정의
본 발명에서 사용되는 바와 같이, "D-자일로오스 및 D-자일룰로오스 이외의 탄소원"은, D-자일로오스 및 D-자일룰로오스 이외의 자일리톨 생성을 위한 탄소 기질 또는 그의 중합체 또는 올리고머 또는 혼합물(예를 들어, 자일란 및 헤미셀룰로오스)을 의미한다. 상기 탄소원에는 바람직하게는 D-글루코오스, 및 다양한 D-글루코오스-함유 시럽 및 D-글루코오스와 기타 당류의 혼합물이 포함된다.
본 발명에서 사용되는 바와 같이, "유전자"는, 단백질을 코드화할 수 있는 핵산 서열, 특히 DNA 서열을 의미한다.
본 발명에서 사용되는 바와 같이, "벡터"는, 유전자 정보, 특히 DNA를 숙주 세포로 전달할 수 있는 플라스미드 또는 임의의 기타 DNA 서열을 의미한다. 벡터는 벡터로 형질전환된 세포의 동정에 사용하기에 적합한 마커 또는 리포터 및 하나 이상의 원핵 또는 진핵 숙주에서의 벡터의 유지 및 복제를 가능하게 하는 복제 기점을 추가로 포함할 수 있다. "플라스미드"는 적어도 하나의 숙주 세포에서 자체적으로 유지 및 복제되는 벡터, 일반적으로는 원형 DNA이다.
본 발명에서 사용되는 바와 같이, "발현 벡터"는, 벡터와 유사하나, 숙주로 형질전환된 후, 클로닝된 유전자 또는 인코딩 핵산의 발현을 지원하는 벡터를 의미한다. 클로닝된 유전자 또는 인코딩 핵산은 통상적으로, 벡터에 의해 또는 클로닝된 유전자의 재조합 작제물에 의해 제공될 수 있는 특정 조절 서열, 예를 들어 프로모터 서열의 조절하에 배치된다(즉, 작동적으로 연결된다). 발현 조절 서열은, 벡터가 원핵 또는 진핵 숙주에서 작동적으로 연결된 유전자를 발현하도록 설계되었는지에 따라 달라지며, 전사 인자, 예를 들어 인핸서 인자(상류 활성화 서열) 및 종결 서열 및/또는 번역 개시 및 종결 부위를 추가로 포함할 수 있다.
본 발명에서 사용되는 바와 같이, "숙주"는 재조합 물질의 수용체 및 운반체로서 이용되는 세포, 원핵 또는 진핵 세포를 의미한다.
본 발명에서 사용되는 바와 같이, "5탄당-포스페이트 경로의 산화 분지"는, D-글루코오스-6-포스페이트 탈수소효소(EC 1.1.1.49) 글루코놀락토나아제(EC 3.1.1.17), 및 6-포스포-D-글루코네이트 탈수소효소(EC 1.1.1.44)에 의해 촉매되는 반응과 같은 산화 반응을 촉매하며, 5탄당 포스페이트의 생성을 위해, 6탄당 기질을 이용하는 5탄당-포스페이트 분로의 일부가 포함된다는 것을 의미한다.(또한, D-글루코오스로부터 리보오스의 순 생성을 촉매하는) 5탄당-포스페이트 경로의 "비산화" 부분은, 트랜스케톨라아제(EC 2.2.1.1), 리보오스-5-포스페이트 이성질화 효소(EC 5.3.1.6), D-리불로오스-5-포스페이트-3-에피머라아제(EC 5.1.3.1) 및 트랜스알돌라아제(EC 2.2.1.2)에 의해 촉매되는 반응과 같은 비산화 이성질화를 특징으로 한다. 문헌[Biological Chemistry, H.R. Mahler & E.H.Cordes, Harper & Row, publishers, New York, 1966, pp. 448-454]을 참조한다.
본 발명에서 사용되는 바와 같이, "인코딩 핵산"은 핵산 분자(바람직하게는 DNA)를 의미한다. 인코딩 핵산은 단백질을 인코딩할 수 있으며, 다양한 공급원으로부터 제조될 수 있다. 이러한 공급원에는 게놈 DNA, cDNA, 합성 DNA 및 그의 조합이 포함된다
본 발명에서 사용되는 바와 같이, "이종"은, 유전자 또는 인코딩 서열이 유전자 조작에 의해 세포 내로 도입되는 것을 의미하는 것으로 이해된다. 이것은 에피솜 또는 염색체 형태로 존재할 수 있다. 유전자 또는 인코딩 서열은 도입된 숙주 세포와 다른 공급원으로부터 유래할 수 있다. 그러나, 이것은 또한, 도입되었으나, 자연적이지 않은 그의 환경으로 인해, 이종으로 간주되는 숙주 세포와 동일한 종류로부터 유래할 수 있다. 예를 들어, 유전자 또는 인코딩 서열은, 그의 천연 프로모터가 아닌 프로모터의 제어하에 있으며, 자연적 위치와는 다른 위치에 도입되기 때문에, 이종으로 지칭된다. 숙주 세포는 이종 유전자의 도입 전에 상기 유전자의 내인성 카피를 함유할 수 있거나 또는 내인성 카피를 함유하지 않을 수 있다.
본 발명의 목적
본 발명에 따르면, 특이적 미생물 숙주의 천연 대사 경로는, 자일리톨 생성 이외의 목적으로의 탄소의 이용이 감소 또는 제거되도록, 조작된다.
따라서, 이러한 유전자 변형 숙주 균주는 하나의 발효 단계에서 자일리톨을 고 수율로 생성할 수 있다. 예를 들어, 48시간 배양 후, 상청액의 자일리톨 역가는 15g/l 이상, 바람직하게는 25g/l 이상, 더욱 바람직하게는 50, 60, 70, 80, 90 또는 100g/l 이상이다.
본 발명의 실제 구현에서, 본 발명의 유전자 변형 숙주는 또한, 바로 D-자일로오스 및/또는 D-자일룰로오스로부터가 아닌, 구조적으로 유사하지 않은 탄소원, 예를 들어 D-글루코오스로부터 자일리톨을 합성할 수 있는 그의 능력을 특징으로 한다.
바람직하게는, 본 발명의 유전자 변형 숙주는 또한 합성 자일리톨을 배지로 분비할 수 있다.
구체적으로, 예시적인 바람직한 실시형태에서, 본 발명의 유전자 변형 숙주는, 아라비톨이 자일리톨 생성의 중간체인 경로를 특징으로 한다.
따라서, 본 발명의 재조합 숙주 균주는 하기와 같은 유전자 변이를 특징으로 한다:
(1) NAD+-특이적 D-아라비톨 4-산화환원효소(D-자일룰로오스-생성) 활성을 가지는 단백질을 인코딩하는 이종 핵산이 숙주 세포로 도입되어 - 따라서, D-아라비톨의 D-자일룰로오스로의 전환이 제공되며;
(2) NADPH-특이적 자일리톨 탈수소효소 활성을 가지는 단백질을 인코딩하는 이종 핵산이 숙주 세포로 도입되어 - 따라서, D-자일룰로오스의 자일리톨로의 전환이 제공된다.
미생물의 선택
본 발명에 적합한 미생물 또는 숙주 균주는 글루코오스로부터 D-아라비톨을 생성할 수 있는 것이다. 더욱 특히, 이들은 고삼투압 배지에서, 글루코오스로부터 다량의 D-아라비톨을 생성할 수 있다.
본 명세서에서, "고삼투압 배지"는 10-60% D-글루코오스, 바람직하게는 약 25% D-글루코오스를 포함하는 배지를 지칭하는 것으로 의도된다.
"다량의 D-아라비톨"은 적어도 100 g/L의 D-아라비톨을 의도한다. 특히, 미생물 또는 숙주 균주가 회분 조건(batch condition)에서 25 % D-글루코오스를 함유하는 배지에서, 100g/L D-아라비톨을 생성하는 경우, 상기 미생물 또는 숙주 균주는 다량의 D-아라비톨을 생산하는 것으로 간주된다.
글루코오스로부터 다량의 D-아라비톨을 생성할 수 있는 숙주 균주의 예에는 호삼투성 또는 내삼투성 효모, 특히 피치아(Pichia), 코다마에아(Kodamaea), 칸디다(Candida), 지고아카로마이세스(Zygoaccharomyces), 데바로마이세스(Debaromyces), 메트쉬니코비아(Metschnikowia) 및 한세눌라(Hansenula) 종에 속하는 것; 또는 D-아라비톨 생성 균류, 특히 덴드리피 엘라(Dendryphiella) 및 스키조필룸(Schizophyllum ) 종에 속하는 것, 특히 덴드리피엘라 살리나(Dendryphiella salina) 및 스키조필룸 콤뮨(Schizophyllum commune)이 포함된다.
피치아 속의 미생물의 예에는 피치아 오메리, 피치아 스티피티스, 피치아 파 리노사(Pichia farinosa ), 피치아 하플로필라(Pichia haplophila)이 포함된다. 칸디다 속의 미생물의 예에는 칸디다 폴리모르파 및 칸디다 트로피칼리스(Candida tropicalis)가 포함된다. 지고아카로마이세스 속의 미생물의 예에는 지고아카로마이세스 룩시이(Zygoaccharomyces rouxii )가 포함된다. 기타 예에는 토룰롭시스 칸디다 및 토룰라스포라 한세니이(Torulaspora hansenii)가 포함된다. 메트쉬니코비아 속의 미생물의 예에는 메트쉬니코비아 풀케리마(Metschnikowia pulcherrima), 메트쉬니코비아 레우카우피 ( Metschnikowia reukaufii), 메트쉬니코비아 비쿠스피다타 ( Metschnikowia bicuspidata ), 메트쉬니코비아 루나타(Metschnikowia lunata) 및 메트쉬니코비아 조벨리이(Metschnikowia zobellii)가 포함된다. 특이적 균주로서, 메트쉬니코비아 풀케리마 ATCC 18406, 메트쉬니코비아 레우카우피 ATCC 18407, 메트쉬니코비아 비쿠스피다타 ATCC 24179, 메트쉬니코비아 루나타 ATCC 22033, 메트쉬니코비아 조벨리이 ATCC 22302 및 메트쉬니코비아 풀케리마 FERM BP-7161이 언급될 수 있다. 이들 균주는 아메리칸 타입 컬쳐 콜렉션(American Type Culture Collection)(주소: 미국 메릴랜드 20852, 록빌, 12301 파크런 드라이브)에서 구할 수 있다. 메트쉬니코비아 풀케리마 FERN BP-7161은 처음에, 수탁번호 FERM P-16592로, 1998년 1월 16일자로, 통상산업성, 산업기술종합연구소, 생명공학공업기술연구소(National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry)(주소: 일본 이바라키-켄, 츠쿠바-시, 1-3 히가시 1-초메(우편번호: 305-8566))에 기탁되었으며, 2000년 5월 15일자로, 원 기탁기관으로부터 부다페스트 조약을 기초로 한 국제 기탁기관으로 이송되었고, 수탁번호 FERM BP-7161로서, 기탁되었다. 구체적 측면에서, 상기 미생물은 등록번호 FERM BP-7161을 가진다. 더 많은 정보를 위해, EP1065276을 참조한다.
상기 미생물은 D-아라비톨을 생성하고/거나, 자일리톨 생성 이외의 목적을 위해, D-아라비톨을 이용할 수 있는 그의 능력을 감소시키는 그의 능력을 향상시키기 위해, 유전자 조작될 수 있다.
본 발명에서, 숙주 균주는 하기와 같은 그의 특이적 대사 특성에 의해, 유리하게 선택된다:
- 상기 숙주 균주는 특히 고삼투압 배지, 예를 들어, 10-60% D-글루코오스, 바람직하게는 25% D-글루코오스를 포함하는 배지("정상" 배지는 일반적으로 단지 2-3% 글루코오스를 포함함) 하에, 전술된 바와 같이, 글루코오스로부터 다량의 D-아라비톨의 생산자일 수 있으며;
- 상기 숙주 균주는 단일 탄소원으로서, D-아라비톨을 소비하지 않을 수 있고;
- 상기 숙주 균주의 산화 환원 반응 평형은 상응하는 케토5탄당/5탄당 알코올 전환에 필요한 보인자의 생성을 가능하게 한다.
본 발명의 일 실시형태에서, 호삼투성 효모 피치아 오메리(및, 그의 돌연변이 유도체)가 모델 및 바람직한 숙주로서, 사용되었다. 피치아 오메리는 처음에, 오이 염수에서 분리되었으며, 식품 산업에서, 피클, 껍질 및 과일의 발효를 위해 일반적으로 사용되었다.
피치아 , 지고사카로마이세스 , 데바로마이세스 및 한세눌라와 같은 효모 종이 사카로마이세스 세레비시애(Saccharomyces cerevisiae)와 반대로, 저 수분 활성 환경에서 성장할 수 있다는 것은 당업자에게 알려져 있다. 이들 내삼투성 또는 호삼투성 효모는, 효소를 보호하고 안정화시켜서, 따라서, 삼투성 성장 조건에서, 세포 기능을 가능하게 하는 글리세롤, D-아라비톨, 에리트리톨 및 만니톨과 같은 상용성 용질을 축적시킨다. 생성된 폴리올은 산화 환원 반응 평형에서, 역할을 한다.
바람직한 측면에서, 미생물은 피치아 오메리이다. 사실, 숙주 균주 피치아 오메리의 주된 특징은, 지고사카로마이세스 룩시이가 글리세롤과 D-아라비톨을 생산하는 것과는 대조적으로, D-아라비톨만을 상용성 용질로 생성한다는 것이다. 또한, 글루코오스에서 D-아라비톨에 이르는 대사 경로는 피치아 오메리에서 잘 알려져 있다.
지고아카로마이세스 룩시이에서 개시된 바와 같이(J.M.INGRAM and W.A.WOOD, 1965, Journal of Bacteriology, Vol.89, N°5, 1186-1194), 피치아 오메리에서의 탄소 흐름은, D-글루코오스가 D-리불로오스-5-P로 전환되면서, 동시에 2 분자의 NADPH가 생성되는 5탄당-포스페이트 경로(PPP)의 산화 부분을 통해 진행된다. D-리불로오스-5-P는 D-리불로오스로 탈인산화된 후, D-아라비톨로 환원된다. 피치아 오메리 숙주 균주에서, 5탄당 포스페이트 경로(PPP)는 매우 활성이며, 50%를 초과하는 것으로 측정되었다.
바람직한 실시형태에서, 숙주 세포는 기탁번호 I-4605로, 2012년 3월 7일자로, 인스티튜트 파스퇴르(Institut Pasteur)의 내셔널 콜렉션 오브 마이크로오가니즘 컬쳐스(Collection of Microorganism Cultures, CNCM(Collection Nationale de Cultures de Microorganismes; 25 rue du Docteur Roux, 75724 PARIS Cedex 15))에 기탁된 돌연변이 피치아 오메리이다.
산화 환원 반응 및 효소
보인자 NADH 및 NADPH는 이른바 산화 환원 반응으로 작용하는 다수의 생물학적 기능에 필수적이며, 하나의 반응에서 다른 반응으로의 전자 운반체로서, 작용한다. NADH/NADP+ 쌍이 NADH/NAD+ 쌍보다 더 환원된 상태로 유지되는 것을 볼 때, 세포는 열역학 추진력의 제공을 위해, 두 산화 환원 쌍 NADH/NAD+ 및 NADPH/NADP+의 대사 평형을 유지할 필요가 있다. NADH는 산화된 형태 NAD+로 주로 발견되며, 영양소로부터 에너지의 산화 방출에 관여하는 이화 반응의 주된 보인자이다. NADH와 달리, NADPH는 단백 동화 반응시 또는 산화 스트레스 기간 동안만, 재산화된다
산화 환원 반응을 수반하는 모든 대사 공학 전략은 이러한 세포의 제약 하에서 기능해야 한다. 본 발명의 목적인 유전자 변형 균주에서 수행되었다.
본 발명자에 의해 발견된 바와 같이(박사 논문("Contribution
du metabolisme des pentitols chez Pichia ohmeri"(Sophie Huchette, University of Sciences and Technics of Lille, 1992))에 명백히 기술됨), 케토5탄당의 산화 환원에 관여되는 반응이 두 개의 상이한 효소들에 의해 촉매된다는 것이 입증된 바 있다.
따라서, 숙주 균주는, D-리불로오스로부터 D-아라비톨을 생성하고, D-자일룰로오스로부터 자일리톨을 생성하는 NADPH-특이적 D-케토5탄당-산화환원효소로서 정의된 효소를 가진다. 숙주 균주는 또한, D-리불로오스 및 D-자일룰로오스로오스로부터 각각 리비톨 및 자일리톨을 생성하는 NADH-특이적 D-케토5탄당-산화환원효소를 가진다. 이러한 효소는 잘 알려진 피치아 스티피티스(XYL2)의 NAD+-특이적 자일리톨 탈수소효소 E.C 1.1.1.9와 유사하다. D-자일룰로오스와 달리, 세포 내 D-리불로오스만이 이용 가능하므로, 숙주 균주는 D-리불로오스로부터 D-아라비톨의 세포질 생성을 통해, NADPH/NADP+ 산화 환원 쌍을 직접 NADPH의 재산화와 평형화한다. 그 후, D-아라비톨은 수동적 확산을 통해 브로쓰(broth)로 분비된다.
본 발명자들은, 피치아 오메리가 NADH- 또는 NADPH-특이적-D-케토5탄당-산화환원효소를 통해, 이들 폴리올을 생성하는 모든 효소적 도구를 보유하고 있음에도 불구하고, 세포 내 D-자일룰로오스의 결핍이 숙주 균주에 의한 자일리톨의 비생성의 주된 이유가 된다는 것을 밝혀냈다.
사실, 세포질 D-아라비톨의 D-자일룰로오스 및 NADH로의 전환을 가능하게 하는 것으로서, NAD+-특이적 D-아라비톨 4-산화환원효소(D-자일룰로오스-생성) 활성(E.C.1.1.1.11)을 가지는 단백질을 인코딩하는 유전자의 야생형 숙주 균주 피치아 오메리로 클로닝하는 것이 선택되었다.
따라서, 세포 내 D-자일룰로오스는 유전자 변형 균주로 이용 가능하며, 고유의 NADH- 및 NADPH-특이적-D-케토5탄당-산화환원효소에 의해, 환원될 수 있다. 그러나, 이러한 균주에는 D-자일룰로오스를 자일리톨로 효율적으로 변형시킬 수 있는 내인성 효소가 존재하지 않는다. 따라서, 이종 자일리톨 탈수소효소를 도입하기 위해, 이들 균주를 유전적으로 조작할 필요가 있다.
특허 WO 94/10325에서, 자일리톨의 생성을 가능하게 하고, 이전 대사 단계에 의해 생성된 NADH의 산화와 NADH/NAD+ 산화 환원 쌍을 평형화시키는 것으로서, 피치아 스티피티스(XYL2)로부터 NAD+-특이적 자일리톨 탈수소효소(E.C 1.1.1.9)의 클로닝이 선택되었다. 그러나 앞서 언급했듯이, 그 결과는 정말로 설득력이 없다.
본 발명자들은, NADPH-특이적 자일리톨 탈수소효소를 가지는 돌연변이 단백질을 인코딩하는 유전자의 클로닝에 의해, D-자일룰로오스가 자일리톨로 전환되어, D-리불로오스로부터 D-아라비톨의 고유한 생성에 의해, 수행되는 바와 같이, NADPH/NADP+ 산화 환원 쌍이 평형화된다는 것을 밝혀냈다.
피치아 오메리 야생형 숙주 균주는, D-아라비톨에 대한 NADPH-특이적 D-케토5탄당-산화환원효소의 친화도가 낮기 때문에, 세포 외 D-아라비타톨을 소비하지 않는다.
유전자 변형 균주 내로의 NAD+-특이적 D-아라비톨 4-산화환원효소(D-자일룰로오스-생성) 활성의 도입으로 인해, 브로쓰 내로 생성된 D-아라비톨은 세포질 D-아라비톨과 동일한 방식으로, 변형 균주에 의해 잘 소비될 수 있다.
따라서, 자일리톨은 세포 내 및 세포 외 D-아라비톨로부터 동시에 생성된다.
그 생산은, 자일리톨 경로 확장의 효율을 증대시킴으로써, 중간 D-아라비톨의 유출을 완전히 피할 수 있게함으로써, 향상될 수 있다.
따라서, D-아라비톨과 동일한 생리적 효과를 가지는 D-글루코오스로부터, 단지 자일리톨만이 생성될 것이다. 이러한 향상은 유전자 변형의 결과일 수 있으나, 또한 배양 조건의 적응의 결과일 수 있다.
숙주 균주에서
클로닝되는
2가지 효소 활성의 선택
이러한 2가지 효소 활성의 선택은 전술된 바와 같이, 그의 보인자 특이성에 의해 지원된다.
첫 번째 효소는 D-아라비톨을 D-자일룰로오스로 산화시킨다.
2종의 D-아라비톨 탈수소효소가 공지되어 있다: D-자일룰로오스-생성(EC 1.1.1.11)(D-아라비니톨 NAD+ 4-산화환원효소) 및 D-리불로오스-생성(EC 1.1.1.250). 달리 명시되지 않는 한, 본 명세서에서 아라비톨 탈수소효소로 의도 및 지칭되는 것은 D-자일룰로오스-생성 아라비톨 탈수소효소이다. D-리불로오스-생성 탈수소효소는 야생형 효모 및 균류에서 발견된다.
D-자일룰로오스-생성 아라비톨 탈수소효소는 주로 박테리아에서 알려져 있다. 예를 들어, 이들은 장내세균과(Enterobacteriaceae), 특히 이. 콜라이, 클레브시엘라 에어로게네스 ( Klebsiella aerogenes ), 및 에어로박터 에어로게네스(Aerobacter aerogenes) 균주 PRL-R3, 글루코노박터 옥시단스, 및 추가로 또한 피치아 스티피티스에서 정의된 바 있다. 특히, 클레브시엘라 뉴모니애(Klebsiella pneumoniae)(# O52720), 랄스토니아 솔라나세아룸(# P58708), 예르시니아 페스티스(Yersinia pestis)(# P58709), 에어로박터 에어로게네스(#L8BEF0), 이. 콜라이(# K3EX35, I2ZSJ5, W1BYD6, W1H8N7, E7U4R7)와 같은 다수의 효소가 UniprotKB 데이터베이스에서 참조된다.
본 발명의 목적을 위해, 에스케리치아 콜라이(Escherichia coli)는 NAD+-특이적 D-아라비톨 4-산화환원효소(D-자일룰로오스-생성) 유전자의 바람직한 공급원이다. 더욱 구체적으로, 그의 아미노산 서열은 SEQ ID NO 2로 개시된다. 특히, SEQ ID NO 1 및 SEQ ID NO 3은 에스케리치아 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 핵산을 개시한다. 상기 인코딩 서열은 그의 코돈 특이성을 고려하여, 피치아 오메리에 대해 최적화되었다.
또한, 랄스토니아 솔라나세아룸은 또한 NAD+-특이적 D-아라비톨 4-산화환원효소(D-자일룰로오스-생성) 유전자의 바람직한 공급원이다. 더욱 구체적으로, 그의 아미노산 서열은 SEQ ID NO 43로 개시된다. 특히, SEQ ID NO 42는 랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 핵산을 개시한다. 상기 인코딩 서열은 그의 코돈 특이성을 고려하여, 피치아 오메리에 대해 최적화되었다.
두 번째 효소는 D-자일룰로오스를 자일리톨로 전환시킨다.
대부분의 효모와 균류가 내인성 자일리톨 탈수소효소(EC 1.1.1.9) 유전자를 가진다할지라도, NADH에서 NADPH로의 그의 보인자 특이성의 변화는 본 발명의 구현을 위해, 필요하다. 사실, 본 발명의 주요 측면은 NADPH-특이적 자일리톨 탈수소효소의 사용이다. 또한, 이러한 효소는 바람직하게는 숙주에서 과발현된다.
다수의 자일리톨 탈수소효소가 알려져 있으며, 다수의 과학 문헌이 NADH에서 NADPH로의 보인자 특이성의 변화 방법을 교시하고 있다. 문헌[Watanabe et al(J; Biol. Chem., 2005, 280, 10340-10345)]은 변형 보인자 특이성, 특히 삼중 돌연변이(triple mutant)(D207A/I208R/F209S) 및 사중 돌연변이(quadruple mutant)(D207A/I208R/F209S/N211R)를 가지는 피치아 스티피티스의 돌연변이 자일리톨 탈수소효소를 개시한다. 상기 사중 돌연변이의 아미노산 서열은 SEQ ID NO 5로 개시되어 있다. NADPH 보인자 특이성을 가지는 글루코노박터 옥시단스의 자일리톨 탈수소효소의 이중 돌연변이(D38S/M39R)가 문헌[Ehrensberger et al(2006, Structure, 14, 567-575)]에 개시되어 있다. 상기 이중 돌연변이의 아미노산 서열은 SEQ ID NO 8로 개시되어 있다.
본 발명자들은, NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 피치아 스티피티스 XYL2 핵산 서열의 돌연변이 유발 및 클로닝을 제조하였다. 특히, SEQ ID NO 4 및 SEQ ID NO 6에서, 피치아 스티피티스의 특이적 NADPH 자일리톨 탈수소효소를 인코딩하는 핵산이 개시된다.
대안적으로, 본 발명자들은 또한, NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 글루코노박터 옥시단스 핵산 서열의 돌연변이 유발 및 클로닝을 수행하였다. 특히, SEQ ID NO 7 및 SEQ ID NO 9에서, 글루코노박터 옥시단스의 NADPH 특이적 자일리톨 탈수소효소를 인코딩하는 핵산이 개시된다. 상기 인코딩 서열은 그의 코돈 특이성을 고려하여, 피치아 오메리에 대해 최적화되었다.
발현 카세트, 벡터 및 재조합 숙주 세포
특정 측면에서, 본 발명은 SEQ ID NO 1, SEQ ID NO 3, SEQ ID NO 7, SEQ ID NO 9 및 SEQ ID NO 42로 이루어진 그룹으로부터 선택되는 피치아 오메리에 대해 최적화된 인코딩 서열을 포함하는 핵산에 관한 것이다.
본 발명은 또한, SEQ ID NO 1, SEQ ID NO 3, SEQ ID NO 7, SEQ ID NO 9 및 SEQ ID NO 42로 이루어진 그룹으로부터 선택되는 피치아 오메리에 대해 최적화된 인코딩 서열을 포함하는 핵산을 포함하는 발현 카세트에 관한 것이다.
본 발명은 또한, SEQ ID NO 4의 핵산 작제물 및 상기 핵산 작제물을 포함하는 핵산에 관한 것이다.
또한, 본 발명은 상기 핵산 또는 발현 카세트를 포함하는 재조합 벡터, 특히 발현 벡터에 관한 것이다. 일반적으로, 발현 카세트는 단백질로의 유전자 전사 및 번역에 필요한 모든 인자를 포함한다. 특히, 여기에는 프로모터, 임의로 인핸서, 전사 종결자 및 번역 인자가 포함된다. 더욱 특히, NADPH-특이적 자일리톨 탈수소효소의 발현의 조절에 이용되는 프로모터는 강력한 발현을 유도하기 위해, 선택된다. 실제로, 이러한 효소는 바람직하게는 숙주 세포에서 과발현된다. 이러한 프로모터는 당업계에 잘 알려져 있다. 예를 들어, 프로모터는 피치아 오메리 리불로오스 환원 효소 프로모터(poRR) 또는 피치아 오메리 포스포글리세레이트 키나아제(poPGK1)일 수 있다.
본 발명은, NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 핵산 및 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 핵산을 포함하는 재조합 벡터, 특히 발현 벡터에 관한 것이다. 본 발명은 또한, NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 핵산을 포함하는 재조합 벡터, 특히 발현 벡터 및 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 핵산을 포함하는 재조합 벡터, 특히 발현 벡터를 포함하는 키트에 관한 것이다.
바람직하게는, 상기 NAD+-특이적 D-아라비톨 4-산화환원효소 및 NADPH-특이적 자일리톨 탈수소효소는 전술된 효소 중 선택된다. 특히, 상기 NAD+-특이적 D-아라비톨 4-산화환원효소는 SEQ ID NO 2 또는 SEQ ID NO 42의 아미노산 서열 또는 1 내지 3개의 아미노산의 첨가, 치환 또는 결실을 가지는 서열을 포함하거나 또는 그로 이루어진다. 특히, 상기 NADPH-특이적 자일리톨 탈수소효소는 SEQ ID NO 5 또는 SEQ ID NO 8의 아미노산 서열 또는 1 내지 3개의 아미노산의 첨가, 치환 또는 결실을 가지는 서열을 포함하거나 또는 그로 이루어진다.
바람직한 벡터는 플라스미드이다. 적절한 플라스미드는 당업자에게 잘 공지되어 있으며, 예를 들어, 실시예에 구체적으로 개시된 것들 중에서 선택될 수 있다.
본 발명의 유전자 변형 숙주는 우선, 적절합 프로모터의 조절 하에, NAD+-특이적 D-아라비톨 4-산화환원효소 및 NADPH-특이적 자일리톨 탈수소효소를 코딩하는 유전자를 재조합 벡터로 클로닝한 후, 형질전환에 의해, D-아라비톨 생성 유기체의 숙주 세포로 도입시킴으로써, 생성된다.
본 발명은 NAD+-특이적 D-아라비톨 4-산화환원효소(EC 1.1.1.11)를 인코딩하는 이종 핵산 서열 및 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 이종 핵산 서열을 포함하는 재조합 또는 유전자 조작 숙주 세포에 관한 것이다. NAD+-특이적 D-아라비톨 4-산화환원효소는 기질로서, D-아라비톨을 이용하며, 생성물로서, D-자일룰로오스를 생성한다. NADPH-특이적 자일리톨 탈수소효소는 기질로서 D-자일룰로오스를 이용하며, 자일리톨을 생성한다. NADPH-특이적 자일리톨 탈수소효소 및 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 서열은 에피솜일 수 있거나, 또는 숙주 세포의 염색체로 혼입될 수 있다. 실제로, 유전적으로 안정한 형질전환체는 바람직하게는, 벡터를 이용한 형질전환 시스템을 통해 제작되며, 원하는 DNA가 숙주 염색체로 혼입된다. 이러한 혼입은 세포 내에서, 새로 유발되거나 또는 염색체에서, DNA 서열의 혼입을 촉진하는 DNA 인자를 가지는 것으로서, 숙주 염색체로 그 자체가 기능적으로 삽입되는 벡터에 의한 형질전환에 의해, 보조될 수 있다.
재조합 또는 유전자 조작 숙주 세포는, 바람직하게는 숙주 세포 염색체로 혼입된 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 서열의 복수의 카피 및/또는 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 서열의 복수의 카피를 포함할 수 있다. 특히, 재조합 또는 유전자 조작 숙주 세포는 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 2, 3 또는 4개의 서열 및/또는 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 2, 3 또는 4개의 서열을 포함할 수 있다. 예를 들어, 숙주 세포는 이. 콜라이의 2 또는 3개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및/또는 랄스토니아 솔라나세아룸의 1 또는 2개의 NAD+-특이적 D-아라비톨 4-산화환원효소, 더욱 구체적으로는 이. 콜라이의 2 또는 3개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및/또는 랄스토니아 솔라나세아룸의 하나의 NAD+-특이적 D-아라비톨 4-산화환원효소를 포함할 수 있다. NAD+-특이적 D-아라비톨 4-산화환원효소는 동일한 유기체 또는 다른 유기체로부터 유래한 것일 수 있다. NADPH-특이적 자일리톨 탈수소효소는 동일한 유기체 또는 다른 유기체의 것일 수 있다. 예를 들어, 숙주 세포는 피치아 스티피티스의 1, 2 또는 3개의 NADPH-특이적 자일리톨 탈수소효소 및/또는 글루코노박터 옥시단스의 1, 2 또는 3개의 NADPH-특이적 자일리톨 탈수소효소, 더욱 구체적으로는 피치아 스티피티스의 하나의 NADPH-특이적 자일리톨 탈수소효소 및/또는 글루코노박터 옥시단스의 3개의 NADPH-특이적 자일리톨 탈수소효소를 포함할 수 있다.
본 발명의 특정 측면에서, 재조합 또는 유전자 조작 숙주 세포는 하기의 것을 포함하는 피치아 오메리 균주이다:
- 2개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 2개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 이. 콜라이의 2개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 하나는 피치아 스티피티스의 것이고, 다른 하나는 글루코노박터 옥시단스의 것인 2개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 2개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 3개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 이. 콜라이의 2개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 하나는 피치아 스티피티스의 것이며, 2개는 글루코노박터 옥시단스의 것인 3개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 3개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 3개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 2개는 이. 콜라이의 것이고, 나머지 하나는 랄스토니아 솔라나세아룸의 것인 3개의 NAD+-특이적 D-아라비톨 4-산화환원효소, 및 하나는 피치아 스티피티스의 것이고, 2개는 글루코노박터 옥시단스의 것인 3개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 4개의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 4개의 NADPH-특이적 자일리톨 탈수소효소; 또는
- 3개는 이. 콜라이의 것이고, 나머지 하나는 랄스토니아 솔라나세아룸의 것인 4개의 NAD+-특이적 D-아라비톨 4-산화환원효소, 하나는 피치아 스티피티스의 것이고, 3개는 글루코노박터 옥시단스의 것인 4개의 NADPH-특이적 자일리톨 탈수소효소.
숙주 세포는 전술된 유기체 중 선택된다. 바람직한 실시형태에서, 숙주 세포는 피치아 오메리이다. 출발 숙주 세포는 바람직하게는, 기탁번호 I-4605 하에, CNCM에 기탁된 돌연변이 피치아 오메리이다.
본 발명의 특정 측면에서, 숙주 세포는 CNCM에 기탁된 균주 I-4982, I-4960 및 I-4981로부터 선택되는 균주이다.
본 발명은, 배양 배지에서, 재조합 또는 유전자 조작 숙주 세포를 배양하는 단계 및 생성된 자일리톨을 회수하는 단계를 포함하는 자일리톨의 생성 방법에 관한 것이다. 바람직하게는, 배양 배지는 미생물에 간편한 탄소원을 제공한다. 탄소원에는 바람직하게는 D-글루코오스, 및 다양한 D-글루코오스-함유 시럽 및 D-글루코오스와 당류의 혼합물이 포함된다. 상기 방법은 자일리톨의 정제 단계를 추가로 포함할 수 있다.
본 발명은 본 명세서에 개시된 바와 같이, 재조합 또는 유전자 조작 숙주 세포의 자일리톨의 생성을 위한 용도에 관한 것이다.
이러한 유전자 변형 균주에 의해 생성된 자일리톨은, 당업계에 공지된 임의의 기술에 따라, 본 발명의 숙주의 배지로부터 정제될 수 있다. 예를 들어, 본 명세서에서, 참조로서 포함되는 US 5,081,026은 효모 배양액으로부터 자일리톨을 크로마토그래피에 의해 분리하는 방법을 개시한다. 따라서, US 5,081,026에 개시된 바와 같이, 크로마토그래피 단계 후, 결정화 단계를 이용하여, 발효 단계의 자일리톨을 배양 배지로부터 정제할 수 있다.
하기 실시예를 읽어본다면, 본 발명의 기타 특징적 특성 및 이점이 분명해질 것이다. 그러나, 본 명세서에서, 이들은 단지 예시로서 제공되는 것이며, 한정하고자 하는 것은 아니다.
실시예
실시예
1. 유전자 조작을 위한 바람직한 숙주로서, 피치아 오메리 균주의 선택
선택 숙주 균주로서, 피치아 오메리는:
- 고삼투압 배지, 예를 들어, 10-60% D-글루코오스, 바람직하게는 25% D-글루코오스를 포함하는 배지("정상" 배지는 일반적으로 단지 2-3% 글루코오스를 포함함) 하에, 글루코오스로부터 다량의 아라비톨의 생산자이며;
- 필요한 보인자의 생성을 가능하게 하는 산화 환원 평형을 가진다.
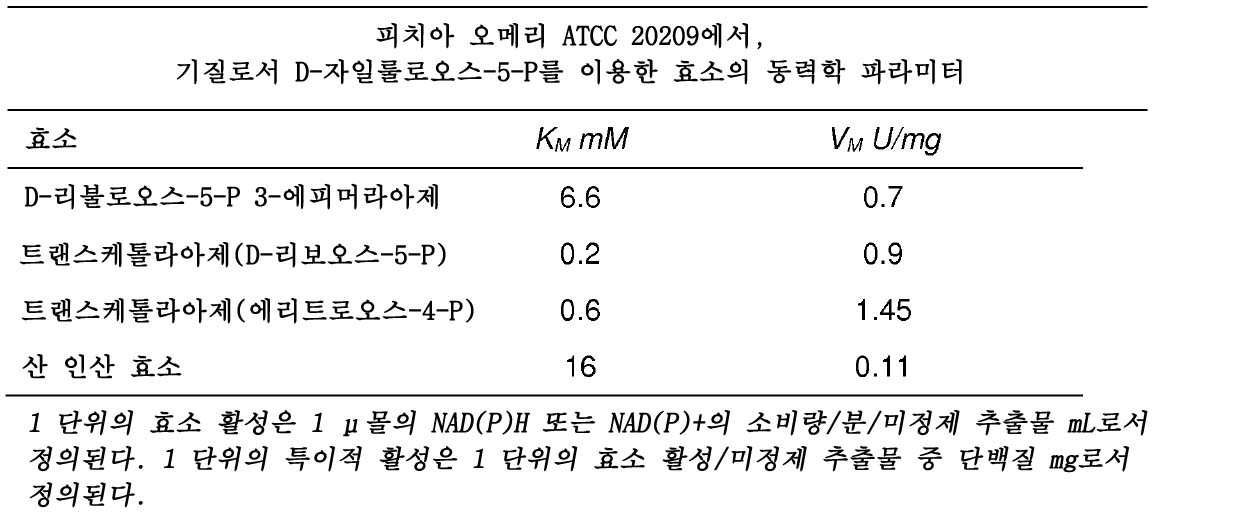
그의 성능의 예시로서, 하기 표는 피치아 오메리의 아라비톨 대사 경로에 관여하는 효소 활성을 나타낸다(Sophie HUCHETTE의 논문, 1992).
6탄당 모노포스페이트 경로: 글루코오스-6-P 내지 D-리불로오스-5-P 및 D-자일룰로오스-5-P
PPP의 산화 부분, 또한 이른바 6탄당 모노포스페이트 경로(HMP)는 NADPH-생성 경로이다. 2개의 NADP+-의존적 효소, 글루코오스-6-P 탈수소효소(E.C.1.1.1.49) 및 6-P-글루코네이트 탈수소효소(E.C.1.1.1.44)는 1 몰의 D-리불로오스-5-P에서, 1 몰의 글루코오스-6-P의 산화 과정에 관여하며, 2 몰의 NADPH를 생성시킨다.
하기 효소의 동력학 파라미터를 측정하였다: D-리불로오스-5-P 3-에피머라아제(E.C 5.1.3.1), D-리보오스-5-P 케토-이성질화 효소(E.C.5.3.1.6), 트랜스케톨라아제(E.C.2.2.1.1) 및 산성 인산 효소(E.C. 3.1.3.2).
생체 내에서, D-리불로오스-5-P의 에피머화로부터 합성된 D-자일룰로오스-5-P는 케토 전달 과정을 통해, PPP의 비산화 부분으로 효과적으로 진입한다. 결과적으로, D-자일룰로오스-5-P는 D-자일룰로오스로의 그의 탈인산화에 이용 가능하지 않다.
NADH
및
NADPH
특이적 D-
케토5탄당
-산화환원효소
D-리불로오스-5-P 및 D-자일룰로오스-5-P의 탈인산화에 의해, D-리불로오스 및 D-자일룰로오스가 생성된다.
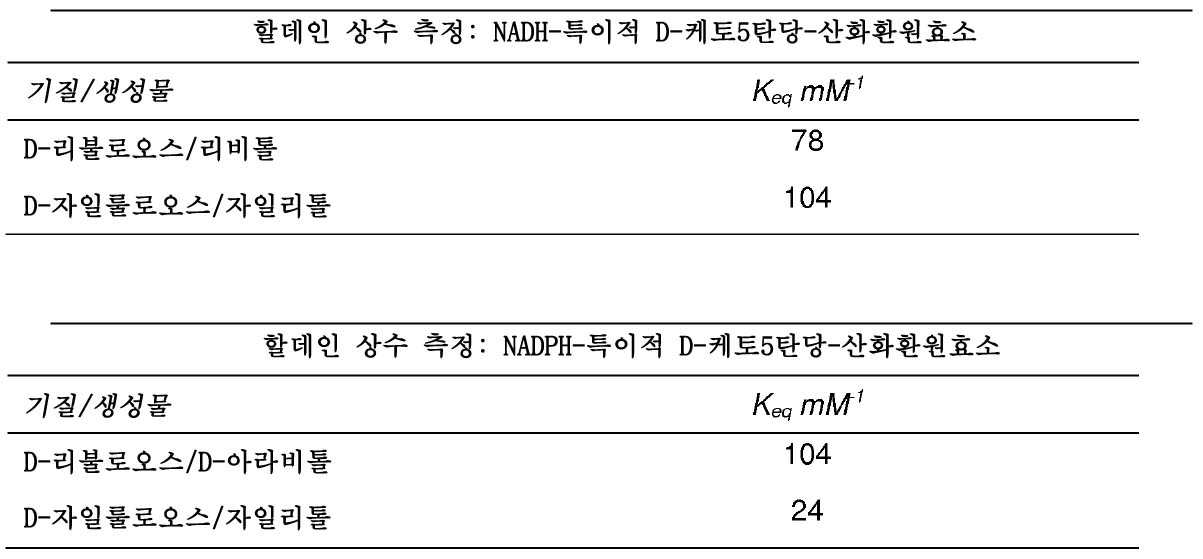
미카엘리스-멘텐 상수는 각각의 기질에 대한 NADH 및 NADPH-D-케토5탄당-산화환원효소의 친화성 및 상응하는 최고 속도를 강조한다.
각각 D-리불로오스 및 D-자일룰로오스로부터 리비톨 및 자일리톨을 생성하는 NADH-특이적 D-케토5탄당-산화환원효소는 D-리불로오스 보다 D-자일룰로오스에 대해 더 큰 친화성을 나타낸다. 역반응은 자일리톨 및 리비톨에 대해 양호한 친화성을 나타내며, 이는 이들 2개의 폴리올에서의 숙주 균주의 양호한 성장을 설명해준다.
D-리불로오스로부터 D-아라비톨을 생성하고, D-자일룰로오스로부터 자일리톨을 생성하는 NADPH-특이적 D-케토5탄당-산화환원효소는 D-자일룰로오스 보다 D-리불로오스에 대해 더 큰 친화성을 나타낸다. 역반응은 D-아라비톨에 대해 매우 작은 친화성을 나타내며, 이는 이들 폴리올에서의 숙주 균주의 비성장을 설명해준다.
숙주 균주의 2개의 케토5탄당-산화환원효소를 특성화하였으며, 문헌[Saccharomyces rouxii by Ingram and Wood, 1965(Journal of Bacteriology, vol.89, n°5, 1186-1194)]에 개시된 종래의 효소와 달랐다. 사실, 사카로마이세스 룩시이(Saccharomyces rouxii)에서, D-리불로오스 및 NADH에서는, 정반응이 검출되지 않았으며, NADPH와 D-아라비톨에서는 역반응이 검출되었다.
할데인(Haldane) 관계에 의해, 생체 내 효소 동력학 거동이 예측된다.
2가지 효소에서, 역반응(펜티톨 환원)에 비해, 정반응(D-케토5탄당 산화)이 우세하다.
숙주 균주의 PPP는 매우 효과적이며, 소비된 1 몰의 글루코오스로부터 2 몰의 NADPH가 생성된다. 결과적으로, NADPH는 근육 강화 반응과 유지 반응 모두에서 과량으로 이용 가능할 것이다. 숙주 균주는 D-리불로오스로부터 D-아라비톨을 생성하거나 또는 D-자일룰로오스로부터 자일리톨을 생성하여, NADPH/NADP+ 산화 환원 쌍의 평형이 이루어져야 한다.
NADPH-특이적 D-케토5탄당-산화환원효소에 대한 NADP+의 억제 효과를 시험관 내에서 측정하였다. NADP+가 과량으로 첨가되는 경우, 활성은 80%이다. 이러한 농도가 세포 내 NADP+ 농도와 상용성이 아니라 할지라도, 이러한 결과는, NADPH/NADP+ 산화 환원 쌍의 평형에 대한 NADPH-특이적 D-케토5탄당 산화환원효소의 역할에 대한 부분적 개괄을 제공한다.
PPP의 비산화 부분으로의 D-자일룰로오스-5-P의 유입으로 인해, D-자일룰로오스가 이용될 수 없었기 때문에, 숙주 균주는 D-리불로오스로부터 D-아라비톨만을 생성한다.
D-아라비톨 생성에 대한 글루코오스-6-P 탈수소효소의 과발현의 영향을 평가함으로써, 숙주 균주에서, D-아라비톨의 생성과 NADPH/NADP+ 산화 환원 평형 사이의 관계를 입증하였다. 따라서, 획득된 균주는 1.5배 더 큰 G6PDH 활성을 가지며, 숙주 균주(FR2772788)에 비해, 10% 이상의 D-아라비톨을 생성한다.
실시예
2.
피치아
오메리
코돈 선호도(
codon
usage
)
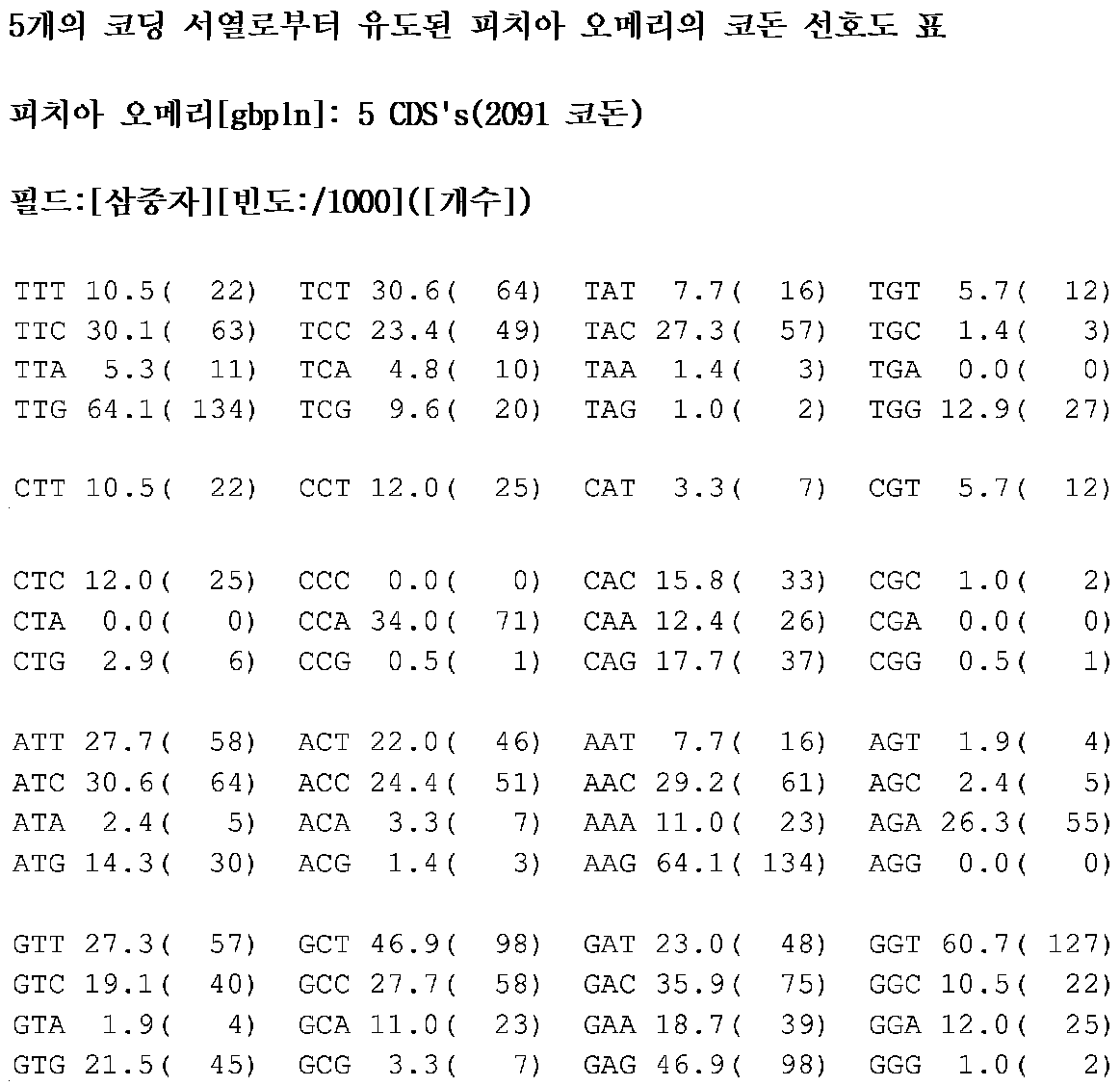
5개의 피치아 오메리 유전자의 상응하는 아미노산 서열 및 이용 가능한 DNA로부터 피치아 오메리의 코돈 선호도를 측정하였다: 트랜스케톨라아제, 글루코오스-6-포스페이트 탈수소효소(FR 2772788), 리불로오스 환원 효소, 베타-이소프로필말레이트 탈수소효소 - LEU2(Piredda and Gaillardin, Yeast, vol.10:1601-1612(1994) 및 오로티딘-5'-포스페이트 탈카르복실화효소 - URA3(Piredda and Gaillardin, 1994, 상기).
모든 개별 유전자를 단일 아미노산을 인코딩하는 뉴클레오티드 삼중자로 분류하였다. 상기 5개의 유전자는 총 2091개의 코돈으로 이루어져 있다.
각 아미노산에서, 5개의 유전자에 존재하는 모든 코돈의 수를 계수한 후, 2091로 나눈 다음, 1000을 곱하였다. 이러한 방식으로, 1000개의 코돈에서, 특이적 코돈의 빈도를 예측하였다.
피치아 오메리의 예비 코돈 선호도를 표 7에 도시하였다.
피치아 스티피티스의 자일리톨 탈수소효소를 제외하고, 피치아 오메리에서 발현되는 모든 이종 유전자를 http://genomes.urv.es/OPTIMIZER/에서 얻은 Optimizer 프로그램 및 하기 표를 이용하여, 코돈 최적화하였다.
상기 효소를 인코딩하는 서열의 각각의 5' 및 3' 말단에, 제한 효소의 인식 부위를 수작업으로 첨가한 후, 얻어진 서열을 유전자 합성 과정에 제공하였다.
실시예
3. 이. 콜라이 박테리아
NAD
+
-특이적 D-
아라비톨
4-산화환원효소(D-
자일룰로오스
-생성) 유전자의
클로닝
이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 altD를 인코딩하는 DNA 단편을 SEQ ID NO: 1의 제시 서열에 따라, 화학적으로 합성하였다(GeneArt® Gene Synthesis, Life Technologies, Regensburg, Germany).
altD 유전자를 코딩하는 서열 AF378082.1(http://www.ncbi.nlm.nih.gov/nuccore/AF378082에서 입수함)의 뉴클레오티드 1441 내지 2808을 주형으로 사용하였으며, http://genomes. urv.es / OPTIMIZER /에서 입수한 Optimizer 프로그램을 이용하여, 실시예 2의 표 7에 따라, 피치아 오메리 ATCC 20209에서 사용하기 위한 코돈 최적화에 적용하였다.
추가적 클로닝을 용이하게 하기 위해, 생성된 서열의 5' 및 3' 말단에, 각각 제한 효소 AscI(GGCGCGCC) 및 SphI(GCATGC)의 인식 부위를 인코딩하는 뉴클레오티드를 첨가하였다.
또한, 효모의 Kozak-형 서열의 -3 위치에 아데노신을 추가하기 위해, 아데노신 삼중자를 개시 ATG 앞에 포함시켰다.
그 후, 합성을 위해 최종 서열(SEQ ID NO: 1)을 제출하였다(GeneArt, Regensburg, Germany).
이. 콜라이로부터의 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 합성된 DNA 단편을 pMK-RQ 유도 벡터(12ABYWMP, 도 1)에 5 ㎍ 동결건조된 플라스미드 DNA로서, 전달하였다.
추가적 서브클로닝을 위해, AscI 및 SphI 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, 상기 유전자를 방출시켰다.
실시예
4. 피치아 스티피티스
NADH
및
NADPH
-특이적
자일리톨
탈수소효소의 돌연변이 유발 및
클로닝
피치아 스티피티스 NADH -특이적 자일리톨 탈수소효소 유전자의 클로닝
자일리톨 탈수소효소를 인코딩하는 효모(피치아 스티피티스) 유전자 XYL2의 인지의 뉴클레오티드 서열(Koter et al., Curr. Genet. 18:493-500(1990))을 FR 2 765 589(해당 특허의 실시예 4 및 도 7 참조)에 따라, 플라스미드 벡터 lig 7.78에 클로닝하였다. 상기 벡터의 제한효소 지도는 도 2a에서 제공된다.
피치아 스티피티스 NADPH -특이적 자일리톨 탈수소효소 유전자의 돌연변이 및 클로닝
피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 XYL2를 인코딩하는 DNA 단편을 SEQ ID NO: 4의 서열에 따라, 화학적으로 합성하였다(GeneArt® Gene Synthesis, Life Technologies, Regensburg, Germany).
XYL2 유전자를 코딩하는 서열 X55392.1(http://www.ncbi.nlm.nih.gov/nuccore/X55392.1에서 얻음)의 뉴클레오티드 319 내지 1410을 주형으로 사용하였다.
Watanabe 등의 논문(J; Biol. Chem., 2005, 280, 10340-10345)에 따르면, 자일리톨 탈수소효소의 보인자 선호는 4개의 공개된 아미노산 돌연변이의 도입에 의해, NADH에서 NADPH로 변화될 수 있다: D207A/I208R/F209S/N211R(http://www.uniprot.org/uniprot/P22144에서 얻은 P22144 단백질 서열을 기초로 번호 매김).
따라서, D207, I208, F209 및 N211을 인코딩하는 코돈을 각각 해당 서열에서, GCT, AGA, TCA 및 AGA로 수작업 교체 하였다.
또한, 추가적 클로닝을 용이하게 하기 위해, 각각 제한 효소 HindIII(AAGCTT) 및 SacII(CCGCGG)의 인식 부위를 코딩하는 뉴클레오티드를 각각 5' 및 3' 말단에, 수작업 첨가하였다.
또한, 효모의 Kozak-형 서열의 -3 위치에 아데노신을 추가하기 위해, 아데노신 삼중자를 개시 ATG 앞에 포함시켰다. 최종 서열(SEQ ID NO: 4)을 합성을 위해, 제공하였다(GeneArt, Regensburg, Germany).
피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 합성 DNA 단편을 pMA-T 유도 벡터(12AALQTP, 도 2b)에 5 ㎍ 동결건조 플라스미드 DNA로서, 전달하였다.
실시예
5. 글루코노박터 옥시단스
NADPH
-특이적
자일리톨
탈수소효소 유전자의 돌연변이 유발 및 클로닝
글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 Xdh를 인코딩하는 DNA 단편을 SEQ ID NO: 7의 서열에 따라, 화학적으로 합성하였다(GeneArt® Gene Synthesis, Life Technologies, Regensburg, Germany).
Xdh 유전자를 코딩하는 서열 AB091690.1(http://www.ncbi.nlm.nih.gov/nuccore/AB091690.1에서 얻음)의 뉴클레오티드 1063 내지 1851을 주형으로 사용하였으며, http://genomes.urv.es/OPTIMIZER/에서 얻은 Optimizer 프로그램을 이용하여, 표 7(실시예 2)에 따라, 피치아 오메리 ATCC 20209에서 사용하기 위한 코돈 최적화에 적용하였다.
Ehrensberger 등의 문헌(Structure, 2006, 14, 567-575)에 따르면, 상기 효소의 보인자 특이성은 2개의 공개된 아미노산 돌연변이의 도입에 의해, NADH에서 NADPH로 변화될 수 있다: D38S/M39R(http://www. uniprot.org / uniprot / Q8GR61)에서 얻은 Q8GR61 단백질 서열을 기초로 번호 매김).
따라서, D38 및 M39를 인코딩하는 코돈을 해당 서열에서, 각각 TCT 및 AGA로 수작업으로 교체 하였다. 또한, 추가적 클로닝을 용이하게 하기 위해, 각각 제한 효소 AscI(GGCGCGCC) 및 SphI(GCATGC)의 인식 부위를 코딩하는 뉴클레오티드를 각각 5' 및 3' 말단에, 수작업으로 첨가하였다.
또한, 효모의 Kozak-형 서열의 -3 위치에 아데노신을 추가하기 위해, 아데노신 삼중자를 개시 ATG 앞에 포함시켰다. 최종 서열(SEQ ID NO: 7)을 합성을 위해, 제공하였다(GeneArt, Regensburg, Germany).
글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 합성 DNA 단편을 pMA-T 유도 벡터(13AAYSYP, 도 3)에 5 ㎍ 동결건조 플라스미드 DNA로서, 전달하였다. 추가적 서브클로닝을 위해, AscI 및 SphI 효소를 이용한 제한 효소 절단에 의해, 상기 유전자를 방출시켰다(New England Biolabs, Ipswich, Massachusetts).
실시예
6.
po
URA3
선택
마커를
이용한 이종 유전자 발현을 위한 피치아 오메리 벡터의 제작
교체 가능한:
- 프로모터,
- 오픈 리딩 프레임, 및
- 종결자 인자
를 가지는 벡터의 클로닝을 3개의 개별 단편의 2회의 연속 중첩 PCR에 의해 수행하였다(도 4).
재조합 피치아 오메리 균주에서, 타가토스 3-에피머라아제 유전자의 클로닝 및 과발현을 시험하기 위한 발현 모델로서, 상기 벡터를 처음에 계획하였다.
하기에서 개시되는 바와 같이, 타가토스 3-에피머라아제 유전자를 특이적 AscI - SphI 제한효소 부위 카세트에 클로닝하여, 이러한 동일한 삽입 부위를 이용한 목적 유전자의 클로닝을 가능하게 하였다.
하기의 방식에 의한 클로닝이 고려되었다.
첫 번째 PCR(PCR1)에서, SpeI 및 AscI 부위(프라이머 서열 내 밑줄 부분)에 플랭킹된 피치아 오메리의 490 bp 길이의 리불로오스 환원 효소 프로모터 단편을 하기의 것을 이용하여 증폭시켰다:
- 프라이머 EV2960:
및
- 프라이머 EV2961:
또한, 역방향 프라이머 EV2961의 5' 말단에, 타가토스-3-에피머라아제 유전자의 5' 말단을 나타내는 13개의 뉴클레오티드 길이의 단편을 첨가하였다.
AscI 부위의 8개의 뉴클레오티드 및 리불로오스 환원 효소 프로모터의 3' 말단의 10개의 이후 뉴클레오티드를 함께 가지는 이들 단편이 하기 개시되는 PCR2의 단편과 PCR1의 단편의 융합을 위한 중첩으로 필요하였다. 피치아 오메리 ATCC 20209의 게놈 DNA를 주형으로서 사용하였다.
이러한 목적을 위해, 새로 스트리킹(streaking)된 피치아 오메리 콜로니를 30 ㎕의 0.2% SDS 중에 재현탁시킨 후, 4분 동안 95℃에서 가열하였다. 전속 원심분리 후, 0.5 ㎕의 상청액을 PCR에 사용하였다.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/50℃에서 20초/72℃에서 15초의 25회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
제2 PCR(PCR2)에서, AscI 및 SphI 부위(프라이머 서열 내 밑줄 부분)에 플랭킹된 슈도모나스 치코리이 ST24의 타가토스-3-에피머라아제의 911 bp 길이의 단편을 하기의 것을 이용하여, 증폭시켰다:
- 프라이머 EV2962:
및
- 프라이머 EV2963:
프라이머 EV2962의 5' 말단은 리불로오스 환원 효소 프로모터의 3'을 나타내는 9개의 뉴클레오티드 길이의 단편을 포함한다.
AscI 부위의 8개의 뉴클레오티드 이후, 타가토스-3-에피머라아제 오픈 리딩 프레임의 12개의 뉴클레오티드를 함께 가지는 이들 단편을 전술된 PCR1 생성물과 PCR2 생성물을 융합하기 위한 중첩 PCR에 사용한다.
또한, 역방향 프라이머 EV2963의 5' 말단은 피치아 오메리의 리불로오스 환원 효소 종결자의 5' 말단을 나타내는 12개의 뉴클레오티드 길이의 단편을 포함한다.
SphI 부위의 6개의 뉴클레오티드 이후, 타가토스-3-에피머라아제 오픈 리딩 프레임의 3' 말단의 12개의 뉴클레오티드를 함께 가지는 이들 단편이 전술된 PCR3의 PCR 단편과 PCR2의 융합을 위해, 중첩으로 필요하다.
주형으로서, 슈도모나스 치코리이 ST24의 타가토스-3-에피머라아제 유전자의 합성 카피를 포함하는 25 ng의 벡터 12AAMCJP(도 5)(GeneArt, Regensburg, Germany)를 사용하였다(http://www.ncbi.nlm.nih.gov/nuccore/AB000361의 AB000361.1의 뉴클레오티드 719 내지 1591) - SEQ ID NO: 11.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/48℃에서 20초/72℃에서 30초의 25회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다.
제3 PCR(PCR3)에서, SphI 및 SacII 부위(프라이머 서열 내 밑줄 부분)에 플랭킹된 피치아 오메리의 리불로오스 환원 효소 종결자의 380 bp 길이의 단편을 하기의 것을 이용하여, 증폭시켰다:
- 프라이머 EV2964
및
- 프라이머 EV2965
프라이머 EV2964의 5' 말단은 타가토스-3-에피머라아제 오픈 리딩 프레임의 3' 말단의 12 뉴클레오티드 길이의 단편을 포함하며, SphI 부위의 6개의 뉴클레오티드 이후, 피치아 오메리의 리불로오스 환원 효소 종결자의 12개의 뉴클레오티드와 함께, 전술된 PCR2와 PCR3의 융합에 사용된다.
피치아 오메리 ATCC 20209의 게놈 DNA를 주형으로 사용하였다. 전속 원심분리 후, 0.5 ㎕의 상청액을 PCR에 사용하였다. 이러한 목적을 위해, 새로 스트리킹된 피치아 오메리 콜로니를 30 ㎕의 0.2% SDS 중에 재현탁시킨 후, 4분 동안 95℃에서 가열하였다.
적절한 1X 완충액에, 200 μM의 각 dNTP, 0.5 μM의 각 프라이머 및 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/50℃에서 20초/72℃에서 15초의 25회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
3개의 개별 PCR 단편의 융합을 하기와 같이 수행하였다: PCR1 및 PCR2의 각 겔 정제 생성물 50 ng을 EV2960 및 EV2963을 이용한 PCR 반응의 주형으로서, 사용하였다.
전술된 프라이머 설계로부터 생성된 2개의 단편의 30개의 뉴클레오티드 길이의 상동 절편을 융합 반응에서, 중첩으로 사용하였다.
이러한 방식으로, SpeI 및 AscI 부위에 플랭킹된 피치아 오메리의 리불로오스 환원 효소 프로모터로 이루어진 1.4 kb 길이의 단편을 슈도모나스 치코리이 ST24의 타가토스-3-에피머라아제의 오픈 리딩 프레임에 융합시켰다.
적절한 1X 완충액에, 200 μM의 각 dNTP, 0.5 μM의 각 프라이머 및 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/62℃에서 20초/72℃에서 45초의 30회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
정제된 단편을 제2 중첩 PCR에서, PCR3 생성물과 융합시켰다: 각 단편 40 ng을 주형으로 사용하고, EV2960 및 EV2965를 사용하여, 증폭시켰다.
전술된 프라이머 설계로부터 생성된 2개의 단편의 30개의 뉴클레오티드 길이의 상동 절편을 융합 반응에서, 중첩으로 사용하였다.
이러한 방식으로, SpeI 및 AscI 부위에 플랭킹된 피치아 오메리의 리불로오스 환원 효소 프로모터 및 AscI 및 SphI 부위에 플랭킹된 슈도모나스 치코리이 ST24의 타가토스-3-에피머라아제의 오픈 리딩 프레임으로 이루어진 1.8 kb 길이의 단편을 피치아 오메리의 리불로오스 환원 효소 종결자에 융합시켰다.
적절한 1X 완충액에, 200 μM의 각 dNTP, 0.5 μM의 각 프라이머 및 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/65℃에서 20초/72℃에서 55초의 30회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 슈도모나스 치코리이 ST24의 타가토스-3-에피머라아제의 1.7 kb 길이의 단편으로 이루어진 최종 PCR 생성물을 제한 효소 SpeI 및 SacII(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 분해시킨 후, 겔 정제한 다음, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, lig 7.78 벡터 백본의 9.8 kb 길이의 분리 SpeI/SacII 단편에 의해, 16℃에서 밤새 결찰시켰다(도 6).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(ultracompetent cell)(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리하였다. 정제된 플라스미드 DNA를 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가적 특성화에 사용하였다.
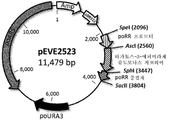
새로 클로닝된 발현 플라스미드 pEVE2523(도 7)는, 효모에서의 선택을 위한 것으로서, 박테리아(이. 콜라이) 복제기점 및 암피실린 내성 유전자, 효모(피치아 오메리) 자가 복제 서열, 및 poURA3(피치아 오메리) 유전자로 이루어진 셔틀 이. 콜라이 - 피치아 오메리 벡터이다.
또한, 여기에는, 슈도모나스 치코리이의 타가토스-3-에피머라아제의 오픈 리딩 프레임에 플랭킹된 대체성 피치아 오메리 리불로오스 환원 효소 프로모터 인자(SpeI 및 AscI 제한을 통해) 및 종결자 인자(SphI 및 SacII를 통해)(AscI 및 SphI 제한을 통해 대체)를 포함한다.
실시예
7.
po
LEU2
선택
마커를
이용한 이종 유전자 발현을 위한 피치아 오메리 벡터의 제작
제2 피치아 오메리 발현 벡터의 제작을 위해, 실시예 6에서 전술된 플라스미드 pEVE2523의 발현 카세트(도 7)를 피치아 오메리 poLEU2 선택 마커를 포함하는 벡터(도 6)로 클로닝하였다.
SpeI 및 SacII(New England Biolabs, Ipswich, Massachusetts)에 의해 절단된 벡터 pEVE2523(도 7)의 1.7 kb 평활 단편을 삽입물로서 사용하였다. 실온에서, 15분 동안 평활화 효소 믹스(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화(Blunting)를 수행한 후, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다.
SalI(New England Biolabs, Ipswich, Massachusetts)에 의해 선형화된 poARS 벡터(plig3 - FR 2772788)로부터 벡터 백본을 얻은 후, 평활화시키고, 앤타크틱(Antarctic) 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서, 1시간 동안, 탈인산화시켰다. Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용한 벡터 백본과 겔 정제 삽입물을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 1시간 동안, 결찰시켰다.
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가적 특성화에 사용하였다.
새로 클로닝된 발현 플라스미드 pEVE2560(도 8)은, 효모에서의 선택을 위한 것으로서, 박테리아(이. 콜라이) 복제기점 및 암피실린 내성 유전자, 효모(피치아 오메리) 자가 복제 서열, 및 poLEU2(피치아 오메리) 유전자를 포함하는 셔틀 이. 콜라이 - 피치아 오메리 벡터이다.
또한, 슈도모나스 치코리이의 타가토스-3-에피머라아제의 오픈 리딩 프레임에 플랭킹된 대체성 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자는 AscI 및 SphI 제한을 통해, 대체 가능하다.
실시예
8. 글루코노박터 옥시단스
NADPH
-특이적
자일리톨
탈수소효소의 과발현을 위한 피치아 오메리 벡터의 제작
글루코노박터 옥시단스 NADPH-특이적 자일리톨 탈수소효소의 과발현을 위한 피치아 오메리 벡터를 제작하였다.
발현 벡터로의 클로닝을 위해, 글루코노박터 옥시단스 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 DNA 단편을 AscI 및 SphI 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 절단하여, 벡터 13AAYSYP(도 3)로부터 방출시켰다.
803 bp 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 2시간 동안, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, pEVE2523(도 7)의 9.8 kb AscI/SphI-분해 겔-정제된 벡터 백본에 결찰시켰다(도 9).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
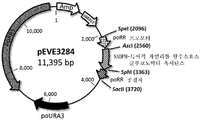
생성된 플라스미드 pEVE3284(도 10)은 피치아 오메리의 리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 글루코노박터 옥시단스의 코돈-최적화 NADPH-특이적 자일리톨 탈수소효소 및 poURA3 선택 마커를 포함한다.
실시예
9. 피치아 스티피티스
NADPH
-특이적
자일리톨
탈수소효소의 과발현을 위한 피치아 오메리 벡터의 제작
발현 벡터로의 서브클로닝을 위해, 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 DNA 단편을 AscI 및 SphI 제한효소 부위에 플랭킹시켜야 한다.
이러한 목적을 위해, 주형으로서, 30 ng의 벡터 12AALQTP(도 2b)와 함께, 하기의 것을 PCR 반응에 사용하였다:
- EV3101 프라이머
(SEQ ID NO 18)(
AscI 부위(밑줄) 포함) 및
- EV3102 프라이머
(SEQ ID NO 19)(
SphI(밑줄) 포함)™
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/55℃에서 20초/72℃에서 30초의 25회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다.
1.1 kb PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제한 다음, AscI 및 SphI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해하였다. DNA Clean & Concentrator™-5 키트(Zymo Research Corporation, Irvine, California)에 의한 칼럼 정제 후, 이를 실온에서, 2시간 동안, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 각각 pEVE2523(도 7)의 10.6 kb AscI/SphI-분해 겔-정제된 벡터 백본 및 pEVE2560(도 8)의 11.8 kb AscI/SphI-분해 겔-정제된 벡터 백본에 결찰시켰다(도 11).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2562 및 pEVE2564(도 12)는 피치아 오메리의 리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 피치아 스티피티스의 코돈-최적화 NADPH-특이적 자일리톨 탈수소효소 및 각각 poURA3 또는 poLEU2 선택 마커를 포함한다.
실시예
10. 피치아 스티피티스
NADH
-특이적
자일리톨
탈수소효소의 과발현을 위한 피치아 오메리 벡터의 제작
발현 벡터로의 서브클로닝을 위해, 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 DNA 단편을 AscI 및 SphI 제한효소 부위에 플랭킹시켜야 한다.
이러한 목적을 위해:
- EV3101
(SEQ ID NO 18)(
AscI 부위(밑줄) 포함) 및
- EV3102
(SEQ ID NO 19)(
SphI(밑줄) 포함)
를 주형으로서, 30 ng의 벡터 lig7.78(도 2a)의 PCR 반응에 사용하였다.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다.
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/55℃에서 20초/72℃에서 30초의 25회 순환 및 72℃에서 10분의 최종 확장 단계의 PCR을 수행하였다.
1.1 kb PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제한 다음, AscI 및 SphI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해하였다.
DNA Clean & Concentrator™-5 키트(Zymo Research Corporation, Irvine, California)에 의한 칼럼 정제 후, 이를 실온에서, 2시간 동안, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, pEVE2560(도 8)의 10.5 kb AscI/SphI-분해 겔-정제된 벡터 백본에 결찰시켰다(도 13).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2563(도 14)은 피치아 오메리의 리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 피치아 스티피티스의 코돈-최적화 NADH-특이적 자일리톨 탈수소효소 및 poLEU2 선택 마커를 포함한다.
실시예
11. 이. 콜라이
NAD
+
-특이적 D-
아라비톨
4-산화환원효소의 과발현을 위한 피치아 오메리 벡터의 제작
이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소의 과발현을 위한 피치아 오메리 벡터를 제작하였다.
발현 벡터로의 서브클로닝을 위해, 코돈-최적화 이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소를 인코딩하는 DNA 단편을 AscI 및 SphI 제한효소(New England Biolabs, Ipswich, Massachusetts)에 의해 절단하여, 벡터 12ABYWMP(도 1)로부터 방출시켰다.
상기 1.4 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 2시간 동안, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, pEVE2523(도 7)의 9.8 kb AscI/SphI-분해 겔-정제된 벡터 백본에 결찰시켰다(도 15).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2839(도 16)는 피치아 오메리의 리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 코돈-최적화 이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소 및 poURA3 선택 마커를 포함한다.
피치아 오메리 리불로오스 환원 효소 프로모터에 더하여, 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하에, 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소를 또한 클로닝하였다.
처음에, 리불로오스 환원 효소 프로모터를 poPGK1 프로모터로 교체한 후, 리불로오스 환원 효소 종결자를 poTKL 종결자로 대체함으로써, 2개의 연속 단계로, 클로닝을 수행하였다.
피치아 오메리 poPGK1 프로모터의 611 bp 길이의 단편을 하기의 것을 이용하여, 피치아 오메리의 게놈 DNA로부터 증폭시켰다:
- 프라이머 EV3177
(SEQ ID NO 20)(
SpeI 부위(밑줄) 포함) 및
- 프라이머 EV3178
(SEQ ID NO 21),(
AscI 부위(밑줄) 포함).
새로 스트리킹된 피치아 오메리 콜로니를 30 ㎕의 0.2% SDS 중에 재현탁시킨 후, 4분 동안 95℃에서 가열하하여, 게놈 DNA 주형을 제조하였다. 전속 원심분리 후, 0.5 ㎕의 상청액을 PCR에 사용하였다.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다.
96℃에서 2분의 초기 변성 단계 후, 96℃에서 10초/58℃에서 10초/72℃에서 30초의 25회 순환 및 72℃에서 2분의 최종 확장 단계의 PCR을 수행하였다.
PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
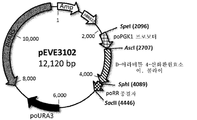
증폭된 610 bp 길이의 poPGK1 프로모터 단편을 AscI 및 SphI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해한 후, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 2시간 동안, pEVE2839(도 16)의 11.5 kb SpeI/AscI-분해 겔-정제된 벡터 백본에 결찰시켰다(도 17).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3102(도 18)는 피치아 오메리의 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 리불로오스 환원 효소 종결자에 플랭킹된 코돈-최적화 이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소 및 poURA3 선택 마커를 포함한다.
다음 단계에서, pEVE3102의 리불로오스 환원 효소 종결자를 피치아 오메리의 트랜스케토라아제(poTKL) 종결자로 교체하였다.
피치아 오메리 poTKL 종결자의 213 bp 길이의 단편을 하기의 것을 이용하여, 피치아 오메리의 게놈 DNA로부터 증폭시켰다:
- 프라이머 EV3817
(SEQ ID NO 22)(
SphI 부위(밑줄) 포함) 및
- 프라이머 EV3818
(SEQ ID NO 23)(
SacII 부위(밑줄) 포함).
게놈 DNA 주형을 전술된 바와 같이 제조하였다.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다.
96℃에서 2분의 초기 변성 단계 후, 96℃에서 10초/57℃에서 10초/72℃에서 30초의 25회 순환 및 72℃에서 2분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
증폭된 213 bp 길이의 poTKL 종결자 단편을 SphI 및 SacII(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해한 후, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 2시간 동안, pEVE3102(도 18)의 11.5 kb SphI/SacII-분해 겔-정제된 벡터 백본에 결찰시켰다(도 17).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3123(도 19)은 피치아 오메리의 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자에 플랭킹된 코돈-최적화 이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소 및 poURA3 선택 마커를 포함한다.
또 다른 선택 과정을 이용하여, 플라스미드로부터 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소가 발현될 수 있도록 하기 위해, pEVE3123의 poURA3 마커를 poLEU2 마커로 교체하였다.
이러한 목적을 위해, PsiI 및 AfeI(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 분해에 의해, poURA3 마커를 벡터 pEVE3123(도 19)로부터 방출시켰다.
상기 9.1 kb 벡터 백본을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안 평활화 효소 믹스(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하고, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서 1시간 동안 탈인산화하였다.
삽입물로서, AseI 및 AfeI 제한 효소 분해에 의해, 벡터 pEVE2560(도 8)로 부터 방출된 poLEU2 마커의 3 kb 평활 겔-정제된 단편을 사용하였다. T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 2시간 동안, 단편의 결찰을 수행하였다(도 20).
상기 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3157(도 21)은 피치아 오메리의 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자에 플랭킹된 코돈-최적화 이. 콜라이 NAD+-특이적 D-아라비톨 4-산화환원효소 및 poLEU2 선택 마커를 포함한다.
실시예
12. 피치아 오메리 균주
ATCC
20209에서, 플라스미드 이.
콜라이
NAD
+
-특이적 D-
아라비톨
4-산화환원효소 및 플라스미드
피치아
스티피티스
NADPH
-특이적
자일리톨
탈수소효소 유전자의 발현
자일리톨로의 아라비톨의 생합성 전환의 경우, NAD+-특이적 이. 콜라이 D-아라비톨 4-산화환원효소 및 피치아 스티피티스의 NADP-특이적 자일리톨 탈수소효소의 동시 발현이 필요하다.
첫 번째 효소는 자일룰로오스의 생성을 초래하고, 두 번째 효소는 자일룰로오스를 자일리톨로 전환시킨다.
ATCC 20209로부터 유도되고 또한 류신 및 우라실에 대해 영양 요구성인 피치아 오메리 균주 SRLU(MATh - leu2 ura3)(Piredda and Gaillardin, 1994, 상기)는, 균주 EYS2755를 초래하는 하기 플라스미드를 이용한 형질전환에 의한 자일리톨 분비 효모 균주의 제작용 숙주로서 이용하였다:
- pEVE2839(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소) 및
- pEVE2564(피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소)
또한, 대조군(WO 94/10325의 교시에 따름)으로서, 균주 EYS2962를 초래하는 것으로서, 하기 플라스미드를 이용한 SRLU 숙주로의 형질전환에 의해, 피치아 스티피티스의 NADH-특이적 야생형 자일리톨 탈수소효소를 발현하는 균주를 제작하였다:
- pEVE2839(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소) 및
- pEVE2563(피치아 스티피티스의 NADH-특이적 자일리톨 탈수소효소)
대조군으로서, 각각 EYS2943, EYS2696 및 EYS2697을 초래하는 것으로, 단일 플라스미드에 의해 형질전환된 균주를 또한 생성시켰다:
- pEVE2839(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소),
- pEVE2563(피치아 스티피티스의 NADH-특이적 자일리톨 탈수소효소), 및
- pEVE2564(피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소)
효모 형질전환은 필수적으로 하기와 같은 수정을 갖는 문헌[Green et al.(Green E.D., Hieter, P., and Spencer F. A., chapter 5 in Genome Analysis: A Laboratory Manual, Vol. 3, Cloning Systems, Birren et al.(eds.)]의 스페로플라스팅 방법(spheroplasting method)에 의해 실시하였다: 스페로플라스트의 생성을 위해, Lyticase 대신에, Zymolyase 100T를 사용하였으며, Zymolyase 처리 전에, 세포 현탁액의 OD가 본래 OD의 20 내지 30%에 도달될 때까지, 37℃에서 효소에 의한 인큐베이션을 수행하였다.
요약하면, 피치아 오메리 세포를 30℃에서 YPD 배지(효모 추출물 1%(w/v), 펩톤 2%(w/v), 덱스트로오스 2%(w/v))에서 밤새 성장시켜서, 최종 OD600이 3 내지 5가 되도록 하였다.
200 OD600 단위를 원심분리에 의해, 수집한 후, 물 및 1M 소르비톨을 이용하여, 1회 세척한 다음, SCE 완충액(1 M 소르비톨, 100 mM 시트르산 삼나트륨 염 2수화물, 10 mM EDTA) 중에 재현탁시켜, 최종 농도가 70 OD/㎖이 되도록 하였다.
DTT 및 Zymolase(LuBio Science, Luzern, Switzerland)를 각각 최종 농도 10 mM 및 0.5 U/OD에 첨가한 후, 혼합물을 저속 진탕에 의해, 37℃에서 인큐베이션하였다.
세포 벽 분해 후, 물에 희석된 용액의 광학 밀도를 측정하였다. 이들 값이 본래 값의 80%로 하락한 경우, 조심스럽게 원심분리한 후, 1 M 소르비톨 및 STC 완충액(0.98 M 소르비톨, 10 mM Tris pH 7.5, 10 mM CaCl2)에 의해, 세척함으로써, 분해 과정을 종결시켰다.
스페로플라스트를 50 ㎍/㎖ 송아지 흉선 DNA(Calbiochem/VWR, Dietikon, Switzerland)가 포함된 STC 완충액 중에 조심스럽게 재현탁시켜서, 최종 농도가 200 OD/㎖이 되도록 하였다. 100 ㎕의 분취량을 100 내지 200 ng의 플라스미드 DNA와 혼합한 후, 10분 동안 실온에서, 인큐베이션하였다.
1 ㎖ PEG 용액(19.6% PEG 8000 w/v, 10 mM Tris pH 7.5, 10 mM CaCl2)을 현탁액에 첨가한 후, 10분 동안 인큐베이션하고, 펠레팅하였다. 스페로플라스트를 30℃에서, 1 내지 2시간 동안, 25% YPD 및 7 mM CaCl2가 포함된 1 M 소르비톨 용액 1 ㎖에 재생시켰다.
상기 재생 세포에, 7 ㎖의 50℃ 가온 탑 한천(warm top agar)(0.67% 효모 질소 베이스 w/o 아미노산, 0.13% 드롭-아웃 분말 - 류신/우라실/히스티딘/트립토판/메티오닌 무함유, 0.086‰ 결여 아미노산 요구, 2% 글루코오스, 1 M 소르비톨, pH5.8 및 2.5% Noble 한천)을 첨가한 후, 혼합물을 예열된 소르비톨 함유 선택 플레이트(0.67% 효모 질소 베이스 w/o 아미노산, 0.13% 드롭-아웃 분말 - 류신/우라실/히스티딘/트립토판/메티오닌 무함유, 0.086‰ 결여 아미노산 요구, 2% 글루코오스, 1 M 소르비톨, pH5.8)에 균일하게 부었다.
플레이트를 30℃에서 3 내지 5일 동안 인큐베이션하였다. 적당한 선택 플레이트에서, 형질전환체를 재선택하였다.
각각의 생성된 균주를 아라비톨, 자일리톨 및 리비톨 생성에 대해, 3중 시험하였다.
이러한 목적을 위해, 클론을 처음에, 시드 배지(seed media)(0.67% 효모 질소 베이스-아미노산 무함유; 0.13% 드롭-아웃 분말-류신/우라실/히스티딘/트립토판/메티오닌 무함유; 0.086‰ 결여 아미노산 요구; 5% 글루코오스; pH5.7)에서, 30℃에서 밤새 성장시켰다.
이렇게 밤새 배양한 것 중, 출발 OD600이 0.2인 것으로서, 생성 배지(0.67% 효모 질소 베이스 - 아미노산 무함유; 0.13% 드롭-아웃 분말-류신/우라실/히스티딘/트립토판/메티오닌 무함유; 0.086‰ 결여 아미노산 요구; 15% 글루코오스; pH5.7)의 주요 배양액을 접종시켰다.
이들 배양액을 37℃에서 48시간 동안 성장시킨 후, 이동상으로서 100% 물의 등용매 조건에서, Aminex® HPX-87 칼럼(Bio-Rad, Hercules, California) 및 Waters® TQ-Detector(삼중 4중극 검출기(triple quadrupol detector)에 연결된 Acquity® UPLC, Waters, Milford, Massachusetts)을 이용한 HPLC/MS에 의해, 상청액의 아라비톨, 자일리톨 및 리비톨 농도를 측정하였다.
모든 시험 균주의 폴리올 역가가 표 8에 제시된다.
피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소를 이용하는 경우, 야생형 NADH-특이적 효소와 비교하여, 자일리톨 역가의 현저한 증가가 초래된다.
실시예
13. 피치아 오메리
에서
, 플라스미드 글루코노박터 옥시단스 NADPH-특이적 자일리톨 탈수소효소 유전자의 발현
피치아 스티피티스의 NADP-특이적 자일리톨 탈수소효소를 이용한 자일리톨 생성 균주에 더하여, 글루코노박터 옥시단스의 NADP-특이적 자일리톨 탈수소효소를 발현하는 제2 균주를 유전자 조작하였다.
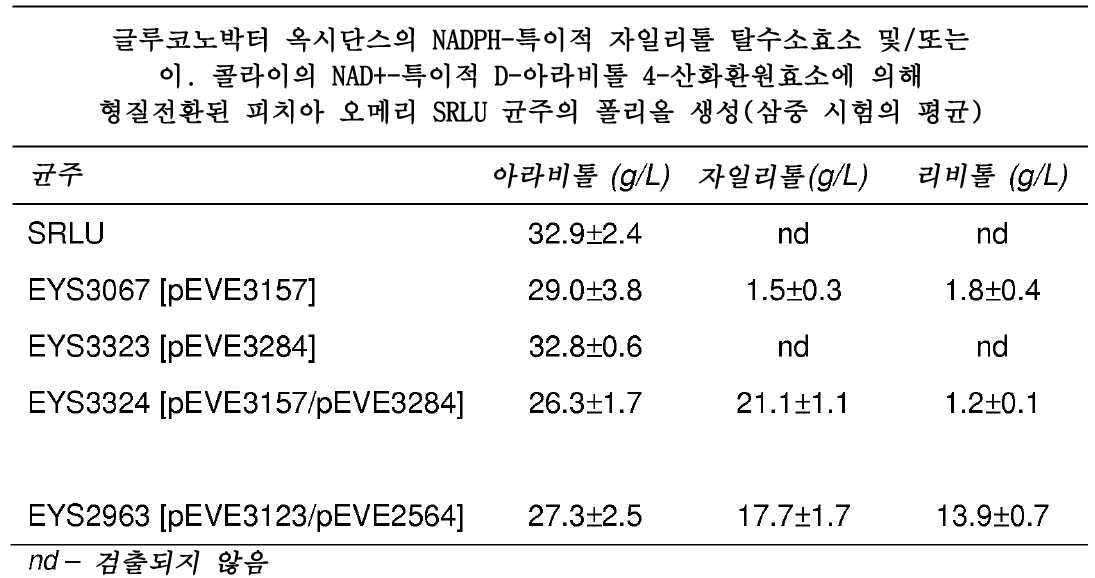
균주 EYS3324를 초래하는 것으로, 플라스미드 pEVE3157(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소) 및 pEVE3284(글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소)를 이용한 형질전환에 의한, 자일리톨 분비 효모 균주의 제작을 위한 숙주로서, ATCC 20209로부터 유도되었으며, 류신 및 우라실에 대해 영양 요구성인 피치아 오메리 균주 SRLU(MATh - leu2 ura3)(Piredda and Gaillardin, 1994, 상기)를 이용하였다.
대조군으로서, 각각 EYS3067 및 EYS3323을 초래하는 것으로서, 하기 단일 플라스미드에 의해 형질전환된 균주를 또한 생성하였다:
- pEVE3157(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소) 및
- pEVE3284(글루코노박터 옥시단스의 NADH-특이적 자일리톨 탈수소효소)
상기 균주의 제작을 위해 이용된 이. 콜라이 D-아라비톨 4-산화환원효소는, 피치아 스티피티스의 자일리톨 탈수소효소를 발현하는 균주에서 이용되는 poRR 프로모터와 달리, poPGK1 프로모터에 의해 조절된다.
그러나, 프로모터의 영향을 배제하고, 따라서, 글루코노박터 옥시단스의 자일리톨 탈수소효소를 발현하는 균주의 폴리올 수준을 피치아 스티피티스의 해당 효소를 발현하는 균주와 비교할 수 있도록 하기 위해, 추가의 균주를 생성하였다.
- pEVE3123(이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소) 및
- pEVE2564(피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소)
를 이용하여, SRLU 숙주를 형질전환시킴으로써, 이러한 균주 EYS2963를 얻었다.
효소 형질전환을 실시예 12에 개시된 바와 같이 실시하였다. 생성된 각각의 균주를 실시예 12에 개시된 바와 같이, 아라비톨, 자일리톨 및 리비톨 생성에 대해, 삼중 시험하였다.
시험된 모든 균주의 폴리올 역가가 표 9에 제시된다.
글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소를 발현하는 균주(EYS3324)의 자일리톨 역가는 피치아 스티피티스의 해당 효소를 발현하는 균주(EYS2963)의 역가와 유사하다. 그러나, 글루코노박터 옥시단스 효소는 더욱 낮은 리비톨 역가를 초래하고, 따라서, 자일룰로오스에 대해 더 큰 기질 특이성을 나타낸다.
실시예
14.
아라비톨
분비가
증가된
돌연변이 피치아 오메리 균주의 생성
더 많은 아라비톨 생산자 돌연변이를 피치아 오메리 ATCC 20209의 UV 방사 현탁액으로부터 선택하였다.
UV-방사 시스템(Vilber Lourmat, France)에 마이크로프로세서-조절 RMX-3W 방사계를 장착시켰다. 피치아 오메리를 37℃에서 YPD 한천(덱스트로오스 20g/L)에서 밤새 성장시켰다.
현탁액을 106 cfu/mL(OD620=0.4)에 도달되도록 제조한 후, 5 mL를 멸균 페트리 디쉬에 넣었다. 디쉬의 뚜껑을 제거한 후, 현탁액을 방사하였다. UV 파장은 254 nm이었으며, 방사 에너지는 1.8 10-2 J/㎠이었다. 90%의 효모 세포 사멸률이 획득되었다. 방사를 중지하고, 디쉬 뚜껑을 교체한 후, 상기 현탁액을 얼음 욕조에 배치된 멸균 튜브로 옮겼다.
20 mL의 YPD 액체 배지에 상기 돌연변이 유발 현탁액을 접종한 후, 37℃에서 12시간 동안 250 rpm으로 인큐베이션하였다.
인큐베이션 후, 돌연변이 유발 배양액을 멸균 40 % 글리세롤(V/V)로 희석하였다. 분취량을 5 mL 바이알에 분배한 후, -80℃에서 동결시켰다.
스크리닝은, 초고농도의 덱스트로오스(최대 600 g/L)에서 성장할 수 있는 피치아 오메리의 호삼투성을 기초로 하였다.
본 발명자들의 목표는 덱스트로오스 600 g/L 또는 700 g/L가 포함된 YPD 한천에서 모 균주 보다 더 빠르게 성장할 수 있는 돌연변이를 선택하는 것이었다.
해동된 분취량을 YPD600 및 YPD700에 확산시키고, 제1 출현 콜로니를 선택하여, 진탕 플라스크에서, 아라비톨의 생성에 대해, 시험하였다.
하위배양액 및 생성 배지는 각각 글루코오스 50g/L 또는 100 g/L, 효모 추출물 3g/L, MgSO4 1 g/L 및 KH2PO4 2g/L, pH 5.7로 구성되었다. 하위배양액(100 mL 플라스크 중 10 mL)을 37℃에서, 24시간 동안, 250 rpm에서 인큐베이션하였다. 생성물(500mL 플라스크 중 40 mL)에, 5 mL의 하위배양액을 접종한 후, 37℃에서, 64시간 동안, 250rpm에서 인큐베이션하였다.
돌연변이 피치아 오메리 균주를 그의 글루코오스의 고속 소비 및 그의 아라비톨의 다량의 생성에 대해 선택한 후, 프랑스에서, 2012년 3월 7일자로, 인스티투트 파스퇴르의 내셔널 콜렉션 오프 마이크로오가니즘 컬쳐스(Collection Nationale de Cultures de Microorganismes(CNCM), 25 rue du Docteur Roux, 75724 PARIS Cedex 15)에 등록번호 I-4605 하에, 기탁하였다.
실시예
15.
LEU2
결실 플라스미드의 제작
플라스미드 선택 및 유전자 혼입을 위해, 새로 생성된 CNCM I-4605 균주를 이용할 수 있도록 하기 위해, LEU2 오픈 리딩 프레임이 결실된 플라스미드를 제작하였다.
제1 단계에서, 피치아 오메리에서 사용될 수 있는 일반적 혼입 벡터를 사카로마이세스 세레비시애 CRE/loxP 시스템에서 적응시켰다. PstI 및 EcoRV 효소(New England Biolabs, Ipswich, Massachusetts)에 의한 제한 효소 절단에 의해, pUG73(Gueldener et al., 2002, Nucleic Acid Res, 30, e23)로부터 벡터 백본을 분리시켰다.
하기의 프라이머 쌍에 의해 생성된 것으로서, loxP 부위에 플랭킹된 피치아 오메리의 LEU2 선택 마커를 포함하는 PCR 단편이 삽입물로서 기능하였다.:
- EV3043
- EV3044
- 주형으로서, poARS(plig3 - FR 2772788 - 도 6 참조).
정방향 프라이머 EV3043은 AscI(밑줄) 부위 이후, SphI 부위(밑줄) 이후, 48 bp 길이의 loxP 단편(볼드체) 및 DraIII 부위(밑줄)를 포함한다. EV3043의 3' 말단은 피치아 오메리 LEU2 유전자의 증폭을 위한 추가의 25 bp 길이의 단편을 포함한다. 한편, 역방향 프라이머 EV3044는 NheI(밑줄) 부위 이후, NcoI 부위(밑줄) 이후, 48 bp 길이의 loxP 단편(볼드체) 및 MluI 부위(밑줄)를 포함한다. EV3044의 3' 말단은 피치아 오메리 LEU2 유전자의 증폭을 위한 추가의 25 bp 길이의 단편을 포함한다. 적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 상기 주형을 증폭시켰다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/65℃에서 10초/72℃에서 50초의 30회 순환 및 72℃에서 7분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
증폭된 단편은 추가의 서브클로닝을 위한 제2 PCR 반응에서, PstI 및 EcoRV 부위에 플랭킹되었다. 적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 하기의 것을 이용하여, 증폭 과정을 수행하였다:
- 프라이머 EV3056
(SEQ ID NO 26)(
PstI 부위(밑줄) 포함) 및
- 프라이머 EV3057
(SEQ ID NO 27)(
EcoRV 부위(밑줄) 포함)
98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/72℃에서 45초의 30회 순환 및 72℃에서 7분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
증폭된 2.5 kb LEU2 마커를 PstI 및 EcoRV 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해시킨 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)로 겔 정제하고, 실온에서, 2시간 동안, 2.4 kb PstI/EcoRV(New England Biolabs, Ipswich, Massachusetts)에 결찰시킨 다음, 겔 정제(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California)하고, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 벡터 pUG73의 백본(Gueldener et al., 2002 Nucleic acid Res, 30, e23)에 결찰시켰다 -(도 22). 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2787(도 23)은 2개의 loxP 부위에 플랭킹된, 내인성 프로모터 및 종결자의 조절 하의 피치아 오메리 LEU2 선택 마커를 포함한다. 또한, 게놈 내 혼입 부위에 상동인 영역의 클로닝을 보조하기 위해, 제1 loxP 및 제2 loxP 하류에 NheI 및 NcoI 부위를 도입시켰다.
그 후, 내인성 LEU2 오픈 리딩 프레임의 결실을 목표로 하였기 때문에, 제2 클로닝 단계에서, 혼입 벡터의 LEU2 마커를 스트렙토마이세스 노우르세이의 nat1 내성 유전자로 교체하였다.
SEQ ID NO 28의 제시 서열에 따라, GeneArt® Gene Synthesis(Life Technologies, Regensburg, Germany)에 의해, 스트렙토마이세스 노우르세이의 nat1 유전자를 인코딩하는 DNA 단편을 화학적 합성하였다.
nat1 유전자를 코딩하는 서열 S60706.1(http://www.ncbi.nlm.nih.gov/nuccore/X55392.1에서 얻음)의 뉴클레오티드 204 내지 776을 주형으로 사용하였으며, http://genomes.urv.es/OPTIMIZER/에서 얻은 Optimizer 프로그램을 이용하여, 표 7(상기)에 따라, 피치아 오메리 ATCC 20209에서 사용하기 위한 코돈 최적화에 적용하였다.
추가의 클로닝을 용이하게 하기 위해, 생성된 서열의 5' 및 3' 말단에, 각각 제한 효소 AscI(GGCGCGCC) 및 SphI(GCATGC)의 인식 부위를 인코딩하는 뉴클레오티드를 텍스트 파일에 수작업으로 첨가하였다. 또한, 효모의 Kozak-형 서열의 -3 위치에 아데노신을 추가하기 위해, 아데노신 삼중자를 개시 ATG 앞에 포함시켰다.
그 후, 최종 서열(SEQ ID NO: 28)을 GeneArt(Regensburg, Germany)에 따른 합성을 위해, 제공하였다. nat1 유전자를 인코딩하는 합성 DNA 단편을 pMA-T 유도 벡터(12ABTV4P, 도 24)에 5 ㎍ 동결건조 플라스미드 DNA로서, 전달하였다.
nat1 유전자의 클로닝을 위해, 리불로오스 환원 효소(poRR) 프로모터 및 종결자를 포함하는 벡터를 이용하였다. 종결자를 오로티딘-5'-포스페이트 탈카르복실화효소(poURA3)로 교체한 후, 프로모터와 종결자 서열 사이에, nat1 유전자를 도입시켰다.
이러한 목적을 위해, the 오로티딘-5'-포스페이트 탈카르복실화효소(poURA3) 종결자를 하기의 것을 이용한 PCR에 의해 생성시켰다:
- 프라이머 EV3393
(SEQ ID NO 29)(
SphI 부위(밑줄) 포함) 및
- 프라이머 EV3394
(SEQ ID NO 30)(
SacII 부위(밑줄) 포함) 및
- 주형으로서, pEVE2523(도 7).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/59℃에서 10초/72℃에서 10초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 239 bp poURA3 종결자를 SphI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해시킨 후, 실온에서, 2시간 동안, SphI 및 SacII 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 선형화된 pEVE2681의 11 kb 벡터 백본에 결찰시키고, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)에 의해, 겔 정제하였다. 결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가로 특성화하였다.
제2 클로닝 단계에서, SphI 및 AscI 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, 12ABTV4P(도 24)로부터 방출시켰다. 또한, 실온에서, 15분 동안 평활화 효소 믹스(New England Biolabs, Ipswich, Massachusetts)에 의해, SphI 부위의 평활화를 수행한 후, SphI 및 AscI 사이의 분해 과정에서, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 그 후, 587 bp 겔-정제된 단편(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California)을 SphI 및 AscI 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 절단된 전술된 벡터의 겔 정제된 10.5 kb 벡터 백본에 결찰시켰다.
또한, 상기 벡터의 SphI 부위를 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 15분 동안 평활화한 후, AscI에 의한 분해의 수행 전에, 10분 동안 70℃에서 열 불활성화 단계를 수행하였다. 또한, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서 1시간 동안, 상기 벡터를 탈인산화시켰다. T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 2시간 동안, 결찰 과정을 수행하였다.
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2798(도 25)은 피치아 오메리 리불로오스 환원 효소(poRR) 프로모터 및 오로티딘-5'-포스페이트 탈카르복실화효소(poURA3) 종결자에 플랭킹된 nat1 약제 내성 마커를 포함한다.
상기 혼입 벡터에서, 피치아 오메리 LEU2 선택 마커의 교체를 위해, nat1 발현 카세트를 사용하였다. 추가의 클로닝을 용이하게 하기 위해, 하기의 것에 의한 PCR에 의해, nat1 카세트가 XbaI(프라이머 EV3643 내 밑줄 부분) 및 MluI(프라이머 EV3644 내 밑줄 부분) 부위에 플랭킹되어야 한다:
- 프라이머 EV3643
- 프라이머 EV3644
(SEQ ID NO 32).
프라이머 EV3643은 XbaI 부위 이후, 추가의 ClaI 부위(점선)를 포함한다. pEVE2798이 주형으로 기능한다(도 25).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/54℃에서 10초/72℃에서 25초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 상기 1.3 kb nat1 발현 카세트를 MluI 및 XbaI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해시킨 후, 실온에서, 2시간 동안, MluI 및 XbaI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 선형화된 pEVE2787(도 23)의 2.6 kb 벡터 백본에 결찰시키고, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)에 의해, 겔 정제하였다(도 26).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가로 특성화하였다.
생성된 플라스미드 pEVE2852(도 27)는 2개의 loxP 부위에 플랭킹된, 리불로오스 환원 효소(poRR) 프로모터 및 오로티딘-5'-포스페이트 탈카르복실화효소(poURA3) 종결자의 조절하의 nat1 선택 마커를 포함한다.
상기 혼입 플라스미드를 일정 부분 게놈 내로의 부위 특이적 혼입에 필요한 피치아 오메리 상동성 단편을 포함하지 않는다. 이러한 부위는 다음 단계에서 부착된다.
LEU2 오픈 리딩 프레임 상류의 5' 상동 영역을 하기의 것을 이용하여, 50 ng poARS 벡터(도 6)로부터 증폭시켰다:
- 프라이머 EV3548
(SEQ ID NO 33)(
PstI 부위(밑줄) 포함) 및
- 프라이머 EV3549
(SEQ ID NO 34)(
SphI 부위(밑줄) 포함).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, PCR을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/61℃에서 10초/72℃에서 15초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 567 bp 단편을 PstI 및 SphI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해시킨 후, 실온에서, 2시간 동안, PstI 및 SphI 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 선형화된 pEVE2852(도 27)의 3.9 kb 벡터 백본에 결찰시키고, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)에 의해, 겔 정제하였다(도 29).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가로 특성화하였다.
생성된 플라스미드 pEVE2855(도 28)는 2개의 loxP 부위에 플랭킹된 nat1 마커 및 LEU2 오픈 리딩 프레임 상류의 5' 영역에 상동인 단편을 포함한다.
LEU2 오픈 리딩 프레임 상류의 3' 상동 영역을 하기의 것을 이용하여, 50 ng poARS 벡터(도 6)로부터 증폭시켰다:
- 프라이머 EV3550
(SEQ ID NO 35)(
NcoI 부위(밑줄) 포함) 및
- 프라이머 EV3551
(SEQ ID NO 36)(
NheI 부위(밑줄) 포함).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, PCR을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/51℃에서 10초/72℃에서 25초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 수행하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 상기 1.3 kb 단편을 NcoI 및 NheI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해시킨 후, 실온에서, 2시간 동안, NcoI 및 NheI 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 선형화된 pEVE2855(도 28)의 4.4 kb 벡터 백본에 결찰시키고, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)에 의해, 겔 정제하였다(도 29).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가로 특성화하였다.
생성된 최종 LEU2 결실 플라스미드 pEVE2864(도 30)는 2개의 loxP 부위에 플랭킹된 nat1 마커 및 LEU2 오픈 리딩 프레임 상류의 3' 영역에 상동인 단편을 포함한다.
실시예
16.
류신에
대해 영양
요구성인
돌연변이 피치아 오메리 균주의 제작
생성된 피치아 오메리 CNCM I-4605 균주가 일정 정도 영양 요구성을 나타내지 않기 때문에, 유전자 혼입을 위해, LEU2 선택 마커를 이용할 수 있도록 하기 위해, LEU2 오픈 리딩 프레임 결실 과정을 수행하였다.
이러한 목적을 위해, 플라스미드 pEVE2864(도 30)를 EcoRV 및 PstI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 2.5시간 동안, 37℃에서, 제한 효소 분해한 후, 상기 혼합물을 실시예 12에 개시된 과정에 따라, Mut165 균주를 형질전환시키는데 사용하였다.
상기 재생 세포에, 25 ㎍/㎖ 나타마이신이 포함된 7 ㎖의 50℃ 가온 탑 한천(1% 효모 추출물, 2% 펩톤, 2% 글루코오스, 1 M 소르비톨, pH 5.8 및 2.5% Noble 한천)을 첨가한 후, 혼합물을 25 ㎍/㎖ 나타마이신이 포함된 예열된 소르비톨 함유 선택 플레이트(1% 효모 추출물, 2% 펩톤, 2% 글루코오스, 1 M 소르비톨, pH 5.8 및 2% 한천)에 균일하게 부었다. 플레이트를 30℃에서 4일 동안 인큐베이션하였다. LEU2 오픈 리딩 프레임의 결실을 류신이 포함되지 않은 선택 플레이트에서의 무성장에 의해 입증한 후, 하기의 것을 이용한 콜로니 PCR에 의해, 확인하였다:
- 프라이머 EV3393
(SEQ ID NO 29) 및
- 프라이머 EV3795
(SEQ ID NO 37).
상기 1.6 kb 단편을 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/51℃에서 10초/72℃에서 25초의 30회 순환 및 72℃에서 5분의 최종 확장 단계에 의해, 증폭시켰다.
생성된 균주는 CNCM I-4605 백그라운드의 LEU2 유전자의 전체 오픈 리딩 프레임 결실을 포함하며, 프랑스에서, 2015년 2월 5일자로, 등록번호 I- 4955 하에, 인스티투트 파스퇴르의 내셔널 콜렉션 오프 마이크로오가니즘 컬쳐스(Collection Nationale de Cultures de Microorganismes(CNCM), 25 rue du Docteur Roux, 75724 PARIS cedex 15)에 기탁되었다.
실시예
17. 피치아 스티피티스
의
NADPH
-특이적
자일리톨
탈수소효소 및 이. 콜라이
의
NAD
+
-특이적 D-
아라비톨
4-산화환원효소를 포함하는 이중 발현 플라스미드의 제작
류신에 대해서만 영양 요구성인 돌연변이 피치아 오메리 균주에서, 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 및 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소가 발현될 수 있도록 하기 위해, 이중 발현 플라스미드의 제작이 필요하였다.
피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소를 포함하는 발현 카세트를 SpeI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, pEVE2562(도 12)로부터 방출시켰다. 상기 1.9 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안 평활화 효소 믹스(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 그 후, 삽입물을, 선형화, 평활화, 탈인산화(37℃에서, 1시간 동안, 앤타크틱 포스파타아제 - New England Biolabs, Ipswich, Massachusetts 사용)된 12.1 kb SpeI에 결찰시킨 다음, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소를 포함하는 pEVE3157 백본(도 21)을 겔 정제하였다(도 31).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의한 추가로 특성화하였다.
생성된 플라스미드 pEVE3318(도 32)는 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 리불로오스 환원 효소(poRR) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소의 이중 발현 작제물 및 poLEU2 선택 마커를 포함한다.
실시예
18. 피치아 오메리
에서
, 이. 콜라이
NAD
+
-특이적 D-
아라비톨
4-산화환원효소 유전자 및 피치아 스티피티스
NADPH
-특이적
자일리톨
탈수소효소 유전자의 발현을 위한 혼입 벡터의 제작
이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자는 궁극적으로 피치아 오메리 게놈의 필수 부분이 되어야 한다. 따라서, pEVE2852의 nat1 선택 마커를 교체한 후, 아라비톨 산화환원효소 및 자일리톨 탈수소효소의 이중 발현 작제물을 혼입시킴으로써, LEU2 선택 마커를 가지는 혼입 벡터를 제작해야 한다.
이러한 목적을 위해, AscI 및 SphI 부위에 플랭킹된 피치아 오메리 LEU2 오픈 리딩 프레임을 하기의 것을 이용한 PCR에 의해 생성하였다:
- 프라이머 EV3645
(SEQ ID NO 38) 및
- 프라이머 EV3646
(SEQ ID NO 39).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 50 ng의 poARS(도 6) 주형의 증폭 과정을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/57℃에서 10초/72℃에서 20초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 달성하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 그 후, 증폭된 LEU2 오픈 리딩 프레임을 AscI 및 SphI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해하였다.
또한, 실온에서, 15분 동안 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, SphI 부위의 평활화를 수행한 후, SphI 및 AscI 사이의 분해 과정에서, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 그 후, 1.1 kb 겔-정제된 단편을 SphI 및 AscI 제한 효소(New England Biolabs, Ipswich, Massachusetts)에 의해 절단된 pEVE2811의 겔-정제된 11 kb 벡터 백본에 결찰시켰다. 또한, 상기 벡터의 SphI 부위를 실온에서, 15분 동안 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, AscI에 의한 분해 전에, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 또한, 상기 벡터를 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서 1시간 동안, 탈인산화시켰다. LEU2 오픈 리딩 프레임 및 벡터 백본의 결찰 과정을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 2시간 동안, 수행하였다.
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2862(도 33)는 피치아 오메리 리불로오스 환원 효소(poRR) 프로모터 및 오로티딘-5'-포스페이트 탈카르복실화효소(poURA3) 종결자에 플랭킹된 피치아 오메리 LEU2 마커를 포함한다.
그 후, LEU2 마커를 하기의 것을 이용한 PCR에 의해, 증폭시켰다:
- 프라이머 EV3643
(SEQ ID NO 31)(ClaI 부위 포함) 및
- 프라이머 EV3644
(SEQ ID NO 32)(
MluI 부위(밑줄) 포함) 및 주형으로서, pEVE2862(도 33).
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/54℃에서 10초/72℃에서 30초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 달성하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다. 증폭된 1.8 kb 길이의 LEU2 단편을 ClaI 및 MluI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해한 후, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, pEVE2852(도 27)의 2.6 kb ClaI 및 MluI(New England Biolabs, Ipswich, Massachusetts) 제한 효소 분해되고, 겔-정제된 벡터 백본에, 실온에서, 2시간 동안, 결찰시켰다(도 34).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE2865(도 35)는 2개의 loxP 부위에 플랭킹된 피치아 오메리 LEU2 마커를 포함한다.
혼입 벡터의 클로닝을 위해, pEVE2865를 SalI 효소(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해한 후, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 15분 동안 평활화한 다음, 10분 동안 70℃에서 열 불활성화 단계를 수행하고, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서 1시간 동안, 탈인산화시켰다.
상기 벡터 백본의 4.5 kb 겔-정제된 단편을 결찰 과정에 사용하였다. NdeI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, pEVE3318(도 32)로부터 방출된 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자 및 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소의 이중 발현 작제물이 삽입물로서 기능하였다.
Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 상기 4.4 kb 단편을 겔 정제한 후, 실온에서, 15분 동안 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하고, 추가의 겔 정제 과정을 수행하였다. pEVE2865의 벡터 백본 및 pEVE3318의 삽입물을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 2시간 동안 결찰시켰다(도 34).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3387(도 36)은 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자의 이중 발현 작제물을 포함한다. 2개의 loxP 부위에 플랭킹된 피치아 오메리 LEU2 유전자가 선택 마커로서 기능한다.
실시예
19. 배지로
자일리톨을
분비하는
제1 세대
혼입 피치아 오메리 균주의 제작
피치아 오메리의 게놈내로 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자를 무작위 혼입시키기 위해, 전술된 벡터를 사용하였다.
이러한 목적을 위해, 류신에 대해 영양 요구성인 균주 CNCM I-4955(실시예 16)를 실시예 12에 개시된 절차에 따라, 37℃에서 3시간 동안, NotI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해된 pEVE3387(도 36)에 의해, 형질전환시켰다. 형질전환체를 류신이 포함되지 않은 소르비톨 플레이트에서 선택하였다.
생성된 균주는 피치아 오메리 게놈내로 무작위 혼입된 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자를 포함하며, 프랑스에서, 2015년 5월 20일자로, 등록번호 I-4982 하에, 인스티투트 파스퇴르의 내셔널 콜렉션 오브 마이크로오가니즘 컬쳐스(Collection Nationale de Cultures de Microorganismes(CNCM), 25 rue du Docteur Roux, 75724 Cedex 15)에 기탁되었다.
실시예
20. 글루코노박터 옥시단스
의
NADPH
-특이적
자일리톨
탈수소효소 및 이.
콜라이의
NAD
+
-특이적 D-
아라비톨
4-산화환원효소를 포함하는 이중/삼중 발현 플라스미드의 제작
류신에 대해서만 영양 요구성인 돌연변이 피치아 오메리 균주에서, 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 및 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소가 발현될 수 있도록 하기 위해, 이중 발현 플라스미드의 제작이 필요하였다.
글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소를 포함하는 발현 카세트를 SpeI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, pEVE3284(도 10)로부터 방출시켰다. 상기 1.6 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안 평활화 효소 믹스(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 사용된 벡터 백본은 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소를 포함하는 12.1 kb SpeI-선형화(New England Biolabs, Ipswich, Massachusetts) 및 겔-정제된(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California) pEVE3157 백본(도 21)으로 구성되었다.
상기 백본을 또한, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 실온에서, 15분 동안 평활화한 후, 10분 동안 70℃에서 효소의 열 불활성화를 수행한 다음, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서 1시간 동안, 탈인산화하였다. T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 2시간 동안, 결찰 과정을 수행하였다(도 37).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3322 및 pEVE3324(도 38)는 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소의 이중 발현 작제물 또는 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 2개의 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 유전자의 삼중 발현 작제물 및 poLEU2 선택 마커를 포함한다.
실시예
21. 피치아 오메리
에서
, 이. 콜라이
NAD
+
-특이적 D-
아라비톨
4-산화환원효소 유전자 및 글루코노박터 옥시단스
NADPH
-특이적
자일리톨
탈수소효소 유전자의 발현을 위한 혼입 벡터의 제작
피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 및 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자를 포함하는 혼입 벡터 이외에, 또한, 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소를 포함하는 플라스미드를 생성하였다.
이러한 목적을 위해, 1 또는 2개의 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 및 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소를 포함하는 이중 및 삼중 발현 카세트를 NdeI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, 각각 pEVE3322 및 pEVE3324(도 38)로부터 방출시켰다.
상기 4.1 kb 및 5.7 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 겔-정제된(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California), 5.7 kb SalI-선형화 pEVE2865(도 35)가 벡터로서 기능하였다.
상기 벡터 백본을 또한 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 실온에서, 15분 동안, 평활화한 후, 10분 동안 70℃에서, 열 불활성화를 수행한 다음, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서, 1시간 동안, 탈인산화하였다. 벡터와 삽입물의 결찰 과정을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 실온에서, 2시간 동안, 수행하였다(도 39).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3390 및 pEVE3392(도 40)는 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 1 또는 2개의 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 유전자의 이중 또는 삼중 발현 작제물을 포함한다. 2개의 loxP 부위에 플랭킹된 피치아 오메리 LEU2 유전자가 선택 마커로서 기능한다.
실시예
22.
100 g
/L 이상의
자일리톨을
분비할 수 있는
제2 세대
혼입 균주의 제작
2개의 이종 효소의 추가 카피를 추가로 혼입시키기 위해, 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 피치아 스티피티스의 NADPH-특이적 자일리톨 탈수소효소 유전자의 무작위 혼입 카피를 포함하는 제1 세대 균주 CNCM I-4982를 사용하였다.
그러나, 상기 작제물이 혼입될 수 있도록 하기 위해, LEU2 선택 마커가 제거되어야 한다. 이러한 목적을 위해, 제1 새대 균주 CNCM I-4982를 실시예 12에 개시된 절차에 따라, 벡터 pEVE3163으로 형질전환시켰다. 벡터 pEVE3163은 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 박테리오파지 P1의 CRE 재조합효소(표 7에 따라 최적화된 코돈)를 포함한다. LEU2 선택 마커의 제거를 류신이 포함되지 않은 플레이트에서, 클론의 비성장에 의해, 확인하였다.
생성된 균주 EYS3842를 실시예 12에 개시된 절차에 따라, 37℃에서 3시간 동안, NotI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해된 pEVE3390 또는 pEVE3392(도 40)로 형질전환시켰다. 형질전환체를 류신이 포함되지 않은 소르비톨 플레이트에서 선택하였다.
생성된 제2 세대 균주 EYS3929는 게놈내로 무작위 혼입된 2개의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 하나는 G. oxydan의 것이고, 다른 하나는 피치아 스티피티스의 것인 2개의 NADPH-특이적 자일리톨 탈수소효소 유전자를 포함한다. 한편, 균주 EYS3930은 추가의 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 유전자를 포함한다.
실시예
23. 이. 콜라이
의
NAD
+
-특이적 D-
아라비톨
4-산화환원효소 및 글루코노박터 옥시단스의
NADPH
-특이적
자일리톨
탈수소효소의 추가적 유전자 카피의 혼입을 위해 이용되는 추가적 벡터의 제작
추가의 혼입 벡터의 제작을 위해, 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소의 이중 발현 카세트를 하기의 것을 이용한 PCR에 의해, 증폭시켰다:
- 프라이머 EV4904
(SEQ ID NO 40)(
SmaI 부위 포함) 및
- 프라이머 EV4905
(SEQ ID NO 41)(
SmaI 부위(밑줄) 포함) 및
주형으로서, pEVE3321.
적절한 1X 완충액에, 0.02 U/㎕의 iProof™ 중합효소(BIO-RAD, Hercules, California)를 함유하며, 200 μM의 각 dNTP 및 0.5 μM의 각 프라이머로 이루어진 반응 믹스에서, 증폭 과정을 수행하였다. 98℃에서 30초의 초기 변성 단계 후, 98℃에서 10초/68℃에서 10초/72℃에서 75초의 30회 순환 및 72℃에서 5분의 최종 확장 단계의 PCR을 달성하였다. PCR 생성물을 1% 아가로오스 겔에서 분리하고, 추출한 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 사용하여, 정제하였다.
증폭된 3.9 kb 길이의 단편을 SmaI(New England Biolabs, Ipswich, Massachusetts)에 의해, 제한 효소 분해한 후, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 4.4 kb PvuII(New England Biolabs, Ipswich, Massachusetts) 선형화된 후, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts) 탈인산화되고, 겔-정제된 pEVE2865(도 35)의 벡터 백본에, 실온에서, 2시간 동안, 결찰시켰다(도 41).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE4390(도 42)은 피치아 오메리 리불로오스 환원 효소 프로모터 및 종결자(poRR)에 플랭킹된 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 유전자 및 피치아 오메리 포스포글리세레이트 키나아제(poPGK1) 프로모터 및 트랜스케톨라아제(poTKL) 종결자의 조절 하의 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소의 이중 발현 작제물을 포함한다. 2개의 loxP 부위에 플랭킹된 피치아 오메리 LEU2 유전자가 선택 마커로서 기능한다.
실시예
24.
글루코노박터
옥시단스의
NADPH
-특이적
자일리톨
탈수소효소 및 랄스토니아
솔라나세아룸의
NAD
+
-특이적 D-
아라비톨
4-산화환원효소의 혼입에 이용되는 벡터의 제작
글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 및 랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소의 발현을 위한 추가적 혼입 벡터를 하기와 같이 제작하였다: 제1 단계로, 2개의 상기 유전자를 포함하는 이중 발현 벡터를 생성하였다. 이들 이중 발현 카세트를 혼입 loxP 벡터에 클로닝하였다.
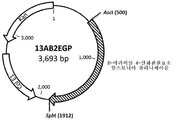
랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자를 인코딩하는 DNA 단편을 SEQ ID NO 42의 제시 서열에 따라, GeneArt® Gene Synthesis(Life Technologies, Regensburg, Germany)에 의해, 화학적으로 합성하였다.
dalD 유전자를 코딩하는 서열 AL646052.1의 뉴클레오티드 2310548 내지 2309151(http://www.ncbi.nlm.nih.gov/nuccore/AL646052에서 얻음)을 주형으로 사용하였으며, http://Genomes.urv.es/OPTIMIZER/에서 얻은 Optimizer 프로그램을 이용하여, 표 7(상기)에 따라, 피치아 오메리 ATCC 20209에서 사용하기 위해, 코돈 최적화에 적용하였다. 추가적 클로닝을 용이하게 하기 위해, 생성된 서열의 5' 및 3' 말단에, 각각 제한 효소 AscI(GGCGCGCC) 및 SphI(GCATGC)의 인식 부위를 인코딩하는 뉴클레오티드를 텍스트 파일에, 수작업 첨가하였다. 또한, 효모의 Kozak-형 서열의 -3 위치에 아데노신을 추가하기 위해, 아데노신 삼중자를 개시 ATG 앞에 포함시켰다.
그 후, 최종 서열(SEQ ID NO 42)을 합성을 위해, GeneArt(Regensburg, Germany)에 제공하였다. dalD 유전자를 인코딩하는 합성 DNA 단편을 pMA-RQ 유도 벡터(13AB2EGP, 도 43)에, 5 ㎍ 동결건조 플라스미드 DNA로서 전달하였다.
랄스토니아 솔라나세아룸의 D-아라비톨 4-산화환원효소의 1.4 kb 단편을 AscI 및 SphI(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 분해에 의해, 벡터 13AB2EGP(도 43)로부터 방출시킨 후, Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)에 의해, 겔 정제하였다. 그 후, 삽입물을 AscI 및 SphI(New England Biolabs, Ipswich, Massachusetts)에 의해, 선형화된 pEVE2560(도 8)의 11.8 kb 백본과 결찰시킨 후, T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 겔 정제하였다(도 44).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE3898(도 45)은 피치아 오메리의 리불로오스 환원 효소 프로모터 및 종결자에 플랭킹된 코돈-최적화된 랄스토니아 솔라나세아룸 NAD+-특이적 D-아라비톨 4-산화환원효소 및 poLEU2 선택 마커를 포함한다.
다음 단계에서, 포스포글리세레이트 키나아제 프로모터(poPGK) 및 리불로오스 환원 효소 종결자(poRR)에 플랭킹된 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소를 포함하는 발현 카세트를 SpeI 및 SacII 효소(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, pEVE3960로부터 방출시켰다. 상기 1.8 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 겔-정제된(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California) 13.2 kb SalI-선형화된 pEVE3898가 벡터로서, 기능하였다. 상기 벡터 백본을 또한 실온에서, 15분 동안, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하고, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서, 1시간 동안, 탈인산화하였다. 벡터와 삽입물의 결찰 과정을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 2시간 동안 수행하였다(도 44).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE4077(도 46)은 피치아 오메리 리불로오스 환원 효소 프로모터 및(poRR) 종결자의 조절 하의 랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소 및 피치아 오메리 포스포글리세레이트 키나아제 프로모터(poPGK) 및 리불로오스 환원 효소 종결자(poRR)에 플랭킹된 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소의 이중 발현 작제물 및 poLEU2 선택 마커를 포함한다.
마지막으로, 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 및 랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소의 이중 발현 카세트를 SapI(New England Biolabs, Ipswich, Massachusetts)를 이용한 제한 효소 절단에 의해, pEVE4077(도 46)로부터 방출시켰다. 상기 5.9 kb 단편을 Zymoclean™ 겔 DNA 회수 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 겔 정제한 후, 실온에서, 15분 동안, 평활화 효소 믹스 키트(New England Biolabs, Ipswich, Massachusetts)에 의해, 평활화한 다음, 10분 동안 70℃에서, 효소의 열 불활성화를 수행하였다. 겔-정제된(Zymoclean™ 겔 DNA 회수 키트 - Zymo Research Corporation, Irvine, California) 4.4 kb EcoRV-선형화된 pEVE2865(도 35)가 벡터로서, 기능하였으며, 앤타크틱 포스파타아제(New England Biolabs, Ipswich, Massachusetts)를 이용하여, 37℃에서, 1시간 동안, 탈인산화하였다. 벡터와 삽입물의 결찰 과정을 T4 DNA 리가아제(New England Biolabs, Ipswich, Massachusetts)를 사용하여, 실온에서, 2시간 동안 수행하였다(도 44).
결찰 혼합물을 이용한 XL10 Gold 초형질전환성 세포(Agilent Technologies, Santa Clara, California)의 형질전환 후, Zyppy™ 플라스미드 미니프렙 키트(Zymo Research Corporation, Irvine, California)를 이용하여, 플라스미드 DNA를 분리한 후, 제한 효소 분해 및 서열 분석(Microsynth, Balgach, Switzerland)에 의해, 추가로 특성화하였다.
생성된 플라스미드 pEVE4377(도 47)은 2개의 loxP 부위에 플랭킹된 poLEU2 선택 마커 및 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소 및 랄스토니아 솔라나세아룸의 NAD+-특이적 D-아라비톨 4-산화환원효소의 이중 발현 작제물을 포함한다.
실시예
25.
자일리톨의
생산성이 증대된
제3 세대
혼입 균주의 제작
제2 세대 균주 EYS3929 및 EYS3930의 LEU2 마커(실시예 22)를 벡터 pEVE3163을 이용하여, 실시예 18에 개시된 바와 같이, 록스 아웃(lox out)시켰다. 생성된 균주 EYS4118 및 EYS4119를 각각 pEVE4377(도 47) 및 pEVE4390(도 42)으로 형질전환시켰다. 상기 벡터를 실시예 12에 개시된 절차에 따라, 37℃에서 3시간 동안, NotI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해하였다. 형질전환체를 류신이 포함되지 않은 소르비톨 플레이트에서 선택하였다.
생성된 제3 세대 균주 EYS4353은 게놈내로 무작위 혼입된 것으로서, 2개는 이. 콜라이의 것이고, 하나는 랄스토니아 솔라나세아룸의 것인 3개의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 2개는 글루코노박터 옥시단스의 것이고, 하나는 피치아 스티피티스의 것인 3개의 NADPH-특이적 자일리톨 탈수소효소 유전자를 포함한다.
한편, 두 번째 제3 세대 균주는 백그라운드의 피치아 스티피티스의 하나의 카피, 이. 콜라이의 NAD+-특이적 D-아라비톨 4-산화환원효소의 3개의 카피, 및 글루코노박터 옥시단스의 NADPH-특이적 자일리톨 탈수소효소의 3개의 카피를 포함하며, 프랑스에서, 2015년 3월 5일자로, 등록번호 I-4960 하에, 인스티투트 파스퇴르의 내셔널 콜렉션 오브 마이크로오가니즘 컬쳐스(Collection Nationale de Cultures de Microorganismes)(CNCM), 25 rue du Docteur Roux, 75724 PARIS Cedex 15)에 기탁되었다.
실시예
26.
제4 세대
혼입 균주의 제작
제3 세대 균주 CNCM I-4960의 LEU2 마커(실시예 25)를 벡터 pEVE3163을 이용하여, 실시예 18에 개시된 바와 같이, 록스 아웃시켰다. 생성된 균주 EYS4955를 실시예 12에 개시된 절차에 따라, 37℃에서 3시간 동안, NotI(New England Biolabs, Ipswich, Massachusetts)에 의해 제한 효소 분해된 pEVE4377(도 47)로 형질전환시켰다. 형질전환체를 류신이 포함되지 않은 소르비톨 플레이트에서 선택하였다.
생성된 제4 세대 균주는 게놈내로 무작위 혼입된 것으로서, 3개는 이. 콜라이의 것이고, 하나는 랄스토니아 솔라나세아룸의 것인 4개의 NAD+-특이적 D-아라비톨 4-산화환원효소 유전자 및 3개는 글루코노박터 옥시단스의 것이고, 하나는 피치아 스티피티스의 것인 4개의 NADPH-특이적 자일리톨 탈수소효소 유전자를 포함하며, 프랑스에서, 2015년 5월 20일자로, 등록번호 I-4981 하에, 인스티투트의 내셔널 콜렉션 오브 마이크로오가니즘 컬쳐스(Collection Nationale de Cultures de Microorganismes(CNCM), 25 rue du Docteur Roux, 75724 PARIS Cedex 15)에 기탁되었다.
실시예
27. 피치아 오메리 균주를 이용한 폴리올 생성(합성 배지)
전술된 바와 같이 제작된 효모 균주 CNCM I-4605, CNCM I-4982, CNCM I-4960 및 CNCM I-4981을 하기 프로토콜에 따라 발효시켰다.
발효 과정은 질소-제한 하에 진행되며, 성장 단계 및 생성 단계로 구분될 수 있다. 성장 단계 동안, 배지 내의 암모니아는 완전히 소비되고, 바이오매스가 생성되며, 바이오매스 생성이 중단되는 경우, 생성 단계가 개시되고, 폴리올 수준이 증가한다. 개시된 발효 과정에 사용되는 플랫폼은 INFORS HT의 Multifors 2이며, 1 L 작업량의 용기가 이용되었다. 발효기에는 2개의 Rushton 6-날 디스크 터빈(Rushton six-blade disc turbines)이 장착되었다. 발효기의 스파징(sparging)에는 공기가 이용되었다.
배양 전 과정 동안, 온도, pH, 교반 및 용존 산소량이 조절되었다. 온도는 36 ℃에서 유지되었다. pH는 5M KOH의 자동 첨가에 의해, 3으로 유지되었다.
용존 산소량은 1.0 vvm으로 유지되었으며, 초기 교반기 속도는 300 rpm으로 설정되었다. 용존 산소(DO)가 20% 이하로 하강하는 것을 방지하기 위해, 자동 교반 연속 과정(automatic stirring cascade)을 사용하였다. 발효 과정에 사용된 작업 조건은 표 10에 정리되어 있다.
발효기의 접종을 위해, 1-단계 증식 배양이 이용되었다. 이용된 증식 배양 배지의 조성은 표 11에 개시된다. 증식 배양액은, 4개의 칸막이가 구비된 500-㎖ 진탕 플라스크(주문 제작)에서, 100 ㎖의 배지를 접종함으로써, 제조하였다. 진탕 플라스크를 30℃, 150 rpm의 진탕 테이블에서 인큐베이션하였다. 세포를 중기-지수 단계까지, 약 24시간 동안, 성장시켰다.
접종 전에, 접종물과 동량의 발효기 내 배지 양을 제거한 후, 증식 배지 분취량을 1 L의 최종 용적 및 약 0.2(CDW 약 0.03 g/L)의 OD600-개시로, 발효기의 접종을 위해, 사용하였다. 발효기에 사용된 배지의 조성은 표 12에 개시된다.
일정한 간격으로 샘플을 채취하고, 전체 발효 브로쓰를 글루코오스 소비량 및 세포 외 폴리올(자일리톨, 아라비톨 및 리비톨) 형성에 대해, 분석하였다. 추가의 통상 발효 대사물(글리세롤, 아세테이트, 에탄올, 피루베이트, 말레이트, 푸마레이트 및 숙시네이트)을 측정하였다. 바이오매스의 증가를 한편으로, OD600에 의해, 추적 조사하고, 다른 한편으로, 세포 건조 중량(CDW) 측정에 의해, 추적 조사하였다. 폴리올 생성, 아라비톨 또는 자일리톨 수율 및 생산성의 측정을 위해, 전술된 측정치를 이용하였으며, 그 결과는 표 13에 제시된다.
피치아 오메리 CNCM I-4605는 아라비톨만을 생성한다.
피치아 오메리 CNCM I-4982는 아라비톨, 자일리톨 및 리비톨을 생성한다. 이들 균주에서, NAD+-D-아라비톨 4-산화환원효소 유전자의 하나의 카피 및 NADPH-특이적 자일리톨 탈수소효소 유전자의 하나의 카피를 혼입시켰다. 상기 변형 균주는 이 때 아라비톨을 소비할 수 있다. 결과적으로, 글루코오스의 총 소비 후, 아라비톨 및 리비톨는 CNCM I-4982에 의해, 재소비되어, 더 많은 자일리톨이 생성된다.
피치아 오메리 CNCM I-4960(제3 세대) 및 CNCM I-4981(제4 세대)은 자일리톨 및 리비톨을 생성하나, 아라비톨은 더이상 생성하지 않는다. 자일룰로오스 및 자일리톨로의 아라비톨의 세포 내 전환은 브로쓰로의 아라비톨의 배출을 피하기에 충분하다. NAD+-특이적 D-아라비톨 산화환원효소 및 NADPH-특이적 자일리톨 탈수소효소를 인코딩하는 유전자의 더 많은 카피가 피치아 오메리에 도입될수록, 자일리톨의 역가, 수율 및 생산성은 더 크다.