결합을 FACS로 측정하였다. 인간 TGFβ1 LAP에의 세포 부착을 차단하는 m3G9의 능력은 세포 부착 분석에서 평가되었다. 마지막으로, 잠복 TGFβ의 NHP ανβ6-매개 활성화를 차단하는 m3G9의 능력은 동시배양 분석을 이용하여 측정하였다.
재료 및 방법:
시약. 마우스 단일클론 항체 6.3G9(m3G9) 및 6.4B4는 참고문헌 1 및 상기 기재한 바와 같이 발생시키고 정제하였다. 재조합 인간 LAP(LAP)는 R&D 시스템스(카탈로그 # 246-LP)로부터 구입하였다. 인간 β6-형질감염 SW480(인간 결장직장 선암종) 세포주(SW480β6)는 딘 쉐파드(Dean Sheppard)(UCSF)에 의해 제공되었다.
NHP 세포주. 하기 세포주들을 아메리칸 타입 컬쳐 콜렉션(ATCC)으로부터 입수하였다:
형광-활성화 세포 분류(FACS). 세포를 트립신처리에 의해 수거하고, 인산염 완충 식염수 중에서 한번 세척한 다음, FC 완충제(PBS, 2% FBS, 0.1% NaN3, 1 mM CaCl2, 및 1 mM MgCl2) 중에 재현탁시켰다. 이어서, 1 x 106 세포를 얼음 상의 총 50 ㎕ FC 완충제 중에서 10 μg/㎖의 m3G9와 함께 0.5시간 동안 항온처리하였다. 항온처리 후, 세포를 빙냉 FACS 완충제(PBS, 2% FBS, 0.1% NaN3)로 2회 세척하고, Alexa488-접합 염소 항-마우스 IgG(Jackson ImmunoResearch)의 1:200 희석액을 함유하는 100 ㎕의 FC 완충제 중에 재현탁시키고, 얼음 상에서 30분 동안 항온처리하였다. 그 후, 세포를 빙냉 FC 완충제로 2회 세척하고, 100 ㎕의 FC 완충제 및 50 ㎕의 4% 파라포름알데히드 중에 재현탁시켰다. 표지된 2차 항체의 결합을 유세포분류기(Biogen Idec Research core facility)로 모니터링하였다.
세포 부착 분석. 96-웰 마이크로타이터 플레이트를 4℃에서 50 mM 중탄산나트륨(pH 9.2) 중에 희석된 50 ㎕/웰의 0.5 μg/㎖ LAP로 밤새 코팅하였다. 플레이트를 PBS(100 ㎕/웰)로 2회 세척하고, 25℃에서 PBS(100 ㎕/웰) 중의 1% BSA로 1시간 동안 차단하고, 100 ㎕/웰의 분석 완충제(50 mM 트리스, pH 7.5, 150 mM NaCl, 1 mM CaCl2, 1 mM MgCl2)로 2회 세척하였다. 12MBr6 또는 4MBr5 세포(4 내지 12 X 106 세포/㎖)를 37℃ 수조 중에서 15분 동안 약하게 진탕하면서 분석 완충제 중의 2 μM 형광 염료(BCECF-AM, Molecular Probes)와 함께 항온처리하고, 원심분리하여 모으고, 4 내지 12 X 106 세포/㎖까지 분석 완충제 중에 재현탁시켰다. 세척된 플레이트의 개개의 웰에 25 ㎕의 2x 농축 m3G9 및 25 ㎕의 표지된 세포를 첨가하고, 플레이트를 25℃에서 0.5시간 동안 항온처리하였다. 플레이트를 분석 완충제(100 ㎕/웰)로 4 내지 6회 세척하고, 플레이트 상에 포획된 세포로 인한 형광도를 96-웰 형광도 플레이트 판독기(CytoFluor Series 4000, Perseptive Biosystems) 상에서 기록하였다. 결합률(%)은 최종 세척 단계 전의 형광도(즉, 첨가된 총 세포)를 세척 후의 형광도(즉, 결합된 세포)와 비교함으로써 측정하였다.
동시배양 분석. TMLC(플라스미노겐-활성화제 억제제 1 단백질의 일부로 안정하게 형질감염된 밍크 폐 상피 세포주 Mv 1 Lu)4를 2 mM L-글루타민, 페니실린-스트렙토마이신 및 200 ㎍/㎖의 G418이 함유된 DMEM + 10% 태아 소 혈청 중에서 증식시켰다. 세포를 PBS + 5 mM EDTA를 사용하여 플라스크로부터 걷어내고, PBS + 0.1% BSA로 세척하고, 혈구계로 세포의 수를 세고 96-웰 플레이트에 플레이팅하였다. TMLC가 DMEM + 0.1% FBS 중의 104 세포/웰로 96-웰 플레이트에 플레이팅되어 있는 동안 ανβ6-발현 세포를 얼음 상에서 2시간 동안 저장하고, 37℃에서 부착되게 한 후, 결합된 TMLC를 DMEM + 0.1% BSA로 1회 세척하였다. 단일클론 항체를 ανβ6-발현 세포에 첨가된 DMEM + 0.1% BSA 중에서 희석하고, 실온에서 20분 동안 예비항온처리하였다. 이어서, ανβ6-발현 세포를 4 x 104/웰로 DMEM + 0.1% BSA(100 ㎕/웰) 중의 TMLC에 첨가하였다. 플레이트를 37℃의 습한 CO2-풍부 항온처리기 내에서 20시간 동안 항온처리하였다. 상청액을 버리고 100 ㎕ PBS + 1 mM Ca+2 및 1 mM Mg+2로 교체하였다. 세포를 용해시키고 루시퍼라제를 마이크로플레이트 발광측정기(luminometer)(Tropix TR717 microplate luminometer, Perkin Elmer Life Sciences)를 사용하여 LucLite 키트(Perkin Elmer Life Sciences, Boston, MA)로 검출하였다.
결과:
1. 영장류 α ν β 6 인테그린에 대한 마우스 6.3G9(m3G9)의 결합 친화성 및 차단 효능 . 비-인간 영장류(NHP)ανβ6 인테그린에 대한 m3G9의 친화성을 연구하기 위해, 4가지 NHP 세포주를 ATCC로부터 입수하였다. 초기 스크린(도 23)은 ανβ6 발현이 12MBr6 및 4MBr5 세포주에 대해 최대이었기 때문에, 이들 두 세포주를 m3G9의 특징규명에 사용하였다. ανβ6 비-차단 항체인 6.4B4 또한 m3G9에 대해 관찰된 평균 형광도와 유사한 평균 형광 강도로 12MBr6 및 4MBr5에 결합하였다.
1.1 세포-발현 인테그린에의 결합(FACS) . m3G9와 12MBr6 및 4MBr5의 결합은 재료 및 방법 단락에 기재된 바와 같은 형광-활성화 세포 분류(FACS)를 이용하여 측정하였다. m3G9의 전체 적정을 각 세포주에 대해 수행하여, ED50(절반-최대 신호를 제공하는 항체의 농도)을 비-선형 회귀를 이용하여 측정하였다. 12MBr6 및 4MBr5에 대한 ED50 값은 각각 0.30 μg/㎖(도 24A) 및 0.38 μg/㎖(도 24B)이었다.
1.2 LAP에의 세포 부착 . LAP에 결합하는 12MBr6 및 4MBr5의 능력은 세포 부착 분석을 이용하여 입증하였다. 이 부착은 m3G9에 의해 차단되었지만, 대조군 단백질(BSA) 또는 αvβ6 비-차단 항체(6.4B4)에 의해서는 차단되지 않았다(도 25). 이 상호작용을 차단하는 m3G9의 효능은 각 세포주에 대한 억제의 농도 의존도를 측정함으로써 평가하였다(도 26). 이 실험들로부터, IC50(절반-최대 억제를 제공하는 항체의 농도)을 측정하였다. 12MBr6 및 4MBr5에 대한 IC50 값은 각각 0.22 μg/㎖ 및 0.29 μg/㎖이었다.
1.3 TGFβ 활성화(동시배양 분석). 잠복 TGFβ를 활성화시키는 12MBr6 및 4MBr5의 능력은 플라스미노겐 활성화제 억제제 1 프로모터의 일부를 안정하게 발현하는 TGFβ 반응성 밍크 폐 상피 레포터 세포(TMLC)와 함께 12MBr6 및 4MBr5 세포를 항온처리하는 동시배양 분석을 이용하여 측정하였다(도 27). αvβ6의 발현은 TGFβ 활성화를 촉진하는 데 충분하지 않은데, 이는 잠복 TGFβ의 세포내 신호전달 및/또는 생성의 차이 또한 이 분석에서 활성에 영향을 줄 것이기 때문이다. 이들 두 세포주 중, 12MBr5는 영향을 나타내지 않고 4MBr5만이 잠복 TGFβ를 활성화시키는 데 효과적이었다. 활성화는 인간 αvβ6을 안정하게 발현하는 β6-형질감염 인간 결장 암종 세포주인 양성 대조군 세포주 SW480β6을 사용한 경우에도 관찰되었다. 형질감염되지 않은 모 대조군 세포주(SW480)는 예상대로 활성화를 전혀 보이지 않았다.
4MBr5에 의한 TGFβ 활성화를 차단하는 m3G9의 능력은 동일한 실험에서 측정되었다. TGFβ 활성화는 1 μg/㎖ 또는 10 μg/㎖의 m3G9의 첨가에 의해 차단되었으나, 비-차단 항-αvβ6 항체 6.4B4의 첨가에 의해서는 차단되지 않았다. 상기 농도에서 4MBr5 세포에 대한 m3G9의 억제 효과는 SW480β6 세포주를 사용하였을 때 관찰된 것과 유사하였다. 별도의 실험에서, 4MBr5-매개 TGFβ 활성화의 m3G9 억제의 투여량 의존도를 측정하였다(도 28). 4MBr5 및 SW480β6에 대한 IC50 값은 각각 0.31 μg/㎖ 및 0.37 μg/㎖이었다.
2. 종 사이의 m3G9 친화성의 비교. 마우스, NHP 및 인간 ανβ6에 대한 m3G9의 결합 및 활성에 대해 얻은 데이타는 하기 표에 요약되어 있다. 하기 표에서 첫 번째 컬럼은 FACS에 의해 측정된 결합 ED50을 보여준다. 각 경우에서, 측정된 ED50은 ≤ 0.38 μg/㎖(2.5 nM)이었다. 하기 표에서 두 번째 컬럼은 m3G9에 의한 세포 부착의 억제에 대한 IC50을 보여준다. 각 경우에서, m3G9는 ≤ 0.29 μg/㎖(1.9 nM)의 IC50으로 세포 부착을 차단하였다. 하기 표의 세 번째 컬럼은 동시배양 분석을 이용하여 측정한, ανβ6-매개 TGFβ 활성화의 억제에 대한 IC50을 보여준다. NHP 세포주 4MBr5는 인간 세포주 SW480β6과 유사한 활성을 보였다(각각 0.40 μg/㎖ 및 0.24 μg/㎖의 IC50 값).
실시예 13: 알포트 (Col4A3-/-) 마우스에서의 뮤린 항-α
ν
β
6
mAb의 효과
요약: ανβ6은 상피 리모델링의 부위에서 우세하게 발현되며 잠복 전구체 TGF-β에 결합하여 이를 활성화시키는 것으로 밝혀진 TGF-β-유도가능성 인테그린이다. 본원에서, 본 발명자들은 ανβ6이 막사구체신염, 진성 당뇨병, IgA 신장병증, 굿파스쳐 및 알포트 증후군을 앓는 인간의 신장 상피에서 과다발현된다는 것을 밝힌다. 신장 질환에 있어서 ανβ6의 잠재적인 조절 역할을 평가하기 위해, 본 발명자들은 알포트 증후군의 마우스 모델인 Col4A3-/- 마우스에서 신장 섬유화증에 대한 ανβ6 mAb의 기능 차단 및 β6 서브유니트의 유전적 제거의 효과를 연구하였다. 알포트 마우스 신장에서의 ανβ6의 발현은 피질 세뇨관 상피 세포에서 주로 관찰되었고 섬유화증의 진행과 상관관계를 가졌다. ανβ6-차단 mAb를 사용한 처리는 활성화된 섬유아세포의 축적 및 간질(interstitial) 콜라겐 매트릭스의 침착을 억제하였다. 신장 섬유화증의 유사한 억제가 β6-결핍 알포트 마우스에서 관찰되었다. 신장 조직의 전사체 프로파일링은 ανβ6 차단 mAb가 섬유화증 및 염증 매개자의 발현에서의 질환-관련 변화를 유의하게 억제한다는 것을 보여주었다. 전사체 조절의 유사한 패턴이 재조합 가용성 TGF-β RII 처리에서 나타났는데, 이는 ανβ6과 TGF-β의 공유된 조절 기능을 암시한다. 이 발견은 ανβ6이 신장 섬유화증의 조절에 기여하고 이 인테그린이 잠재적 치료제임을 시사한다.
도입:
진행성 섬유화증은 말기 신장 질환의 발달을 초래하는 공통된 과정이고 상피 리모델링, 섬유아세포 활성화, 염증, 및 세포외 매트릭스(ECM)와 세포의 상호작용의 재조직화에 의해 촉진된다. 이러한 과정에 기여하는 분자적 기작은 복잡하고 TGF-β 축의 오조절(misregulation), 비정상적인 ECM 리모델링, 및 인테그린 거대족(superfamily)의 세포 부착 수용체들의 변경된 발현 및 기능을 포함한다1-5. 최근 연구는 신장 상피 세포 및 중간엽 세포에서 여러 인테그린 및 관련 분자들의 중요한 조절 기능을 밝혔다3,6-8.
신장 질환에서 그 발현이 크게 증가되는 인테그린들 중에는 TGF-β-유도가능한 인테그린 ανβ6이 있다5,9,10. ανβ6의 발현은 일반적으로 상피 세포에 한정되어 있고, 상피 세포에서 ανβ66은 정상 성체 조직에서 낮은 수준으로 발현되고 발달 과정, 손상 및 신생물에서 상승된다9,11-13. ανβ6이 건강한 성체 신장에서 비교적 낮은 수준으로 발현된다 하더라도, 그의 발현은 발달 중인 마우스 신장, 특히 근위 세뇨관, 헨레 고리(loop of Henle) 및 집합관에서 현저하다11,12,14. 최근에, ανβ6의 상승된 발현은 다양한 형태의 인간 신장의 병리학적 상태에 대해 보고되어 있다10.
조직 리모델링 동안 생체 내에서의 ανβ6의 증가된 발현과 일치하여, 배양된 상피 세포에서 ανβ6 인테그린의 발현은 EGF 및 TGF-β를 비롯한, 상피 리모델링을 조절하는 사이토카인에 의해 유도될 수 있다5,9. 게다가, 형질전환 마우스의 피부에서의 β6의 과다발현은 자연발생적 만성 상처의 형성을 야기하는 것으로 밝혀졌고15, 이는 ανβ6이 상피 조직 리모델링의 조절에 있어서 중요한 역할을 수행할 수 있음을 암시한다.
ανβ6에 대한 공지된 리간드에는 피브로넥틴, 테나신 및 잠복 관련 펩티드 1 및 3(LAP1 및 LAP3), 및 TGF-β1 및 TGF-β3의 잠복 전구체 형태들의 N-말단 단편이 포함된다16-19. 이들 리간드들과의 결합 결과, ανβ6은 잠복 TGF-β의 세포 부착, 퍼짐, 이동 및 활성화를 매개할 수 있다. TGF-β는 성숙 활성 C-말단 TGF-β 사이토카인과 비-공유적으로 연결된 N-말단 LAP로 절단되어 분비되는 잠복 단백질로서 합성된다. 잠복 TGF-β 복합체는 그의 동족 수용체에 결합할 수 없으므로, 프로테아제에 의한 절단, 낮은 pH 또는 이온화 방사선에의 노출, 및 상기 잠복 복합체가 그의 동족 수용체에 결합할 수 있게 하는 상기 잠복 복합체의 구조적 변화를 포함하는 여러 대안적 기작들 중 하나에 의해 활성 형태로 전환될 때까지 생물학적 불활성 상태로 존재한다20-22. 활성화시키는 구조적 변화는 LAP1 및 LAP3 내에 함유된 RGD 모티프와 인테그린의 직접적인 결합을 수반하는 ανβ6에 의해 유도될 수 있다. 이 결합은 TGF-β 전구체를 수용체 결합-능력 상태로 전환시킨다17,19. 이 발견은 상피 세포의 표면 상의 ανβ6 발현의 상향조절이 국소 TGF-β 활성화를 일으킨 후, 방관자(bystander) 세포 내의 TGF-β-의존적 과정의 주변분비 활성화를 일으킬 수 있다는 것을 시사한다. 이것은 ανβ6 발현 부위에서의 초기 염증 및 섬유화증을 변경시킴에 의해 매개될 수 있는 TGF-β 활성에 대한 간접적인 다운스트림(downstream) 효과에 대한 가능성을 포함할 것이다.
TGF-β가 신장 섬유화증의 중추적인 조절자로서 관여하기 때문에, 본 발명자들은 ανβ6에 의한 그의 국소 활성화가 신장 질환의 발병시기 및 진행에 있어서 중요한 과정일 수 있고 ανβ6 기능의 차단이 신장 섬유화증의 발달을 억제할 수 있다는 가설을 세웠다. 본 명세서에 기재된 연구에서, 본 발명자들은 ανβ6이 신장 섬유화증의 마우스 모델 및 섬유화증성 병리학적 상태를 가진 인간 신장 샘플에서 매우 상향조절되어 있음을 보여준다. 인간 알포트 증후군에서 관찰된 것과 유사한 진행성 신장 질환의 모델인 Col4A3-/- 마우스를 사용하여, 본 발명자들은 ανβ6의 리간드 결합 및 TGF-β 활성화 기능을 차단하는 mAb23 뿐만 아니라, β6의 유전적 제거가 사구체 및 세뇨관간질성 섬유화증을 강하게 억제하고 신장 조직 구조의 파괴를 지연시킨다는 것을 입증한다. 본 발명자들은 ανβ6 인테그린의 발현이 신장에서 세뇨관 상피 세포로 한정된다고 할지라도, ανβ6 인테그린이 상기 조직에서 멀리 위치한 부위에서 보호 효과를 제공할 수 있다는 것을 밝혔다. 이 발견은 항-섬유화증성 효과가 세뇨관 상피 세포 상의 ανβ6의 차단이라는 직접적인 효과 이외에 간접적인 신장외 효과를 통해 매개될 수도 있다는 가능성을 제기하였다. 지연 치료 연구는 ανβ6의 치료적 차단이 신장 섬유화증의 진행을 억제할 뿐만 아니라 기존 섬유화증성 병소를 찾을 수 있게 하는 잠재력을 가진다는 것을 보여준다. 신장 질환 진행과 관련되어 있으며 ανβ6 억제에 의해 영향을 받는 분자적 징후(signature)에 대한 본 발명자들의 분석은 ανβ6 차단 항체의 치료 효과가 전신성 TGF-β 차단의 치료 효과와 유사하고 감소된 TGF-β 활성과 기작적으로 관련되어 있다는 것을 보여준다. 이 데이타는 ανβ6이 신장 섬유화증의 조절에 관여하고 그의 치료적 조절에 대한 새로운 표적 분자를 제공할 수 있다는 것을 암시한다.
재료 및 방법:
1. 시약 . ανβ6 mAb는 본 명세서 및 종래 보고된 바와 같이 생성하였다23. 인간/마우스 키메라 2A1 및 3G9 cDNA는 제1 가닥 cDNA 합성 키트(Amersham/Pharmacia, Piscataway, NJ)를 사용하여 중쇄에 대한 불변 영역 프라이머 CDL-739 및 경쇄에 대한 불변 영역 프라이머 CDL-738로 각각의 모 하이브리도마 총 RNA로부터 생성하였다. 중쇄 및 경쇄 가변 영역 유전자는 cDNA 합성에 사용되는 동일한 3' 프라이머, 대부분의 뮤린 항체 유전자 신호 서열(요청 시 이용가능한 서열)에 특이적인 축퇴 프라이머의 풀 및 Pfu DNA 중합효소(Stratagene, La Jolla CA)를 사용하여 중합효소 연쇄 반응으로 증폭시켰다. 클로닝된 중쇄 및 경쇄 가변 영역들을 인간 IgG1 불변 영역과 함께 포유동물 발현 벡터 내로 연결시켰다. 재조합 가용성 뮤린 TGF-β 수용체 타입 II-Ig 융합 단백질(rsTGF-βRII-Ig)은 공지된 바와 같이7 생성하였고 R&D 시스템스(532-R2, Minneapolis, MN)로부터 구입하였다. 항체는 표시된 바와 같이 구입하였다: FITC에 접합된 팬(pan) 항-사이토케라틴 mAb(C-11), 시그마-알드리치(F3418, St. Louis, MO); 항-라미닌 B1 쇄 mAb(LT3), 케미콘(MAB1928, Temecula, CA); 파이코에리쓰린(PE)-접합 항-αν mAb(RMV7), 케미콘(CBL1346P); 토끼 항-αv, 케미콘(AB1930); PE-래트 IgG1, BD 바이오사이언스(Biosciences)(553925, San Jose, CA); 및 항-평활근 액틴(SMA)-Cy3(Sigma-Aldrich (C-6198)). 본 발명자들은 토끼 다중클론 항-TGF-β(Santa Cruz Biotechnology (sc-146, Santa Cruz, CA))가 잠복 TGF-β를 발현하는 293 세포의 이종이식 절편에 비해 TGF-β의 기본적 활성 형태를 발현하는 293 세포의 이종이식 절편에 우세하게 결합하는 항체임을 확인하였다24.
2. 동물 . 129Sv/J 백그라운드에서의 Col4A3+/- 마우스를 도미니크 코스그로브(Dr. Dominique Cosgrove) 박사(Boy's Town National Research Hospital, Omaha, NE)로부터 입수하여 사육하였고, 주사 연구를 위한 Col4A3-/- 마우스를 발생시켰다. 129SV 백그라운에서의 베타6-/- 마우스를 딘 쉐파드 박사(Dr. Dean Sheppard) (University of California San Francisco, CA)로부터 입수하여 Col4A3+/- 마우스와 교배시켰다. 모든 동물들을 바이오겐 아이덱에서 사육하고 모든 동물 연구를 동물 보호 및 사용 조직 위원회로부터 승인받고 수행하였다.
3. 유세포분류기 . 뮤린 β6으로 안정하게 형질감염된 NIH3T3 세포(NIH3T3b6)를 공지된 바와 같이 생성하였다23. 세포를 트립신처리로 수거하고, PBS 중에서 세척하고 FC 완충제(1 X PBS, 2% FBS, 0.1% NaN3, 1 mM CaCl2, 및 1 mM MgCl2) 중에 재현탁시켰다. 0.2 x 105 세포를 총 100 ㎕의 정제된 1차 항체를 함유하는 FC 완충제 중에서 얼음 상에서 1시간 동안 항온처리하였다. 항온처리 후, 세포를 빙냉 FC 완충제로 2회 세척하고, 5 ㎍/㎖의 PE-접합 당나귀 항-마우스 IgG(Jackson ImmunoResearch)를 함유하는 100 ㎕의 FC 완충제 중에 재현탁시키고, 얼음 상에서 30분 동안 항온처리하였다. αν 발현을 모니터링하기 위해, 세포를 PE-접합 래트 항-마우스 αν mAb(RMV-7) 및 PE-접합 래트 IgG1 대조군과 함께 항온처리하였다. 세포를 빙냉 FC 완충제로 2회 세척하고 표지된 2차 항체의 결합을 유세포분류기로 모니터링하였다.
면역조직화학 . 조직 절편으로부터 파라핀을 크실렌 및 에탄올 중에서 제거하고, 증류수 중에서 재수화시킨 후, 0.45% H2O를 함유하는 메탄올에 함침시켰다. 조직을 펩신(00-3009, Zymed, San Francisco, CA)과 함께 항온처리하고 아비딘 및 바이오틴(SP-2001; Vector Laboratories, Burlingame, CA)으로 차단하였다. 1차 항체를 0.1% BSA가 함유된 PBS 중에서 희석시키고 조직을 4℃에서 밤새 항온처리하였다. 마우스 조직 상의 β6을 면역염색하기 위해, 절편을 항-ανβ6 mAb의 인간/마우스 키메라 형태, 즉 2A123, 및 항-인간 바이오티닐화 2차 항체(PK-6103, Vector Laboratories, Burlingame, CA)와 함께 항온처리하였다. 인간 조직 상의 β6을 면역염색하기 위해, 절편을 뮤린 2A123 및 항-마우스-바이오티닐화 2차 항체(PK-6102, Vector Laboratories)와 함께 항온처리하였다. 아비딘-바이오틴 복합체-호스라디쉬 퍼록시다제(Vector Kit, PK-6102)를 절편에 가하고, 실온에서 30분 동안 항온처리하고, 3,3'-디아미노벤지딘(DAB) 기질을 지시대로 준비하고(SK-4100, Vector Laboratories), 실온에서 5분 동안 절편에 가하였다. 조직 절편을 메이어스 헤마톡실린으로 1분 동안 염색하고 물 및 PBS 중에서 린싱하였다.
O.C.T. 화합물(Cat.# 4583, Sakura Tokyo, Japan) 중에 포매된 동결 조직 절편을 아세톤 중에서 고정시키고 PBS 중의 0.5% 카세인/0.05% 티메로살(thimerosal)로 차단하였다. 인간 조직 상의 β6을 면역염색하기 위해, 절편을 뮤린 2A123 및 항-마우스알렉사 플루오르 594 2차 항체(A-11014, Molecular Probes Eugene, Oregon)와 함께 항온처리하였다. 마우스 조직 상의 β6을 면역염색하기 위해, 절편을 뮤린 2A1 및 항-인간알렉사 플루오르 594-접합 2차 항체(A-11014, Molecular Probes Eugene, Oregon)와 함께 항온처리하였다. 라미닌 및 αv의 면역염색을 위해, 항-래트 알렉사 플루오르 488-접합 2차 항체(A-11006, Molecular Probes)를 사용하였다. 모든 다른 항체들은 전술된 바와 같이 직접 접합되었다. 40X에서 찍은 도 2A를 제외하고 모든 화상을 20X로 찍었다. 모든 인간 조직 샘플을 지역 위원회 심사의 승인 및 환자 승인 하에 입수하였다.
5. 면역조직화학적 정량화 . SMA 면역염색을 메타모르프(MetaMorph) v5.0(Universal Imaging Corporation, Sunnyvale, CA)을 이용하여 정량하였고, 총 화상 크기에 대한 % 양성으로서 표시하였다. 각 동물에 대해, 5개 이상의 피질 절편 및 1 또는 2개의 수질 절편으로부터의 20X 화상을 분석하였다. 처리군의 통계적 분석은 ANOVA를 이용하여 수행하였다.
6. mAb 및 rsTGF-βRII-Ig를 사용한 Col4A3-/- 마우스의 처리 . 129Sv/J 배백그라운드에서 Col4A3+/- 마우스를 사육하여 Col4A3-/- 마우스를 발생시켰다. 표시된 바와 같이 3 내지 7주령 또는 8.5주령에서 주당 3회씩 단백질을 마우스의 복강내로 주사하였다. 4 mg/kg의 mAb를 복강내로 주사하고, 2 mg/kg의 rsTGF-βRII-Ig를 주사하였다. 마우스를 마취시키고 RNA 및 면역염색을 위해 신장을 수거하였다. 모든 동물 연구는 동물 보호 및 사용 조직 위원회으로부터 승인받아 수행하였다.
7. 총 RNA 정제 및 cDNA 합성
신장을 TRIzol 중에서 직접 균질화하고(155-96-018, Invitrogen, Carlsbad, CA), 추가 1 ㎖ 산성 페놀:클로로포름:이소아밀 알코올 25:24:1 추출(pH 6.6)을 이용하여 제조자의 프로토콜에 따라 RNA를 추출하였다. 정제된 총 RNA를 디에틸피로카보네이트(DEPC) 중에 재현탁시키고 H2O(Ambion Inc, Austin, TX)로 처리하고, 260 및 280에서 기록하였다(Spectra max Plus, Molecular devices, Sunnyvale, CA). 20℃에서 15분 동안 5 유니트의 DNase I 증폭 등급(cat# 18068-015, Invitrogen)을 사용하여 잔류 DNA를 제거하였다. 제조자의 프로토콜(cat# 4322171, Applied Biosystems Inc, Foster City, CA)에 따라 고 성능 cDNA 보존(archive) 키트를 사용하여 cDNA를 발생시켰다.
8. 택만(Taqman)을 위한 프라이머, 프로브 및 올리고뉴클레오티드 표준 주형 의 디자인. 올리고뉴클레오티드 프라이머 및 택만 MGB 프로브를 프라이머 발현 버젼 2.0.0(Applied Biosystems Inc.)을 사용하여 아피메트릭스(Affymetrix) 컨센서스 서열로부터 디자인하였다. 택만 MGB 프로브를 5'에 공유결합된 형광 레포터 염료(FAM) 및 3' 말단에 공유결합된 마이너 그루브(minor groove) 결합제/비-형광 켄쳐(quencher)(MGBNF)로 디자인하였다. 올리고뉴클레오티드 표준 주형은 앰플리콘의 5' 및 3' 말단에 10 bp의 유전자 특이적 서열을 부가함으로써 디자인하였다. 역상 HPLC로 정제한 프라이머 및 올리고뉴클레오티드 표준 주형을 바이오서치 테크놀로지스 인코포레이티드(Biosearch technologies Inc., Novato, CA)로부터 구입하였다. 뮤린 글리세르알데히드-3-포스페이트 데히드로게나제(GAPDH)에 대한 HPLC 정제 프라이머 및 프로브는 바이오겐 아이덱에서 합성하였다[CATGGCCTTCCGTGTTCCTA(서열번호 147), GCGGCACGTCAGATCC(서열번호 148), 6FAM-CCCCAATGTGTCCGTC(서열번호 149)].
9. 택만 열적 사이클링(Taqman thermal cycling) . 샘플 및 표준물에 대한 4회 반복 PCR 반응을 하기 조건 하에 7900HT(Applied Biosystems Inc.) 열적 사이클러 내에서 순환시켰다: 50℃에서 2분(우라실 N-데글리코실라제 절단), 95℃에서 10분(Taq 열안정성 중합효소의 활성화), 및 95℃에서 15초 및 60℃에서 60초로 구성된 40 사이클. 형광 방출을 각 반응 웰에 대해 수행 기간 동안 7초마다 수거하였다. 서열 검출 소프트웨어(Applied Biosystems Inc.)를 사용하여 올리고뉴클레오티드 표준 곡선과 비교함으로써 상대적 전사체 양을 각 샘플에 대해 측정하였다.
10. 프로브 표지, 혼성화 및 전사체 프로파일링에 대한 스캐닝 . 샘플 표지, 혼성화 및 염색을 유전자칩® 발현 분석(Affymetrix, Santa Clara, CA)에 대한 아피메트릭스 테크니칼 메뉴얼(701021 rev 1)의 진핵 표적 준비(Eukaryotic Target Preparation) 프로토콜에 따라 수행하였다. 요약하면, 5 μg의 정제된 총 RNA를 42℃에서 200 U 수퍼스크립트 II(Superscript II)(cat #, 18064-022, Invitrogen) 및 0.5 μg의 (dT)-T7 프라이머(서열번호 150)[5'-GGCCAGTGAATTGTAATACGACTCACTATAGGGAGGCGG(T)24]와 함께 20 ㎕ 제1 가닥 반응 중에서 1시간 동안 사용하였다. 40 U의 이. 콜라이 DNA 중합효소(cat # 18010-025, Invitrogen), 2 U의 이. 콜라이 RNase H(cat #18021-071, Invitrogen) 및 10 U의 이. 콜라이 DNA 리가제(cat # 18052-019, Invitrogen)를 첨가한 후 16℃에서 2시간 동안 항온처리함으로써 제2 가닥 합성을 수행하였다. 제2 가닥 합성 반응액은 제조자의 프로토콜에 따라 유전자칩® 샘플 클린업 모듈(cat #900371, Affymetrix, Santa Clara, CA)을 사용하여 정제하였다. 정제된 cDNA는 제조자의 프로토콜에 따라 바이오어레이 고수율 RNA 전사 표지 키트(cat #42655-40, Enzo Life Sciences, Inc., Parmingdale, NY)를 사용하여 증폭시켜 바이오틴으로 표지된 cRNA(상보적 RNA) 70 내지 120 μg을 생성하였다. 마우스 MgU74Av2, MgU74Bv2 및 MgU74Cv2 유전자칩® 프로브 어레이를 제조자의 프로토콜에 따라 유전자칩® 혼성화 오븐 640(Affymetrix, Santa Clara, CA)에서 예비혼성화시켰다. 단편화되고 표지된 cRNA를 100 mM 2-모르폴리노에탄설폰산, 1 M [Na+], 20 mM EDTA, 0.01% Tween 20, 0.5 mg/㎖ 아세틸화 BSA, 0.1 mg/㎖ 헤링 정자 DNA, 대조군 올리고 B2 및 대조군 전사체 바이오B 1.5 pM, 바이오C 5 pM, 바이오D 25 pM 및 cre 100 pM이 함유된 300 ㎕ 1X 혼성화 완충제 중에 재현탁시키고, 제조자의 프로토콜(Affymetrix, Santa Clara, CA)에 따라 유전자칩® 프로브 어레이에 혼성화시켰다. 혼성화된 유전자칩® 프로브 어레이를 세척하고 스트렙타비딘-파이코에리쓰린(cat # S866, Molecular Probes, Eugene, OR)을 사용하여 염색하고, 유전자칩® 플루딕스 스테이션 400(GeneChip® Fluidics Station 400)(Affymetrix, Santa Clara, CA)을 사용하여 바이오티닐화 항-스트렙타비딘(BA-0500, Vector Laboratories, Burlingame, CA)으로 증폭시켰다. 유전자칩® 프로브 어레이를 진어레이(GeneArray) 스캐너(Hewlett Packard, Corvallis, OR)를 사용하여 스캐닝하였다.
11. 전사체 프로파일링 데이타 분석 . 어레이 스캔을 아피메트릭스로 전환하였다. CEL 파일 및 생성된 데이타 세트(완전한 실험을 나타내는 CEL 파일의 군)를 로버스트 마이크로어레이 애버리지(Robust Microarray Average; RMA) 방법을 이용하여 표준화하였다. 통계적 및 클러스터링 분석은 진스프링(GeneSpring)(Agilent) 및 스팟파이어(Spotfire) 데이타 분석 수단을 이용하여 수행하였다. 본 발명자들은 미처리된 Col4a3-/- 군에 비해 실험 처리에 의해 신호 강도가 p<0.05에서 2-배 이상 변경된 프로브 세트를 확인하기 위해 2-단계 ANOVA 및 배수-변화 필터링(fold-change filtering)을 이용하였다. 유사하게, p<0.01의 통계적 컷오프 및 2의 신호 배수-변화 컷오프를 사용하여, 미처리된 Col4a3-널(null) 군과 천연 야생형 군 사이의 차등적 발현에 대해 질환-관련 전사체를 선별하였다. 생성된 유전자 군의 프로파일 및 실험 조건의 분류를 계층학적 클러스터링으로 분석하고 가시화하였다. IPA 데이타베이스(Ingenuity Systems)를 이용하여 실제 경로 분석을 수행하였다.
결과:
1. 섬유화증성 병리학적 상태를 가진 인간 신장 샘플에서의 α ν β 6 의 발현. 염증/섬유화증성 병리학적 상태와 관련된 여러 다양한 유형의 인간 신장 질환은 신장 조직에서의 TGF-β의 상응하는 증가된 발현을 보였다25-27. 본 발명자들은 면역조직화학적 분석을 이용하여, TGF-β의 증가된 활성화를 이끌어 내는 잠재적인 기작으로서 만성 염증 및 섬유화증과 관련된 인간 신장 생검 샘플에서의 ανβ6의 발현을 조사하였다(도 29A 및 29B). 막 사구체신염, 진성 당뇨병, IgA 신장병증, 굿파스쳐, 알포트 및 루프스로부터의 조직 샘플들은 모두 확장되고 손상된 세뇨관의 상피 라이닝(lining)에서 현저한 ανβ6 염색을 보였다. 반대로, 형태학적으로 정상인 신장(신장 암종 및 정상 조직)의 샘플은 세뇨관에서 종종 최소한의 면역 염색을 보였다. 사구체 염색은 분석된 모든 신장 샘플에서 존재하지 않았다. 이 발견은 ανβ6이 건강한 성체 상피에서는 낮은 수준으로 발현되지만, 조직 손상 및 회복 도중에는 상향조절된다는 종래 보고와 일치한다10,11,13,15,17.
2. Col4A3-/- 마우스의 신장에서의 α ν β 6 의 발현은 신장 섬유화증의 진행과 상관관계를 가진다. 인간 알포트 질환의 마우스 모델인 Col4A3-/- 마우스는 다수의 섬유화증 표준 마커의 증가된 발현에 동반되는 신장의 사구체 및 간질 영역 둘다에서의 ECM의 축적을 초래하는 진행성 사구체신염을 발병시킨다28,29. 종래, rsTGF-βRII-Ig를 사용한 Col4A3-/- 마우스의 처리가 신장 섬유화증을 억제한다고 이미 보고되었다7. Col4A3-/- 마우스로부터의 신장은 대략 5 또는 6주령에서 섬유화증의 조직학적 징후를 보이기 시작한다. 상기 질환은 연령에 따라 급속히 진행되고, 마우스는 대략 11주째에 신부전으로 사망한다. 이종접합 Col4A3+/- 마우스는 사구체신염을 발병시키지 않으며, 그의 신장은 야생형 한배새끼(littermate)의 신장과 조직학적으로 구별될 수 없다. 연령이 증가함에 따른, Col4A3+/- 및 Col4A3-/- (알포트) 마우스의 신장에서의 ανβ6 발현의 동역학적 변화를 조사하기 위해, 본 발명자들은 4주, 7주 및 8주령의 마우스로부터 단리된 신장에서의 ανβ6 발현의 면역조직화학적 분석을 수행하였다(도 30A 내지 30C). 4주령에서, Col4A3+/- 및 Col4A3-/- 마우스 둘다의 신장 세뇨관에서 ανβ6의 간헐적 발현이 있었다. 7주까지, ανβ6의 발현은 Col4A3-/- 마우스의 세뇨관 상피 세포에서 현저히 증가하였지만 Col4A3+/- 신장에서는 그러하지 않았다. ανβ6의 이러한 증가된 발현은 8주령보다 더 많은 연령의 Col4A3-/- 마우스에서 지속되었다. 본 발명자들은 6주령 후의 Col4A3-/- (알포트) 마우스에서 확장되고 손상된 세뇨관의 상피 세포에서의 ανβ6 염색의 강도가 증가되며, 이것이 강한 ανβ6 발현을 나타내는 신장 조직 영역에서의 유의한 증가에 동반된다는 것도 관찰하였다. 7주령 또는 8주령 기간 동안의 증가된 발현은 Col4A3-/- 마우스에서의 신장 섬유화증의 급속한 진행과 일치한다. 반대로, 최소한의 ανβ6 발현, 및 그의 염색 강도의 미미한 검출가능한 연령-의존적 증가가 기간 전체에 걸쳐 Col4A3+/- 마우스의 신장에서 검출되었다. Col4A3-/- 마우스 신장에서의 ανβ6의 발현이 섬유화증의 진행과 상관관계를 가지기 때문에, 본 발명자들은 ανβ6의 차단이 섬유화증성 병소의 개시 및 진행을 억제할 수 있는지를 확인하고자 하였다.
3. Col4A3-/- 마우스의 신장에서 α ν β 6 에 결합하는 mAb의 특이성 . 본 발명자들은 리간드와 결합하는 ανβ6의 능력에 영향을 주지 않으면서 ανβ6에 결합하는 mAb(비-차단 mAb) 및 리간드 결합 및 ανβ6-매개 TGF-β 활성화 둘다를 차단하는 mAb(차단 mAb)를 비롯한 강력한 선택적 항-ανβ6 mAb23의 발생을 이미 보고하였다. 생체 내 연구에 사용되는 ανβ6 차단 mAb가 ανβ6 인테그린과의 결합에 대해 선택성을 가진다는 것을 입증하기 위해, 본 발명자들은 ανβ6 mAb와 형질감염되지 않은 NIH3T3 모세포의 결합과 ανβ6 mAb와 뮤린 β cDNA로 형질감염된 NIH3T3 세포(NIH3T3b6)의 결합을 비교하는 FACS 분석을 수행하였다(도 31A). 대조군 항-αν mAb인 RMV7은 형질감염되지 않은 NIH3T3 세포 및 ανβ6-발현 NIH3T3 세포 둘다를 염색하였지만, 항-ανβ6 mAb는 NIH3T3b6 세포에만 선택적으로 결합하였다. 신장에서 ανβ6과의 결합 특이성을 확인하기 위해, 본 발명자들은 차단 ανβ6 mAb 중 하나인 3G9의 인간/마우스 키메라 형태를 발생시키고, 토끼 항-αν 다중클론 항체를 사용하여 생성한 면역염색의 패턴을 비교하였다(도 31B 및 31C). 3G9의 키메라 뮤린 형태와 3G9의 원래의 뮤린 형태는 FACS 및 ELISA에 의한 측정 시 필적할만한 표적 결합 친화성을 가졌다(데이타는 나타내지 않음). 3G9의 키메라 형태는 Col4A3-/- 마우스의 신장에서 세뇨관 상피 세포를 특이적으로 면역염색하였고 β6-/- 마우스와 교배된 Col4A3-/- 마우스(Col4A3-/-;β6-/-)로부터 얻은 신장 절편의 면역염색을 전혀 보이지 않았다. 항-αν 항체를 사용한 신장의 면역염색은 Col4A3-/- 마우스 사이 또는 Col4A3-/-;β6-/- 마우스 사이에 유의한 차이를 보이지 않았다.
4. 항-α ν β 6 mAb를 사용한 Col4A3-/- (알포트) 마우스의 처리는 신장 섬유 화증을 억제한다. 신장 섬유화증의 조절에 있어서 ανβ6의 잠재적인 기능적 관여를 확인하기 위해, 본 발명자들은 진행된 섬유화증성 병소의 개시 및 진행에 영향을 주는 차단 ανβ6 mAb의 능력을 시험하였다. ανβ6 차단의 예방적 효과를 평가하기 위해, Col4A3-/- 마우스를 3주령 내지 7주령 또는 3주령 내지 8.5주령에서 2가지 상이한 차단 ανβ6 mAb, 즉 3G9 또는 8G6; 비-차단 ανβ6 mAb, 즉 6.8B3; 또는 이소타입-일치 음성 대조군 mAb, 즉 1E6으로 처리하였다. 표현형에 대한 기준으로 사용하고 전신 TGF-β 억제의 효과를 모니터링하기 위해, 이들 연구는 rsTGFβRII-Ig로 처리된 Col4A3-/- 마우스도 포함하였다. RNA의 조직학적 평가 및 단리를 위해 신장을 수거하였다. 섬유화증의 조직학적 특징뿐만 아니라 SMA 발현도 7주 및 8.5주에서 연령-일치 Col4A3+/- 마우스의 신장에 비해 Col4A3-/- 신장에서 현저히 증가되었다. 음성 대조군 mAb로 처리된 Col4A3-/- 마우스의 신장은 보만(Bowman) 캡슐 내에서 팽창된 섬유화증성 사구체 토리사이질 및 반월(crescent) 형성을 제공하였다(도 32A, 1E6). 또한, 이 신장은 세뇨관 상피 손상 및 확장과 관련된 현저한 근섬유아세포 활성화 및 간질성 섬유화증을 보였다. 차단 ανβ6 mAb, 즉 6.3G9 또는 6.8G6를 사용한 Col4A3-/- 마우스의 처리는 사구체 및 간질성 손상 및 섬유화증을 현저히 감소시켜, 신장 구조를 전체적으로 상당히 보존시켰다(도 32A, 3G9 및 8G6). 차단 ανβ6 mAb의 이 효과는 사구체에서 65% 초과의 SMA 발현 감소 및 간질 영역에서의 90% 초과의 SMA 발현 감소에 동반된다(도 32B 및 32C). 사구체에서의 SMA 발현에 대한 차단 mAb의 효과는 ανβ6 인테그린의 발현이 세뇨관 상피 세포에 한정되지만, 그의 기능의 차단은 상기 조직에서 보다 멀리 떨어진 부위에서 효과를 나타낼 수 있음을 암시한다. 이것은 적어도 부분적으로는 차단 ανβ6 mAb의 간접적인 전신적 효과로 인해 매개될 수 있다. 비-차단 ανβ6 mAb, 즉 8B3을 주사한 Col4A3-/- 마우스의 신장에서는 섬유화증의 진행에 대해 효과가 전혀 관찰되지 않았다. TGF-β의 차단을 통한 신장 섬유화증의 종래 보고된 억제와 일치하여7,30-33, rsTGFβRII-Ig를 사용한 Col4A3-/- 마우스의 처리는 신장 조직의 조직학적 외관 및 SMA 함량에서의 변화에 의해 판단될 때, 차단 ανβ6 mAb에 의해 나타나는 신장 섬유화증의 억제와 유사한 신장 섬유화증의 억제를 나타내었다. SMA 발현에 대한 ανβ6 차단 mAb의 효과는 콜라겐1α1 및 콜라겐1α2 mRNA의 총 신장 조직 수준에서의 감소에 동반되었다(도 33A 및 33B). 3G9, 8G6 또는 rsTGF-βRII-Ig를 사용한 Col4A3-/- 마우스의 처리는 콜라겐1α1 및 콜라겐1α2 mRNA 존재도를 유의하게 감소시킨 반면, 비-차단 ανβ6 mAb 및 동위원소 대조군 mAb는 이 전사체들의 수준에 대해 유의한 효과를 전혀 나타내지 않았다.
진행된 신장 섬유화증에 대한 ανβ6 차단의 영향을 시험하기 위해, 본 발명자들은 신장 병리학적 상태가 SMA-양성 활성화 섬유아세포의 측정가능한 손상 및 축적에 의해 분명해지는 시기인 6주 연령의 Col4A3-/- 마우스에서 신장 섬유화증에 대한 ανβ6 차단 mAb의 효과를 연구하였다. 마우스는 2.5주령에서 ανβ6 차단 mAb, 즉 3G9 또는 이소타입 대조군 mAb, 즉 1E6으로 처리된 후, 8.5주령에서 희생되었다. SMA 면역염색의 정량화는 이소타입 대조군 mAb로 처리된 마우스에 비해 3G9로 처리된 Col4A3-/- 마우스의 신장에서 SMA 양성 섬유아세포의 존재의 감소를 보여주었다(도 34A). 마찬가지로 중요한, 지연된 3G9 처리를 이용한 SMA 면역염색의 강도 및 영역은 처리 개시시점(6주)에서 관찰된 SMA의 수준에 비해 감소되었다. 3G9를 사용한 Col4A3-/- 마우스의 지연된 처리는 1E6으로 처리된 Col4A3-/- 마우스로부터 단리된 신장에 비해 간질에서의 손상된 세뇨관 및 섬유화증의 상당한 감소를 비롯한 병리학적으로 현저한 복구를 유도하였다.
이 결과는 ανβ6의 치료적 차단이 신장 섬유화증의 진행을 억제할 뿐만 아니라 기존 섬유화증성 병소를 찾을 수 있게 할 수 있다는 것을 암시한다.
5. 항-α ν β 6 mAb에 의한 신장 유전자 발현의 조절 . ανβ6 기능과 관련된 질환 기작을 더 파악하기 위해, 본 발명자들은 야생형 마우스 및 알포트 마우스의 신장 조직에서의 유전자 발현의 아피메트릭스 유전자칩 분석을 수행하였다. Col4a3-/- 신장에서의 발현이 변경된 유전자 군은 7주령의 Col4a3-/- 연령-일치 야생형 신장 사이의 표준화된 신호 강도에서 2배 이상의 평균 차이를 보이는 395개의 유전자칩 프로브세트로서 확인되었다(p<0.01, 도 35). 차등적으로 발현되는 유전자들의 기능적 해석은 정교 경로 분석(IPA) 수단을 이용하여 수행하였고 Col4a3-/- 신장에서 과다발현되는 유전자가 염증 및 백혈구 기능의 조절과 주로 관련되어 있다는 것을 보여주는 반면, 발현이 감소된 유전자들은 대사 조절과 주로 관련되어 있었다(도 36).
ανβ6-차단 mAb를 사용한 동물의 처리는 Col4A3-/- 신장에서 유전자 하위세트의 차등적 발현을 약화시켰다. 본 발명자들은 p<0.05에서 실험적 처리에 의해 영향을 받는 전사체를 확인하기 위해 편차 분석(Welch ANOVA)을 이용하였고, 처리에 반응하여 신호 강도에서 2-배 이상의 차이를 보이는 프로브세트를 선별하기 위해 생성된 프로브세트를 더 여과하였다. 이 절차는 3G9, 8G6 및 TGFβRII-Fc 각각에 의해 유의하게 영향을 받는 56개, 42개 및 28개의 프로브세트를 제공하였다. 이 프로브 세트 군들은 상당한 중첩을 보였고, 상기 군들 각각은 Col4a3-/- 신장에서 차등적으로 발현되는 전사체들에 상응하는 395개의 완전히 포함된 하위세트를 대표하였다(도 37A). 본 발명자들은 2가지 상이한 생화학적 부류에 속하는 것으로 이미 밝혀진 ανβ6 차단 mAb, 즉 3G9 및 8G6(도 35, 도 37A)에 의한 유전자 발현의 유사한 조절을 관찰하였다23. 비-차단 항-ανβ6 mAb 8B3 및 이소타입 대조군 mAb 1E6 둘다가 유전자 발현에 대해 유의한 효과를 나타내지 않았다. 따라서, 신장 유전자 발현에 대한 ανβ6 mAb의 영향은 인테그린 신호전달의 활성화 또는 비-특이적 과정에 기인한 것이라기 보다는 ανβ6 기능의 차단에 기인하는 듯하였다. 본 발명자들은 정상 야생형 신장 조직에서 유전자 발현에 대한 차단 ανβ6 mAb 3G9의 유의한 효과를 전혀 발견하지 못하였다. 이 관찰결과는 본 발명자들의 실험에서 관찰된 ανβ6-차단 mAb의 효과가 주로 ανβ6의 질환-특이적 조절 기능을 반영한다는 것을 암시하였다.
ανβ6-mAb에 의해 조절되는 유전자와 주요 조절 경로의 잠재적 관계를 조사하기 위해, 본 발명자들은 IPA 소프트웨어를 사용하여 각 유전자 리스트에 대해 실제 조절 네트워크 분석을 수행하였다(도 37). IPA는 유전자와 단백질 사이의 공지된 물리적 및 조절적 상호작용의 관리된 데이타베이스에 대한 입력정보로서 제공된 유전자 리스트를 비교한다. 이 분석은 소정의 조절된 유전자 리스트에 의해 가장 잘 반영될 것 같은 조절 경로의 순위화된 리스트 및 개개의 구성을 제공한다. 3G9 및 8G6에 의해 조절되는 유전자 리스트는 완전히 동일하지 않지만, IPA는 3G9(도 37A)에 의해서 뿐만 아니라 8G6(도 37B)에 의해서도 영향을 받는 최고-스코어링 조절 네트워크로서 TGFβ-의존적 네트워크를 보였다. 이 발견과 일치하여, 실험 군의 평균 유전자 발현 프로파일의 계층적 클러스터링은 ανβ6 mAb의 의한 유전자 조절 패턴과 rsTGFβRII-Ig에 의한 유전자 조절 패턴 사이의 유사성을 입증하였다(도 35).
6. α ν β 6 의 차단은 Col4A3-/- 신장에서 TGFβ의 발현을 감소시킨다 . 3G9 및 8G6 mAb 처리에서 검출된 감소된 신장 섬유화증이 감소된 TGF-β 발현과 관련되어 있는지를 확인하기 위해, 신장 절편을 항-TGF-β1 mAb로 면역염색하였다(도 38A). 면역염색에 사용된 mAb는 잠복 TGF-β에 비해 활성 TGF-β를 기본적으로 발현하는 조직 절편에 우세하게 결합하는 mAb이었다(데이타는 나타내지 않았음)24. 3G9 또는 8G6을 사용한 마우스의 처리는 신장의 간질 영역 및 사구체 영역 둘다에서 TGF-β1 면역염색을 유의하게 감소시켰다. TGF-β 발현의 이러한 변화는 TGF-β mRNA의 총 신장 조직 수준의 유사한 변화에 동반되었다(도 38B). TGF-β1 면역염색의 패턴은 ανβ6의 발현이 신장 세뇨관의 상피 라이닝에 한정되어 있다 하더라도 ανβ6 기능의 억제가 ανβ6-발현 영역에 바로 인접하지 않는 멀리 떨어진 부위 예컨대, 사구체 영역에서 TGF-β 발현을 감소시킬 수 있다는 것을 보여주었다. 이것은 ανβ6이 국소 TGF-β 활성화의 초기 유발제로서 작용할 수 있다 하더라도 TGF-β에 대한 장기간 조절 효과 또한 나타낼 수 있다는 것을 암시할 수 있다. 이러한 장기간 효과의 한 가능한 기작은 TGF-β 발현 조직 영역을 팽창시키는 자가분비 또는 주변분비 방식으로 그 자신의 발현을 활성화시키는 TGF-β의 능력에 기초할 수 있다. 상기 가능한 기작은 ανβ6의 차단에 의한 TGF-β 활성화의 초기 국소 억제가 염증 및 섬유화증의 과정을 방해하고, 이러한 방해가 TGF-β 활성을 간접적으로 더 하향 조절할 수 있다는 가능성도 포함한다.
7. Col4A3-/- 마우스에서의 β 6 유전자의 유전적 제거. mAb 실험으로부터 얻은 발견을 검증하기 위해, 본 발명자들은 Col4A3 및 β6 이중 넉아웃 마우스(Col4A3-/-;β6-/-)를 발생시켰다. 연령-일치 Col4A3-/-;β6+/- 마우스 및 Col4A3-/-;β6-/- 마우스의 신장의 조직학적 검사는 ανβ6-차단 mAb를 사용한 연구에서 얻은 결과를 확인시켜 주었다. 7 내지 10주 연령의 Col4A3-/-;β6+/- 마우스에 비해 연령-일치 Col4A3-/-;β6-/- 마우스의 신장에서 SMA 면역염색이 유의하게 감소되어 있었다(데이타는 나타내지 않았음). 이는 신장의 사구체 및 간질 영역에서의 섬유화증의 현저한 감소에 동반되었다(도 39). 이는 10주 연령의 Col4A3-/-;β6+/- 마우스의 신장에 비해 연령-일치 Col4A3-/-;β6-/- 마우스의 트리크롬-매손 염색 신장에서 관찰된 콜라겐 발현의 감소 및 잘 보존된 신장 구조에 의해 입증된다. β6 기능의 유전적 제거에 비해 차단 ανβ6 mAb에서 관찰된 항-섬유화증성 효과의 일관성은 mAb 처리가 생체 내에서 ανβ6 기능을 차단하는 효율적인 방법임을 의미한다.
7. Col4A3-/- 마우스에서의 신장 섬유화증의 3G9 억제의 투여량 반응 . Col4A3-/- 마우스를 증가하는 농도의 3G9로 주당 3회씩 처리하였다. 마우스를 3주령 내지 7주령에서 처리한 후 희생시켰다. SMA의 면역조직화학적 분석은 신장의 피질 및 수질 영역 둘다에서 분석하였다. 투여량-적정은 3G9가 Col4A3-/- 마우스에서 0.3 내지 0.4 mg/kg의 ED50으로 SMA 발현을 억제한다는 것을 입증하였다(도 40).
논의:
신장 질환의 진행은 궁극적으로 신장 조직 구조의 파괴 및 신장 기능의 상실을 초래하는 강한 조직 리모델링, 염증, 및 섬유화증성 병소의 형성에 동반된다. 섬유화증은 이 과정에서 중추적이고 섬유아세포의 활성화 및 팽창, 병든 조직의 신생혈관형성 및 세포외 매트릭스의 대규모 침착을 수반한다. 이러한 사건들의 주요 유발자로서 관여하는 분자적 기작은 세포 집단화 섬유화증성 병소에서의 ECM 단백질 및 이의 수용체의 변경된 발현에 동반되는 TGF-β 축의 오조절을 포함한다34-36. TGF-β의 상승된 발현은 섬유화증성 인간 조직의 특징이며25-27, 조직 섬유화증을 촉진하는 데 있어서 TGF-β의 기능적 중요성은 시험관 내 및 동물 질환 모델에서 증명되었다. TGF-β의 과다발현은 생체 내에서 섬유아세포 활성화 및 혈관신생을 유도하고 기관형적 세포 배양물에서 ECM의 과도한 생성을 활성화시키기에 충분하다34,37,38. 반대로, TGF-β 신호전달의 유전적 또는 약리학적 파괴는 폐, 피부 및 신장 섬유화증 모델에서 섬유화증으로부터의 효율적인 보호를 제공한다30,32,33,39-41.
TGF-β 기능(차단 mAb, rsTGF-βRII-Ig, TGF-β 수용체 키나제 억제제)의 전신적 억제에 대한 여러 연구는 동물 질환 모델에서 섬유화증을 약화시키는 것으로 밝혀졌다. 그러나, 이 방법들 모두가 TGF-β의 활성화된 형태를 차단하는 것이 목적이지만, TGF-β의 활성화를 차단할 수 있는 잠재적인 치료제는 덜 연구된 상태로 남아있다. TGF-β는 잠복 전구체로서 발현되며, 반응성 산소 종, pH, 트롬보스폰딘-1, 세포외 프로테아제 및 인테그린을 수반하는 다수의 기작에 의해 생물학적 활성 사이토카인으로 전환될 수 있다21,42-44. 이들 중 특히 흥미로운 것은 상피 리모델링 부위에서 발현되는 TGF-β-유도가능한 인테그린인 ανβ6이며 이는 잠복 TGF-β의 수용체 및 활성화제로서 작용하는 것으로 밝혀졌다11,17,45. β 서브유니트는 여러 형태의 신장 질환에서 상향조절되어 있고10, 그의 유전적 제거는 편측 요관 폐색(UUO)의 마우스 모델에서 손상-유도 신장 섬유화증으로부터 현저한 보호를 제공하는 것으로 밝혀졌다46. 섬유화증으로부터의 β6-결핍 마우스의 유사한 보호가 블레오마이신 폐 섬유화증 모델에서 관찰되었는데, 이는 ανβ6이 다양한 조직에서 섬유화증을 매개할 수 있다는 것을 암시한다17,47. 흥미롭게도, TGF-β 신호전달의 중추적 매개자인 SMAD2의 UUO-유도 인산화는 β6-결핍 신장에서 현저히 약화되었는데, 이는 ανβ6이 생체 내에서 사실상 TGF-β 조절 회로의 일부로서 작동할 수 있다는 것을 의미한다46.
β6 넉아웃 마우스를 사용한 종래 연구는 ανβ6이 섬유화증의 개시에서 일정한 역할을 할 수 있다는 증거를 제공하였고, 이는 이 인테그린이 잠재적인 새로운 치료 표적임을 암시한다. 본 발명자들은 차단 mAb를 사용한 ανβ6 기능의 약리학적 억제가 상염색체 열성 알포트 증후군의 동물 모델인 Col4A3-/- 마우스에서 신장 섬유화증을 약화시킬 수 있는지를 확인하고자 하였다28,29. 알포트 증후군은 Col4A3, Col4A4 또는 Col4A5 유전자에서의 돌연변이에 의해 야기되는 유전성 질환이다48. 이 유전자들에서의 결핍은 콜라겐 IV 네트워크의 비정상적인 조립 및 사구체 및 세뇨관 기저막의 비정상적인 형성을 초래한다. 알포트 환자는 말기 신장 질환을 초래하는 진행성 사구체신염을 발병시킨다. 본 발명자들은 인간 알포트 뿐만 아니라 마우스 Col4A3-/- 신장 조직에서 ανβ6의 현저한 상향조절을 관찰하였다. Col4A3-/- 마우스 신장에서, ανβ6의 증가된 발현은 세뇨관 상피에서 특히 현저하였고, 세뇨관 상피에서 SMA-양성 세포의 광범위한 팽창 및 콜라겐 침착을 동반하되 먼저 일어났다. 이러한 발현 패턴에 기초하여, 본 발명자들은 ανβ6이 손상에 대한 상피 반응의 사이클에서 초기에 상향조절되고 지속적인 상피 손상의 셋팅에 있어서 섬유화증의 개시 및 유지 둘다에서 중요한 매개자일 수 있다는 가설을 세웠다. 본 발명자들의 연구 결과는 ανβ6의 항체-매개 차단이 신장 섬유화증의 개시 뿐만 아니라 초기 진행을 억제할 수 있고 그의 유지도 억제할 수 있다는 것을 보여준다. UUO의 β6-결핍 마우스 모델로부터 얻은 종래 발견46과 일치하여, 본 실험에서 관찰된 ανβ6-차단 mAb의 항-섬유화증성 효과는 감소된 TGF-β 활성 및 발현과 상관관계를 가졌다. 흥미롭게도, ανβ6 mAb 처리 후의 TGF-β 및 SMA 발현의 명백한 감소는 ανβ6 양성 세포의 바로 인접한 곳에서 일어날 뿐만 아니라 비교적 멀리 떨어져 있는 조직 영역에서도 검출될 수 있었다. 이 발견은 ανβ6이 직접적 및 간접적인 방식으로, 예를 들면, TGF-β 발현의 주변분비 자가-활성화를 통해 TGF-β 축의 활성화에 기여할 수 있다는 것을 시사할 수 있지만, ανβ6 차단이 전신성 면역 기능의 변경을 비롯한 신장외 효과를 통해 보호를 제공할 수 있다는 가능성을 배제하지 않는다. 본 발명자들은 ανβ6 mAb 처리로부터 4주 후 마우스에서 다수의 케모카인 및 사이토카인 및 말초혈 집단의 혈청 수준을 평가하였고 유의한 변화를 발견하지 못하였다. 또한, Col4A3-/- 신장의 신장 내 단핵구에서의 최소한의 변화만이 ανβ6 mAb 처리를 사용한 면역조직화학적 분석 또는 β6 유전자의 유전적 넉아웃에 의해 검출되었다. ανβ66 mAb 처리 또는 이식 연구를 이용한 면역 시스템의 기능적 상태를 평가하는 추가 연구는 이를 더 완벽하게 해결할 수 있을 것이다.
TGF-β 경로의 오조절 및 과활성은 Col4A3-/- 마우스에서 신장 질환의 진행에 관여하는 중요한 기작으로서 관련되어 있다6. 섬유화증성 질환에서 이 사이토카인의 명백히 우세한 역할을 설명하는 데 도움을 줄 수 있는 TGF-β 회로의 한 가지 흥미로운 특징은 그 자신의 발현을 유도하는 TGF-β의 능력이다. 이것은 ανβ6 차단의 항-섬유화증성 효과가 적어도 부분적으로 간접적인 기작에 의해 매개될 수 있다는 가능성을 제기한다. 이것은 염증 및 섬유화증을 국소적으로 변경시키고 추후 증가된 TGF-β 활성을 방해함으로써 TGF-β 발현에 대한 다운스트림 효과를 포함할 수 있다. ανβ6이 TGF-β에 의해 유도될 수 있고 잠복 TGF-β 활성화를 촉진할 수 있기 때문에, 본 발명자들은 Col4A3-/- 신장의 병리학적 상태를 매개하는 데 있어서 TGF-β와 ανβ6 사이의 기능적 관계를 조사하였다. 본 발명자들은 Col4A3-/- 마우스의 신장에서 질환-관련 전사체의 발현에 대한 ανβ6 mAb의 영향과 rsTGF-βRII-Ig의 영향을 비교하였다. 이 비교는 TGF-β와 ανβ6-의존적 유전자의 명백한 기능적 관련성뿐만 아니라 ανβ6 mAb 및 rsTGF-βRII-Ig에 의한 유전자 조절 패턴의 상당한 유사성을 보여주었다. 게다가, ανβ6-차단 mAb를 사용한 Col4A3-/- 마우스의 처리는 TGF-β의 신장 발현을 억제하였다. 이 발견은 억제 ανβ6 mAb의 질환-개선 효과가 가능하게는 병든 조직에서 잠복 TGF-β의 ανβ6-매개 활성화 억제를 통한 TGF-β 기능의 억제로부터 기인할 수 있다는 것을 보여준다. 상기 데이타의 한 흥미로운 양태는 ανβ6 또는 TGF-β의 차단을 통한 전구-염증성 유전자 발현의 억제이다. TGF-β는 소염 기능 및 면역억제 기능을 잘 확립하였지만, 본 발명자들의 실험에서 rsTGF-βRII-Ig 및 항-ανβ6 mAb에 의한 유전자 조절의 패턴은 알포트 질환 모델에서 TGF-β의 전구-염증성 기능을 표시하였다. 이 명백한 전구-염증성 효과의 실제 기작은 더 조사될 필요가 있지만, 상기 기작은 상피 세포의 성장 정지 및 사멸을 유도하는 TGF-β의 공지된 능력에 기초할 수 있다31,49-51. 상피 손상이 조직 손상에 대한 초기 선천적 면역 반응의 활성화에 중요한 기작을 제공하기 때문에, 본 발명자들의 데이타에 의해 시사되는 TGF-β의 명백한 전구-염증성 기능은 간접적인 것일 수 있고 신장 상피에 대한 TGF-β-촉진 손상에 의해 매개될 수 있다. 이 모델에 따르면 ανβ6은 상피 리모델링의 TGF-β-의존적 기작의 중요한 성분으로서 작용할 수 있고, 질환에 있어서 그의 기능의 오조절은 질환-관련 조직 손상 및 염증을 더 촉진할 수 있다.
본 발명자들의 연구 결과는 ανβ6이 인간 신장 질환에서 고도로 상향조절되어 있고 기능-차단 항체를 사용한 ανβ6의 표적화가 신장 섬유화증의 치료적 조절에 대한 효과적인 새로운 방법을 제공할 수 있다는 것을 입증한다. ανβ6의 발현이 병든 조직에서 상피 세포에 주로 한정되어 있기 때문에, 이 방법은 TGFβ-기능의 선택적인 국소적 억제를 허용한다. TGFβ는 다양한 유형의 세포 및 조직에서 발현되고, 다수의 상이한 항상성 과정을 조절하는 데 있어서 중요한 역할을 하기 때문에, ανβ6 기능의 차단은 ανβ6 인테그린이 상향조절되어 있는 질환에서 TGF-β의 전신적 억제에 대한 잠재적으로 보다 안정한 대안을 제공한다.
결론
ανβ6은 염증성 및 섬유화증성 병리학적 상태와 관련된 인간 신장 질환에서 과다발현된다.
ανβ6-차단 mAb는 신장 섬유화증의 Col4A3-/- (알포트) 모델에서 섬유화증을 억제한다.
지연된 치료 연구는 ανβ6의 치료적 차단이 신장 섬유화증의 진행을 억제할 뿐만 아니라 기존 섬유화증성 병소를 찾을 수 있게 한다.
β6의 유전적 넉아웃은 Col4A3-/- 마우스에서 보호를 이끌어 낸다.
신장 조직의 전사체 프로파일링은 ανβ6 차단 mAb가 섬유화증 및 염증 매개자의 발현에 있어서 질환-관련 변화를 유의하게 억제한다는 것을 보여주었다.
전사체 조절의 유사한 패턴이 재조합 가용성 TGF-βRII 처리에서 나타났는데, 이는 ανβ6과 TGF-β의 공유된 조절 기능을 암시한다.
참고문헌:
실시예 14: 신장 섬유화증의 마우스 편측 요관 폐쇄 모델에서의 뮤린 3G9(mu3G9)의 효능
요약
편측 요관 폐쇄(UUO)는 가속화된 신장 세뇨관간질성 섬유화증을 초래하는 신장 손상의 잘-확립된 동물 모델이다. 요관 폐쇄는 신장 실질 손상을 초래하는 요관 및 신장 세뇨관의 관내압을 증가시켰다. UUO는 수신증, 세뇨관 확장, 신장 세뇨관 아폽토시스, 진행성 신장 위축증, 간질성 세포 침입, 신장 TGF-β의 증가 및 신장 간질성 섬유화증을 특징으로 한다1. 인테그린 ανβ6 기능은 TGF-β 활성화 및 상피-중간엽 변환 과정에 참여할 수 있다. 이 과정은 질환의 진행에 기여하기 때문에, UUO모델을 사용하여 급속 진행성 신장 섬유화증에 대한 항-ανβ6 단일클론 항체 mu3G9 및 재조합 가용성 뮤린 TGF-β 수용체 타입 II-Ig 융합 단백질(rsTGF-βRII-Ig)의 효능을 평가하였다. 10일간의 실험 기간 동안 주당 3회씩 4 mg/kg의 양으로 복강내로 투여된 mu3G9를 사용한 평활근 액틴 염색의 평균 억제율(%)은 32%이었다.
도입
진행성 섬유화증은 말기 신장 질환을 발병시키는 공통된 과정이고 상피 리모델링, 섬유아세포 활성화, 염증, 및 세포와 세포외 매트릭스(ECM)의 상호작용의 재조직화에 의해 촉진된다. 이러한 과정에 기여하는 분자적 기작은 복잡하고 TGF-β 축의 오조절, 비정상적인 ECM 리모델링, 및 인테그린 거대족1-5의 세포 부착 수용체들의 변경된 발현 및 기능을 포함한다2-9. 최근 연구는 상피 세포 및 중간엽 세포에서 여러 인테그린 및 관련 분자들의 중요한 조절 기능을 밝혔다8,10-13.
신장 질환에서 그 발현이 크게 증가되는 인테그린들 중에는 TGF-β-유도성 인테그린 ανβ6이 있다3,14,15. ανβ6의 발현은 일반적으로 상피 세포에 한정되어 있고, 상피 세포에서 ανβ6은 정상 성체 조직에서 낮은 수준으로 발현되며 발달 과정, 손상 및 신생물에서 상승된다14,16-18. ανβ6이 건강한 성체 신장에서 비교적 낮은 수준으로 발현된다 하더라도, 그의 발현은 발달 중인 마우스 신장, 특히 근위 세뇨관, 헨레 고리 및 집합관에서 현저하다16,17,19. 최근에, ανβ6의 상승된 발현은 다양한 형태의 인간 신장의 병리학적 상태에 대해 보고되어 있다15.
조직 리모델링 동안 생체 내에서의 ανβ6의 증가된 발현과 일치하여, 배양된 상피 세포에서 ανβ6 인테그린의 발현은 EGF 및 TGF-β를 비롯한, 상피 리모델링을 조절하는 사이토카인에 의해 유도될 수 있다3,14. 게다가, 형질전환 마우스의 피부에서의 β6의 과다발현은 자연발생적 만성 상처의 형성을 야기하는 것으로 밝혀졌고20, 이는 ανβ6이 상피 조직 리모델링의 조절에 있어서 중요한 역할을 수행할 수 있음을 암시한다.
ανβ6에 대한 공지된 리간드에는 피브로넥틴, 테나신 및 잠복 관련 펩티드 1 및 3(LAP1 및 LAP3), 및 TGF-β1 및 TGF-β3의 잠복 전구체 형태들의 N-말단 단편이 포함된다21-25. 이들 리간드들과의 결합 결과, ανβ6은 잠복 TGF-β의 세포 부착, 퍼짐, 이동 및 활성화를 매개할 수 있다. TGF-β는 성숙 활성 C-말단 TGF-β 사이토카인과 비-공유적으로 연결된 N-말단 LAP로 절단되어 분비되는 잠복 단백질로서 합성된다. 잠복 TGF-β 복합체는 그의 동족 수용체에 결합할 수 없으므로, 프로테아제에 의한 절단, 낮은 pH 또는 이온화 방사선에의 노출, 및 상기 잠복 복합체가 그의 동족 수용체에 결합할 수 있게 하는 상기 잠복 복합체의 구조적 변화를 포함하는 여러 대체가능한 기작들 중 하나에 의해 활성 형태로 전환될 때까지 생물학적 불활성 상태로 존재한다26-29. 활성화시키는 구조적 변화는 LAP1 및 LAP3 내에 함유된 RGD 모티프와 인테그린의 직접적인 결합을 수반하는 ανβ6에 의해 유도될 수 있다. 이 결합은 TGF-β 전구체를 수용체 결합-능력 상태로 전환시킨다22,25. 이 발견은 상피 세포의 표면 상의 ανβ6 발현의 상향조절이 국소 TGF-β 활성화를 이끌어낸 후, 방관자 세포에서의 TGF-β-의존적 과정의 주변분비 활성화를 이끌어낼 수 있다는 것을 시사한다.
TGF-β가 신장 섬유화증의 중추적인 조절자로서 관여하기 때문에, 본 발명자들은 ανβ6에 의한 그의 국소적 활성화가 신장 질환의 발병시기 및 진행에 있어서 중요한 과정일 수 있고 ανβ6 기능의 차단이 신장 섬유화증의 발병을 억제할 수 있다는 가설을 세웠다. 본 명세서에 기재된 연구에서, 본 발명자들은 신장 섬유화증의 마우스 편측 요관 폐쇄 모델에서 유의하게 상향조절되어 있음을 보여준다. 본 발명자들은 ανβ6의 리간드 결합 및 TGF-β 활성화를 차단하는 mAb가 이 모델에서 섬유화증을 억제한다는 것을 보여준다30.
재료 및 방법
1. 동물 . 8 내지 12주령의 25.5 ± 0.2 g의 바이러스 항원-무함유 수컷 C57BL 마우스(Jackson Laboratories, Bar Harbor, ME)를 본 연구에 사용하였다. 동물들은 바이오겐 아이덱 바이러스-무함유 실험 동물 시설 내에서 환기된 격리 우리 선반에서 사육하였고 본 연구를 시작하기 전에 7일 동안 수용하였다. 수용 및 실험의 전체 기간 동안 마우스는 방사선을 조사받은 표준 마우스 먹이(LabDiet Prolab® 5P75 Isopro® RMH 3000) 및 멸균수에 자유롭게 접근하였다. 체중은 동물 건강 모니터링의 일부로서 주기적으로 측정하였다.
2. 항체 및 시약 . ανβ6 mAb는 본 명세서 및 종래 보고된 바와 같이 생성하였다30. 인간/마우스 키메라 2A1 및 3G9 cDNA는 제1 가닥 cDNA 합성 키트(Amersham/Pharmacia, Piscataway, NJ)를 사용하여 중쇄에 대한 불변 영역 프라이머 CDL-739 및 경쇄에 대한 불변 영역 프라이머 CDL-738로 각각의 모 하이브리도마 총 RNA로부터 생성하였다. 중쇄 및 경쇄 가변 영역 유전자는 cDNA 합성에 사용되는 동일한 3' 프라이머, 대부분의 뮤린 항체 유전자 신호 서열(요청 시 이용가능한 서열)에 특이적인 축퇴 프라이머의 풀 및 Pfu DNA 중합효소(Stratagene, La Jolla CA)를 사용하여 중합효소 연쇄 반응으로 증폭시켰다. 클로닝된 중쇄 및 경쇄 가변 영역들을 인간 IgG1 불변 영역과 함께 포유동물 발현 벡터 내로 연결시켰다. 재조합 가용성 뮤린 TGF-β 수용체 타입 II-Ig 융합 단백질(rsTGF-βRII-Ig)은 종래 보고된 바와 같이11 생성하였고 R&D 시스템스(532-R2, Minneapolis, MN)로부터 구입하였다.
3. 면역조직화학 . 조직 절편으로부터 파라핀을 크실렌 및 에탄올 중에서 제거하고, 증류수 중에서 재수화시킨 후, 0.45% H2O를 함유하는 메탄올에 함침시켰다. 조직을 펩신(00-3009, Zymed, San Francisco, CA)과 함께 항온처리하고 아비딘 및 바이오틴(SP-2001; Vector Laboratories, Burlingame, CA)으로 차단하였다. 1차 항체를 0.1% BSA가 함유된 PBS 중에서 희석시키고 조직을 4℃에서 밤새 항온처리하였다. 마우스 조직 상의 β6을 면역염색하기 위해, 절편을 항-ανβ6 mAb의 인간/마우스 키메라 형태, 즉 2A130, 및 항-인간 바이오티닐화 2차 항체(PK-6103, Vector Laboratories, Burlingame, CA)와 함께 항온처리하였다. 마우스 조직 상의 β6을 면역염색하기 위해, 절편을 2A130 및 항-마우스-바이오티닐화 항체(PK-6102, Vector Laboratories)와 함께 항온처리하였다. 아비딘-바이오틴 복합체-호스라디쉬 퍼록시다제(Vector Kit, PK-6102)를 절편에 가하고, 실온에서 30분 동안 항온처리하고, 3,3'-디아미노벤지딘(DAB) 기질을 지시대로 준비하고(SK-4100, Vector Laboratories), 실온에서 5분 동안 절편에 가하였다. 조직 절편을 메이어스 헤마톡실린으로 1분 동안 염색하고 물 및 PBS 중에서 린싱하였다.
4. 신장 섬유화증의 편측 요관 폐쇄 유도 . 연구를 위한 수술은 2일에 걸쳐 수행하였고, 마우스에 대한 투여 스케쥴은 요관 연결을 한 날과 관련하여 조절하였다. 좌측 요관은 케타민:크실라진(1000:10 mg/kg 피하) 마취 하에 좌측-중간 개복술을 통해 무균 상태로 단리하였다. 2개의 단단한 폐색 6-0 명주 봉합사를 신장의 하부 극(lower pole)의 수평선에서 요관 상에 배치하였고 봉합사 사이에서 요관을 절개하였다. 복부 벽을 4-0 비크릴(Vicryl) 봉합사로 봉합하고 피부를 4-0 나일론으로 봉합하였다. 동물을 가열 패드 상에서 회복시키고 0일 및 1일째 날에 매일 2회씩 0.05 mg/kg의 부프레노르핀을 피하 투여하였다. 동물은 수술 전 날부터 mu3G9를 주당 3회씩 또는 sTGF-βRII-Ig를 주당 2회씩 투여받았다. 절차는 앞서 기재된 보고로부터 채택되었다31.
5. 조직 샘플 채취 및 질환의 지표에 대한 조직학적 분석 . 동물을 봉합 후 10일째 날에 이산화탄소로 마취시켰다. 양쪽 신장(좌측은 연결되었으나, 우측은 연결되지 않음)을 적출하고 신우의 중심을 통해 횡단하여 이등분하였다. 고정된-조직 염색을 위해 각 신장의 한쪽 절반을 10% 중성 완충 포르말린에 넣었다. 각 신장의 나머지 절반을 15% 수크로스에 넣은 후, 평활근 액틴의 면역조직화학적 염색을 위해 30% 수크로스에 넣었다.
콜라겐 함량을 측정하기 위해, 포르말린-고정 신장 절편을 매손 트리크롬 염색으로 염색하였고, 해부학적 구조를 조사하기 위해 H & E로 염색하였다. 트리크롬-염색 절편을 레이카 퀸 화상 분석 시스템(leica Qwin image analysis system)을 사용한 명시야 현미경으로 포착한 화상에서 형태계측학적으로 정량하였다. 표준화된 조명 조건 및 디지탈 카메라 노출 셋팅을 이용하여 화상을 포착하고, 백그라운드에 대해 수정하고 거리 표준에 대해 보정하였다.
매손 트리크롬-염색 슬라이드에서 콜라겐 염색에 대해 어두운 청색이 검출되도록 역치를 설정하였다. 콜라겐 영역은 200X에서 촬영된 화상에서 분석되었다. 전체 좌측 신장 절편을 덮는 인접한 필드의 화상을 정량을 위해 각 동물로부터 얻었다.
6. 통계적 분석 . 측정된 각각의 필드에서의 콜라겐 함량은 200X 필드(임의의 백색 공간을 제외함) 내의 총 조직 면적의 비율, 즉 청색 면적(%)으로서 표시되었다. 16 내지 35개의 개별 필드를 각 신장으로부터 측정하였다. 이들은 절편으로부터의 모든 피질 및 수질 조직을 포함하였고 신장 유두를 배제하였다. 좌측 연결 신장에서 모든 필드로부터 얻은 평균 청색 면적(%)을 각 동물에 대해 계산하고, 통계적 시험을 위한 동물의 섬유화증 스코어로서 사용하였다. 여러 처리군들 중에서 청색 면적(%)에서의 처리-관련 차이의 통계적으로 유의성은 일방(oneway) 편차 분석 후 쌍별 다중 비교에 대한 스튜던트-뉴만-컬스(Student-Newman-Keuls) 절차로 결정하였다. 차이는 p<0.05일 때 통계적으로 유의한 것으로 간주되었다.
결과
1. UUO 후 신장에서의 α ν β 6 의 발현 . UUO 후의 신장에서의 ανβ6의 발현은 면역조직화학적 분석에 의해 조사되었다. 최소한의 ανβ6 발현이 손상되지 않은 (정상) 신장에서 검출되었지만, 유의하게 상향조절된 발현은 UUO 후 7일, 10일 및 14일째 날에 나타났다(도 41). 검출가능한 발현은 UUO 후 3일째 날만큼 초기에 측정되었다.
2. mu3G9 처리를 사용한 UUO 신장에서의 평활근 액틴 면역염색의 억제. 뮤린 UUO 모델에서의 섬유화증의 정도는 면역조직화학적 α-평활근 액틴 염색(갈색 염색) 또는 매손 트리크롬 콜라겐 매트릭스 염색(청색 염색)의 조직형태계측학적 분석에 의해 측정되었다. 각각의 연구에서, 섬유화증이 차지하는 조직 면적의 비율은 비히클 또는 이소타입 대조군 mAb(음성 대조군)를 투여받은 UUO 마우스, 시험 치료적 처리를 제공받은 군 및 연결되지 않은 정상 마우스에서 측정되었다. 치료 효과는 음성 대조군 마우스와 연결되지 않은 정상 마우스 사이의 차이에 대한, 음성 대조군과 시험 치료적 처리군 사이의 염색된 면적의 차이의 비를 계산함으로써 매손 트리크롬 콜라겐 매트릭스(청색) 염색 또는 평활근 액틴(갈색) 염색의 억제율(%)로서 표현된다.
다수의 연구에 걸친 결과들을 연관시키기 위해, 치료 효과는 α-평활근 액틴 염색의 억제율(%)로서 표현된다. 10일간의 실험 기간 동안 mu3G9를 주당 3회씩 4 mg/kg의 양으로 복강내로 투여한 경우, 평활근 액틴 염색의 평균 억제율(%)은 32.8%인 반면, rsTGF-βRII-Ig는 2 mg/kg에서 13.2%의 평균 억제율(%)을 보였다(표 14.1).
[표 14.1]
다수의 연구로부터의 데이타를 조합하는, mu3G9 및 rsTGF-βRII-Ig에 대한 α-평활근 액틴 염색의 평균 억제율(%)
[표 14.2]
[표 14.3]
[표 14.4A]
[표 14.4B]
[표 14.5]
[표 14.6]
[표 14.7]
[표 14.8]
참고문헌
실시예 15: 방사선조사-유도 폐 섬유화증의 뮤린 모델에서의 억제성 항-α νβ6의 용도
도입
폐 섬유화증은 폐 손상 후 매트릭스 리모델링에 장애가 생겼을 때 일어난다((Chapman 2004). 폐 섬유화증에서 이상조절되는 많은 신호전달 인자들 중에서, 사이토카인 TGFβ는 특히 중요한 역할을 한다. 동물 모델에서 TGFβ 신호전달의 억제는 폐, 신장, 간 및 피부에서 섬유화증을 예방한다.
세포내 가공 후, TGFβ 및 그의 프로도메인은 비공유적으로 연결된 복합체로서 분비된다(Annes, Munger et al., 2003). 그의 프로도메인에 결합된 TGFβ는 잠복 상태에 있으므로, 즉 TGFβ는 TGFβ 수용체에 결합할 수 없으므로, 프로도메인은 잠복-관련 펩티드(LAP)로서 지칭된다. LAP는 TGFβ 억제제로서 작용하는 것 이외에 이황화 결합을 통해 잠복 TGFβ 결합 단백질(LTBP) 족의 단백질과 상호작용한다. LTBP는 매트릭스 단백질이며, ECM에서 잠복 TGFβ를 고정시킨다. LAP로부터의 TGFβ의 방출, 즉 잠복 TGFβ 활성화로 지칭되는 과정은 TGFβ 신호전달 경로에서 필요한 단계이다. 이 활성화 단계는 TGFβ 신호전달을 감소시키는 방법에 대한 잠재적 표적이다.
인테그린 ανβ6 및 ανβ8은 각 LAP의 C-말단 근처에 위치한 RGD 아미노산 서열과 상호작용함으로써 잠복 TGFβ1 및 TGFβ3을 활성화시킨다(Munger, Huang et al., 1999; Annes, Rifkin et al., 2002; Mu, Cambier et al., 2002). (최종 TGFβ 이소폼인 TGFβ2는 RGD 서열을 가지지 않으며 이 인테그린들에 의해 활성화될 수 없다). 폐 내에서, 인테그린 ανβ6에 의한 TGFβ 활성화는 항상성 및 손상에 대한 반응에서 대체될 수 없는 역할을 수행한다. ανβ6은 정상 폐 상피에서 소량으로 발현되나, 손상 후 급속히 상향조절된다. β6 유전자가 결핍된 마우스(Itgb6-/-)는 감소된 TGFβ 신호전달로 인해 폐염증 및 폐기종을 발병시킨다. 블레오마이신에의 마우스 폐의 노출은 ανβ6 발현에서의 큰 증가에 동반되는 급성 폐 손상을 야기한 후, TGFβ-의존적 폐 섬유화증을 야기하며; Itgb6-/- 마우스는 블레오마이신 처리 후 폐 섬유화증을 발병시키지 않는다.
이온화 방사선은 폐 섬유화증을 야기한다(Franko and Sharplin 1994; Movsas, Raffin et al., 1997; Martin, Lefaix et al., 2000; Abratt, Morgan et al., 2004). 섬유화증이 폐 손상 후 수일 내에 시작되는 블레오마이신 폐 섬유화증 모델과 대조적으로, 방사선조사-유도 폐 섬유화증(RILF)은 손상 후 수개월 내에 시작된다. 뮤린 RILF는 종-의존적이고; 본 실험들에서 사용된 C57BL/6 종은 감수성을 가진다. 뮤린 RILF 모델에서, 대략적으로 섬유화증의 발병 시점에서 일어나는 실질적인 말기 사망도 있으며; 이 사망은 아마도 폐 관류의 상실로 인한 것으로 보인다(Franko, Nguyen et al., 1996; Haston, Zhou et al., 2002). 억제성 항-ανβ6 mAb(3G9)가 개발되었다(Weinreb, Simon et al., 2004). 이 연구들의 목적은 (1) 흉선 방사선에 대한 Itgb6+/+의 반응과 Itgb6-/-의 반응을 비교함으로써 감수성 마우스 종에서 RILF의 ανβ6-의존성을 확립하는 것, (2) 방사선을 조사받은 마우스를 TGFβ 길항제(가용성 TGFβ 수용체)로 처리함으로 뮤린 RILF 모델의 TGFβ-의존성을 확인하는 것, 및 (3) 방사선을 조사받은 마우스에게 투여된 다양한 투여량의 3G9의 효과를 평가하는 것이었다.
재료 및 방법
1. 동물. 사용된 모든 동물은 암컷이었다. Itgb6-/- 마우스는 딘 쉐파드(UCSF)로부터 제공받아 C57BL/6 백그라운드에 대한 본 출원인의 시설에서 사육되었다. 야생형 마우스는 잭슨 레이보레이토리(Jackson Laboratory, Bar Harbor, ME)로부터 구입한 C57BL/6이었고, 도착 시 7 내지 9주령이었으며, 방사선조사 전에 본 출원인의 동물 시설에서 1주일 동안 적응시켰다. 모든 동물 취급 절차 및 실험은 뉴욕 의과대학의 동물 보호 위원회로부터 승인받았고 실험 동물의 보호 및 사용에 대한 NIH 지침에 따라 수행하였다. 우리는 동물 시설 프로토콜 당 우리 당 최대 5 마리의 마우스로 한정되었다. 마우스는 이환률 및 사망률에 대해 매일 모니터링되었다. 죽어가는 마우스는 희생되었다.
2. 항체 . 2개의 항체를 바이오겐 아이덱(Cambridge, MA)으로부터 입수하고, 본 명세서의 다른 부분에 기재된 바와 같이 준비하였다. 시험된 1차 항체인 3G9는 ανβ6-매개 TGFβ 활성화를 차단하는 항-ανβ6 mAb이다. 상기 항체는 IgG1 하위유형이며, 양이온-독립적 방식으로 ανβ6에 결합한다. 대조군 Ab(1E6)는 마우스 LFA-3과 상호작용하지 않는 마우스 항-인간 LFA-3 IgG1 단일클론 Ab이었다. 정상 마우스에서 4주 동안 최대 200 mg/kg/주의 상기 대조군 Ab의 투여량은 독성을 전혀 나타내지 않았다(바이오겐 아이덱으로부터 얻은 데이타). 항체 분취액은 총 부피 200 ㎕ 중의 0.3, 1, 3, 6 및 10 mg/kg의 투여량을 달성하도록 멸균 PBS 중의 희석액으로서 준비하였다. 실험에 따라 우측 측면에서의 피하 주사 또는 복강내 주사를 방사선조사 후 15주가 되는 시점에서 시작하였다.
3. 방사선 프로토콜 . 마우스를 8 내지 10주령에서 방사선조사하였다. 방사선조사 전, 복강내로 전달된 15 ㎕/g의 2.5% 용액으로 아버틴(Avertin)(2,2,2-트리브로모에탄올; Acros Organics, NJ)을 사용하여 마취를 달성하였다. 그 후, 테이프를 사용하여 마우스를 플렉시글라스 표면 상에 눕혔다. 납 차폐를 적절하게 위치시켜 방사선이 흉선에만 조사되게 하였다. 폐 첨부 수평면부터 검상 돌기까지의 필드는 1.8 cm이었다. 60Co 공급원을 사용하여 14-Gy 방사선을 전달하였다. 공급원으로부터 피부까지의 거리는 65 cm이었다. 이 공급원을 사용한 노출 시간은 약 11분이었다. 방사선에 노출시킨 후, 마우스들을 그들의 우리로 돌려보내고 대면하도록 위치시키고 회복 기간 동안 모니터링하였다.
4. 희생된 마우스로부터의 샘플 채취 . 방사선조사 후 소정의 시점(항체 연구의 경우, 26주, 28주 또는 32주)까지 생존한 마우스를 희생시키고 하기 방식으로 처리하였다. 아버틴으로 깊은 마취를 달성한 후, 70% 에탄올을 흉선 및 복부 피부 상에 분무하였다. 횡경막을 통해 흉강을 개방하였다. 심실로부터 400 내지 500 ㎕의 혈액을 직접 흡입하였다. 기관을 노출시키고 22-게이지 혈관도관(angiocatheter)으로 기관에 캐뉼러를 꽂았다. 폐를 700 ㎕의 PBS 분취액으로 2회 세척하였다. 우측 주관(mainstem) 기관지를 문(hilum)에서 결찰시키고, 각 엽(lobe)을 제거하고 분리된 튜브 내에 넣고, 액체 질소 중에서 급속히 동결시키고, -80℃에서 저장하였다. 좌측 폐를 400 ㎕의 10% 포르말린으로 팽창시키고, 10% 포르말린 중에서 밤새 넣어 두고, 파라핀 중에 포매시켰다. 희생 시 얻은 기관지-폐포 세척액(BAL)을 2개의 분취액, 즉 200 ㎕와 나머지로 나누었다. 튜브 둘다를 200 RPM으로 3분 동안 원심분리하였다. 두 튜브로부터 얻은 상청액을 합하고, 액체 질소 중에서 동결시키고, -80℃에서 저장하였다. 보다 큰 튜브로부터 얻은 세포 펠릿을 급속 동결시키고 동일한 방식으로 저장하였다. 200 ㎕의 분취액 세포 펠릿을 200 ㎕의 RBC 용해 완충제로 재현탁시키고 철저히 혼합하였다. 50 ㎕를 혈구계에서의 세포 계수에 사용하였다. 남은 150 ㎕를 사이토스핀(cytospin) 제조에 사용하였다. 심장 천자로 얻은 혈액을 사용하여 다음과 같이 혈청 또는 혈장을 수득하였다. 캐피젝트(Capiject) 튜브 또는 헤파린으로 처리된 1.5 cc 마이크로원심분리 튜브 중에서 먼저 혼합한 후, 샘플을 14,000 RPM에서 20분 동안 원심분리하였다. 상청액을 제거하고 -80℃까지 즉시 동결시켰다.
5. 죽은 상태로 발견된 또는 죽어가는 마우스로부터의 샘플 채취 . 희생 일이 되기 전에 죽은 상태로 발견된 또는 죽어가는 마우스를 해부하여 폐 표본을 얻었다. 죽어가는 마우스를 해부 전에 아버틴으로 마취시켰다. 횡경막을 통해 흉강을 개방하였다. 기관까지의 해부는 완전히 노출된 상태에서 수행하였다. 22-게이지 혈관도관으로 기관에 캐뉼러를 꽂았다. 폐를 800 내지 1000 ㎕의 10% 포르말린으로 팽창시켰다. 흉강의 내용물을 일괄적으로 제거하고, 폐 분리 및 파라핀 포매 전에 24시간 동안 10% 포르말린 중에 넣어 두었다.
6. 세포 분화 . 사이토스핀 제제를 디프퀵(DiffQuik) 방법(고정액, 1% 에오신-Y, 1% 아주르(Azure) A 및 탈이온수 중에 순차적으로 15-초 동안 함침시킴)으로 염색하였다. 그 후, 슬라이드를 탈수시키고 현미경에 장착시켰다. 호중구, 림프구 및 대식세포의 수를 2개의 분리된 고배율(400x) 필드에서 수동으로 세었다.
7. 면역조직화학 . 일부 포르말린-고정 샘플을 β6의 면역조직화학적 검출에 사용하였다. 내인성 퍼록시다제 활성을 메탄올 중에서 15분 동안 3% 과산화수소로 켄칭하였고, 항원 보충은 다이제스트-올 3 펩신(Digest-All 3 Pepsin; Zymed, South San Francisco)을 5 내지 7분 동안 가함으로써 달성하였다. 아비딘/바이오틴 차단 용액(Vector Laboratories, Burlingame, CA)을 제조자의 지시에 따라 사용하였다. 0.5% 카세인 용액으로 15분 동안 차단하였다. 인간 IgG 내로 클로닝된 마우스 항-β6 mAb의 가변 영역으로 구성된 항-β6 단일클론 항체 ch2A1(Biogen Idec)을 0.1% BSA 중에서 1:500 희석비로 실온에서 1시간 동안 사용하였다. 제조자의 지시에 따라 항-인간 2차 항체와 함께 벡타스테인(Vectastain) ABC 키트(Vector Laboratories)를 사용하여 검출하였다. DAB 키트(Sigma, St. Louis, MO)를 사용하여 크로모겐(Chromogen) 발생을 수행한 후, 헤마톡실린 반대염색을 수행하였다. 상기 절차의 특이성은 동시에 처리된 절편에서의 1차 항체의 생략(이는 통상적으로 수행됨) 및 Itgb6-/- 마우스로부터 얻은 폐 절편에 대한 예비 시험(이는 음성 결과를 제공함)에 의해 확인되었다.
8. 폐 절편, 트리크롬 염색 및 % 섬유화증 측정 . 포르말린으로 고정되고 파라핀에 포매된 폐를 5-마이크론 두께로 횡으로 절개하였다. 가시적 평가를 통해 폐 절편을 문에서 또는 문 근처에서 수득하였다. 슬라이드로부터 파라핀을 탈이온수로 제거하고(각각 3분 동안 크실렌 욕조 x 2, 각각 3분 동안 100% 에탄올 x 2, 각각 3분 동안 95% 에탄올 x 2, 3분 동안 70% 에탄올, 3분 동안 탈이온수). 절편을 매손 트리크롬 염색으로 염색하였다(매염 상태까지 밤새 보우인(Bouin) 용액, 5분 동안 웨이게르트(Weigert) 철 헤마톡실린, 5분 동안 비에브리크 스카렛-산 푸친(Biebrich's Scarlet-Acid Fucshin), 5분 동안 포스포텅스틱(Phosphotungstic)/포스포-몰리브딕 산 용액, 5분 동안 아닐린 블루 용액, 중간에 탈이온수를 사용한 린스, 및 2분 동안 아세트산 1%). 이어서, 슬라이드를 탈수시키고(70% 에탄올, 95% 에탄올, 100% 에탄올 및 크실렌) 퍼마운트(Permount)를 사용하여 현미경에 장착하였다.
문헌[Haston et al., (Cancer Res 2002, 62: 372-8)]에 기재된 % 섬유화증 기법을 이용하였다. 트리크롬으로 염색된 폐 절편의 저배율(2X 내지 3X) 화상을 얻어 디지탈 포맷으로 저장하였다. 폐 절편의 고배율(100X 내지 200X) 수동 검사를 광학 현미경으로 수행하여 (증가된 콜라겐 침착 및 구조의 상실로 정의된) 섬유화증 영역을 확인하였다. 폐 절편의 총 횡단 면적과 함께 섬유화증의 횡단 면적을 NIH 화상 1.62 소프트웨어를 사용하여 디지탈 화상으로 윤곽을 그렸다. 섬유화증 면적의 합계를 폐의 총 면적으로 나누어 % 섬유화증을 확립하였다. 폐의 한 무작위 횡단면은 10개의 무작위 절편과 비교될 때 전체 폐에서의 % 섬유화증을 반영하는 것으로 밝혀졌다(Haston, Amos et al., 1996).
9. 히드록시프롤린 측정 . 히드록시프롤린 함량의 측정은 레디(Reddy) 및 엔웨멤카(Enwememka)의 방법으로부터 채택되었다. 우측 폐를 분리하여 중량을 측정하였다. 폐 조직(약 20 mg의 습윤 중량)을 400 ㎕의 2N NaOH 용액 중에서 12시간 동안 (실온) 항온처리한 후, 균질화하였다. 균질화물 및 표준 히드록시프롤린 용액을 120℃까지 30분 동안 가열하여 가수분해하였다. 클로르아민-T 용액(0.056 M, 450 ㎕)을 50 ㎕의 가수분해물에 첨가하고, 25분 동안 산화를 진행시켰다. 에를리히(Ehrlich) 시약(1 M, 500 ㎕)을 첨가하고, 65℃에서 20분 동안 색상이 나타나게 하였다. 그 다음, 550 nm에서 흡광도를 측정하였다. 최종 농도는 ㎍ 히드록시프롤린 / mg 습윤 폐 조직으로서 표현된다.
10. RV/LV 질량 비 측정 . 본 실험에서 사용된 마우스는 방사선조사 후 32주에서 희생된 무리 중에 있었고, 방사선이 조사되지 않은 7마리의 추가 C57BL/6 마우스를 대조군으로서 사용하였다. 방사선을 조사받은 마우스로부터 얻은 심장은 28주와 32주 사이에 죽은 마우스 또는 32주째에 희생될 때까지 생존한 마우스로부터 얻은 것이었다. 심장은 포르말린 중에서 고정시켰다. 해부 현미경 하에, 심방을 제거하고, 우측 심실 자유벽(RV)을 좌측 심실 및 사이막(LV)으로부터 절개하여 떼어내었다. 심실 조직의 조각 각각의 중량을 측정하고, 상기 비를 계산하였다. 방사선이 조사되지 않은 7마리의 C57BL/6 마우스를 대조군으로 사용하였다.
11. 통계 . 군 사이의 % 섬유화증 차이의 통계적 유의성은 비-매개변수적 데이타에 대한 만-휘트니(Mann-Whitney) 검정을 이용하여 수행하였다. 사망 날짜를 기록하고 이를 이용하여 카플랜-메이에르 곡선을 작도하였다. 개개의 군 비교 뿐만 아니라 카플랜-메이에르 곡선의 총 비교를 로그-순위(Log-rank)(맨텔-콕스(Mantel-Cox)) 검정을 이용하여 수행하였다. 측정치의 평균값은 평균의 상응하는 표준 오차 또는 표준 편차를 이용하여 보고하였다. RV/LV 질량 비 측정치의 평균의 비교를 위해, 스튜던트 t-검정(비쌍별(non-paired), 양방(2-tailed))을 이용하였다. 피셔 정확도 검정을 이용하여 Itgb6-/- 마우스와 Itgb6+/+ 마우스 사이의 섬유화증의 존재 또는 부존재를 비교하였다. 통계적 유의성은 p<0.05로서 정의되었다.
결과
1. 뮤린 RILF는 α ν β 6 발현을 필요로 한다: 흉선 방사선조사 후 Itgb6-/- 마우스와 Itgb6+/+ 마우스의 비교. 본 실험은 기준선에서의 ανβ6 발현과 방사선조사 후의 ανβ6 발현을 비교하고, ανβ6의 부재가 섬유화증을 예방하는 지를 확인하기 위해 디자인하였다. 본 발명자들은 Itgb6-/- 마우스 및 Itgb6+/+ 마우스에게 방사선을 조사하고 이들을 28주 전의 다양한 시점 및 28주째가 되는 시점에서 희생시켰다.
(a) β 6 은 방사선조사 후 18 내지 20주째에 상향조절된다. 방사선조사 후 18주 전에 희생된 마우스는 면역조직화학적 기법에 의한 측정 시 정상적인 낮은 ανβ6 발현을 보였다. 그러나, 18주째에서, 폐포 상피 전체에 걸친 β6의 퍼져 있는 증가된 발현이 관찰되었다(도 42).
(b) 높은 α ν β 6 발현은 섬유화증 영역에서 지속된다. 본 발명자들은 방사선조사 때문에 시간과 관계없이 섬유화증성 병소 내의 상피 세포에서 ανβ6이 높은 수준으로 발현되었음을 일관되게 관찰하였다(도 43). 그러나, 18주째에 관찰된 ανβ6의 퍼져 있는 증가된 발현은 종종 후기 시점에서 덜 분명하였다(도 43, 24주와 27주를 비교함).
(c) α ν β 6 이 결핍되어 있는 마우스는 RILF를 발병시키지 않는다. 대조군 마우스에서, 섬유화증 영역이 식별될 수 있는 최초 시점은 방사선조사 후 20 내지 22주이었다(나타내지 않음). 섬유화증 영역은 전형적으로 잘 식별되었고 늑막 아래에 있었다. 본 발명자들은 방사선조사 후 27주째에 희생된 Itgb6-/- 마우스(N=17)의 폐 절편에서는 섬유화증의 영역을 전혀 발견하지 못하였다. 대조적으로, 본 발명자들은 23마리의 Itgb6+/+ 마우스 중 21마리의 절편에서 섬유화증 영역을 발견하였고, 이는 통계적으로 유의한 차이이다(p<0.001, 양방 피셔 정확도 검정)(도 44). Itgb6+/+ 마우스(방사선조사 후 27주)의 절편의 평균 % 섬유화증 영역은 17% +/- 3%이었다. 본 발명자들은 방사선조사 후 27주째에 Itgb6+/+ 및 Itgb6-/- 마우스로부터 얻은 폐의 히드록시프롤린 함량을 측정함으로써 조직학적 발견을 확인하였다(도 45).
(d) α ν β 6 의 부재는 폐 방사선조사 후에 생존에 영향을 주지 않는다. 14 Gy의 흉선 방사선조사 후, 사망률은 방사선조사 후 18주까지 무시될 수 있는 정도이었으나, 방사선조사 후 대략 25주에서 50%에 도달하였다. Itgb6+/+ 마우스의 생존 곡선과 Itgb6-/- 마우스의 생존 곡선 사이에는 유의한 차이가 없었다(도 46).
2. 뮤린 RILF 모델에서 3G9(0.3, 1 및 1 mg/kg/주 IP 투여) 및 가용성 TGFβ R의 효과. 이전 결과는 RILF가 ανβ6 인테그린의 발현에 의존하고, ανβ6의 결핍이 방사선조사 후 사망률을 악화시키지 않는다는 것을 보여준다. ανβ6에 대한 필요성은 잠복 TGFβ1의 활성화제로서의 그의 공지된 기능과 일치한다. 또한, 이 결과는 항섬유화증 방법으로서 ανβ6 억제의 이용가능성을 암시한다. 이 개념을 시험하고 RILF의 뮤린 모델이 TGFβ-의존적임을 확인하기 위해, 본 발명자들은 방사선조사를 받은 마우스를 대조군 Ab, 가용성 TGFβ 수용체, 또는 3가지 투여량의 3G9 중 하나(군당 N=27)로 처리하였다. 보다 적은 수의 마우스(N=15)를 PBS 주사액(200 ㎕)으로만 처리하였다. 3G9는 0.3 1 및 10 mg/kg의 투여량으로 사용되었고, 대조군 Ab는 10 mg/kg의 투여량으로 사용되었으며, 가용성 TGFβ 수용체는 5 mg/kg의 투여량으로 사용되었다. 처리는 방사선조사 후 15주째(ανβ6 상향조절 전 대략 3주째)에 시작하였고 방사선조사 후 26주째에 희생될 때까지 주마다 계속 수행하였다(세부사항은 방법 단락을 참조함).
(a) 3G9 및 가용성 TGFβ 수용체는 섬유화증을 감소시킨다. 모든 생존 마우스를 방사선조사 후 26주째에 희생시켰다. 0.3-mg/kg 군이 대조군에 비해 % 섬유화증에서 감소를 전혀 나타내지 않은 반면, 1-mg/kg 군은 섬유화증에서 유의한 감소를 나타내었다. 가용성 TGFβ 수용체 군 및 10-mg/kg 군도 대조군보다 덜한 섬유화증을 보였지만, 결과는 희생된 마우스의 분석에서만 통계적으로 유의한 수준에 도달하지 못하였다(도 47). 계획된 희생 시점 전에 죽은 마우스는 생존 마우스와 생물학적으로 상이할 가능성이 있다. 그러므로, 본 발명자들은 연구 기간 동안 죽었거나 희생된 모든 마우스에 대해 유사한 분석을 수행하였다. 모든 마우스(희생된 마우스 및 희생 전에 죽은 마우스)를 고려할 때, 대조군에 비해 유의하게 감소된 섬유화증이 1-mg/kg 군, 10-mg/kg 군 및 가용성 TGFβ 수용체 군에서 존재하였다. 0.3-mg/kg 군은 대조군에 비해 유의한 차이를 나타내지 않았다(도 48).
(b) 10-mg/kg 투여량의 3G9는 호중구 및 림프구성 폐포염을 야기한다. 모든 희생된 마우스에 대해 수행된 BAL은 대조군에 비해 10-mg/kg 군에서 호중구 및 림프구의 증가된 비율을 보여주었다(도 49). 다른 군은 유사한 증가를 보이지 않았다.
(c) 보다 낮은 투여량에서 항체를 통한 α ν β 6 의 차단은 폐 방사선조사 후 생존에 영향을 주지 않는다. 모든 군이 비교될 때 생존에서의 유의한 차이는 없었다(p = 0.088; 로그-순위 맨텔-콕스 검정에 의한 모든 군의 비교). 그러나, 10-mg/kg 군에서 사망률이 증가하는 경향이 명백하였다(도 50). 10-mg/kg/주 군과 대조군만이 비교된 경우(즉, 다수의 비교에 대한 보정이 없음), 생존 곡선의 차이는 통계적으로 유의하다.
3. 뮤린 RILF 모델에서의 3G9의 효과(1, 3, 6 및 10 mg/kg/주 피하 투여 ). 이전 결과는 뮤린 모델에서 RILF가 TGFβ에 의해 매개되며 1 mg/kg 및 10 mg/kg 투여량에서 3G9에 의해 유의하게 감소된다는 것을 보여준다. 그러나, 증가된 폐포 염증 및 사망률이 증가하는 경향은 10 mg/kg 투여량에서 존재하였다. 본 실험에서, 본 발명자들은 3G9 처리가 RILF의 발병을 방지하기보다는 수주 동안 RILF의 발병시기를 단순히 지연시킬 가능성을 시험하기 위해 후기 시점(방사선조사 후 최대 32주)에서 섬유화증을 평가하였다. 또한, 폐포 염증의 차이 및 가능하게는 1 mg/kg 투여량과 10 mg/kg 투여량에서의 생존율의 차이를 더 잘 정의하기 위해, 본 발명자들은 1 mg/kg 내지 10 mg/kg의 추가 투여량을 시험하였다. 본 발명자들은 270마리의 마우스에게 방사선을 조사하고 이들을 동등한 군으로 나누어 4가지 투여량(1, 3, 6 및 10 mg/kg) 중 한 투여량의 3G9 또는 대조군 Ab(10 mg/kg)를 투여하였다. 처리는 방사선조사 후 15주째에 시작하여 추후 희생 시점까지 주마다 계속 수행하였다. 본 실험은 약 1개월 간격으로 방사선을 조사받은 마우스로 구성된 2개의 군을 사용하여 수행하였다. 한 군에서, 처리를 시작한 마우스가 방사선조사 후 28주까지 생존하였다면 이들을 희생시켰고(군 1), 다른 군에서 이들이 32주까지 생존하였다면 이들을 희생시켰다(군 2). 항체는 피하 투여되었다(이전 실험에서와 같이 IP가 아님).
(a) 1, 3, 6 및 10 mg/kg의 투여량에서 3G9는 섬유화증을 감소시킨다. 유의하게 감소된 수준의 섬유화증은 대조군에 비해 3G9를 투여받은 모든 군에서 관찰되었다(p<0.01). 이 차이는 죽은 상태로 발견된 또는 죽어가는 마우스, 최종 시점에서 희생된 마우스 및 조합된 모든 마우스에 대해 유의하였다(도 51).
(b) 보다 높은 투여량에서 3G9는 호중구성 및 림프구성 폐포염을 야기한다. 모든 희생된 마우스(N=101)에 대해 수행된 BAL은 대조군에 비해 3-mg/kg, 6-mg/kg 및 10-mg/kg 군에서 호중구 및 림프구의 보다 높은 비율(도 52)을 보였다(p<0.02). 또한, 1-mg/kg 군에 비해 상당히 더 높은 비율이 10-mg/kg 군에서 존재하였다(p<0.001). 정량적으로, "거품 형태의" 대식세포의 증가된 수(Itgb6-/- 마우스에서 관찰된 것과 동일함) 및 세포 데브리스가 보다 높은 투여량에서 관찰되었다(도 53).
(c) 6 mg/kg의 투여량에서 3G9는 감소된 생존율과 관련되어 있다. 대조군을 비해 1-mg/kg 군 및 3-mg/kg 군과 비교할 때, 대조군과 상기 군들 사이에는 사망률의 차이가 없었다(도 54 및 55). 그러나, 대조군에 비해 6-mg/kg 군에서 사망률이 상당히 더 높았다(p < 0.05). 10-mg/kg 군은 대조군 마우스에 비해 생존율에서의 감소를 보이지 않았다. 6-mg/kg/주 3G9 군은 28-주 희생 집단(군 1)과 32-주 희생 집단(군 2) 둘다에서 더 낮은 생존율을 나타내었지만, 차이는 28-주 군에서 더 현저하였다(도 54 및 55).
(d) RV/LV 질량비 및 폐 관류에 대한 폐 방사선조사의 효과. 종래 연구는 폐 방사선조사 후 섬유화증 주기 동안의 사망이 기공 폐쇄(주로 섬유화증에 기인함)와 폐포 관류(Sharplin and Franko 1989)의 상실의 조합으로부터 비롯된 호흡 곤란증에 기인한다는 것을 암시한다. 관류의 상실은 폐 동맥 고혈압 및 우측 심실 비대증을 초래할 것이고, 증가된 RV 질량은 이 모델에서 보고되어 있다. 본 발명자들은 32주까지 생존한 마우스의 RV/LV 질량비와 28주와 32주 사이에서 죽은 동일한 집단의 마우스의 RV/LV 질량비를 측정하였다. 죽은 마우스는 생존한 마우스 및 방사선을 조사받지 않은 마우스에 비해 유의하게 증가된 RV/LV 비를 나타내었다(도 56). 또한, 죽은 일부 마우스에서, 폐포 벽 내에서 적혈구가 완전히 존재하지 않는 영역이 명백히 확인되었고, 이는 폐포 관류의 완전한 상실을 표시하는 발견이다(도 57).
논의
TGFβ는 전구-섬유화증 매개자인 것으로 공지되어 있다. 종래 연구는 인테그린 ανβ6이 잠복 TGFβ1 및 TGFβ3을 활성화시키고 Itgb6-/- 마우스가 블레오마이신-유도 폐 섬유화증으로부터 보호된다는 것을 보여주었다. 여기에 제공된 결과는 Itgb6-/- 마우스가 방사선조사에 의해 유도된 폐 섬유화증으로부터도 보호된다는 것을 의미한다. 이 결과는 ανβ6 인테그린을 표적화하는 항-섬유화증 요법에 대한 이론적 근거를 제공한다.
본 연구에서, 본 발명자들은 1 mg/kg/주 이상의 투여량으로 투여된 3G9가 RILF를 일관되게 효과적으로 감소시킨다는 것을 발견하였다. 보다 높은 투여량, 특히 6 내지 10 mg/kg/주의 투여량은 BAL 액에서 호중구 및 림프구의 증가된 비율과 관련되어 있었다. 이 변화는 Itgb6-/- 마우스의 표현형과 일치하고 상기 투여량의 3G9가 상기 동물의 표현형이 넉아웃 동물의 표현과 동일한 정도까지 ανβ6-매개 TGFβ 활성화를 억제한다는 것을 시사한다.
또한, 보다 높은 투여량의 3G9는 뮤린 RILF 모델에서 감소된 생존과 변동가능하게 관련되어 있다. 첫 번째 시험에서, 10 mg/kg/주의 투여량에서 사망률이 증가하는 경향이 강하였지만, 1 또는 0.3 mg/kg/주의 투여량으로 처리된 마우스에서는 그러한 경향이 없었다. 두 번째 시험에서, 6-mg/kg/주 군에서 사망률이 증가되었지만, 다른 군들(10 mg/kg/주를 포함함)에서는 그러하지 않았다. 투여량/반응에서의 차이에 대한 이유는 명확하지 않지만, 일반적인 결과는 시험된 최대 투여량에서 생존율이 감소되는 경향이 있다는 것이다.
RILF 모델에서 일어나는 사망은 호흡 곤란증으로 인한 것으로 보인다. 사망률은 종-의존적이고 성별-의존적이다(Haston, Zhou et al., 2002). 뮤린 RILF 모델에서 폐 기능장애 및 사망의 원인은 광범위하게 연구되어 왔다(Sharplin and Franko 1989). 이 연구 보고의 저자들은 폐 관류가 방사선조사 후에 감소되고, 이 결손이 사망에 기여할 가능성이 있다는 결론을 내렸다. 또한, 방사선조사에 의해 유도된 폐 섬유화증에 대한 내성을 가진 마우스는 방사선조사에 의해 유도된 폐 섬유화증 모델에서 감소된 생존율을 나타낸다. 이 마우스들에서 감소된 폐 관류는 이 모델에서 그들의 사망에 기여하는 주요 인자인 것으로 밝혀졌다. 관류 결손의 패턴은 종에 달려있다. 섬유화증이 발병되기 쉬운 마우스에서, (희생 30초 전에 정맥 내로 주사된 콜로이드성 탄소의 존재 또는 부재에 의해 판단될 때) 관류의 완전한 상실은 섬유화증 영역 및 섬유화증성 병소에 인접한 간헐적 소영역에 한정되어 있다. 또한, 방사선을 조사받지 않은 대조군보다 방사선을 조사받은 마우스에서 관류된 영역 내의 탄소의 양에서 더 큰 영역-대-영역 변동가능성이 있었는데, 이는 관류가 감소되었으나 부재하지는 않는 실질적인 영역이 있다는 것을 암시한다. 다수의 마우스 종에서 관찰된 폐 섬유화증의 상실에 대한 다른 증거는 병소-부재 폐에서의 작은 혈관의 감소된 수, 및 RV/LV 두께 비에 의한 평가 시 RV 비대증이었다. 또한, 폐포 벽에서의 적혈구의 수(관류를 평가하는 상이한 방법)가 방사선을 조사받은 마우스(A/J 종)에서 감소되었다. 대부분의 C57BL/6 마우스는 흉막 유출을 보이지 않았다. C57BL/10J 종을 제외하고, 심근 손상은 방사선조사의 결과로서 일어나지 않았다.
사망률에 대한 본 발명자들의 관찰결과는 샤플린(Sharplin) 및 프랜코(Franko)의 보다 광범위한 관찰결과와 일치한다. 대조군 마우스에서, 섬유화증의 정도는 희생될 때까지 생존한 마우스보다 죽은 마우스에서 더 높았는데(도 51), 이는 보다 심한 폐 기능이상이 사망과 관련되어 있다는 것을 암시한다. 죽은 마우스는 우측 심실 비대증(RV/LV 질량 지수에서의 증가로서 정의됨)을 가진 반면, 희생될 때가진 생존한 마우스는 그러하지 않았는데(도 56), 이는 폐 관류의 상실이 사망과 관련되어 있음을 암시하는 관찰결과이다. 죽은 상태로 발견된 일부 마우스에서, 폐포 벽 내에 적혈구가 없는 폐 영역이 조직학적 절편 상에서 분명히 나타났는데(도 57), 이는 그 영역에서의 관류의 완전한 상실과 일치하였고 종래 보고와 일치한다(Sharplin and Franko 1989). 본 발명자들은 사망의 원인이 될 수 있는 식도 기능장애(흉터형성, 천공)의 증거를 발견하지 못하였을 뿐만 아니라 심근 괴사도 발견하지 못하였다. 따라서, 종래 연구에 비추어 해석하건대, 본 발명자들의 증거는 심지어 섬유화증이 예방될 때에도 폐 관류의 상실이 C57BL/6 마우스에서 일어나고 섬유화증이 아니라 폐 관류의 상실이 사망의 원인이라는 것을 시사한다. 또한, 본 발명자들은 마우스가 (대조군 Ab 또는 3G9로) 주사한 후 2일 동안 죽을 가능성이 더 높다는 것을 발견하였는데, 이는 취급의 스트레스 및 아마도 주사로부터의 과도한 유체 부피가 의식 상태와 무의식 상태의 경계에 있는 마우스에서 사망을 촉진한다는 것을 암시한다.
유전자 제거에 의한 ανβ6의 상실이 이 모델에서 사망률에 영향을 주지 않을지라도, 높은 투여량의 3G9(6 내지 10 mg/kg/주)는 보다 낮은 투여량의 3G9 또는 대조군 Ab로 처리된 마우스에 비해 더 이른 사망과 관련되어 있었다. 가장 명백한 해석은 보다 높은 투여량의 3G9가 사망률을 악화시킨다는 것이다. 그러나, 본 발명자들은 대조군 Ab 및 보다 낮은 투여량의 3G9가 사실상 생존을 개선할 가능성을 배제할 수 없다. 3가지 실험에 대해 시간 대 50% 생존율의 비교는 2개의 군을 보여주는 듯하다. 야생형 및 Itgb6-/- 마우스(도 46), IP 실험에서의 PBS-처리 및 10-mg/kg/주 3G9-처리 마우스(도 50), 및 SC 실험에서의 6-mg/kg/주 3G9-처리 마우스(도 54)는 약 22.5 내지 25주의 평균 생존 시간을 가진 반면, 모든 다른 처리 군들은 약 24.5 내지 30주의 평균 생존 시간을 가졌다(도 50 및 54). 이 점에 대한 명확한 결론은 내릴 수 없었는데, 이는 실험적 조건이 다르고 다른 조건들이 실험 사이에서 변경되어 기준 사망률의 변화를 초래할 수 있기 때문이다. 6 내지 10 mg/kg/주의 투여량으로 3G9로 처리된 마우스에서, 본 발명자들은 해부된 마우스의 총체적인 검사 또는 폐의 조직학적 검사 시 생존율의 변화를 설명할 (폐 염증 이외의) 새로운 이상증상을 발견하지 못하였다. RILF 모델에서 보다 낮은 투여량의 3G9와 보다 높은 투여량의 3G9 사이의 생존율의 차이에 대한 원인은 알려지지 않았다.
본 연구는 ανβ6-매개 TGFβ 활성화를 최대한 감소시키지 않을 것으로 예측되는 보다 낮은 투여량의 3G9가 뮤린 RILF 모델에서 폐 섬유화증을 안전하게 예방한다는 결론을 지지한다.
참고문헌
실시예 16: 폐 섬유화증의 블레오마이신 모델에서 항-α
ν
β
6
인테그린 단일클론 항체의 효능
요약
폐 섬유화증에 대한 현행 치료는 크게 효과적이지 않아서 새로운 치료법을 개발한 필요성이 충분하다. TGF-β 경로를 차단하는 물질은 섬유아세포 활성화 및 증식, 및 세포외 매트릭스 분자의 발현을 포함하는, 폐 섬유화증의 특징적인 많은 병리학적 과정을 작동시키는 데 있어서 TGF-β의 중추적인 역할 때문에 특히 흥미롭다. TGF-β는 그의 전구-섬유화증성 활성 이외에 중요한 소염 사이토카인이므로, TGF-β의 치료적 억제는 과도한 염증을 촉진하지 않으면서 섬유화증을 이상적으로 차단해야 한다. 종래 연구는 인테그린 ανβ6이 생체 내, 특히 폐에서 TGF-β 활성화의 핵심 매개제라는 것을 입증하였다. ανβ6은 세포외 공간에서 잠복 TGF-β 복합체에 직접 결합하고, 이 결합은 많은 경우에서 활성 형태의 유리(liberation)에 필요하다. ανβ6이 결핍된 마우스는 폐에서의 손상된 TGF-β 신호전달로 인해 경미한 폐 염증을 가지며 블레오마이신에 의해 유도된 폐 섬유화증에 대한 내성을 가진다. 본 발명자들은 ανβ6과 잠복 TGF-β의 결합을 차단하고 TGF-β 활성화 및 후속 신호전달을 억제하는 단일클론 항체를 개발하였다. 여기서, 본 발명자들은 이 항체가 마우스에서 블레오마이신에 의해 유도된 섬유화증을 약화시키는 데 효과적임을 입증한다. 또한, 본 발명자들은 폐 콜라겐 발현을 약화시키는 데 있어서 거의 최대 효능이 기관지폐포 세척액 중의 염증 세포의 수에 의해 측정된 때 추가 염증을 나타내지 않는 투여량에서 달성된다는 것을 보여준다. 섬유화증을 약화시키는 보다 높은 투여량의 항체는 ανβ6-결핍 마우스에서 관찰된 것과 일치하는 염증을 유도할 수 있다. 이 발견은 ανβ6-매개 TGF-β의 차단의 전구-염증성 효과와 항-섬유화증성 효과는 이 모델에서 분리될 수 있고 섬유화증의 억제는 전구-염증 효과보다 더 낮은 투여량에서 일어난다는 것을 입증한다.
도입
TGF-β1 사이토카인은 병든 조직이 과량의 세포외 매트릭스(ECM)로 치환되어 궁극적으로 기관 흉터형성 및 부전을 초래하는 것에 의해 표시되는 병리학적 과정인 섬유화증의 개시 및 유지 둘다에서 중추적이다. TGF-β1은 ECM을 분비하고 섬유화증 과정의 진행을 유지하는 데 책임이 있는 섬유아세포의 증식 및 근섬유아세포의 활성화를 촉진한다[1-6]. TGF-β1은 상처 치유 동안에 일어나는 조직 리모델링 과정에서 잘 조절된 역할을 수행하지만, 많은 질환에서 이러한 조직 리모델링 과정은 비정상적이고 연장된 상향조절된 TGF-β 신호전달, 과도한 섬유아세포 축적, ECM 침착 및 흉터형성을 특징으로 한다. 생체 내 섬유화증의 진행에 있어서 TGF-β1의 중요성은 기능 획득(gain-of-function) 연구뿐만 아니라 차단을 통해서도 밝혀졌다[1, 7-12]. 폐에서의 다양한 사이토카인의 아데노바이러스 형질전환 과다발현은 TGF-β1이 상당한 염증의 부재 하에 섬유화증을 촉진하는 그의 능력 면에서 독특하다는 것을 보여준다. 섬유화증을 촉진하는 다른 사이토카인은 조직에서 TGF-β1 발현을 상향조절함으로써 종종 섬유화증을 촉진한다. 또한, 연구는 TGF-β 신호전달의 매개자인 SMAD3이 결핍된 넉아웃 마우스가 폐 섬유화증의 발병에 대한 내성을 가짐을 보여준다[13]. 항-TGF-β 물질을 사용한 다수의 연구는 질환 모델에서 섬유화증으로부터 충분한 보호를 보여준다[8, 9, 11, 14-17]. 결과적으로, TGF-β1은 섬유화증의 병리학적 상태와 관련된 질환의 치료에 대한 잠재적인 치료적 표적으로서 확인되었다.
ανβ6 인테그린은 TGF-β1 활성화의 중추적 조절자로서 확인되었다. TGF-β1은 성숙 활성 C-말단 TGF-β 사이토카인과 비공유적으로 연결되어 있는 N-말단 LAP와 함께 분비되며 절단되는 잠복 단백질로서 합성된다. 잠복 TGF-β1 복합체는 그의 동족 수용체에 결합할 수 없으므로, 프로테아제에 의한 절단, 낮은 pH 또는 이온화 방사선조사에의 노출, 및 상기 잠복 복합체가 그의 동족 수용체에 결합할 수 있게 하는 상기 잠복 복합체의 구조적 변화를 포함하는 여러 대안적 기작들 중 하나에 의해 활성 형태로 전환될 때까지 생물학적 불활성 상태로 남아있다[18-21]. ανβ6 인테그린은 잠복 TGF-β1 복합체 내의 RGD 모티프에 결합하여 이것을 활성 형태로 전환시킨다[18, 22-25]. TGF-β 활성화에 대한 여러 다른 기작들이 확인되었지만, 베타6 인테그린-결핍 마우스(β6 널 마우스)에서의 연구는 TGF-β의 ανβ6-매개 활성화가 폐 및 신장에서 섬유화증의 발병에 필요하다는 것을 암시한다[18, 26]. ανβ6은 정상 성체 조직에서 낮은 수준 또는 검출불가능한 수준으로 발현되지만, 염증/섬유화증성 질환에서 강하게 상향조절되며, 일반적으로 상피 세포에 한정되어 있다[27-30]. 따라서, 조직 손상 동안에 상피 세포에서의 ανβ6의 상향조절된 발현은 TGF-β의 증가된 국소적 활성화 및 방관자 세포에서의 후속 TGF-β-의존적 과정에 대한 기작을 제공한다. ανβ6 리간드 결합의 차단은 ανβ6의 발현이 상향조절되어 있는 조직에서 특이적으로 TGF-β 활성화를 국소적으로 억제하는 방법을 제공한다[31]. 이 방법은 TGF-β 경로의 전체적 억제와 관련된 임상적 안전성 위험을 감소시키는 잠재력을 제공한다.
본 명세서에 기재된 연구에서, 본 발명자들은 ανβ6이 특발성 폐 섬유화증(IPF)을 비롯한, 염증성 및 섬유화증성 병리학적 상태와 관련된 인간 폐 질환에서 유의하게 상향조절되어 있다는 것을 보여준다. 종래 연구는 ανβ6 기능이 결핍된 β6 널 마우스가 블레오마이신에 의해 유도된 폐 섬유화증으로부터 보호된다는 것을 입증하였다[18]. 본원에서, 본 발명자들은 ανβ6의 리간드 결합 및 TGF-β 활성화 기능을 차단하는 단일클론 항체가 섬유화증의 많은 다양한 척도에 의한 확인 시 다양한 마우스 종에서 블레오마이신-유도 섬유화증을 강력하게 억제한다는 것을 보여준다. 또한, 본 발명자들은 폐포 세포 집단이 블레오마이신에 의해 손상된 폐에서 낮은 유효 투여량에서 변경되지 않으므로 예측된 바와 같이 항-섬유화증성 작용 기작이 염증의 억제에 의해 매개되지 않는다는 것을 입증한다. 높은 투여량을 빈번히 투여한 경우에만 본 모델에서 추가 염증이 관찰되었고, 이는 β6 널 마우스에서의 추가 염증의 발견과 일치한다.
재료 및 방법
1. 시약 . ανβ6 mAb는 본 명세서의 다른 곳에 기재되고 종래 보고된 바와 같이 생성하였다[31]. 인간/마우스 키메라 2A1 및 3G9 cDNA는 제1 가닥 cDNA 합성 키트(Amersham/Pharmacia, Piscataway, NJ)를 사용하여 중쇄에 대한 불변 영역 프라이머 CDL-739 및 경쇄에 대한 불변 영역 프라이머 CDL-738로 각각의 모 하이브리도마 총 RNA로부터 생성하였다. 중쇄 및 경쇄 가변 영역 유전자는 cDNA 합성에 사용되는 동일한 3' 프라이머, 대부분의 뮤린 항체 유전자 신호 서열(요청 시 이용가능한 서열)에 특이적인 축퇴 프라이머의 풀 및 Pfu DNA 중합효소(Stratagene, La Jolla CA)를 사용하여 중합효소 연쇄 반응으로 증폭시켰다. 클로닝된 중쇄 및 경쇄 가변 영역들을 인간 IgG1 불변 영역과 함께 포유동물 발현 벡터 내로 연결시켰다. 재조합 가용성 뮤린 TGF-β 수용체 타입 II-Ig 융합 단백질(rsTGF-βRII-Ig)은 종래 보고된 바와 같이 생성하였다[32]. 연구-등급 mu3G9, 1E6 및 rsTGF-βRII-Ig(포스페이트 완충 식염수 중의 정제된 단백질)를 모든 실험에서 사용하였다.
2. 동물 .
마우스. SV129 마우스는 판독대상으로서 폐 히드록시프롤린을 사용한 실험에 사용하였다(Sheppard Laboratory, UCSF). C57B16 마우스는 판독방법으로서 조직형태계측 또는 BAL 수거 및 분석을 이용한 실험에 사용하였다(Biogen Idec). 판독대상으로서의 콜라겐 유전자 발현의 정량적 분석을 위해, 형질전환 레포터 마우스를 사용하였다(Biogen Idec). colIα2 유전자 프로모터의 17 kb 영역의 조절 하에 루시퍼라제 레포터 유전자를 보유하는 형질전환 마우스는 이미 공지되어 있다[33]. 이 마우스는 형질전환 수컷을 C57B1/6 X DBA/2 F1 하이브리드 암컷(Jackson Laboratories)과 교배시킴으로써 유지한다. (꼬리 루시퍼라제 발현에 의한 평가 시) 형질전환유전자(transgene)에 대해 양성을 나타내는 자손을 하기 요약한 바와 같은 블레오마이신 챌린지(challenge) 실험을 위해 선별하였다.
햄스터(Giri Laboratory). 중량이 90 내지 110 g인 수컷 골든 시리안(Golden Syrian) 햄스터를 시몬센스 인코포레이티드(Simonsens, Inc.)(Gilroy, CA)로부터 구입하였다. 햄스터를 공기가 여과되어 있으면서 온도와 습도가 일정하게 유지된 시설 내에서 4개의 군으로 나누어 사육하였다. 모든 보호는 동물 복지 약정에 대한 국립 보건소의 지침(National Institutes of Health Guide for Animal Welfare Act)에 따라 행하였다. 햄스터를 임의의 처리 전에 1주일 동안 시설 내에서 적응시켰다. 12시간의 명/암 사이클을 유지하였다.
3. 면역조직화학 . 마우스 폐를 10% 완충 포르말린 중에서 수거하여 통상의 관행에 따라 파라핀 조직학적 분석을 위해 처리하였다. 섬유화증성 병리학적 상태를 가진 폐 질환을 앓는 환자의 폐로부터 얻은 파라핀 조직 절편을 지. 데이비스(G. Davis)(U. Vermont), 알. 라피아티스(R. Lafyatis)(Boston University), 아르다이스 코포레이션(Lexington, MA) 및 아스테란드 인코포레이티드(Asterand Inc.)(Detroit, MI)로부터 입수하였다. 조직 절편으로부터 파라핀을 크실렌 및 에탄올 중에서 제거하고, 조직 절편을 증류수 중에서 재수화한 후, 0.45% H2O를 함유하는 메탄올 중에 함침시켰다. 조직을 펩신(00-3009, Zymed, San Francisco, CA)과 함께 항온처리하고 아비딘 및 바이오틴으로 차단하였다(SP-2001; Vector Laboratories, Burlingame, CA). 1차 항체를 0.1% BSA가 함유된 PBS 중에서 희석하고, 조직을 4℃에서 밤새 항온처리하였다. 절편을 항-ανβ6 mAb의 인간/마우스 키메라 형태인 2A1[31], 및 마우스 조직에 대한 항-인간-바이오티닐화 2차 항체(PK-6103, Vector Laboratories, Burlingame, CA)와 함께 항온처리하였다.
절편을 뮤린 2A1[31], 및 인간 조직에 대한 항-마우스-바이오티닐화 2차 항체(PK-6102, Vector Laboratories)와 함께 항온처리하였다. 아비딘-바이오틴 복합체-호스라디쉬 퍼록시다제(Vector Kit, PK-6102)를 절편에 가하고, 실온에서 30분 동안 항온처리하고, 3,3'-디아미노벤지딘(DAB) 기질을 지시된 바와 같이 준비하고(SK-4100, Vector Laboratories), 실온에서 5분 동안 절편에 가하였다. 조직 절편을 메이어스 헤마톡실린으로 1분 동안 염색하고 물 및 PBS 중에서 린싱하였다. 모든 인간 조직 샘플을 지역 위원회 심사의 승인 및 환자 승인 하에 입수하였다.
3.1 마우스에서의 블레오마이신 주입.
SV129 종(D. Sheppard, UCSF). 종래 보고된 바와 같이[18], 블레오마이신 또는 식염수를 마우스의 기관 내로 주입하였다. 요약하면, 연령 및 성별이 일치하는 8주령 내지 12주령의 129/terSVEMS 마우스 종을 특정 병원체-무함유 환경 하에서 유지하였다. 블레오마이신(Mead Johnson, Princeton, NJ)을 멸균 식염수(60 ㎖ 중의 0.03 또는 0.05 유니트) 중에 용해시켰다. 블레오마이신 또는 식염수를 메톡시플루란 마취 하에 직접적인 절개에 의해 경기관(transtracheal)으로 투여하였다.
C57B1/6 종 및 콜라겐 레포터 마우스((Biogen Idec). 100 mg/kg의 케타민 및 10 mg/kg의 크실라진을 복강내로 주사함으로써 마우스를 마취시켰다. 멸균 #15 스칼펠(scalpel)을 사용하여 목에 0.5 내지 1.0 cm의 중앙 절개를 행하여 가시화를 위한 기관을 노출시켰다. 기관을 노출시키고 분무 팁을 구강을 통해 기관 내에 위치시킨 후, 펜 센츄리 마이크로분무 장치(Penn Century microspraying device)로 블레오마이신을 주입하였다. 식염수를 대조군 동물 내에 주입하였다. 주입 후, 멸균 상처치유용 클립으로 수술 부위를 폐쇄시켰다. 수술 후 통증 때문에, 0.05 mg/kg의 부프레노르핀을 피하 주사하였다.
다수의 치료 프로토콜을 활용하여 이 마우스 블레오마이신 연구에서 시험 제품을 평가하였으므로 이는 결과 단락에 기재될 것이다.
3.2 햄스터에서의 블레오마이신 주입. 펜토바르비탈 마취 하에, 0일, 7일 및 14일째 날에 식염수(SA; 4 ㎖/kg) 또는 블레오마이신(BL; 6.5 U/4 ㎖/kg)을 햄스터에 IT 주입하였다. 동물을 하기 6개의 실험군으로 무작위적으로 나누었다: PBS로 처리된 SA-주입 군(SA + PBS); PBS로 처리된 BL-주입 군(BL + PBS); 0일째 날에 시작한 mu3G9로 처리된 BL-주입 군(BL + Ab1); 7일째 날에 시작한 mu3G9로 처리된 BL-주입 군(BL + Ab2); 14일째 날에 시작한 mu3G9로 처리된 BL-주입 군(BL + Ab3); 및 0일째 날에 시작한 1E6으로 처리된 BL-주입 군(BL + 1E6). 동물들을 28일째 날에 희생시키고 이들의 폐를 분리하여 히드록시프롤린 및 지질 과산화 분석을 위해 처리하였다.
4. 콜라겐 함량에 대한 히드록시프롤린 측정 . 폐를 유리 튜브 내의 1 ㎖ dH2O 중에서 균질화시켰다(Fisher #14961). 125 ㎕의 50% 트리클로로아세트산(TCA)을 균질화물에 첨가하고 얼음 위에서 20분 동안 항온처리하였다. 샘플을 4℃에서 1000 rpm으로 5분 동안 원심분리하였다. 상청액을 따라 버리고 1 ㎖의 12 N HCl을 유리 튜브 내의 펠릿에 첨가하였다. 그 다음, 샘플을 (유리 비이커 내에서) 110℃에서 24시간 동안 베이킹하였다. 6개의 히드록시프롤린 표준물(Sigma-H6002)을 0.25 mg/㎖부터 만들었다. 건조된 펠릿을 2 ㎖의 dH2O로 재구성하였다. 500 ㎕ 클로르아민 T(0.5 M 아세트산나트륨 및 10% 이소프로판올 중의 1.4% 클로르아민 T)를 함유하는 1.5 ㎖의 에펜도르프 튜브에 200 ㎕의 샘플을 첨가하고 실온에서 20분 동안 항온처리하였다. 그 후, 500 ㎕의 에를리히/pDMBA(70% 이소프로판올 및 30% 과염소산 중의 1 M p-DMBA(p-디메틸아미노벤즈알데히드))를 첨가하고 65℃에서 15분 동안 항온처리하였다. 100 ㎕의 최종 반응 용액을 96-웰 플레이트로 옮기고, 각 샘플에 대해 3회 반복 측정을 수행하고, 2시간 후에 샘플을 550 nm에서 판독하였다.
5. 조직학 지수 . 판독방법으로서 조직형태계측을 이용한 실험을 위해, 희생되었을 때 각 마우스의 전체 폐를 조직학적으로 평가하였다. 횡단 절편을 잘라 내어 표준 관행에 따라 매손 트리크롬으로 염색하였다. 횡단 절편을 각 마우스로부터 얻은 폐의 다수의 엽을 포함하도록 선택하였다. 전체 트리크롬-염색 절편(마우스 당 약 30의 평균)을 덮도록 고-배율(100X) 필드에서 사진을 촬영하였다. 각 사진을 메타모르프 6.0.5 소프트웨어로 (트리크롬-염색 조직학적 절편에서 청색으로 나타내는) 콜라겐 함량에 대해 평가하였다. 청색 영역은 색상 역치(threshold)에 의해 선별되었고 총 조직 면적의 비율로서 표시되었다.
6. 루시퍼라제 분석 . 폐를 수거하고 1 ㎖의 용해 완충제 중에서 균질화하였다(0.1M KH2PO4-pH7.8 및 1 mM DL-디티오트레이톨). 이어서, 샘플을 얼음 상에서 10분 동안 놓아 두고, 4℃에서 12,000 rpm으로 10분 동안 원심분리한 후, 각각의 샘플 100 ㎕를 웰락 이소플래터(Wallac Isoplatter)로 옮겼다. 샘플을 얼음 상에서 다시 15분 동안 놓아둔 후, 100 ㎕의 루클라이트(Luclite) 기질(Perkin Elmer #601911)을 첨가하였다. 그 다음, 루시퍼라제 활성을 발광측정기 상에서 판독하였다.
7. 기관지폐포(BAL) 수거 및 BAL 세포의 차등 염색 . 과량의 이낵틴(Inactin)(Sigma) 복강내 주사(IP)를 이용하여 마우스를 마취시켰다. 가장 무딘 해부(blunt dissection)를 이용하여 기관을 노출시켰다. 그 후, 기관을 두 개의 연골 고리 사이에서 가위로 개방하고, 23-게이지의 무딘 말단 바늘을 기관 내로 삽입하였다. 슈와르츠 임시 클립(Roboz)으로 약하게 클램핑함으로써 바늘을 제자리에 고정시켰다. Ca2+ 또는 Mg2+를 함유하지 않는 포스페이트 완충 식염수(PBS) 0.8 ㎖를 폐 내로 주사함으로써 BAL을 수행하였다. 이어서, 높은 압력을 인가하지 않으면서 유체를 시린지 내로 빨아들이고, 얼음 상의 15 ㎖ 폴리프로필렌 팔콘 튜브 내로 옮겼다. 그 후, 상기 절차를 반복하고, 과도한 음의 압력을 발휘하지 않으면서 BAL 액을 시린지 내로 다시 빨아들였다. 샘플을 얼음 상에서 저장하고 세포 계수 및 분화를 위해 처리하고, BAL 펠릿 및 유체 둘다를 수거하고 -80℃에서 저장하였다.
1. 그 다음, BAL을 4℃에서 탁상용 원심분리기 상에서 180 g 및 1000 rpm(Beckman GPR)으로 10분 동안 원심분리하였다. 그 후, 상청액(BAL 액)을 분리하여 -80℃에서 동결시켰다. 1.0 ㎖의 RBC 용해 용액(Sigma)을 펠릿에 첨가하고 20초 동안 볼텍싱하였다. 이어서, 세포 계수 및 사이토스핀을 행하였다.
2. 샘플을 얼음 상에서 저장하고, 세포를 혈구계에 가한 후 판독하기 전에 세포가 1분 동안 가라앉게 함으로써 혈구계를 이용하여 세포의 수를 세었다.
3. 100 ㎕의 세포 현탁액을 실온에서 500 RPM으로 5분 동안 사이토스피닝하였다(ThermoShandon). 사이토스핀 제제는 세포 용액을 사이토스핀 장치 내에 넣고 500 rpm에서 5분 동안 원심분리함으로써 준비하였다. 세포 농도가 슬라이드 상에서 너무 높지 않도록 점검하였다. 슬라이드를 밤새 건조한 후, 디프퀵(Fisher)으로 염색하였다. 제조자(DiffQuik)의 프로토콜에 따라 염색을 수행하였다. 일단 슬라이드가 건조되고 퍼마운트(Fisher)로 커버 슬립을 덮으면, 세포를 유형별로 분류하고 실험실 세포 계수기(Fisher)를 이용하여 100개의 무작위로 선택된 세포의 수를 세었다.
8. 통계적 분석 . 편차 분석(ANOVA)을 이용하여 비히클 대조군 및/또는 이소타입 대조군과 시험 제품 사이에 통계적 비교를 수행하였다. 통계적으로 유의한 차이가 ANOVA를 이용하여 p<0.05의 확률에서 확립되면, 군 사이의 유의한 차이를 던넷(Dunnet) 다중 비교 검정으로 평가하였다.
결과
1. 인간 폐 질환 및 블레오마이신 모델에서의 α ν β 6 의 발현 . 상향조절된 TGF-β 발현 및 신호전달은 섬유화증성 또는 염증성 병리학적 상태를 수반하는 다양한 인간 폐 질환에서 보고되어 있다[20, 34]. 그러나, ανβ6 발현은 섬유화증성-염증성 폐 질환의 작은 샘플에서만 보고되어 있다[28]. 본 발명자들은 섬유화증성 및/또는 염증성 변화를 가지는 것을 특징으로 하는 폐 질환을 앓는 환자로부터 얻은 41개의 폐 조직 샘플에서 ανβ6의 발현을 평가하였다(표 16-1). 추가로, 본 발명자들은 암 환자의 폐 생검의 외관상 정상적인 영역으로부터 얻은 폐 조직 어레이를 염색하였다(Imgenex). 정상 폐에서의 ανβ6의 발현은 면역조직화학적 분석에 의해 거의 검출될 수 없었다. "정상" 폐 조직 어레이 상의 일부 조직 절편은 양성 ανβ6 염색을 보였지만, 이들 절편들 각각은 거의 염증성 병리학적 상태도 보였다. 모든 41개의 병든 폐 샘플에서, 섬유화증 및/또는 염증성 변화를 가진 폐의 영역은 ανβ6 발현의 강한 상향조절을 보였다(도 58). ανβ6은 명백한 섬유화증 영역 상에 있는 상피 세포, 또는 염증성 침입물에 인접한 영역에 편재되어 있었다. 상향조절된 ανβ6의 존재는 특발성 폐 섬유화증, 피부경화증 폐 질환 및 만성 폐색성 폐 질환을 비롯한 광범위한 섬유화증성-염증성 질환에 걸쳐 관찰되었다.
발현과 특정한 병리학적 변화의 상관관계를 더 잘 파악하기 위해, 본 발명자들은 염색된 조직 샘플을 외부의 훈련된 폐 병리학자에게 보냈다. 그가 상업적으로 입수한(Ardais and Asterand) 많은 샘플의 병리학적 진단을 검증할 수 없을지라도, 그는 보다 약한 섬유화증 및 보다 우수한 예후와 관련된 병리학적 상태인 비-특이적 간질성 폐렴(NSIP)과 비교할 때 섬유화증 및 진행성 질환과 관련된 병리학적 상태인 "ανβ6 염색의 더 높은 강도가 일반적인 간질성 폐렴에서 관찰되었다"는 것을 일관되게 확인하였다. UIP를 앓는 환자로부터 얻은 생검 모두에서, 강한 염색이 폐포 관과 폐포의 경계를 형성하는 폐세포 타입 II 및 타입 I 내에서 관찰되었지만, 큰 기도는 주로 음성이었고, 폐포 내 대식세포는 음성이었다. 요약하면, "높은 수준의 염색은 섬유화증 영역 및 UIP와 관련되었고", 보다 덜 강한 염색의 유사한 패턴은 NSIP를 비롯한 섬유화증의 다른 경우에서 관찰되었다. UIP가 특발성 폐 섬유화증(IPF)을 앓는 환자에서 우세한 병리학적 상태이기 때문에, 이 병리학적 상태를 가진 환자에서 전구-섬유화증 사이토카인 TGF-β의 활성화제인 ανβ6의 강한 과다발현은 ανβ6이 섬유화증의 진행을 유발하는 데 있어서 기능적인 역할을 수행할 수 있다는 것을 암시한다.
2. 마우스 블레오마이신 모델에서의 α ν β 6 의 발현 . 더불어, ανβ6이 폐 섬유화증의 블레오마이신 마우스 모델에서 상향조절되었다는 것을 검증하기 위해, 본 발명자들은 블레오마이신 주입 후 1일, 5일 및 15일째 날에 채취한 조직 절편 상의 ανβ6 단백질의 존재에 대해 면역염색하였다. 5일째 날, ανβ6 발현은 블레오마이신 챌린지에 의해 손상된 폐의 영역 전체에 걸쳐 폐포 상피 상에서 상향조절되어 있다(도 59). 15일째 날, 섬유화증이 현저한 영역이 분명해졌을 때, ανβ6은 이 섬유화증 영역들 내의 폐포 상피에서 보다 강하게 상향조절되어 있다.
3. SV129 마우스에서의 히드록시프롤린에 의한 섬유화증의 평가 . ανβ6이 유전적으로 결핍된 마우스(β6 널 마우스)는 SV129 마우스 종에서 블레오마이신-유도 폐 섬유화증으로부터의 보호를 이미 나타내었다[18]. 본 발명자들은 이와 동일한 종에서 블레오마이신-유도 섬유화증을 약화시키는 데 있어서 항-ανβ6 단일클론 항체 mu3G9의 효능을 평가하고자 하였다. 일련의 4가지 실험을 본 발명자들의 동료인 UCSF의 딘 쉐파드 박사의 실험실에서 수행하였다(표 16-2). 0일째 날, 블레오마이신을 SV129 마우스의 기관 내로 주입하고, 블레오마이신 주입 후 0일, 15일 또는 30일째 날부터 주당 3회씩 4 mg/kg의 mu3G9를 복강내로 주사하였다. 대조군 마우스를 PBS 또는 음성 대조군 항체 1E6으로 주사하였다. 1E6은 인간 LFA-3에 대한 뮤린 IgG1 항체이며, 임의의 마우스 항원에 결합하지 않는다. 마우스를 30일째 또는 60일째 날에 희생시키고, 섬유화증을 총 조직 콜라겐의 척도인 히드록시프롤린 함량으로 평가하였다. 4개의 실험들 중 3개 실험에서, mu3G9-처리 마우스에서 히드록시프롤린의 통계적으로 유의한 감소가 있었고, 이는 블레오마이신-유도 섬유화증을 약화시키는 데 있어서의 효능을 나타낸다(도 60). 블레오마이신 주입 후 30일까지 처리를 지연하였을 경우에만(도 60C), mu3G9-처리 마우스의 폐에서의 히드록시프롤린 함량이 PBS-처리 또는 이소타입-대조군 처리 마우스에 비해 유의하게 감소되지 않았다.
4. 46일 동안 처리된 블레오마이신-챌린지 마우스에서의 생존율의 평가 . 블레오마이신 모델에서의 항-섬유화증 효능은 개선된 생존율과 대체적으로 상관관계를 가지지 않으므로, 본 발명자들은 mu3G9-처리 마우스에서 생존율이 개선되리라고 예측하지 못하였다. 그러나, 15일 내지 60일 동안 주당 3회씩 4 mg/kg의 mu3G9로 처리된 마우스(도 60D)가 본 발명자들이 시험한 가장 긴 처리 기간(46일)을 대표하기 때문에, 본 발명자들은 그 실험에서 시험된 군들에서 생존율의 임의의 차이가 있는지를 알아보기 위해 분석하였다. mu3G9로 처리된 마우스를 PBS로 처리된 마우스 및 4B4 비-차단 대조군 항체로 처리된 마우스와 비교하였을 때, 총 생존율에서의 유의한 차이는 없었다(표 16-2).
5. C57B16 마우스에서의 섬유화증의 조직형태계측적 분석 . 상이한 마우스 종 및 상이한 실험실에서 mu3G9의 항-섬유화증 효능을 검증하기 위해, 바이오겐 아이덱에서의 일련의 실험에서 C57B16 마우스 종을 사용하여 mu3G9를 평가하였다. 이 종은 블레오마이신 모델에서 종종 사용되고 있으며 주입 후 14일째 날에 측정될 수 있는 급성 섬유화증을 나타낸다. 처음 3개의 실험에서, 0일째 날에 마우스의 기관 내로 블레오마이신을 주입하고, 마우스를 블레오마이신 챌린지 1일 전(-1일)부터 주당 3회씩 mu3G9로 처리하였고, 폐 수거를 위해 14일째 날에 마취시켰다. 4번째 실험에서, 14일째 날까지 처리를 지연시키고, 폐를 28일째 날에 수거하였다. 이 실험들 각각에서(도 61), 매손 트리크롬-염색 조직 절편에서 청색 염색 영역으로서 조직형태계측적으로 측정될 때, mu3G9는 PBS-처리 대조군에 비해 섬유화증 폐 조직의 비율을 일관되게 감소시켰다. 1E6 mAb가 음성 IgG 대조군으로서 사용된 2개의 실험들 중 하나에서, 1E6 mAb도 섬유화증성 조직의 비율을 유의하게 감소시켰다(도 61A). 1E6 mAb의 이 효과는 판독대상으로서 히드록시프롤린을 사용한 초기 실험들에서 관찰되지 않았고(도 60), 지연된 처리(14일 내지 28일) 실험에서도 관찰되지 않았다(도 61D). 요약하면, 다수의 실험들이 C57B16 마우스에서 블레오마이신에 의해 유도된 섬유화증성 조직의 비율을 감소시키는 데 있어서 mu3G9 mAb의 효능을 입증하였다. 그러나, 조직형태계측 판독의 노동-집약적 성질 때문에, 본 발명자들은 섬유화증을 측정하는 보다 신속한 정량적 방법을 찾고자 하였다.
6. 정량적 판독대상으로서의 콜라겐-루시퍼라제 레포터 형질전환유전자의 용 도. 루시퍼라제 레포터 유전자가 콜라겐 Iα2 프로모터의 조절 하에서 발현되는 형질전환유전자를 보유하는 형질전환마우스는 섬유화증 모델에서 콜라겐 발현의 정량적 판독정보를 제공하기 위해 이미 사용되고 있다[33];[35]. 14일째 날, 식염수 대조군에 비해 블레오마이신-챌린지 마우스에서의 폐 루시퍼라제 수준의 증가는 대략 10배이었기 때문에, 이는 폐 루시퍼라제 수준은 히드록시프롤린 측정보다 훨씬 더 민감한 판독대상이 된다. 이 시스템을 사용하여, 본 발명자들은 주당 1회 투여를 이용하여 3G9 항체의 투여량-적정을 수행하되, 히드록시프롤린 및 트리크롬 조직형태계측을 판독방법으로 이용한 실험에서 사용된 주당 3회씩 4 mg/kg의 투여 섭생법으로 처리된 마우스 군을 포함시켰다. 투여량 적정은 각 실험에서 PBS-처리 대조군(총 n=17에 대해 n=6, 5 및 6)이 사용된 3개의 실험에서 평가되었다. 루시퍼라제 측정치에서의 실험-대-실험 편차를 보정하기 위해, 각 실험에서 모든 군에 대한 루시퍼라제 값을 PBS 대조군의 평균으로 표준화하였다. 블레오마이신-챌린지 레포터 마우스의 mu3G9 처리는 콜라겐 루시퍼라제 레포터에서 투여량-의존적 감소를 나타내었으며(도 62), 이때 유의한 효능이 주당 0.3 mg/kg의 투여량에서 관찰되었고, 최대 효능이 1 내지 3 mg/kg에서 관찰되었다.
7. 기관지폐포 세척 세포 조성물의 분석 . 본 발명자들은 섬유화증의 억제가 감소된 염증으로 인한 것이지 아니면 폐 내의 염증 세포의 주요 하위집단에서의 변화로 인한 것인지를 확인하기 위해 2일, 5일, 8일 및 11일째 날에 블레오마이신 모델의 기관지폐포 세척(BAL) 세포 집단을 분석하였다. 예측된 바와 같이, 식염수-주입 마우스와 비교할 때 블레오마이신의 기관내 투여로 인해 BAL 세포의 총 수에서의 증가가 있었다. 그러나, 기간 전체에 걸쳐, 이소타입 대조군 항체 1E6으로 처리된 마우스와 비교할 때 0.3, 1.0 및 3 mg/kg의 유효 투여량의 3G9로 처리된 마우스에서 관찰된 BAL 세포의 총 수 뿐만 아니라 대식세포, 호중구 또는 림프구의 수 또는 비율에서의 유의한 차이는 없었다. 이어서, 본 발명자들은 히드록시프롤린 및 조직형태계측 판독을 이용하여 효능을 입증하기 위해 먼저 사용된 주당 3회 처리를 이용하여 훨씬 더 높은 투여량을 시험하였다. 마우스는 블레오마이신 챌린지를 기준으로 -1일, +1일 및 +3일째 날에 3가지 투여량, 즉 4, 20 및 40 mg/kg의 m3G9를 제공받았다. 마우스를 5일째 날에 마취시키고, BAL 세포 함량을 분석하였다. 4 mg/kg의 투여량(총 12 mg/kg의 투여량)에서, BAL 세포의 총 수 뿐만 아니라 대식세포, 호중구 또는 림프구의 수 또는 비율에서의 유의한 변화는 없었다. 20 및 40 mg/kg의 투여량(총 60 및 120 mg/kg)에서, IgG1 대조군이 아닌 PBS 대조군에 비해 대식세포의 수의 유의한 증가가 있었다(도 62). 40 mg/kg의 투여량에서, PBS 및 IgG 대조군과 비교할 때 BAL에서 호중구의 비율의 유의한 증가가 있었지만, 호중구의 총 수의 변화는 유의한 수준까지 도달하지 못하였다. 매우 높은 투여량(총 120 mg/kg)에서 일어나는 호중구의 이러한 증가는 방사선조사 섬유화증 모델에서 높은 투여량의 장기 처리(6 및 10 mg/kg 처리, 3-4개월)에서 관찰된 것과 유사하였다(실시예 15). 요약하면, 항-ανβ6 항체는 0.3 mg/kg만큼 낮은 주당 투여량에서 블레오마이신-유도 콜라겐 발현을 약화시킬 수 있지만, 총 12 mg/kg 이하의 투여량은 이 모델에서 BAL 세포 집단에서 유의한 변화를 나타내지 않는다. 보다 높은 투여량은 총 대식세포 수 및 호중구의 비율을 증가시킬 수 있다.
8. 다중 블레오마이신 투여 햄스터 모델에서의 평가 . 본 발명자들은 0일, 7일 및 14일째 날에 블레오마이신을 기관 내로 3회 연속적으로 투여한 햄스터 폐 섬유화증 모델에서 mu3G9의 효능을 조사하였다. 3개의 햄스터 군들을 0일, 7일 또는 14일째 날부터 주당 3회씩 5 mg/kg의 mu3G9(주당 총 15 mg/kg)로 처리하였다. 28일째 날, 동물을 희생시키고 히드록시프롤린(도 65A) 및 지질 과산화 분석(도 65B)으로 섬유화증에 대해 평가하였다(캘리포니아-데이비스 대학(U. California-Davis)(Davis, CA)에서 수행됨). 예상외로, 이 모델에서 섬유화증을 약화시키는 데 있어서의 효능이 관찰되지 않았다. 이 군들 중 하나에서, 햄스터는 7일 및 14일째 날부터 mu3G9로 처리되었고, 마우스는 PBS 및 IgG 대조군 둘다에 비해 폐 히드록시프롤린에서의 통계적으로 유의한 증가를 보였다. 지질 과산화 값은 IgG 대조군에 비해 유의하게 상승되지 않았다. 본 발명자들은 이 명백한 악화가 햄스터의 생존에 대해 영향을 주는지를 알아보기 위해 다양한 처리군들의 생존 곡선을 분석하였다. 0일째 날부터 처리된 mu3G9 마우스(도 65C에서 BL+AB1 군)는 PBS 및 IgG 대조군보다 다소 더 일찍 죽기 시작하였으나, B1+Ab1 생존 곡선을 PBS 및 IgG 대조군에 대하여 개별적으로 비교하였을 때, 차이는 유의하지 않았다(로그 순위 시험; PBS에 대해 p=0.15, 및 IgG 대조군 mAb에 대해 p=0.25). mu3G9 단일클론 항체가 햄스터와 교차-반응하는지는 알 수 없으며, 햄스터는 이 모델에서 사용된 뮤린 항체에 대한 항체 반응을 나타낼 수 있는 가능성이 있다. 이 모델에서 결과 해석의 어려움 때문에, 효능에 대한 다양한 투여량의 평가는 수행하지 않았는데, 이는 일치된 효능이 2개의 상이한 뮤린 폐 섬유화증 모델(블레오마이신-유도 폐 섬유화증 모델 및 방사선조사-유도 폐 섬유화증 모델)에서 관찰되었기 때문이다.
[표 16-1]
ανβ6 발현에 대해 면역염색된 인간 폐 질환 샘플
[표 16-2]
mu3G9로 46일 동안 처리된 블레오마이신-챌린지 마우스의 생존
결론
mu3G9는 블레오마이신 섬유화증 모델에서 질환 심각성의 강력한 억제제이다. 이 일련의 실험들은 하기 사항들을 입증한다:
(a) ανβ6 발현은 섬유화증성 및/또는 염증성 병리학적 상태를 가진 매우 광범위한 인간 폐 질환에서 상피 세포 상에서 상향조절된다. ανβ6 발현은 마우스 블레오마이신 섬유화증 모델에서 유사하게 상향조절된다.
(b) mu3G9는 분석에 대한 다수의 판독대상을 이용하여 다수의 실험에서 블레오마이신-유도 섬유화증을 유의하게 감소시켰다. 효능은 시험된 마우스의 모든 종, 즉 SV129, C57B16 및 C57B16 X DBA 하이브리드에서 관찰되었다. 콜라겐 발현을 억제하는 효능은 주당 0.3 mg/kg만큼 낮은 투여량에서 관찰되었다.
(c) mu3G9의 작용 기작은 소염성을 가진 전구-섬유화증성 사이토카인인 TGF-β의 억제를 통한 것이다. 예측된 바와 같이, 섬유화증 억제는 염증을 약화시키지 않으면서 일어난다. 5일째 날, 높은 투여량의 mu3G9의 빈번한 투여(이틀마다 20 및 40 mg/kg)는 블레오마이신-챌린지 마우스의 기관지폐포 세척액 중의 총 BAL 세포, 대식세포 및 호중구에서 PBS에 비해 약간의 증가를 유도할 수 있지만, IgG 대조군에 비해서는 그러한 증가를 유도할 수 없다.
(d) mu3G9는 다중 투여 블레오마이신 햄스터 모델에서 효과적이지 않았다. 하나의 mu3G9-처리 군에서의 섬유화증의 분명한 악화가 시험-제품과 관련되어 있는지는 불분명하다.
(e) 46일 동안 주당 3회씩 4 mg/kg의 투여량에서의 mu3G9 처리는 블레오마이신 모델에서 생존에 영향을 주지 않았다.
참고문헌
용어 정리
부록 A: mu3G9 투여의 변이
부록 B: mu3G9 투여의 변이
부록 C: mu3G9 투여의 변이
부록 D: 블레오마이신에서 BAL 조성: 주마다 투여
부록 E: BAL 세포 조성물: 주마다 3회 투여
실시예 17: 정상 및 병든 마우스 폐에서 항-α
ν
β
6
인테그린 단일클론 항체의 효과의 분자적 분석
요약
폐 섬유화증에 대한 동물 모델은 인간 특발성 폐 섬유화증의 섬유화증성 병리학적 상태의 일부 양태를 모델링함에 있어서 효과적이지만, IPF의 정확한 병리학적 상태를 정확히 모방하는 동물 모델은 없다. 또한, IPF에서의 질환 개시 및 진행 기작이 불완전하게 이해되어 있기 때문에, 다수의 질환 모델에서 잠재적 치료제를 시험하고 섬유화증성 병리학적 상태의 개선 면에서 뿐만 아니라 상기 인간 질환에서 관련되어 있는 것으로 생각되는 분자적 경로에서의 중재 면에서 상기 치료제의 잠재적 효능을 평가하는 것은 중요하다. TGF-β 경로는 IPF에서 핵심 경로로서 연루되어 있다. 인간 IPF 폐의 전체적인 전사체 프로파일링은 이 질환에서의 우성 징후가 섬유아세포 활성화 및 증식, 및 세포외 매트릭스 분자의 발현을 비롯한, 폐 섬유화증의 특징이 되는 많은 병리학적 과정을 작동시키는 데 있어서 TGF-β의 공지된 역할과 일치하는 활성화된 TGF-β 경로 중 하나라는 것을 보여준다. TGF-β의 전구-섬유화증성 활성 이외에, TGF-β는 중요한 소염성 사이토카인이므로, TGF-β의 치료적 억제는 과도한 염증을 촉진하지 않으면서 섬유화증을 이상적으로 차단해야 한다. 인테그린 ανβ6은 잠복 TGF-β 복합체에 직접 결합하고 TGF-β가 활성 상태로 전환하는 데 필요하다. 그러나, ανβ6-매개 TGF-β 활성화가 일부 조직에서만 중요하기 때문에, ανβ6 기능이 완전히 결핍된 마우스는 폐에서만 병리학적 상태를 보이는 반면, TGF-β가 결핍된 마우스는 다수의 기관 시스템에서 염증을 보인다. 그러므로, 여기에 요약된 치료 전략은 TGF-β 경로의 전체적인 억제를 피하도록 ανβ6 기능을 차단하는 것과, 증가된 TGF-β 신호전달과 관련된 섬유화증성 병리학적 상태를 차단하는 효능이 TGF-β의 완전한 상실과 관련된 염증을 유도하지 않는 투여량에서 달성될 수 있다는 것을 입증하는 것이다. 여기서, 본 발명자들은 섬유화증성 및 염증성 병리학적 상태와 관련된 mRNA 및 단백질 수준에서의 분자적 변화를 특징규명한다. 본 발명자들은 높은 투여량의 항-ανβ6 단일클론 항체 mu3G9, 즉 임상 후보 BG00011의 뮤린 모 항체가 ανβ6-결핍 마우스와 일치하는, 폐 내의 염증성 마커의 mRNA 및 단백질 변화를 나타낸다는 것을 입증한다. 또한, 본 발명자들은 섬유화증을 약화시키는 데 있어서 효과적인 낮은 투여량의 mu3G9가 마우스에서 이들 염증성 변화를 나타내지 않는다는 것을 입증한다.
도입
TGF-β1 사이토카인은 섬유아세포가 궁극적으로 기관 흉터형성 및 부전을 초래하는, 과도한 세포외 매트릭스(ECM)를 생성하도록 자극하는 것으로 알려져 있는 전구-섬유화증성 사이토카인이다(Roberts 1986). 폐에서 다양한 사이토카인의 아데노바이러스 및 형질전환 과다발현은 TGF-β1이 상당한 염증의 부재 하에 섬유화증을 촉진하는 그의 능력 면에서 독특하다는 것을 보여준다. TGF-β 경로에서의 유전자의 넉아웃 마우스 모델(Bonniaud 2004, Munger 1999) 및 항-TGF-β 물질을 사용한 다수의 연구는 섬유화증을 약화시키는 수단으로서 TGF-β의 억제 효능을 입증하였다(George, J. 1999; Sharma, K. 1996; Bonnidaud, P. 2004; Zheng, H. 2000; Kasuga, H. 2001; Ziyadeh, F. N. 2000; Laping, N. J. 2003). 2개의 서브유니트, 즉 αν 인테그린과 β6 인테그린으로 구성된 ανβ6 인테그린은 특히, 폐에서의 TGF-β1 활성화의 중추적인 조절자이다. ανβ6 인테그린은 손상, 염증 및 섬유화증 동안에 상피 세포 상에서 상향조절되며, 잠복 TGF-β1 결합체에 결합하여 이 결합체를 활성 형태로 전환시킨다[Huang 1998; Munger 1999; Busk 1992; Yokoaski 1996; Annes 2002]. 잠복 TGF-β1에 결합하는 ανβ6의 차단은 ανβ6의 발현이 상향조절된 부위에서 TGF-β 활성화를 국소적으로 억제하는 방법을 제공함으로써, 모든 조직에서 TGF-β 경로의 전체적인 억제의 잠재적인 임상적 안전성 위험을 피할 수 있다.
상기 실시예에서, 본 발명자들은 2개의 뮤린 폐 섬유화증 모델, 즉 블레오마이신-유도 섬유화증(실시예 16) 및 방사선조사-유도 섬유화증(실시예 15)에서 뮤린 항-ανβ6 단일클론 항체, 즉 mu3G9 처리의 효능을 특징규명하였다. 또한, 정상 마우스에서 mu3G9의 효과는 독성학적 보고에서 상세히 기재되어 있다. 본 실시예에서, 본 발명자들은 정상 마우스 및 폐 섬유화증의 질환 모델에서 폐 내의 mRNA 및 단백질 수준에 대한 mu3G9 처리의 효과를 특징규명한다.
폐 조직의 전사체 프로파일링은 ανβ6 차단이 블레오마이신-유더 폐 섬유화증과 관련되어 있는 TGF-β 표적 유전자의 변화를 약화시킨다는 것을 입증한다. 이 데이타는 ανβ6이 폐 섬유화증의 조절에 관여하고 그의 치료적 조절을 위한 새로운 분자 표적을 제공할 수 있다는 것을 시사한다.
재료 및 방법
1. 시약 .
ανβ6 mAb는 본 명세서의 다른 곳에 기재되어 있고 종래 보고된 바와 같이 생성하였다(29). 인간/마우스 키메라 2A1 및 3G9 cDNA는 제1 가닥 cDNA 합성 키트(Amersham/Pharmacia, Piscataway, NJ)를 사용하여 중쇄에 대한 불변 영역 프라이머 CDL-739 및 경쇄에 대한 불변 영역 프라이머 CDL-738로 각각의 모 하이브리도마 총 RNA로부터 생성하였다. 중쇄 및 경쇄 가변 영역 유전자는 cDNA 합성에 사용되는 동일한 3' 프라이머, 대부분의 뮤린 항체 유전자 신호 서열(요청 시 이용가능한 서열)에 특이적인 축퇴 프라이머의 풀 및 Pfu DNA 중합효소(Stratagene, La Jolla CA)를 사용하여 중합효소 연쇄 반응으로 증폭시켰다. 클로닝된 중쇄 및 경쇄 가변 영역들을 인간 IgG1 불변 영역과 함께 포유동물 발현 벡터 내로 연결시켰다. 재조합 가용성 뮤린 TGF-β 수용체 타입 II-Ig 융합 단백질(sTGF-βRII-Ig)은 종래 보고된 바와 같이 생성하였다(10). 연구-등급 mu3G9, 1E6 및 sTGF-bRII-Ig(포스페이트 완충 식염수 중의 정제된 단백질)를 모든 실험에서 사용하였다.
2. 동물 .
(a) RNA 분석에 대한 정상 마우스에서의 mu3G9 처리: 정상 C57B16 마우스를 5 mg/kg의 가용성 TGFbRII-Ig, PBS 또는 하기 투여량의 mu3G9로 4주 동안 주당 1회씩 (1일, 8일, 15일 및 22일) 처리하였다: 0.3, 1, 3, 10 및 30 mg/kg. 처리된 마우스의 한 무리를 29일째 날에 (마지막 투여 후 1주) 모으고, 다른 무리는 78일째 날에 (마지막 투여 후 8주 = 7-주 회수) 모았다. 이 실험은 CD-1 마우스 종이 시험되고 8주 회수가 이용된 mu3G9 마우스 독성학적 보고에 기재된 연구와는 독립적으로 수행되었다.
(b) BAL 단백질의 다중-분석물 프로파일링을 위한 정상 마우스에서의 mu3G9 처리: 정상 C57B16 마우스는 PBS 또는 하기 투여량의 mu3G9로 4주 동안 (1일, 8일, 15일 및 22일) 주당 1회씩 처리하였다: 0.1, 0.3, 1, 3 및 10 mg/kg. 기관지폐포 세척(BAL) 수거를 위해 29일째 날에 (마지막 투여 후 1주) 마우스를 모았다. 이 실험들은 CD-1 마우스 종이 시험된 mu3G9 마우스 독성학적 보고에 기재된 연구와는 독립적으로 수행되었다.
(c) 방사선조사 섬유화증 모델에서의 mu3G9 처리: 방사선조사-유도 섬유화증을 약화시키는 효능을 평가하는 데 사용된 처리군 및 판독대상에 대한 세부사항은 BIIB 보고서 #Rsch-2006-007에 포함되어 있다. 요약하면, 마우스의 흉선에 방사선을 조사하고 방사선조사 후 15주째부터 마우스를 주마다 0.3 내지 10 mg/kg의 다양한 투여량의 mu3G9로 처리한 3개의 연구를 뉴욕 의대(New York School of Medicine)의 존 멍거(John Munger) 박사의 연구실에서 수행하였다. 마우스 폐는 마우스가 본 연구 동안 죽은 상태로 발견되면 조직학적 검사/섬유화증 측정을 위해 수거하였다. 생존 마우스는 방사선조사 후 26주, 28주 및 32주째에 수거하였다. BAL 액은 하나의 폐 엽으로부터 수거하여 바이오겐 아이덱에 보낸 반면, 다른 폐 엽은 조직학적/섬유화증 측정을 위해 처리하였다. BAL 액 중의 단백질의 분석은 바이오겐 아이덱에서 mu3G9로 처리된 정상 마우스로부터의 BAL 액의 분석과 비교하기 위해 이 보고에 포함된다.
3. 기관지폐포 세척 수거 . 마우스를 과량의 이낵틴(Sigma) 복강내 주사(IP)로 마취시켰다. 가장 무딘 해부를 이용하여 기관을 노출시켰다. 그 후, 기관을 두 개의 연골 고리 사이에서 가위로 개방하고, 23-게이지의 무딘 말단 바늘을 기관 내로 삽입하였다. 슈와르츠 임시 클립(Roboz)으로 약하게 클램핑함으로써 바늘을 제자리에 고정시켰다. Ca2+ 또는 Mg2+를 함유하지 않는 포스페이트 완충 식염수(PBS) 0.8 ㎖를 폐 내로 조사함으로서 BAL을 수행하였다. 이어서, 많은 압력을 인가하지 않으면서 유체를 시린지 내로 빨아들이고, 얼음 상의 15 ㎖ 폴리프로필렌 팔콘 튜브 내로 옮겼다. 그 후, 상기 절차를 반복하고, 과도한 음의 압력을 발휘하지 않으면서 BAL 액을 시린지 내로 다시 빨아들였다. 샘플을 얼음 상에서 저장하고 세포 계수 및 분화를 위해 처리하고, BAL 펠릿 및 유체 둘다를 수거하고 -80℃에서 저장하였다.
1. 그 다음, BAL을 4℃에서 탁상용 원심분리기 상에서 180 g(1000 rpm)(Beckman GPR)으로 10분 동안 원심분리하였다. 그 후, 상청액(BAL 액)을 분리하여 -80℃에서 동결시켰다. 1.0 ㎖의 RBC 용해 용액(Sigma)을 펠릿에 첨가하고 20초 동안 볼텍싱하였다. 이어서, 세포 계수 및 사이토스핀을 행하였다.
2. 샘플을 얼음 상에서 저장하고, 혈구계를 이용하여 세포의 수를 세었다. 세포를 혈구계에 가한 후 판독하기 전에 세포가 1분 동안 가라앉게 하였다.
3. 100 ㎕의 세포 현탁액을 실온에서 500 RPM으로 5분 동안 사이토스피닝하였다(ThermoShandon). 사이토스핀 제제는 세포 용액을 사이토스핀 장치 내에 넣고 500 rpm에서 5분 동안 원심분리함으로써 준비하였다. 세포 농도가 슬라이드 상에서 너무 많지 않도록 점검하였다. 슬라이드를 밤새 건조한 후, 디프퀵(Fisher)으로 염색하였다. 염색은 제조자(DiffQuik)의 프로토콜에 따라 수행하였다. 일단 슬라이드가 건조되고 퍼마운트(Fisher)로 커버 슬립을 덮으면, 세포를 유형별로 분류하고 실험실 세포 계수기(Fisher)를 이용하여 100개의 무작위로 선택된 세포의 수를 세었다.
4. RNA를 위한 폐 수거 . 마우스를 과량의 이낵틴(Sigma) 복강내 주사(IP)로 마취시켰다. 70% ETOH을 동물에게 분무하고, 1쌍의 멸균 가위를 사용하여, 흉골부터 시작하여 머리를 향하여 피부를 절개하였다. 일단 피부가 제거되면, 흉골 및 늑골을 절개하여 심장 및 폐를 노출시켰다. 폐를 분리하여 멸균된 거즈(gauze) 조각 위에 배치하여 임의의 혈액 생성물을 제거하고 14 ㎖ 폴리프로필렌 둥근 바닥 튜브 17x100 mm(Fisher) 내로 신속히 넣었다. 액체 질소를 폐가 함유된 폴리프로필렌 튜브에 첨가하고 드라이 아이스 상에 배치하였다. 그 다음, 폐를 -80℃에 저장하였다.
5. RNA 준비 . 제조자의 프로토콜에 따라 퀴아졸 시약(Qiagen)을 사용하여 급속-동결 폐 조직 샘플로부터 총 RNA를 정제하였다. RNA 질은 바이오어날라이저(Bioanalyzer) 2000(Agilent) 상에서의 모세관 전기영동에 의해 검증되었다.
6. 프로브 표지, 혼성화 및 전사체 프로파일링을 위한 스캐닝 .
샘플 표지, 혼성화 및 염색을 유전자칩® 발현 분석(Affymetrix, Santa Clara, CA)에 대한 아피메트릭스 테크니칼 메뉴얼(701021 rev 1)의 진핵 표적 준비(Eukaryotic Target Preparation) 프로토콜에 따라 수행하였다. 요약하면, 5 μg의 정제된 총 RNA를 200 U 수퍼스크립트 II(Superscript II)(cat # 18064-022, Invitrogen) 및 0.5 μg의 (dT)-T7 프라이머[5'-GGCCAGTGAATTGTAATACGACTCACTATAGGGAGGCGG(T)24]와 함께 42℃에서 20 ㎕ 제1 가닥 반응 중에서 1시간 동안 사용하였다. 40 U의 이. 콜라이 DNA 중합효소(cat # 18010-025, Invitrogen), 2 U의 이. 콜라이 RNase H(cat # 18021-071, Invitrogen) 및 10 U의 이. 콜라이 DNA 리가제(cat # 18052-019, Invitrogen)를 첨가한 후 16℃에서 2시간 동안 항온처리함으로써 제2 가닥 합성을 수행하였다. 제2 가닥 합성 반응액은 유전자칩® 샘플 클린업 모듈(cat # 900371, Affymetrix, Santa Clara, CA)을 제조자의 프로토콜에 따라 사용하여 정제하였다. 정제된 cDNA는 바이오어레이 고수율 RNA 전사 표지 키트(cat # 42655-40, Enzo Life Sciences, Inc., Parmingdale, NY)를 제조자의 프로토콜에 따라 사용하여 증폭시킴으로써 바이오틴으로 표지된 cRNA(상보적 RNA) 70 내지 120 μg을 생성하였다. 마우스 MgU74Av2, MgU74Bv2 및 MgU74Cv2 유전자칩® 프로브 어레이를 제조자의 프로토콜에 따라 유전자칩® 혼성화 오븐 640(Affymetrix, Santa Clara, CA)에서 예비혼성화시켰다. 단편화된 표지 cRNA를 100 mM 2-모르폴리노에탄설폰산, 1 M [Na+], 20 mM EDTA, 0.01% Tween 20, 0.5 mg/㎖ 아세틸화 BSA, 0.1 mg/㎖ 헤링 정자 DNA, 대조군 올리고 B2 및 대조군 전사체 바이오B 1.5 pM, 바이오C 5 pM, 바이오D 25 pM 및 cre 100 pM이 함유된 300 ㎕ 1X 혼성화 완충제 중에 재현탁시키고, 제조자의 프로토콜(Affymetrix, Santa Clara, CA)에 따라 유전자칩® 프로브 어레이에 혼성화시켰다. 혼성화된 유전자칩® 프로브 어레이를 세척하고 스트렙타비딘-파이코에리쓰린(cat # S866, Molecular Probes, Eugene, OR)을 사용하여 염색하고, 유전자칩® 플루딕스 스테이션 400(GeneChip® Fluidics Station 400)(Affymetrix, Santa Clara, CA)을 사용하여 바이오티닐화 항-스트렙타비딘(BA-0500, Vector Laboratories, Burlingame, CA)으로 증폭시켰다. 유전자칩® 프로브 어레이를 진어레이 스캐너(Hewlett Packard, Corvallis, OR)를 사용하여 스캐닝하였다.
7. 전사체 프로파일링 데이타 분석 .
어레이 스캔을 아피메트릭스로 전환하였다. CEL 파일 및 생성된 데이타 세트(완전한 실험을 대표하는 CEL 파일 군)를 로버스트 마이크로어레이 애버리지(Robust Microarray Average; RMA) 방법을 이용하여 표준화하였다. 통계적 분석 및 클러스터링 분석은 BRB 어레이 수단 v. 3.4.0 - 베타 2(NCI), 진스프링(GeneSpring)(Agilent) 및 스팟파이어(Spotfire) 데이타 분석 수단을 이용하여 수행하였다. 신뢰 한계(CL)가 0.95를 초과하지 않도록 설정된 가짜 발견율(False Discovery Rate(FDR)) 역치를 가진 PBS-처리 군에 비해 실험 처리들 중 임의의 처리에 의해 그 신호 강도가 변경되지 않은 프로브세트를 확인하기 위해 마이크로어레이의 유의성 분석(SAM)을 사용하였다. 필요하다면, 신호 강도에서 2-배 이상의 변화를 보이는 유의하게 영향을 받은 (FDR<0.01, CL 0.95) 프로브세트를 선택함으로써 추가 여과를 수행하였다. 생성된 유전자 군의 프로파일 및 실험 조건의 분류를 계층적 클러스터링으로 분석하고 가시화하였다. IPA 데이타베이스(Ingenuity Systems)를 이용하여 실제 경로 분석을 수행하였다.
8. BAL 액 단백질 수준의 다중 분석 . 200 ㎕의 측정용 분취액을 전술한 바와 같이 수거한 BAL 액으로부터 직접 얻었다. 이 분취액을 룰스 베이스드 메디슨 인코포레이티드(Rules Based Medicine, Inc.)(Austin, TX)에 보내고, 거기서 루미넥스-기초 기술(Luminex-based technology)을 이용하여 마우스 단백질의 표준 패널에 대해 분석하였다.
9. 통계적 분석 . 전사체 프로파일링 분석을 위한 통계적 분석은 전술한 바와 같다. BAL 액 중의 단백질 수준을 분석함에 있어서, 일방 편차 분석(ANOVA)을 이용하여 비히클 대조군 및/또는 이소타입 대조군과 다양한 투여량의 시험 제품을 통계적으로 비교하였다. 통계적으로 유의한 차이가 ANOVA를 이용하여 p<0.05의 확률에서 확립되면, 군 사이의 유의한 차이를 던넷(Dunnet) 다중 비교 시험으로 평가하였다.
결과
1. mu3G9를 사용한 정상 마우스의 처리 효과: 폐 전사체 분석 . CD-1 마우스에서의 독성학적 연구의 결과는 폐가 mu3G9에 대한 독성의 표적 기관임을 확인하였다. 이 연구에서 상세히 기재되는 바와 같이, (ανβ6 인테그린이 결핍된) 인테그린 β6 널 마우스에서의 발견과 일치하는 폐 염증은 mu3G9로 처리된 마우스에서 관찰된다. 조직병리학적으로 평가하였을 때, 이 염증은 1 mg/kg의 투여량에서 흔하지 않게 관찰되지만, 주당 10 mg/kg의 투여량에서는 일관되게 관찰된다.이 염증을 C57B16 종(실시예 15 및 16에서 전술한 바와 같은 효능 연구에서 사용된 종)에서 평가하기 위해, 마우스를 주당 0.3 내지 30 mg/kg의 투여량의 mu3G9로 처리하고, 폐를 RNA 및 마이크로어레이 분석을 위해 처리하였다.
FDR 역치가 0.01이고 CL이 0.95인 마이크로어레이의 유의성 분석(SAM)을 이용하여, PBS-처리 대조군과 각각의 실험적 처리군(3G9로 처리된 군 및 sTGFbRII-Ig 군을 포함함) 사이의 일련의 쌍별(pair-wise) 비교에서 차등적으로 발현되는 유전자를 검색하였다. 이러한 쌍별 SAM 분석은 그들의 각 PBS 대조군을 사용하여 처리군 및 회수군에 대해 개별적으로 수행하였다. 수득된 유전자 목록을 추가 여과 단계로 처리하여, 신호 강도가 PBS 대조군과 처리군 중 임의의 처리군 사이에 2-배 이상 변화되어 있는 유의한 영향을 받은 프로브세트의 유전자 목록을 확인하였다. 결과는 하기 표 17-1 내지 17-6에 요약되어 있다. 상기 선별 기준을 만족시키는 유전자 발현의 유의한 변화는 처리군의 10 mg/kg 및 30 mg/kg 하위군에서 관찰되었다(표 17-1 및 17-2). 선별된 프로브 세트에 대한 유전자 발현의 프로파일은 10 mg/kg 및 30 mg/kg의 3G9 처리군들에서 상응하는 유전자들의 발현 수준에서의 강한 쌍방향 변동을 보여주었다(도 66).
유전자 발현의 통계적으로 유의한 변화는 임의의 다른 처리 하위군 또는 회수군에서 전혀 검출되지 않았다(표 17-1 및 17-2). 그러나, MMP12, 및 이 MMP12의 발현 프로파일과의 피어슨 상관관계가 0.95를 초과하는 발현 프로파일을 보이는 선별된 다른 유전자들은 3 mg/kg의 3G9 처리군에서 검출가능한 상승 경향을 보였다(도 67; 표 17-7).
3G9에 의해 유의하게 영향을 받는 유전자의 기능적 해석은 IPA 데이타베이스를 사용하여 수행하였고, 이 유전자들과 면역 반응 및 면역조절 사이토카인 신호전달의 강한 관련성을 보였다(도 68). 유사하게, 실제 조절 경로 분석은 유전자 발현에서의 3G9-유도 변화와 케모카인, TGF-β 및 인터페론 신호전달에서의 변경의 관련성을 암시한다. 이 관련성은 2개의 최대-스코어링 네트워크의 구조의 당연한 결과이다(도 69A 및 69B).
2. MMP-12 전사체의 정량적 PCR 분석 . MMP-12는 mu3G9-처리 마우스로부터 얻은 폐 내의 임의의 전사체의 최대 배수-상향조절을 보인다(표 17-3). MMP-12는 β6 널 마우스의 폐에서 가장 많이 상향조절된 전사체로서 이미 보고되었다. MMP-12 발현과 mu3G9 처리 사이의 관계를 더 밝히기 위해, 본 발명자들은 β6 널 마우스의 폐로부터 얻은 RNA를 비롯한 MMP-12 전사체의 상대적 수준을 정량적 PCR로 분석하였다. mu3G9 처리는 10 및 30 mg/kg에서 유의한 MMP-12 전사체의 투여량-의존적 증가를 나타내었다. MMP-12 발현의 배수-변화는 상기 투여량에서 β6 널 마우스에서 관찰된 것과 필적할만하였다. 마이크로어레이 분석에 대해 전술한 결과와 유사하게, qPCR에 의한 MMP-12 수준은 3 mg/kg 투여량에서 상승되어 있었지만, 0.3 및 1 mg/kg 투여량에서는 변화하지 않았다.
3. mu3G9로 처리된 정상 마우스의 폐로부터 얻은 BAL의 단백질 분석 . mu3G9 처리에 기인하는 폐 내의 분자적 변화를 더 특징규명하기 위해, 본 발명자들은 0.1 내지 10 mg/kg의 투여량의 mu3G9로 4주 동안 처리된 마우스의 또 다른 무리로부터 얻은 BAL 액 중의 60종의 단백질의 수준을 분석하였다(표 17-8). 단백질 분석은 룰스 베이스드 메디슨 인코포레이티드(Austin, TX)에서의 루미넥스 (다중 단백질 분석) 측정에 의해 수행되었다. 전사체 수준에서의 발견과 일치하여, 폐 염증과 관련된 다수의 단백질들이 10 mg/kg 투여량으로 처리된 마우스의 BAL 액 중에서 상승되었다(표 17-9). 추가로, 이 변화들 중 일부는 3 mg/kg의 투여량에서도 ANOVA에 의해 유의한 것으로 확인되었으나, 패널 상의 단백질은 1 mg/kg 투여량에 의해 상승되었다.
4. 방사선조사-유도된 섬유화증 모델로부터의 BAL 액 단백질의 분석 . 방사선조사-유도 폐 섬유화증을 약화시키는 데 있어서 mu3G9의 효능은 실시예 15에서 전술되어 있다. 본 연구에서 수거된 BAL 액(BALF) 샘플은 mu3G9로 처리된 정상 마우스로부터의 BALF에서 사용된 것과 동일한 마우스 패널을 사용하여 다중-분석물 단백질 프로파일링으로 분석하였다. 28주(13주 처리) 및 32주(17주 처리) 시점의 BALF 분석이 본원에서 분석되었다. ANOVA에 의한 측정 시 유의하게 변화된 단백질은 표 17-11에 요약되어 있다. 정상 마우스에서의 전사체 프로파일링 및 단백질 결과와 마찬가지로, 방사선조사 섬유화증 모델에서 변경된 단백질들의 대부분은 10 mg/kg 투여량에서만 유의한 수준에 도달하였지만, 일부는 3 mg/kg에서 유의하였고, 모든 단백질들이 3 mg/kg에서 변화 경향을 보였다. 이 단백질들의 대다수는 폐 섬유화증과 관련된 것으로 공지된 사이토카인 또는 케모카인이다. 10 mg/kg mu3G9로 처리된 정상 마우스에서 2-배 이상까지 상향조절된 20종의 단백질들(표 17-19) 중 14종의 단백질들은 방사선조사 모델에서도 일관되게 상향조절된다. mu3G9 처리 기간 및 손상/염증 상태에서의 차이에도 불구하고, 정상 마우스에서의 발견과 방사선을 조사받은 마우스에서의 발견 사이의 일치는 놀라울 정도이다. 상기 단백질들 중 대부분이 방사선조사 모델에서 더 긴 처리 기간과 일치하는 다소 더 높은-배수 상향조절을 보이지만(표 17-12), 전체적인 발견은 이들 2가지 매우 상이한 셋팅에서 일치한다. 또한, 정상 마우스와 방사선을 조사받은 마우스에서의 상이한 기준 수준에도 불구하고 (방사선조사 손상은 이 단백질들의 대부분을 상향조절함), 이 염증성 표지들이 상향조절되는 투여량 범위는 정상 마우스와 방사선을 조사받은 마우스에서 동일하다. 구체적으로, 이들은 3 및 10 mg/kg 투여량에서 상향조절되지만, 정상 마우스 및 병든 마우스 둘다에서 1 mg/kg의 투여량에서는 상향조절되지 않는다(도 70에서 28주에 가장 높게 상향조절된 4종의 단백질들로 예시됨). 따라서, 방사선조사 모델에서 약물 표적의 훨씬 더 높은 발현에도 불구하고 (상기 실시예 15 참조), mu3G9 처리에 특이적인 BAL 단백질 변화를 나타내는 투여량은 두 셋팅에서 동일하다. mu3G9에 의해 유도된 단백질들 모두가 전구-염증성 및/또는 전구-섬유화증성 효과를 가지는 것으로 생각되지는 않는다는 것을 인식하는 것이 중요하다. 예를 들면, CXCL 케모카인 IP-10/CXCL1O은 마우스 모델에서 강력한 항-섬유화증성 효능을 나타내며, IP-10 또는 그의 수용체 CXCR3이 결핍된 마우스는 블레오마이신에 대한 악화된 섬유화증성 반응을 보인다. 그러나, 일치하는 변화가 최대 유효 투여량인 1 m/kg 투여량 근처에서 이 사이토카인 또는 다른 사이토카인의 경우에는 발견되지 않으므로, 폐의 주위에 있는 사이토카인에서의 이 변화들 중 어떠한 변화도 mu3G9 처리의 효능에 필요한 것 같지 않다.
정상 마우스에서 상향조절되었으나 방사선조사 모델에서는 상향조절되지 않은 단백질들 중에는 ApoA1, 피브리노겐 및 TIMP-1이 있었다. 이 단백질들의 수준 및 제4 단백질, 즉 햅토글로빈의 수준은 정상 마우스에 비해 방사선을 조사받은 마우스의 BAL 액에서 증가되지만, mu3G9 처리에 의해 감소된다(도 71). 따라서, 유효 투여량에서 이 단백질들에 대한 수준의 표준화는 이들이 28주 시점에서 효능의 대리 마커로서 작용할 수 있다는 것을 시사한다(도 72). 더 낮은 투여량에서 유의하게 변화된 패널 상의 단백질만이 처리, 즉 TIMP-1 및 햅토글로빈에 의해 감소된 단백질이었다. TIMP-1은 공지된 TGF-β 표적이고 섬유화증성 질환에서 종종 상승되어 있다. 이의 하향조절은 mu3G9의 작용 기작, 즉 ανβ66-매개 TGF-β 활성화의 억제와 일치한다.
[표 17-1]
mu3G9의 생체 내 투여량 상승에 대한 폐 전사체 변화의 분석. 프로브 세트의 수는 실험 처리에 의해 유의하게 영향을 받았다(FDR < 0.01, CL 0.95).
[표 17-2]
2-배 초과의 변화를 보이는 폐 전사체의 분석. 실험 처리에 반응하여 신호 강도에서 2-배 초과의 유의한 변화를 보이는 프로브세트의 수.
[표 17-3]
[표 17-4]
[표 17-5]
[표 17-8]
다중-분석물 단백질 프로파일링에 의해 분석된 총 단백질
[표 17-9]
다양한 투여량의 mu3G9 처리에서 변화된 단백질
[표 17-10]
정상 마우스에서 2-배 초과의 변화를 보이는 BALF 단백질
[표 17-11]
방사선조사 모델에서 유의하게 변화된 BAL 액 단백질
[표 17-12]
정상 마우스와 방사선을 조사받은 마우스에서 BAL 단백질에서의 배수-변화의 비교
결론
mu3G9-처리의 효과는 정상 마우스에서 하기 전사체 프로파일링 및 다중 분석물 단백질 프로파일링을 특징으로 한다:
(a) 높은 투여량의 mu3G9-처리 마우스의 폐의 전사체 프로파일링 분석은 폐 염증과 관련된 다수의 전사체의 유의한 변화를 유도한다. 이 변화들은 인테그린 β
6
널 마우스(α
ν
β
6
-결핍 마우스)로부터 얻은 전사체 프로파일링 결과와 일치한다. 잘못 조절된 특정한 표적은 mu3G9의 작용 기작으로 인해 폐에서의 감소된 TGF-β 신호전달과 일치한다.
(b) 단백질 프로파일링 결과도 마찬가지로 mu3G9의 작용 기작과 관련된 일부 사이토카인 및 케모카인에서 일관된 증가를 나타낸다.
(c) 다수의 전사체가 3 m/kg의 투여량에서 상승되는 경향이 있다 하더라도, 전사체에서의 변화는 10 mg/kg 투여량에서만 유의한 수준에 도달하였다. 일부 단백질 변화는 3 mg/kg 투여량에서 유의하지만, 대부분은 10 mg/kg 투여량에서 유의한 수준에 도달한다. 1 mg/kg 투여량에서는 단백질 또는 전사체에서의 변화가 전혀 유의하지 않았다.
mu3G9-처리의 효과는 방사선조사 섬유화증 모델에서 하기 다중 분석물 단백질 프로파일링을 특징으로 한다:
(a) 정상 마우스에서 높은 투여량의 mu3G9에 의해 유도되는 동일한 사이토카인 및 케모카인의 대다수도 방사선조사 섬유화증 모델에서 높은 투여량의 mu3G9에 의해 상승된다.
(b) mu3G9의 표적, 즉 인테그린 α
ν
β
6
의 훨씬 더 높은 발현에도 불구하고, 이 염증 마커들에서의 상승은 방사선조사 모델 및 정상 마우스에서 동일한 투여량에서, 즉 3 및 10 mg/kg에서 관찰되지만, 이 모델에서 거의 최대 유효 투여량인 1 mg/kg에서는 관찰되지 않는다(실시예 15 참조).
섬유화증과 관련된 일부 단백질들, 특히 TIMP-1 및 햅토글로빈은 28주 시점에서 유효 투여량의 mu3G9 처리에 의해 정상화되었다.
참고문헌
본 발명은 명확한 이해의 목적으로 설명 및 실시예로서 다소 상세히 전체적으로 기재되어 있지만, 본 발명의 범위 또는 이의 임의의 구체적 실시양태에 영향을 주지 않으면서 넓은 동등한 범위의 조건, 제제 및 다른 매개변수 내에서 본 발명을 변형시키거나 변화시킴으로써 본 발명을 수행할 수 있다는 것과, 이러한 변형 또는 변화가 첨부된 청구의 범위 내에 포함된다는 것은 당업자에게 자명할 것이다.
본 명세서에서 언급된 모든 문헌, 특허 및 특허 출원은 본 발명이 속하는 분야에서 숙련된 자의 기술 수준을 의미하고, 마치 각 개별 문헌, 특허 또는 특허 출원이 참고로 도입되도록 구체적 및 개별적으로 기재되는 것처럼 참고로 본 명세서에 도입된다.