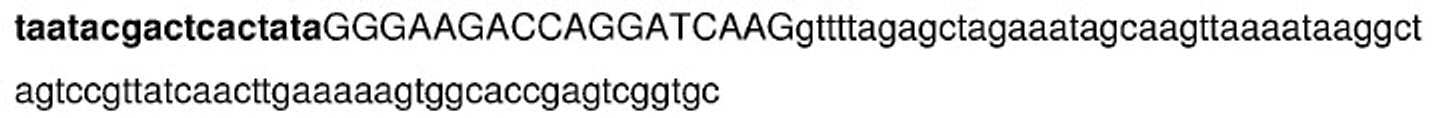본 개시는 진균 세포 내 게놈좌를 갖는 공여체 DNA의 상동성 재조합을 촉진하는데 사용되는 조성물과 방법을 포함한다. 이러한 방법은 원하는 표적 부위를 인식하여 이 부위에서 이중 가닥 절단 또는 닉을 도입하는 기능적 가이드 RNA/Cas 엔도뉴클레아제 복합체를 채용함으로써, 표적 부위에서 또는 표적 부위 근처에서 상동성 재조합을 촉진하고/촉진하거나 향상시킨다. 특정 양태에서, 진균 세포 내 표적 부위에서 비상동성 말단 결합(NHEJ) 메커니즘은 활성화되지 않거나, 비기능적이거나, 감소되는데, 본원에서 나타내길, 이는 원하는 상동성 재조합 이벤트의 효율을 개선한다.
본 조성물과 방법을 더욱 상세히 설명하기 전에, 본 조성물과 방법은 기재된 특별한 구현예에 한정되지 않으며, 이와 같이 당연히 변화될 수 있음을 이해하여야 한다. 또한, 본 조성물과 방법의 범위는 첨부된 청구항에 의해서만 한정될 것이기 때문에, 본원에서 사용된 용어는 특별한 구현예들만을 기재하기 위한 것이고, 한정하려는 의도가 아니라는 것을 이해하여야 한다.
값의 범위가 제공되는 경우, 문맥상 명확하게 달리 지시되지 않는다면, 이 범위의 상한치와 하한치 및 임의의 달리 언급된 값 또는 이 언급된 범위에서의 사이 값 사이에서 하한치의 단위의 1/10까지, 각각의 사이 값이 본 조성물 및 방법에 포함되는 것이 이해된다. 이들 더 작은 범위들의 상한치 및 하한치는 더 작은 범위에 독립적으로 포함될 수 있고, 이 언급된 범위에서 특별히 제외되는 임의의 제한을 받는 본 조성물 및 방법에도 포함된다. 언급된 범위가 제한들 중 하나 또는 양자 모두를 포함한다면, 이러한 포함된 제한들 중 하나 또는 둘 모두를 제외하는 범위들도 본 조성물 및 방법에 포함된다.
수치값 앞에 “약(about)”이라는 용어가 붙은 특정 범위들이 본원에 제시된다. “약”이라는 용어는 뒤에 놓인 정확한 숫자뿐만 아니라 그 용어의 뒤에 놓인 숫자와 거의 동일한 숫자 또는 근사적인 숫자에 대한 문자적 지원을 제공하기 위해 본원에 사용된다. 숫자가 구체적으로 언급된 숫자와 거의 동일한 숫자 또는 근사적인 숫자인지를 판단함에 있어서, 거의 동일하다고 언급되지 않은 숫자 또는 근사적이라고 언급되지 않은 숫자는 그것이 제시된 문맥에서, 구체적으로 언급된 숫자와 실질적으로 동일한 숫자일 수 있다. 예를 들어, 수치와 관련하여, 용어가 문맥에서 달리 구체적으로 정의되지 않으면, “약”이라는 용어는 이 수치의 -10% 내지 +10%의 범위를 지칭한다. 다른 예에서, pH 값이 구체적으로 달리 정의되지 않으면, “약 6의 pH 값”이라는 문구는 5.4 내지 6.6의 pH 값들을 지칭한다.
본원에 제공된 표제들은 전체적으로 본 명세서에 대한 참조로 가질 수 있는 본 조성물 및 방법의 다양한 양태 및 구현예의 제한은 아니다. 따라서, 바로 아래에 정의된 용어들은 전체적으로 명세서에 대한 참조로 보다 완전하게 정의된다.
본 문서는 읽기 쉽도록 수많은 부분들로 구성되지만, 독자는 하나의 부분에 있는 언급이 다른 부분에도 적용될 수 있다는 것을 이해할 것이다. 이런 방식으로, 본 개시의 다른 부분에 사용된 표제어는 한정하는 것으로 해석되어서는 안된다.
달리 정의되지 않으면, 본원에 사용된 기술적 및 과학적인 모든 용어들은 본 조성물 및 방법이 속하는 기술의 당업자가 일반적으로 이해하는 것과 동일한 의미를 가진다. 본원에 기재된 것과 유사하거나 동일한 임의의 방법들 및 물질들이 본 조성물 및 방법을 실시하거나 시험하는데 사용될 수 있지만, 대표적인 예시 방법 및 물질이 이제 기재된다.
본 명세서에서 언급된 모든 간행물과 특허는, 각 개별적인 간행물 또는 특허가 구체적으로 그리고 개별적으로 참조로 통합되도록 표시되듯이 그리고 이 간행물이 언급되는 것과 관련된 방법 및/또는 물질을 개시하고 기재하기 위해 본원에 참조로 통합되듯이, 본원에 참조로 통합된다. 임의 간행물의 인용은 출원일 전 이의 개시를 위한 것이고 본 조성물 및 방법이 선행 발명에 의해 이러한 간행물에 선행할 자격이 없다는 인정으로 해석되어서는 안된다. 또한, 제공된 공개일은 독립적인 확인이 필요할 수 있는 실제적인 공개일과 상이할 수 있다.
발명을 실시하기 위한 구체적인 내용에 따르면, 다음의 약어들 및 정의들이 적용된다. 문맥상 달리 명확하게 지시하지 않으면, 단수형 “a”, ”an”및 “the”는 복수의 대상들을 포함함에 주목하라. 따라서, 예를 들어, “한 효소(an enzyme)”에 대한 참조는 다수의 이러한 효소들을 포함하고, “상기 투여량(the dosage)”에 대한 참조는 하나 이상의 투여량 및 이 기술의 당업자에게 알려진 이의 등가량 등을 포함한다.
또한 주목할 점은 청구항이 임의의 선택적 요소를 제외하도록 작성될 수 있다는 것이다. 따라서, 이 진술은 청구항 요소의 인용 또는 "부정적인” 제한의 사용과 관련하여 "단독으로", "오직"등과 같은 배타적인 용어의 사용에 대한 선행 기준으로 작용하기 위한 것이다.
본 개시를 읽을 때 당업자에게 명백한 바와 같이, 본원에 기술되고 예시된 개별적인 구현예들 각각은 본원에 기재된 본 조성물 및 방법의 범위 또는 사상으로부터 벗어나지 않고서 다른 몇몇 구현예들 중 임의의 구현예의 특징과 쉽게 분리되거나 결합될 수 있는 개별 성분 및 특징을 갖는다. 언급된 임의의 방법은 열거된 사건의 순서로 또는 논리적으로 가능한 다른 순서로 수행될 수 있다.
정의
본원에 사용된, “Cas 엔도뉴클레아제” 또는 “Cas 엔도뉴클레아제 활성”을 갖는 것으로 언급되는 폴리펩티드는, 하나 이상의 가이드 폴리뉴클레오티드와 기능적으로 결합시, Cas 단백질이 표적 DNA 서열을 절단할 수 있는 Cas 유전자에 의해 인코딩된 CRISPR 관련(Cas) 폴리펩티드에 관한 것이다("CRISPR-Cas systems and methods for altering expression of gene products"라는 제목을 가진 미국 특허 제8697359 참조). 닉킹 엔도뉴클레아제 활성을 갖는 Cas 변이체들, 즉 이중 가닥 DNA 표적 부위에서 단일 가닥 닉을 도입하는 Cas 변이체들을 포함하여, 가이드 폴리뉴클레오티드 유도 엔도뉴클레아제 활성을 보유한 Cas 엔도뉴클레아제의 변이체들도 본 정의에 포함된다(아래의 정의 참조). (주목할 점은 날짜로 확인된 야생형 Cas 엔도뉴클레아제가 표적 부위에서 이중 가닥 절단을 도입한다는 것이다.) Cas 엔도뉴클레아제는, 예컨대 세포의 게놈 내 표적 부위에서 이중 가닥 DNA 내 특정 표적 부위를 인식하여 절단하는 가이드 폴리뉴클레오티드에 의해 안내된다. 몇몇 상이한 유형의 CRISPR-Cas 시스템이 기재되었고 유형 I, 유형 II 및 유형 III CRISPR-Cas 시스템으로 분류될 수 있다(예컨대, Liu and Fan, CRISPR-Cas system: a powerful tool for genome editing. Plant Mol Biol (2014) 85:209-218의 설명 참조). 특정 양태에서, CRISPR-Cas 시스템은 Cas9 엔도뉴클레아제 또는 이의 변이체(예컨대, Cas 닉카아제 포함)를 채용한 유형 II CRISPR-Cas 시스템이다. Cas9 엔도뉴클레아제는 다음 세균 종에서 Cas9 엔도뉴클레아제 및 이의 기능성 단편들을 포함하지만, 이들에 한정되지 않는 임의의 편리한 Cas9 엔도뉴클레아제일 수 있다: 스트렙토코쿠스 종(예컨대, S. 파이오제네스, S. 뮤탄스, 및 S. 더모필루스), 캄필로박터 종(예컨대, C. 제주니), 나이세리아 종(예컨대, N. 메닝기티데스), 프란시셀라 종(예컨대, F. 노비시다), 및 파스튜렐라 종(예컨대, P. 물토시다). Cas9의 수많은 기타 종들이 사용될 수 있다. 예를 들어, 서열 번호 45 및 서열 번호 48 내지 53 중 어느 하나와 적어도 70 %의 동일성을 갖는 아미노산 서열을 포함하는 기능성 Cas9 엔도뉴클레아제 또는 이의 변이체가 채용될 수 있고, 예컨대, 서열 번호 45 및 서열 번호 48 내지 53 중 어느 하나와 적어도 80 % 동일성, 적어도 90 % 동일성, 적어도 95 % 동일성, 96 % 이상의 동일성, 97 % 이상의 동일성, 98 % 이상의 동일성, 99 % 이상의 동일성 및 100 %까지의 동일성을 갖는 아미노산 서열을 포함하는 기능성 Cas9 엔도뉴클레아제 또는 이의 변이체가 사용될 수 있다. 다른 구현예에서, Cas 엔도뉴클레아제 또는 이의 변이체는 유형 II CRISPR-Cas 시스템의 Cpf1 엔도뉴클레아제이다. Cpf1은 Cas9과 다른 특징을 가진 견고한 DNA 간섭을 매개한다. Cpf1은 tracrRNA가 부족하여 T-풍부 프로토스페이서-인접 모티프를 이용한다. 이는 엇갈린 DNA 이중 가닥 절단을 통해 DNA를 절단한다. 예컨대, Zetsche 등, Cell (2015) 163:759-771 참조.
본원에서 사용된 바와 같이, "Cas 닉카아제"는, 하나 이상의 가이드 폴리뉴클레오티드와 기능적으로 결합시, 단일 가닥 닉을 표적인 이중 가닥 DNA 서열 내로 도입할 수 있는 Cas 엔도뉴클레아제이다. Cas 닉카아제는 (예컨대, 부위-특이적 돌연변이에 의해) 부모 Cas 엔도뉴클레아제 내 2개의 뉴클레아제 도메인 중 하나를 비활성화하여 재조합적으로 생성될 수 있다. Cas 닉카아제의 비제한적인 일 예로는 RuvC 도메인이 D10A 돌연변이에 의해 비활성화되는 Sander and Joung(Nature Biotechnology, 2013, 1-9)에 기재된 Cas9 닉카아제가 있다. 위에서 언급되었듯이, 일반적인 용어 “Cas 엔도뉴클레아제”는 이중 가닥 절단 및 니킹 Cas 폴리펩티드 둘 모두를 포함한다. 예컨대, 가이드 RNA가 Cas 엔도뉴클레아제를 원하는 표적 부위에 유도할 수 있는 것으로 기재되면, (아래에 정의된 것처럼) 이중 가닥 절단 Cas 엔도뉴클레아제 및 니킹 Cas 폴리펩티드 둘 모두에 대해 그렇게 할 것이다.
본원에 사용된 용어 "가이드 폴리뉴클레오티드"는 Cas 엔도뉴클레아제와 복합체를 형성할 수 있고, 이러한 Cas 엔도뉴클레아제가 DNA 표적 부위를 인식하고 선택적으로 절단할 수 있게 하는 폴리뉴클레오티드 서열에 관한 것이다. 가이드 폴리뉴클레오티드는 단일 분자 또는 이중 분자일 수 있다. 가이드 폴리뉴클레오티드 서열은 RNA 서열, DNA 서열, 또는 이의 조합(RNA-DNA 조합 서열)일 수 있다. 선택적으로, 이러한 가이드 폴리뉴클레오티드는 적어도 하나의 뉴클레오티드, 포스포디에스테르 결합 또는 연결 변형, 예컨대 고정 핵산(LNA), 5-메틸 dC, 2,6-디아미노퓨린, 2'-플루오로 A, 2'-플루오로 U, 2'-O-메틸 RNA, 포스포로티오에이트 결합, 콜레스테롤 분자에 대한 연결, 폴리에틸렌 글리콜 분자에 대한 연결, 스페이서 18(헥사에틸렌 글리콜 사슬) 분자에 대한 연결, 또는 고리화를 초래하는 5'에서 3'으로의 공유 연결을 포함할 수 있으나, 이에 한정되지 않는다. 단독으로 리보핵산을 포함하는 가이드 폴리뉴클레오티드는 "가이드 RNA"라고도 지칭된다.
가이드 폴리뉴클레오티드는 표적 DNA(아래에서 “포토스페이서” 또는 “표적 부위”로도 불림)의 뉴클레오티드 서열에 상보적인 제1 뉴클레오티드 서열 도메인(가변 표적화 도메인 또는 VT 도메인으로 지칭됨) 및 Cas 엔도뉴클레아제 폴리펩티드와 상호 작용하는 제2 뉴클레오티드 서열 도메인(Cas 엔도뉴클레아제 인식 도메인 또는 CER 도메인으로 지칭됨)을 포함하는 이중 분자(듀플렉스 가이드 폴리뉴클레오티드라고도 지칭됨)일 수 있다. 이중 분자 가이드 폴리뉴클레오티드의 CER 도메인은 상보성 영역을 따라 혼성화된 두 개의 별도의 분자들을 포함한다. 이러한 두 개의 별도의 분자들은 RNA, DNA, 및/또는 RNA-DNA- 조합 서열일 수 있다. 일부 구현예에서, CER 도메인에 연결된 VT 도메인을 포함하는 듀플렉스 가이드 폴리뉴클레오티드의 제1 분자는 "crDNA"(연속된 DNA 뉴클레오티드들로 구성되는 경우) 또는 "crRNA"(연속된 RNA 뉴클레오티드들로 구성되는 경우), 또는 "crDNA-RNA"(DNA와 RNA 뉴클레오티드의 조합으로 구성되는 경우)라 지칭된다. cr뉴클레오티드는 박테리아 및 고세균류에서 자연적으로 발생하는 crRNA의 단편을 포함할 수 있다. 일 구현예에서, 박테리아 및 고세균류에서 자연적으로 발생하고 본원에 기재된 cr뉴클레오티드에 존재하는 crRNA의 단편의 크기는 2, 3, 4, 5, 6, 7, 8, 9,10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20개 이상의 뉴클레오티드 범위를 가질 수 있지만, 이들 범위에 제한되지 않는다. 일부 구현예에서, CER 도메인을 포함하는 듀플렉스 가이드 폴리뉴클레오티드의 제2 분자는 "tracrRNA"(연속된 RNA 뉴클레오티드들로 구성되는 경우) 또는 "tracrDNA"(연속된 DNA 뉴클레오티드들로 구성되는 경우) 또는 "tracrDNA-RNA"(DNA와 RNA 뉴클레오티드의 조합으로 구성되는 경우)라 지칭된다. 특정 구현예에서, RNA/Cas9 엔도뉴클레아제 복합체를 가이드하는 RNA는 듀플렉스 crRNA-tracrRNA를 포함하는 듀플렉스화된 RNA이다.
또한, 이러한 가이드 폴리뉴클레오티드는 표적 DNA의 뉴클레오티드 서열에 상보적인 (가변 표적화 도메인 또는 VT 도메인으로 지칭되는) 제1 뉴클레오티드 서열 도메인 및 Cas 엔도뉴클레아제 폴리펩티드와 상호 작용하는 (Cas 엔도뉴클레아제 인식 도메인 또는 CER 도메인으로 지칭되는) 제2 뉴클레오티드 도메인을 포함하는 단일 분자일 수 있다. "도메인"은 RNA, DNA, 및/또는 RNA-DNA-조합 서열일 수 있는 뉴클레오티드들이 연속되어 있는 것을 의미한다. 단일 가이드 폴리뉴클레오티드의 VT 도메인 및/또는 CER 도메인은 RNA 서열, DNA 서열, 또는 RNA-DNA-조합 서열을 포함할 수 있다. 일부 구현예에서, 이러한 단일 가이드 폴리뉴클레오티드는 (CER 도메인을 포함하는) tracr뉴클레오티드에 연결된 (CER 도메인에 연결된 VT 도메인을 포함하는) cr뉴클레오티드를 포함하며, 이때, 이러한 연결은 RNA 서열, DNA 서열, 또는 RNA-DNA 조합 서열을 포함하는 뉴클레오티드 서열이다. cr뉴클레오티드 및 tracr뉴클레오티드로부터의 서열들로 구성되는 단일 가이드 폴리뉴클레오티드는 (연속된 RNA 뉴클레오티드들로 구성되는 경우) "단일 가이드 RNA" 또는 (연속된 DNA 뉴클레오티드들로 구성되는 경우) "단일 가이드 DNA" 또는 (RNA와 DNA 뉴클레오티드의 조합으로 구성되는 경우) "단일 가이드 RNA-DNA"라 지칭될 수 있다. 본 개시의 일 구현예에서, 단일 가이드 RNA는 유형 II Cas 엔도뉴클레아제와 복합체를 형성할 수 있는 유형 II CRISPR/Cas 시스템의 crRNA 또는 crRNA 단편 및 tracrRNA 또는 tracrRNA 단편을 포함하고, 여기서 가이드 RNA/Cas 엔도뉴클레아제 복합체는 Cas 엔도뉴클레아제를 진균 세포 게놈 표적 부위에 유도하여, Cas 엔도뉴클레아제가 게놈 타겟 부위 내로 이중 가닥 절단을 도입할 수 있게 한다.
단일 가이드 폴리뉴클레오티드 대 듀플렉스 가이드 폴리뉴클레오티드를 사용하는 일 양태는 하나의 발현 카세트만이 표적 세포 내에서 단일 가이드 폴리뉴클레오티드를 발현하기 위해 만들어질 필요가 있다는 것이다.
용어 "가변 표적화 도메인" 또는 "VT 도메인"은 본원에서 상호교환적으로 사용되며, 이중 가닥 DNA 표적 부위의 한 가닥(뉴클레오티드 서열)에 상보적인 뉴클레오티드 서열을 포함한다. 제1 뉴클레오티드 서열 도메인(VT 도메인)과 표적 서열 사이의 백분율 보완성은 적어도 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 63%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100% 상보적이다. VT 도메인의 길이는 적어도 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 또는 30개 뉴클레오티드 길이일 수 있다. 일부 구현예에서, 이러한 VT 도메인은 연속된 12개 내지 30개의 뉴클레오티드를 포함한다. VT 도메인은 DNA 서열, RNA 서열, 변형된 DNA 서열, 변형된 RNA 서열, 또는 이들의 임의 조합으로 구성될 수 있다.
용어 가이드 폴리뉴클레오티드의 "Cas 엔도뉴클레아제 인식 도메인" 또는 "CER 도메인"은 본원에서 상호교환적으로 사용되며, Cas 엔도뉴클레아제 폴리펩티드와 상호작용하는 뉴클레오티드 서열(예컨대, 가이드 폴리뉴클레오티드의 제2 뉴클레오티드 서열 도메인)을 포함한다. CER 도메인은 DNA 서열, RNA 서열, 변형된 DNA 서열, 변형된 RNA 서열(예컨대, 본원에 기술된 변형 참조), 또는 이의 임의의 조합으로 구성될 수 있다.
단일 가이드 폴리뉴클레오티드의 cr뉴클레오티드 및 tracr뉴클레오티드를 연결하는 뉴클레오티드 서열은 RNA 서열, DNA 서열, 또는 RNA-DNA 조합 서열을 포함할 수 있다. 일 구현예에서, 단일 가이드 폴리뉴클레오티드의 cr뉴클레오티드 및 tracr뉴클레오티드를 연결하는 뉴클레오티드 서열은 그 길이가 적어도 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99 또는 100 개의 뉴클레오티드일 수 있다. 다른 구현예에서, 단일 가이드 폴리뉴클레오티드의 cr뉴클레오티드 및 tracr뉴클레오티드를 연결하는 뉴클레오티드 서열은 테트라루프 서열, 예컨대 GAAA 테트라루프 서열을 포함할 수 있지만, 이에 한정되지는 않는다.
가이드 뉴클레오티드, VT 도메인 및/또는 CER 도메인의 뉴클레오티드 서열 변형은 5’ 캡, 3’ 폴리아데닐화 테일, 리보스위치 서열, 안정성 조절 서열, dsRNA 듀플렉스를 형성하는 서열, 가이드 폴리뉴클레오티드를 세포내 위치에 표적화하는 변형 또는 서열, 트랙킹을 제공하는 변형 또는 서열, 단백질을 위한 결합 부위를 제공하는 변형 또는 서열, 잠김 핵산(Locked Nucleic Acid(LNA)), 5- 메틸 dC 뉴클레오티드, 2,6- 디아미노퓨린 뉴클레오티드, 2'- 플루오로 A 뉴클레오티드, 2'- 플루오로 U 뉴클레오티드; 2’-O-메틸 RNA 뉴클레오티드, 포스포로티오에이트 결합, 콜레스테롤 분자에 대한 연결, 폴리에틸렌 글리콜 분자에 대한 연결, 스페이서 18 분자에 대한 연결, 5 '내지 3'공유 결합, 또는 이들의 임의의 조합으로 이루어진 군으로부터 선택될 수 있지만, 이들에 한정되지는 않는다. 이러한 변형은 적어도 하나의 추가적인 유익한 특징을 초래할 수 있고, 여기서 추가적인 유익한 특징은 변형 또는 조절된 안정성, 세포 내 표적화, 트래킹, 형광 표지, 단백질 또는 단백질 복합체에 대한 결합 부위, 상보적인 표적 서열에 대한 변형된 결합 친화도, 세포 분해에 대한 변형된 저항성, 및 증가된 세포 투과성의 군으로부터 선택된다.
본원에서 사용된 용어 "가이드 폴리뉴클레오티드/카스 엔도뉴클레아제 시스템”(및 등가물)은 DNA 표적 부위에서 이중 가닥 절단을 도입할 수 있는 Cas 엔도뉴클레아제 및 가이드 폴리뉴클레오티드 (단일 또는 이중)의 복합체를 포함한다. Cas 엔도뉴클레아제는 DNA 표적 부위에 아주 근접하여 DNA 듀플렉스를 풀어주고, 가이드 RNA가 표적 서열을 인식할 때 DNA 가닥 둘 모두를 절단하지만, 올바른 프로토스페이서-인접 모티프(PAM)가 표적 서열의 3’ 말단에서 적절하게 배향되는 경우에만 그렇게 한다.
용어 "기능적 단편", "기능적으로 동등한 단편(fragment that is functionally equivalent)", "기능적으로 동등한 단편(functionally equivalent fragment)” 등은 상호 교환적으로 사용되며, 부모 폴리펩티드의 정량적 효소 활성을 유지하는 부모 폴리펩티드의 일부 또는 부서열을 지칭한다. 예컨대, Cas 엔도뉴클레아제의 기능적 단편은 가이드 폴리뉴클레오티드와 함께 이중 가닥 절단을 생성하는 능력을 유지한다. 여기서 주목할 점은 기능적 단편이 부모 폴리펩티드에 비해 변경된 양론적 효소 활성을 가질 수 있다는 것이다.
용어 "기능적 변이체(functional variant)", "기능적으로 동등한 변이체(variant that is functionally equivalent)", "기능적으로 동등한 변이체(functionally equivalent variant)" 등은 상호 교환적으로 사용되며 부모 폴리펩티드의 정량적 효소 활성을 유지하는 부모 폴리펩티드의 변이체를 지칭한다. 예컨대, Cas 엔도뉴클레아제의 기능적 변이체는 가이드 폴리뉴클레오티드와 함께 (대상 변이체에 따라) 이중 가닥 절단 또는 닉을 생성하는 능력을 유지한다. 여기서 주목할 점은 기능적 단편이 부모 폴리펩티드에 비해 변경된 양론적 효소 활성을 가질 수 있다는 것이다.
단편 및 변이체는 부위-특이적 돌연변이 및 합성 구조를 포함하는 임의의 편리한 방법으로 수득될 수 있다.
진균 세포에 적용시 용어 “게놈”은 핵 내에서 발견되는 염색체 DNA뿐만 아니라 세포의 세포 내 성분(예컨대, 미토콘드리아) 내에서 발견되는 세포소기관 DNA를 포함한다.
“코돈 변형 유전자” 또는 “코돈-선호 유전자” 또는 “코돈-최적화 유전자”는 숙주 세포의 선호되는 코돈 사용 빈도를 모방하도록 설계된 코돈 사용 빈도를 갖는 유전자이다. 유전자를 코돈-최적화하도록 이루어진 핵산 변경은 "동의어"이며, 이는 이들이 부모 유전자의 인코딩된 폴리펩티드의 아미노산 서열을 변경시키지 않는다는 것을 의미한다. 그러나, 고유 유전자 및 변이체 유전자 모두는 특정 숙주 세포에 대해 코돈-최적화될 수 있으며, 이와 관련하여 이와 같은 제한은 의도되지 않는다.
“코딩 서열”은 특정 아미노산 서열을 코딩하는 폴리뉴클레오티드 서열을 지칭한다. "조절 서열"은 코딩 서열의 상류(5 '비코딩 서열), 내 또는 하류(3’ 비코딩 서열)에 위치하며, 관련 코딩 서열의 전사, RNA 가공 또는 안정성, 또는 번역에 영향을 주는 뉴클레오티드 서열을 지칭한다. 조절 서열은 프로모터, 번역 리더 서열, 5’ 미번역 서열, 3’ 미번역 서열, 인트론, 폴리아데닐화 표적 서열, RNA 가공 부위, 이펙터 결합 부위, 및 스템-루프(stem-loop) 구조를 포함할 수 있지만, 이에 한정되지는 않는다.
“프로모터”는 코딩 서열 또는 기능적 RNA의 발현을 조절할 수 있는 DNA 서열을 지칭한다. 프로모터 서열은 근접 상류 요소 및 더 먼 상류 요소로 구성되고, 더 먼 상류 요소는 종종 인헨서(enhancer)로 지칭된다. “인헨서”는 프로모터 활성을 자극할 수 있는 DNA 서열이고, 프로모터의 선천적인 요소 또는 프로모터의 수준 또는 조직 특이성을 향상시키기 위해 삽입된 이종 요소일 수 있다. 프로모터는 고유 유전자로부터 그 전체가 유래될 수도 있거나, 자연에서 발견되는 상이한 프로모터들로부터 유래된 상이한 요소들로 구성될 수 있거나, 심지어 합성 DNA 세그먼트를 포함할 수 있다. 상이한 프로모터가 상이한 조직 또는 세포 유형으로, 또는 발달의 상이한 단계에서 또는 상이한 환경 조건에 반응하여 유전자의 발현을 유도할 수 있음이 당업자에게 이해된다. 또한, 대부분의 경우, 조절 서열의 정확한 경계가 완전히 정의되지 않았기 때문에, 일부 변이체의 DNA 단편이 동일한 프로모터 활성을 가질 수 있음이 또한 인식된다. 이 기술에서 잘 알려진 것처럼, 프로모터들은 이들의 강도 및/또는 이들이 활성인 조건, 예컨대 항시성(constitutive) 프로모터, 강한 프로모터, 약한 프로모터, 유도성/억제성 프로모터, 조직 특이적/발달 조절 프로모터, 세포주기 의존성 프로모터 등에 따라 분류될 수 있다.
“RNA 전사물”은 DNA 서열의 RNA 중합효소-효소화 전사로부터 만들어지는 생성물을 지칭한다. “메신저 RNA” 또는 “mRNA”는 인트론이 없고 세포에 의해 단백질로 번역될 수 있는 RNA를 지칭한다. “cRNA”는 역전사 효소를 이용하는 mRNA 템플릿에 상보적이고 그로부터 합성되는 DNA를 지칭한다. “센스” RNA는 mRNA를 포함하고 세포 내 또는 시험관 내에서 단백질로 번역될 수 있는 RNA 전사물을 지칭한다. “안티센스 RNA”는 표적 1차 전사물 또는 mRNA의 전부 또는 일부에게 상보적이고, 특정 조건 하에서, 표적 유전자의 발현을 차단하는 RNA 전사물을 지칭한다(미국 특허 제5,107,065호 참조). 안티센스 RNA의 상보성은 특정 유전자 전사물의 임의의 부분, 즉 5’ 비-코딩 서열, 3’ 비-코딩 서열, 인트론 또는 코딩 서열과 함께 있을 수 있다. "기능적 RNA"는 폴리펩티드로 번역되지 않을 수 있지만 세포 과정에 영향을 미치는 안티센스 RNA, 리보자임 RNA 또는 기타 RNA를 지칭한다. 용어 "보체(complement)” 및 "역 보체(reverse complement)"는 mRNA 전사체에 대하여 본원에서 상호 교환적으로 사용되며, 메시지의 안티센스 RNA를 정의하기 위한 의미이다.
본원에 사용된, "기능적으로 부착된” 또는 "작동 가능하게 연결된"은 프로모터, 인헨서 부위, 종결인자, 신호 서열, 에피토프 태그 등과 같이 공지된 또는 원하는 활성을 갖는 폴리펩티드 또는 폴리뉴클레오티드 서열의 조절 영역 또는 기능적 도메인이, 조절 영역 또는 기능적 도메인이 그의 공지된 또는 원하는 활성에 따라 그 표적의 발현, 분비 또는 기능을 조절할 수 있는 방식으로 표적(예컨대, 유전자 또는 폴리펩티드)에 부착되거나 연결된다는 것을 의미한다. 예를 들어, 프로모터는, 코딩 서열의 발현을 조절할 수 있는 경우(즉, 코딩 서열이 프로모터의 전사 조절 하에 있을 때), 코딩 서열과 작동 가능하게 연결된다.
본원에서 사용된 표준 재조합 DNA 및 분자 복제 기술은 당업계에 공지되어 있다.
"PCR” 또는 "중합 효소 연쇄 반응(polymerase chain reaction)"은 특정 DNA 단편의 합성을 위한 기술이며 일련의 반복적인 변성, 어닐링 및 확장 사이클로 구성되며 당업계에 잘 알려져 있다.
용어 "재조합물"은, 생물학적 성분 또는 조성물(예컨대, 세포, 핵산, 폴리펩티드/효소, 벡터 등)과 관련하여 사용되는 경우, 생물학적 성분 또는 조성물이 자연에서 발견되지 않는 상태로 있다는 것을 나타낸다. 다시 말해서, 생물학적 성분 또는 조성물은 이의 자연 상태로부터 인간 개입에 의해 변형되었다. 예를 들어, 재조합 세포는 이의 천연 부모(즉, 비-재조합) 세포에서 발견되지 않는 하나 이상의 유전자를 발현하는 세포, 이의 천연 부모 세포와 상이한 양으로 하나 이상의 천연 유전자를 발현하는 세포, 및/또는 천연 부모 세포와 상이한 조건 하에서 하나 이상의 천연 유전자를 발현하는 세포를 포함한다. 재조합 핵산은 하나 이상의 뉴클레오티드에 의해 천연 서열과 다를 수 있으며, 이종 서열(예컨대, 이종 프로모터, 비-천연 또는 변이체 신호 서열을 인코딩하는 서열 등)에 작동 가능하게 연결될 수 있고, 인트론 서열이 없을 수 있고/있거나 단리된 형태로 존재할 수 있다. 재조합 폴리펩티드/효소는 하나 이상의 아미노산에 의해 천연 서열과 상이할 수 있고, 이종 서열과 융합될 수 있고, 절단될 수 있거나 아미노산의 내부 결실을 가질 수 있으며, 천연 세포에서 발견되지 않는 방식으로 발현 될 수 있고/있거나(예컨대, 폴리펩티드를 인코딩하는 발현 벡터의 세포 내 존재로 인해 폴리펩티드를 과발현시키는 재조합 세포로부터), 단리된 형태로 존재할 수 있다. 일부 구현예에서, 재조합 폴리뉴클레오티드 또는 폴리펩티드/효소는 이의 야생형 대응물과 동일하지만 (예를 들어, 단리된 또는 농축된 형태의) 비천연 형태로 존재하는 서열을 갖는다는 것이 강조된다.
용어 "플라스미드", "벡터” 및 "카세트"는 관심 폴리뉴클레오티드 서열, 예컨대 세포에서 발현되는 관심 유전자("발현 벡터” 또는 "발현 카세트")를 운반하는 여분의 염색체 요소를 지칭한다. 이러한 요소는 일반적으로 이중 가닥 DNA의 형태를 가지며, 임의의 근원으로부터 유래된 단일 또는 이중 가닥 DNA 또는 RNA의 서열, 게놈 통합 서열, 파지 또는 뉴클레오타이드 서열을 선형 또는 원형 형태로 자동으로 복제할 수 있고, 여기서 다수의 뉴클레오티드 서열이 세포 내로 관심 폴리뉴클레오티드를 도입할 수 있는 독특한 구조로 결합되거나 재조합되었다. 관심 폴리뉴클레오티드 서열은 표적 세포 내에서 발현될 폴리펩티드 또는 기능적 RNA를 인코딩하는 유전자일 수 있다. 발현 카세트/벡터는 일반적으로 숙주 세포 내에서 그 유전자의 발현을 허용하는 작동 가능하게 연결된 요소를 가진 유전자를 포함한다.
본원에서 사용된 용어 “발현”은 전구체 또는 성숙한 형태의 기능적 최종 생성물(예컨대, mRNA, 가이드 RNA 또는 단백질)의 생산을 지칭한다.
폴리뉴클레오티드 또는 폴리펩티드를 세포(예컨대, 재조합 DNA 작제물/발현 작제물)에 삽입하는 문맥에서 "도입된(introduced)"은 이러한 과제 수행을 위한 임의의 방법을 지칭하며, 원하는 생체 분자의 도입을 달성하기 위한 "형질감염", "형질전환", "형질도입", 물리적 수단 등의 임의 수단을 포함한다.
"일시적으로 도입된(introduced transiently)", "일시적으로 도입된(transiently introduced)", "일시적 도입", "일시적으로 발현하는” 등은 비영구적 방식으로 숙주 세포(또는 숙주 세포군)에 생체 분자가 도입되는 것을 의미한다. 이중 가닥 DNA와 관련하여, 일시적인 도입은 도입된 DNA가 숙주 세포의 염색체 내로 통합되지 않아 성장 중에 모든 딸 세포에 전달되지 않는 상황뿐만 아니라 도입되어 염색체로 통합될 수 있었던 DNA 분자가 임의의 편리한 방법(예를 들어, cre-lox 시스템을 채용하고, 에피솜 DNA 작제물에 대한 양성의 선택적인 압력을 제거하고, 선택 매질 등을 사용하여 염색체로부터의 통합된 폴리뉴클레오티드의 전부 또는 일부로부터 루핑 (looping)을 촉진시킴으로써)을 사용하여 원하는 시간에 제거되는 상황을 포함한다. 이 점에 대한 제한은 없다. 일반적으로, 숙주 세포에 RNA(예컨대, 가이드 RNA, 메신저 RNA, 리보자임 등) 또는 폴리펩티드(예컨대. Cas 폴리펩티드)를 도입하는 것은 이러한 생체 분자들이 세포 성장 동안 복제되지 않고 무한히 딸 세포들에게 전달된다는 점에서 일시적인 것으로 간주된다. Cas/가이드 RNA 복합체와 관련하여 일시적인 도입은, 두 가지 생체 분자가 표적화된 Cas 엔도뉴클레아제 활성을 발휘하는 데 필요하므로, 어느 한 구성 요소가 일시적으로 도입되는 상황을 다룬다. 따라서, Cas/가이드 RNA 복합체의 일시적 도입은 Cas 엔도뉴클레아제 및 가이드 RNA 중 어느 하나 또는 둘 모두가 일시적으로 도입되는 구현예들을 포함한다. 예를 들어, 기능적 복합체는 일시적 방식으로 숙주 세포에 존재하기 때문에 가이드 RNA가 일시적으로 도입되는 Cas 엔도뉴클레아제를 위한 게놈-통합 발현 카세트(따라서 일시적으로 도입되지 않음)를 갖는 숙주 세포는 일시적으로 도입된 Cas/가이드 RNA 복합체 (또는 시스템)를 가지고 있다고 말할 수 있다. 특정 구현예에서, 상기 도입 단계는: (i) Cas 엔도뉴클레아제를 안정적으로 발현하는 부모 진균 세포군을 수득하는 단계, 및 (ii) 가이드 RNA를 상기 부모 진균 세포군에 일시적으로 도입하는 단계를 포함한다. 역으로, 상기 도입 단계는: (i) 가이드 RNA를 안정적으로 발현하는 부모 진균 세포군을 수득하는 단계, 및 (ii) Cas 엔도뉴클레아제를 상기 부모 진균 세포군에 일시적으로 도입하는 단계를 포함한다.
“성숙한” 단백질은 번역후 가공된 폴리펩티드(즉, 1차 번역 생성물에 존재하는 임의의 프리폴리펩티드 또는 프로폴리펩티드가 제거된 것)를 지칭한다. “전구체” 단백질은 mRNA의 번역의 1차 생성물(즉, 프리펩티드 및 프로펩티드가 여전히 존재하는)을 지칭한다. 프리펩티드 및 프로펩티드는 세포내 위치 신호일 수 있지만, 이에 한정되지는 않는다.
"안정한 형질전환"은 핵 및 세포소기관 게놈 둘 모두를 포함하여 숙주 생물의 게놈으로 핵산 단편을 이동시켜 유전학적으로 안정한 유전을 수득하는 것을 지칭한다(수득된 숙주 세포는 때로는 본원에서 "안정한 형질전환체"라고도 함). 대조적으로, "일시적 형질전환"은 숙주 유기체의 핵산 또는 다른 DNA- 함유 세포소기관으로 핵산 단편을 옮겨서 통합 또는 안정한 유전없이 유전자 발현을 유도하는 것을 의미한다(때때로 본원에서 "불안정한 형질전환"으로 지칭되고, 수득된 숙주 세포는 때때로 본원에서 "불안정한 형질전환체"라 지칭된다). 형질전환된 핵산 단편을 포함하는 숙주 생물은 “형질전환 생물”로 지칭된다.
본원에서 사용된 "진균 세포", "곰팡이", "진균 숙주 세포” 등은 필라 아스코마이코타(phyla Ascomycota), 바시디오마이코타(Basidiomycota), 키트리디오마이코타(Chytridiomycota), 및 자이고마이코타(Zygomycota)(Hawksworth 등, In, Ainsworth and Bisby 's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK에서 정의됨)뿐만 아니라 오오마이코타(Oomycota)(Hawksworth 등, supra에서 인용) 및 모든 불완전 균류(mitosporic fungi)(Hawksworth 등, supra)를 포함한다. 특정 구현예에서, 진균 세포는 효모 세포이고, 여기서 “효모”는 자낭홀씨생산 효모(Endomyceales), 담자균류 효모, 및 불완전 균류(Fungi Imperfecti)(Blastomycetes)에 속하는 효모를 의미한다. 이와 같이, 효모 숙주 세포는 칸디다(Candida), 한세누라(Hansenula), 클루베로마이세스(Kluyveromyces), 피치아(Pichia), 사카로마이세스(Saccharomyces), 스키나사카로마이세스(Schizosaccharomyces), 또는 야로위아 세포(Yarrowia cell)를 포함한다. 효모 종은 다음을 포함하지만 이에 한정되지 않는다: 사카로마이세스 칼스베르겐시스(Saccharomyces carlsbergensis), 사카로마이세스 세레비시애(Saccharomyces cerevisiae), 사카로마이세스 디아스타티쿠스(Saccharomyces diastaticus), 사카로마이세스 도우글라시(Saccharomyces douglasii), 사카로마이세스 클루베리(Saccharomyces kluyveri), 사카로마이세스 노르벤시스(Saccharomyces norbensis), 사카로마이세스 오비포르미스(Saccharomyces oviformis), 클루베로마이세스 락티스(Kluyveromyces lactis), 및 야로위아 리폴리티카(Yarrowia lipolytica) 세포.
용어 “사상균 세포”는 아문 유마이코티나(Eumycotina) 또는 페치조마이코티나(Pezizomycotina)의 모든 사상 형태를 포함한다. 사상균 속의 적합한 세포들은 아크레모늄(Acremonium), 아스퍼질러스(Aspergillus), 크리소스포리움(Chrysosporium), 코리나스쿠스(Corynascus), 채토미움(Chaetomium), 푸사리움(Fusarium), 기베렐라(Gibberella), 후미콜라(Humicola), 마그나포르테(Magnaporthe), 마이셀리오프토라(Myceliophthora), 뉴로스포라(Neurospora), 패시로마이세스(Paecilomyces), 페니실리움(Penicillium), 스키탈리디움(Scytaldium), 타라로마이세스(Talaromyces), 더모아스쿠스(Thermoascus), 티에라비아(Thielavia), 토리포클라디움(Tolypocladium), 하이포크레아(Hypocrea), 및 트리코데르마의 세포들을 포함하지만, 이들에 한정되지는 않는다.
사상균 종의 적합한 세포들은 아스퍼질러스 아와모리(Aspergillus awamori), 아스퍼질러스 푸미가투스(Aspergillus fumigatus), 아스퍼질러스 포에티두스(Aspergillus foetidus), 아스퍼질러스 자포니쿠스(Aspergillus japonicus), 아스퍼질러스 니두란스(Aspergillus nidulans), 아스퍼질러스 나이거(Aspergillus niger), 아스퍼질러스 오리재(Aspergillus oryzae), 크리소스포리움 루크노웬스(Chrysosporium lucknowense), 푸사리움 박트리디오데스(Fusarium bactridioides), 푸사리움 세레아리스(Fusarium cerealis), 푸사리움 크룩웰렌스(Fusarium crookwellense), 푸사리움 쿨모룸(Fusarium culmorum), 푸사리움 그라미네아룸(Fusarium graminearum), 푸사리움 그라미눔(Fusarium graminum), 푸사리움 헤테로스포룸(Fusarium heterosporum), 푸사리움 네군디(Fusarium negundi), 푸사리움 옥시스포룸(Fusarium oxysporum), 푸사리움 렉티쿠라툼(Fusarium reticulatum), 푸사리움 로세움(Fusarium roseum), 푸사리움 삼부시눔(Fusarium sambucinum), 푸사리움 사르코크로움(Fusarium sarcochroum), 푸사리움 스포로트리키오이데스(Fusarium sporotrichioides), 푸사리움 술푸레움(Fusarium sulphureum), 푸사리움 토루로숨(Fusarium torulosum), 푸사리움 크리코테시오데스(Fusarium trichothecioides), 푸사리움 베네나툼(Fusarium venenatum), 후미콜라 인솔렌스(Humicola insolens), 후미콜라 라누지노사(Humicola lanuginosa), 하이포크레아 제코리나(Hypocrea jecorina), 마이셀리오프토라 더모필라(Myceliophthora thermophila), 뉴로스포라 크라싸(Neurospora crassa), 뉴로스포라 인테르메디아(Neurospora intermedia), 페니실리움 푸르푸로게눔(Penicillium purpurogenum), 페니실리움 카네센스(Penicillium canescens), 페니실리움 소리툼(Penicillium solitum), 페니실리움 푸니쿠로숨(Penicillium funiculosum), 파네로채테 크리소스포리움(Phanerochaete chrysosporium), 타라로마이세스 플라부스(Talaromyces flavus), 티에라비아 테레스트리스(Thielavia terrestris), 트리코데르마 하르지아눔(Trichoderma harzianum), 트리코데르마 코닌지(Trichoderma koningii), 트리코데르마 롱지브라키아툼(Trichoderma longibrachiatum), 트리코데르마 레에세이(Trichoderma reesei), 및 트리코데르마 비리데(Trichoderma viride)를 포함하지만, 이들에 한정되지 않는다.
용어 "표적 부위", "표적 서열", "게놈 표적 부위", "게놈 표적 서열” (및 등가물)은 본원에서 상호 교환 적으로 사용되며, Cas 엔도뉴클레아제 절단이 게놈 변형, 예컨대 공여체 DNA와의 상동성 재조합을 촉진시키도록 요구되는 진균 세포의 게놈 내 폴리뉴클레오티드 서열을 지칭한다. 그러나, 본 용어가 사용되는 문맥은 이의 의미가 약간 변경될 수 있다. 예를 들어, Cas 엔도뉴클레아제의 표적 부위는 일반적으로 매우 특이적이고 종종 정확한 뉴클레오타이드 서열/위치로 정의될 수 있는 반면, 일부 경우에는 원하는 게놈 변형을 위한 표적 부위가 단지 DNA 절단이 일어나는 부위, 예컨대, 상동성 재조합이 요구되는 게놈좌 또는 영역보다 더 광범위하게 정의될 수 있다. 따라서, 특정 경우에서, Cas/가이드 RNA DNA 절단의 활성을 통하여 일어나는 게놈 변형은 표적 부위”에서” 또는 표적 부위 “근처에서” 일어나는 것으로 기재된다. 표적 부위는 진균 세포 게놈 내의 내인성 부위일 수 있거나 또는 대안적으로 표적 부위는 진균 세포에 이종일 수 있어서 게놈에서 자연 발생하지 않을 수 있거나 또는 자연에서 일어나는 경우에 비해 표적 부위가 이종 게놈 위치에서 발견될 수 있다. 특정한 다른 경우에서, 공여체 DNA가 진균 세포의 게놈좌에 대한 상동성을 가진 도메인을 포함할 때, 진균 세포에 도입된 Cas 엔도뉴클레아제 및 가이드 RNA는 Cas 엔도뉴클레아제가 진균 세포의 게놈좌 내 또는 진균 세포의 게놈좌 근처의 표적 부위에서 작용할 수 있는 복합체를 형성할 수 있다. 일부 구현예에서, 게놈 DNA 위의 Cas 엔도뉴클레아제 절단 부위(또는 표적 부위)는 공여체 DNA 및 게놈좌 사이의 상동성 영역 내에 있고, 여기서 상동성 재조합이 일어날 수 있다. 다른 구현예에서, 절단 부위는 상동성 영역으로부터 약 1 bp 내지 약 10 kb 떨어져 있을 수 있는(예컨대, 상동성 영역의 부위로부터 1 bp, 2 bp, 5 bp, 10 bp, 20 bp, 50 bp, 100 bp, 250 bp, 500 bp, 1 kb, 2 kb, 3 kb, 4 kb, 5 kb, 6 kb, 7 kb, 8 kb, 9 kb 또는 10 kb 떨어진) 공여체 DNA와 게놈좌 사이의 상동성 영역 근처에 있다.
본원에서 사용된 “핵산”은 뉴클레오티드를 의미하고 데옥시리보뉴클레오티드 또는 리보뉴클레오티드 염기들의 단일 가닥 폴리머 또는 이중 가닥 폴리머를 포함한다. 또한 핵산은 단편 및 변형된 뉴클레오티드를 포함할 수 있다. 따라서, 용어 "폴리뉴클레오티드", "핵산 서열", "뉴클레오티드 서열” 및 "핵산 단편"은 단일 또는 이중 가닥인 RNA 및/또는 DNA의 폴리머를 나타내기 위해 교환 가능하게 사용되며, 선택적으로 합성, 비-천연 또는 변경된 뉴클레오티드 염기를 포함한다. (보통 이들의 5’-모노포스페이트 형태로 발견되는) 뉴클레오티드는 다음과 같이 이들의 단일 문자 표시에 의해 지칭된다: “A”는 아데노신 또는 데옥시아데노신(각각 RNA 또는 DNA에 대하여), "C"는 시아노 또는 데옥시시토신, "G"는 구아노신 또는 데옥시구아노신, "U"는 우리딘, "T"는 데옥시티미딘, "R"은 푸린 (A 또는 G), “Y”는 피리미딘 (C 또는 T), “K”는 G 또는 T, “H”는 A 또는 C 또는 T, “I”는 이노신, 및 “N”은 임의의 뉴클레오티드.
용어 “에서 유래된”은 용어 “에서 기원하는”, “에서 수득되는”, “에서 수득 가능한”, “에서 단리된”, 및 “에서 생성된”을 포함하고, 일반적으로 하나의 특정된 물질이 이의 기원을 다른 특정된 물질에서 발견하거나 다른 특정된 물질을 참조하여 기재될 수 있는 특징들을 가진다는 것을 나타낸다.
본원에서 사용된 용어 “혼성화 조건”은 혼성화 반응이 수행되는 조건을 지칭한다. 이들 조건들은 전형적으로 혼성화가 측정되는 조건의 “엄격도” 정도에 의해 분류된다. 엄격도의 정도는, 예컨대 핵산 결합 복합체 또는 프로브의 용융 온도(Tm)에 기초할 수 있다. 예를 들어, “최대 엄격도”는 전형적으로 약 Tm - 5℃(프로브의 Tm 아래의 5℃)에서 발생하고; “높은 엄격도”은 Tm 아래의 약 5~10℃에서 발생하고; “중간 엄격도”는 프로브의 Tm 아래의 약 10~20℃에서 발생하고; 그리고 “낮은 엄격도”는 Tm 아래의 약 20~25℃에서 발생한다. 대안적으로, 또는 추가적으로, 혼성화 조건은 혼성화의 염 또는 이온 세기 조건 및/또는 한 번 이상의 엄격한 세척(예: 6X SSC = 매우 낮은 엄격도; 3X SSC = 낮은 엄격도 내지 중간 엄격도; 1X SSC = 중간 엄격도; 및 0.5X SSC = 높은 엄격도)에 기초할 수 있다. 기능적으로, 최대 엄격도 조건은 혼성화 프로브와 엄격한 동일성 또는 거의 엄격한 동일성을 갖는 핵산 서열을 확인하는데 사용될 수 있다; 반면 고엄격도 조건은 프로브와 약 80% 이상의 서열 동일성을 갖는 핵산 서열을 확인하는데 사용된다. 높은 선택성을 필요로 하는 적용에 있어서, 하이브리드를 형성하기 위해 비교적 엄격한 조건을 사용하는 것이 전형적으로 바람직하다(예컨대, 비교적 낮은 염 및/또는 고온 조건이 사용됨).
본원에 사용된 용어 "혼성화"는 당 업계에 공지된 바와 같이 핵산 가닥이 염기쌍을 통해 상보적 가닥과 결합하는 과정을 지칭한다. 보다 구체적으로, "혼성화"는, 블롯 혼성화 기술 및 PCR 기술 동안 발생하는 것과 같이, 핵산의 한 가닥이 상보적 가닥과 듀플렉스, 즉 염기 쌍을 형성하는 과정을 지칭한다. 핵산 서열은 중등도 내지 고 엄격도 혼성화 및 세척 조건 하에 두 서열이 특이적으로 서로 혼성화되는 경우 기준 핵산 서열에 대해 "선택적으로 혼성화 가능"하다고 간주된다. 혼성화 조건은 핵산 결합 복합체 또는 프로브의 용융 온도(Tm)에 근거한다. 예를 들어, "최대 엄격도"는 일반적으로 약 Tm -5℃(프로브의 Tm 아래의 5℃)에서 발생한다. 높은 엄격도는 Tm 아래의 약 5~10℃에서 발생하고; “중간 엄격도”는 프로브의 Tm 아래의 약 10~20℃에서 발생하고; 그리고 “낮은 엄격도"는 Tm 아래의 약 20~25℃에서 발생한다. 기능적으로, 최대 엄격도 조건은 엄격한 동일성 또는 혼성화 프로브와 거의 엄격한 동일성을 갖는 서열을 확인하는데 사용될 수 있고; 반면에 중간 또는 낮은 엄격도 혼성화는 폴리뉴클레오티드 서열 동족체를 확인하거나 검출하는데 사용될 수 있다.
중간 및 높은 엄격도 혼성화 조건은 당업계에 잘 공지되어 있다. 예를 들어, 20% 포름아미드, 5 x SSC (150 mM NaCl, 15 mM 트리소디움 시트레이트), 50 mM 소듐 포스페이트(pH 7.6), 5 x 덴하르트(Denhardt) 용액, 10% 덱스트란 설페이트 및 20 mg/mL 변성 전단 연어 정자 DNA를 포함하는 용액에서 37℃에서 밤새 배양으로 중간 엄격도 혼성화를 수행한 후, 필터를 37~50℃의 1x SSC에서 세척할 수 있다. 고 엄격도 혼성화 조건은 65℃ 및 0.1x SSC(1x SSC=0.15 M NaCl, 0.015 M Na 시트레이트, pH 7.0)에서의 혼성화일 수 있다. 대안적으로, 약 42℃에서 50% 포름아미드, 5X SSC, 5X 덴하르트 용액, 0.5% SDS 및 100 μg/mL 변성 캐리어 DNA에서 고 엄격도 혼성화 조건을 수행한 후 상온에서 2X SSC 및 0.5% SDS에서 2회 세척 및 42℃에서 0.1X SSC와 0.5 % SDS에서 2회 추가로 세척하였다. 매우 높은 엄격도 혼성화 조건은 68℃ 및 0.1X SSC에서의 혼성화일 수 있다. 또한 당업자는 프로브 길이 등과 같은 인자를 수용하기 위해 필요에 따라 온도, 이온 세기 등을 조정하는 방법을 알고있다.
적어도 2 개의 핵산 또는 폴리펩티드와 관련하여 "실질적으로 유사한” 또는 "실질적으로 동일한"이라는 문구는 폴리뉴클레오티드 또는 폴리펩티드가 부모 또는 기준 서열과 적어도 90%, 적어도 91%, 적어도 92%, 적어도 93%, 적어도 94%, 적어도 95%, 적어도 96%, 적어도 97%, 적어도 98%, 또는 심지어 적어도 99% 동일하거나, 기능성을 추가하지 않고서 본 설명을 단지 우회하기 위해 이루어진 아미노산 치환, 삽입, 삭제 또는 변형을 포함하지 않는 것을 의미한다.
핵산 또는 폴리펩티드 서열의 문맥에서 "서열 동일성” 또는 "동일성"은, 특정 비교창에 대한 최대 일치를 위해 정렬될 때, 동일한 두 서열 내의 핵산 염기 또는 아미노산 잔기를 나타낸다.
용어 "서열 동일성 백분율"은 비교창에서 최적으로 정렬된 2개의 서열을 비교하여 결정된 값을 지칭하며, 이때, 비교창 내의 폴리뉴클레오티드 또는 폴리펩티드 서열의 부분은 2개의 서열의 최적 정렬을 위한 (삽입 또는 결실을 포함하지 않는) 기준 서열과 비교하여 삽입 또는 결실(즉, 갭)을 포함할 수 있다. 백분율은, 두 서열에서 동일한 핵산 염기 또는 아미노산 잔기가 나타나는 위치의 개수를 결정하여 일치하는 위치의 개수를 산출하고, 일치하는 위치의 개수를 비교창 내의 위치의 총 개수로 나누고, 그 결과에 100을 곱하여 서열 동일성의 백분율을 산출함으로써 계산한다. 백분율 서열 동일성의 유용한 예는 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% 또는 95% 내지 100%의 임의 정수 백분율을 포함하지만 이들에 한정되지 않는다. 이들 동일성은 본원에 기재된 프로그램중 임의의 것을 사용하여 판단될 수 있다.
서열 정렬 및 백분율 동일성 또는 유사성 계산은 LASERGENE 생물 정보학 컴퓨팅 스위트(DNASTAR Inc., Madison, WI)의 MegAlignTM 프로그램을 포함하지만 이에 한정되지 않는 상동성 서열을 검출하도록 설계된 다양한 비교 방법을 사용하여 결정될 수 있다. 본 출원의 문맥에서, 서열 분석 소프트웨어가 분석에 사용되는 경우, 달리 명시되지 않는 한, 분석 결과는 언급된 프로그램의 "디폴트 값"에 기초할 것이라는 것을 이해할 것이다. 본원에서 사용된 바와 같이, "디폴트 값"은 최초로 초기화 될 때, 원래 소프트웨어로 로딩되는 임의의 값 또는 파라미터 세트를 의미할 것이다.
"클러스탈 V 정렬 방법"은 클러스탈 V(Higgins and Sharp, (1989) CABIOS 5 : 151-153, Higgins 등, (1992) Comput Appl Biosci 8 : 189-191에 기재)로 명명되고 LASERGENE 생물 정보학 컴퓨팅 스위트의 MegAlignTM 프로그램 (DNASTAR Inc., Madison, WI)에서 발견되는 정렬 방법에 해당한다. 다중 정렬의 경우, 디폴트 값은 GAP PENALTY=10 및 GAP LENGTH PENALTY=10에 해당한다. 클러스탈 방법을 사용하는 단백질 서열의 백분율 동일성의 계산 및 쌍 정렬을 위한 디폴트 파라미터는 KTUPLE=1, GAP PENALTY=3, WINDOW=5 및 DIAGONALS SAVED=5이다. 핵산의 경우, 이들 파라미터는 KTUPLE=2, GAP PENALTY=5, WINDOW=4 및 DIAGONALS SAVED=4이다. 클러스탈 V 프로그램을 사용하여 서열을 정렬한 후에는, 동일한 프로그램에서 "서열 거리"표를 보고 "백분율 동일성"을 얻을 수 있다.
"클러스탈 W 정렬 방법"은 클러스탈 W(Higgins and Sharp, (1989) CABIOS 5 : 151-153, Higgins 등, (1992) Comput Appl Biosci 8 : 189-191에 기재)로 명명되고 LASERGENE 생물 정보학 컴퓨팅 스위트의 MegAlignTM v6.1 프로그램(DNASTAR Inc., Madison, WI)에서 발견되는 정렬 방법에 해당한다. 다중 정렬을 위한 디폴트 파라미터(GAP PENALTY=10, GAP LENGTH PENALTY=0.2, 지연 발산 서열(%) = 30, DNA 전이 중량 = 0.5, 단백질 무게 매트릭스 = Gonnet 시리즈, DNA 무게 매트릭스 = IUB). 클러스탈 W 프로그램을 사용하여 서열을 정렬한 후에는, 동일한 프로그램에서 "서열 거리"표를 보고 "백분율 동일성"을 얻을 수 있다.
달리 명시되지 않는 한, 본원에 제공된 서열 동일성/유사성 값은 하기 파라미터를 사용하여, GAP 버전 10(GCG, Accelrys, San Diego, CA)을 사용하여 얻은 값을 나타낸다: 갭 생성 패널티 중량 50 및 갭 길이 연장 페널티 중량 3을 사용하는 뉴클레오타이드 서열에 대한 % 동일성 및 % 유사성, 및 nwsgapdna.cmp 스코어링 매트릭스; GAP 생성 페널티 중량 8 및 갭 길이 연장 패널티 2를 사용하는 아미노산 서열에 대한 % 동일성 및 % 유사성, 및 BLOSUM62 스코어링 매트릭스(Henikoff 및 Henikoff, (1989) Proc. Natl. Acad. Sci.USA89: 10915). GAP는 Needleman 및 Wunsch (1970) J Mol Biol 48 : 443-53의 알고리즘을 사용하여 일치 횟수를 최대화하고 갭의 수를 최소화하는 두 개의 완전한 서열의 정렬을 찾는다. GAP는 가능한 모든 정렬 및 갭 위치를 고려하고, 일치하는 기준 단위로 갭 생성 페널티 및 갭 연장 페널티를 사용하여 가장 많은 수의 일치된 기준과 가장 적은 갭으로 정렬을 생성한다.
서열 동일성의 많은 레벨은 다른 종 또는 천연 변형되거나 합성된 종으로부터 폴리펩티드를 확인하는데 유용하고, 이러한 폴리펩티드는 동일 또는 유사한 기능 또는 활성을 갖는다는 것이 당업자에 의해 잘 이해된다. 백분율 동일성의 유용한 예는 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% 또는 95%, 또는 50% 내지 100%의 임의 정수 백분율을 포함하지만 이들에 한정되지 않는다. 실제로, 50% 내지 100%, 예컨대 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% 또는 99%의 임의 정수 아미노산 동일성은 본 개시를 설명하는데 유용할 수 있다.
"유전자"는, 이들에 한정되지는 않지만 예컨대 특정 폴리펩티드(예컨대, 효소) 또는 기능적 RNA 분자(예컨대, 가이드 RNA, 항-센스 RNA, 리보자임 등)와 같은 기능적 분자를 인코딩하고 발현할 수 있는 핵산 단편을 포함하고, 코딩 서열 앞(5 '비-코딩 서열) 및/또는 코딩 서열 뒤(3’ 비-코딩 서열)의 조절 서열을 포함한다. "천연 유전자"는 자신의 조절 서열과 함께 자연에서 발견되는 유전자를 지칭한다. 재조합 유전자는 상이한 유기체 또는 동일한 유기체로부터 유래될 수 있는 상이한 유전자의 조절 서열에 의해 조절되는 유전자를 지칭한다.
"돌연변이된 유전자"는 인간 개입을 통해 변경된 유전자이다. 이러한 "돌연변이된 유전자"는 적어도 하나의 뉴클레오티드 삽입, 결실, 또는 치환에 의해 상응하는 비 돌연변이된 유전자의 서열과 상이한 서열을 갖는다. 본 개시의 특정 구현예에서, 돌연변이된 유전자는 본원에 개시된 가이드 폴리뉴클레오티드/Cas 엔도뉴클레아제 시스템을 이용하여 만들어진 변경을 포함한다. 돌연변이된 진균 세포는 돌연변이된 유전자를 포함하는 진균 세포이다.
본원에서 사용된 "표적화된 돌연변이"는, 본원에 개시되거나 당업계에 공지된 바와 같이, 표적 서열의 DNA에서 이중 가닥 절단을 유도할 수 있는 이중 가닥 절단 유도제를 포함하는 방법을 사용하여 천연 유전자 내에서 표적 서열을 변경함으로써 만들어진 천연 유전자의 돌연변이이다.
용어 "공여체 DNA” 또는 "공여체 핵산 서열” 또는 "공여체 폴리뉴클레오티드"는 일반적으로 Cas/가이드 폴리뉴클레오티드 복합체(위에서 상세하게 기재된 바와 같이, 가이드 폴리뉴클레오티드는 표적 부위를 정의함)의 활성과 관련하여, 표적 부위에 또는 표적 부위 근처에 삽입되거나 표적 부위에 또는 표적 부위 근처의 영역을 대체할 관심 폴리뉴클레오티드 서열을 포함하는 폴리뉴클레오티드를 지칭한다. 이와 같이, 공여체 DNA에서 관심 폴리뉴클레오티드 서열은, 표적 부위에 또는 표적 부위 부근에서 치환되거나/편집될 뉴클레오타이드 서열과 비교할 때, 표적 부위에 또는 표적 부위 근처에 삽입되는 신규한 영역 및/또는 변형된 폴리뉴클레오티드 서열을 포함할 수 있다. 특정 구현예에서, 공여체 DNA 작제물은 관심 폴리뉴클레오티드 서열에 인접한 상동성의 제1 및 제2 영역을 더 포함한다. 공여체 DNA의 상동성의 제1 및 제2 영역은 각각 진균 세포 게놈의 표적 부위에 또는 그 측면에 존재하는 제1 및 제2 게놈 부위와 상동성을 공유한다. "상동성"이란 유사한 DNA 서열을 의미한다. 예를 들어, 공여체 DNA에서 발견되는 "게놈 영역에 대한 상동성 영역"은 진균 세포 게놈의 주어진 "게놈 영역"과 유사한 서열을 갖는 DNA 영역이다. 상동성의 영역은 절단된 표적 부위에서 상동종 재조합을 촉진시키기에 충분한 임의의 길이일 수 있다. 예를 들어, 상동성의 영역은 해당 게놈 영역과 상동성 재조합을 겪기에 충분한 상동성을 가지도록, 길이가 적어도 5~10, 5~15, 5~20, 5~25, 5~30, 5~35, 5~40, 5~45, 5~50, 5~55, 5~60, 5~65, 5~70, 5~75, 5~80, 5~85, 5~90, 5~95, 5~100, 5~200, 5~300, 5~400, 5~500, 5~600, 5~700, 5~800, 5~900, 5~1000, 5~1100, 5~1200, 5~1300, 5~1400, 5~1500, 5~1600, 5~1700, 5~1800, 5~1900, 5~2000, 5~2100, 5~2200, 5~2300, 5~2400, 5~2500, 5~2600, 5~2700, 5~2800, 5~2900, 5~3000, 5~3100 이상의 염기를 포함할 수 있다. "충분한 상동성"은 2개의 폴리뉴클레오티드 서열이 상동성 재조합 반응을 위한 기질로서 작용하기에 충분한 구조적 유사성을 갖는다는 것을 나타낸다. 구조적 유사성은 각 폴리뉴클레오티드 단편의 전체 길이뿐만 아니라 폴리뉴클레오티드의 서열 유사성을 포함한다. 서열 유사성은 서열의 전체 길이에 걸쳐 백분율 서열 동일성 및/또는 100% 서열 동일성을 갖는 인접 뉴클레오티드와 같은 국소화된 유사성 및 서열 길이의 일부분에 걸쳐 백분율 서열 동일성을 포함하는 보존된 영역에 의해 기술될 수 있다.
표적 및 공여체 폴리뉴클레오티드가 공유하는 상동성 또는 서열 동일성의 양은 변할 수 있으며, 약 1~20 bp, 20~50 bp, 50~100 bp, 75~150 bp, 100~250 bp, 150~300 bp, 200~400 bp, 250~500 bp, 300~600 bp, 350~750 bp, 400~800 bp, 450~900 bp, 500~1000 bp, 600~1250 bp, 700~1500 bp, 800~1750 bp, 900~2000 bp, 1~2.5 kb, 1.5~3 kb, 2~4 kb, 2.5~5 kb, 3~6 kb, 3.5~7 kb, 4~8 kb, 5~10 kb, 또는 표적 부위의 전체 길이까지를 포함하는 범위의 단위 적분 값을 갖는 총 길이 및/또는 영역을 포함한다. 이 범위에는 범위 내의 모든 정수가 포함되고, 예를 들어 1~20 bp 범위는 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 및 20 bp를 포함한다. 상동성의 양은 적어도 약 50%, 55%, 60%, 65%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%의 백분율 서열 동일성을 포함하는 2개의 폴리뉴클에오티드의 전체 정렬 길이에 걸쳐 백분율 서열 동일성에 의해 또한 기술될 수 있다. 충분한 상동성은 폴리뉴클레오티드 길이, 전체 백분율 서열 동일성, 및 임의로 보존된 인접 뉴클레오티드 또는 국소 백분율 서열 동일성의 임의의 조합을 포함하며, 예를 들어 충분한 상동성은 표적좌의 영역에 대해 적어도 80% 서열 동일성을 갖는 75 내지 150 bp의 영역으로 기술될 수 있다. 충분한 상동성은 또한 고 엄격도 조건 하에서 특이적으로 혼성화하는 2개의 폴리뉴클레오티드의 예측된 능력에 의해 기술될 수도 있다(예컨데, Sambrook 등, (1989) Molecular Cloning: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, NY); Current Protocols in Molecular Biology, Ausubel 등, Eds (1994) Current Protocols, (Greene Publishing Associates, Inc. and John Wiley & Sons, Inc); and, Tijssen (1993) Laboratory Techniques in Biochemistry and Molecular Biology--Hybridization with Nucleic Acid Probes, (Elsevier, New York) 참조).
본원에서 사용된 "게놈 영역"은 표적 부위의 어느 한면에 존재하거나 대안적으로 표적 부위의 일부도 포함하는 진균 세포의 게놈에서 염색체의 세그먼트이다. 게놈 영역은 해당 상동성 영역과 상동성 재조합을 겪기에 충분한 상동성을 가지도록, 적어도 5~10, 5~15, 5~20, 5~25, 5~30, 5~35, 5~40, 5~45, 5~50, 5~55, 5~60, 5~65, 5~70, 5~75, 5~80, 5~85, 5~90, 5~95, 5~100, 5~200, 5~300, 5~400, 5~500, 5~600, 5~700, 5~800, 5~900, 5~1000, 5~1100, 5~1200, 5~1300, 5~1400, 5~1500, 5~1600, 5~1700, 5~1800, 5~1900, 5~2000, 5~2100, 5~2200, 5~2300, 5~2400, 5~2500, 5~2600, 5~2700, 5~2800. 5~2900, 5~3000, 5~3100 이상의 염기를 포함할 수 있다.
본원에 사용된 “상동성 재조합”은 상동성 부위에서 2개의 DNA 분자들간 DNA 단편들의 교환을 포함하고, 당업계에 잘 기술되어 있다.
표현형 마커는 시각적 마커 및 그것이 양성의 선택 가능한 마커든 음성의 선택 가능한 마커든 선택 가능한 마커를 포함하는, 선별이 가능하거나 선택 가능한 마커이다. 임의의 표현형 마커가 이용될 수 있다. 구체적으로, 선택 가능하거나 선별 가능한 마커는 종종 특정한 조건 하에서, 그것을 함유하는 분자 또는 세포를 확인할 수 있게 하거나, 그것을 함유하는 분자 또는 세포를 선택 또는 선별할 수 있게 하는 DNA 세그먼트를 포함한다. 이들 마커는 활성, 예컨대, RNA, 펩티드, 또는 단백질의 생산(그러나 이에 한정되지 않음)을 인코딩할 수 있거나, RNA, 펩티드, 단백질, 무기 및 유기 화합물 또는 조성물 등을 위한 결합 자리를 제공할 수 있다.
선택 가능한 마커의 예로는 제한 효소 부위를 포함하는 DNA 세그먼트; 독성 화합물 및 항생제, 예컨대, 클로리무론 에틸, 베노밀, 바스타 및 히그로마이신 포스포트랜스퍼라제(HPT)에 대해 저항성을 제공하는 생성물을 인코딩하는 DNA 세그먼트; 수용 세포에 결여되어 있는 생성물을 인코딩하는 DNA 세그먼트(예컨대, tRNA 유전자, 영양요구성 마커, 우세 이종 마커-amdS); 용이하게 식별될 수 있는 생성물을 인코딩하는 DNA 세그먼트(예를 들어, 표현형 마커, 예컨대, β-갈락토시다제, GUS; 형광 단백질, 예컨대, 녹색 형광 단백질(GFP), 청록색 형광 단백질(CFP), 황색 형광 단백질(YFP), 적색 형광 단백질(RFP) 및 세포 표면 단백질); PCR을 위한 새로운 프라이머 부위의 생성(예컨대, 이전에는 나란히 놓여있지 않았던 두 DNA 서열의 병치), 제한 엔도뉴클레아제 또는 기타 DNA 변형 효소, 화학물질 등에 의해 영향 받지 않거나 영향 받는 DNA 서열의 포함; 및 식별을 가능하게 하는 특이적인 변형(예컨대, 메틸화)에 필요한 DNA 서열의 포함을 들 수 있으나, 이에 한정되지 않는다.
진균 세포 게놈을 변형하기 위한 방법
진균 세포, 예컨대 사상균 세포 내 게놈좌와 공여체 DNA의 상동성 재조합을 촉진하기 위한 가이드 RNA/Cas 엔도뉴클레아제 시스템을 채용한 방법이 제공된다.
본 개시의 양태는 Cas 엔도뉴클레아제/가이드 폴리뉴클레오티드 복합체를 게놈좌와 상동성을 갖는 도메인을 포함하는 공여체 DNA와 함께 세포 내로 일시적으로 도입함으로써 진균 세포의 게놈 내의 게놈좌와 DNA 서열의 상동성 재조합을 위한 방법을 포함한다. Cas 엔도뉴클레아제/가이드 폴리뉴클레오티드 복합체는 진균 세포의 게놈 내 원하는 표적 부위에서 작용할 수 있고, 여기서 “작용”이란 (위에서 정의된 것처럼) 가이드 폴리뉴클레오티드의 서열에 의해 안내된 Cas 엔도뉴클레아제가 표적 부위에서 DNA의 양 가닥 중 하나를 절단하는 것을 의미한다.
Cas 엔도뉴클레아제, 가이드 폴리뉴클레오티드, 및 공여체 DNA의 도입은 형질감염, 형질도입, 형질전환, 전기충격유전자전달법(electroporation), 입자 충격, 세포 융합 기술 등을 포함한 임의의 편리한 방식으로 수행될 수 있다. 이들 성분들 각각은 사용자가 원할 시 동시에 또는 순차적으로 도입될 수 있다. 예를 들어, 진균 세포는 처음에 Cas 발현 DNA 작제물로 안정적으로 형질감염될 수 있고, 이어서 가이드 폴리클레오티드를 (직접 또는 가이드 폴리뉴클레오티드 발현 DNA 작제물을 사용하여) 안정적인 형질전환체에 도입할 수 있다. 이 설정은 사용자가 상이한 가이드 폴리뉴클레오티드가 독립적으로 도입될 수 있는 안정한 Cas 형질전환체 진균 세포군을 생성할 수 있기 때문에 유리할 수 있다(일부 경우에는 필요하다면 하나 초과의 가이드 폴리뉴클레오티드가 동일한 세포 내로 도입 될 수 있다). 일부 구현예에서, Cas 발현 진균 세포는 사용자에 의해 수득되고, 따라서 사용자는 세포 내로 Cas 엔도뉴클레아제를 발현할 수 있는 재조합 DNA 작제물을 도입할 필요가 없으며, 오히려 Cas 발현 세포 내로 가이드 폴리뉴클레오티드를 도입할 필요만 있다.
특정 구현예에서, 가이드 폴리뉴클레오티드는 가이드 폴리뉴클레오티드를 인코딩하는 발현 카세트(또는 유전자)를 포함하는 재조합 DNA 작제물을 진균 세포로 도입함으로써 도입된다. 일부 구현예에서, 발현 카세트는 진핵생물의 RNA 폴 III 프로모터에 작동 가능하게 연결된다. 이들 프로모터는 RNA 폴 III에 의한 전사가 RNA 폴 II 의존 프로모터로부터의 RNA 중합효소 II에 의한 전사시에 발생하는 5 '캡 구조 또는 폴리아데닐화의 첨가로 이어지지 않기 때문에 특히 흥미롭다. 특정 구현예에서, RNA 폴 III 프로모터는 사상균 세포 U6 중합효소 III 프로모터이다(예컨대, 서열 번호 40 및 이의 기능적 변이체, 예컨대 서열 번호 41; 아래에서 더 상세히 기재됨).
이중 가닥 절단이 숙주 세포의 게놈 DNA에서 유도 될 때(예컨대, 표적 부위에서 Cas 엔도뉴클레아제/가이드 RNA 복합체의 활성, 이중 가닥 엔도뉴클레아제 활성을 갖는 복합체), 세포의 DNA 복구 메카니즘이 활성화되어, 오류가 발생하기 쉬운 성질로 인하여 이중 가닥 절단 부위에서 돌연변이를 일으킬 수 있는 절단을 복구한다. 절단된 말단을 하나로 합치는 가장 일반적인 치료 메카니즘은 비상동성 말단 결합(NHEJ) 경로이다(Bleuyard 등, (2006) DNA Repair 5:1-12). 염색체의 구조적 완전성은 전형적으로 치료에 의해 보존되지만, 결실, 삽입 또는 다른 재배열이 가능하다(Siebert and Puchta, (2002) Plant Cell 14 : 1121-31, Pacher 등, (2007) Genetics 175 : 21 -9).
놀랍게도, 사상 곰팡이의 경우 이중 가닥 절단에서 형질전환 된 DNA의 비상동성 삽입이 이중 가닥 절단에서 염색체 DNA의 두 말단 사이의 단순한 말단 결합보다 매우 선호된다는 것을 발견했다. 따라서, Cas 엔도뉴클레아제 또는 가이드 RNA가 DNA 작제물 또는 작제물들을 함유하는 발현 카세트로 형질전환함으로써 제공되는 경우, 이들 DNA 작제물 또는 이의 단편은 높은 빈도로 이중 가닥 절단부에 삽입된다. 이 삽입은 Cas 엔도뉴클레아제의 DNA 서열 또는 가이드 RNA 발현 작제물 및 이중 가닥 절단 주위의 서열 사이에 상동성이 없는 경우에 발생한다. 이 과정은 또한 상동성 재조합보다 전체 공여체 DNA의 삽입이 선호되므로 공여체 DNA와 게놈좌 사이의 상동성 재조합이 바람직할 때 문제가 된다. 진균 세포에서 자율적으로 유지될 것으로 예상되는 텔로미어 서열을 포함하는 벡터의 형태일지라도, 형질전환 된 DNA의 바람직하지 않은 삽입이 일어난다는 것을 발견했다.
형질전환에 의해 흡수된 DNA는 게놈 내에서 안정한 형태로 통합되거나 일시적으로 유지될 수 있다. 일시적인 유지관리는 불안정한 표현형에 의해 인식될 수 있다. 예를 들어, DNA 흡수는 형질전환 DNA 상에 존재하는 마커 유전자에 대한 선택에 의해 인식될 수 있다. 형질전환 및 삽입 후, 형질전환체는 선택적 조건으로 다시 전이되기 전에 여러 세대 동안 비선택적 조건 하에서 성장될 수있다. 안정한 형질전환체는 선택적 조건으로 전이한 후에 성장할 수 있지만, 불안정한 형질전환체는 형질전환된 DNA의 손실로 인해 선택적 조건으로 다시 전이한 후에는 성장할 수 없을 것이다. 아래의 실시예 부분에 나타낸 바와 같이, 진균 세포/불안정한 형질전환체에서 일시적으로 Cas 엔도뉴클레아제 및/또는 가이드 RNA를 발현하는 것이 가능하다는 것을 나타냈다.
불안정한 형질전환체가 요구되는 구현예에서, 자율 복제를 장려하는 텔로미어 서열을 갖는 플라스미드가 사용될 수 있다. 자율 복제 서열, 동위원소 서열 또는 기타 서열을 갖는 것과 같은 자율 복제를 위해 설계된 다른 유형의 플라스미드가 또한 사용될 수 있다. 놀랍게도, 트리코데르마 레에세이에서 복제, 자율 복제 서열, 동위원소 또는 텔로미어 서열의 기원이 알려지지 않은 플라스미드를 사용할 수 있음을 발견했다. 선택 가능한 마커에 대하여 불안정한 표현형을 나타내는 형질전환체를 선별함으로써, 벡터 DNA 삽입없이 효과적인 표적 부위 유전자 변형이 수득된다(예컨대, 공여체 DNA에서 상동성 영역과의 상동성 재조합).
본 개시의 특정 구현예는 반복부에 의해 선택적으로 플랭킹된 Cas 엔도뉴클레아제 발현 카세트 및 제1 선택 가능한 마커를 곰팡이의 게놈에 통합하여 발현 카세트 및 제1 선택 가능한 마커의 후속 제거(루프-아웃)를 가능하게 하여 Cas 엔도뉴클레아제를 발현하는 숙주 세포를 생산한다. 이들 세포는 공여체 DNA와의 상동성 재조합을 포함하여 관심 유전적 변형을 얻기 위해 다양한 방법으로 사용될 수 있다.
예를 들어, Cas 엔도뉴클레아제를 발현하는 숙주 세포는 제2 선택 가능 마커 (및 임의로 별도의 공여체 DNA)를 함유하는 가이드 RNA 발현 카세트를 포함하는 DNA 작제물로 형질전환 될 수 있다. 제2 선택 가능 마커를 사용하기 위해 선택된 숙주 세포는 Cas 엔도뉴클레아제 활성을 가능하게 하고 게놈에서 관심 대상이 되는 정의된 목표 부위에 대한 표적화를 가능하게 하는 이 DNA 작제물로부터 가이드 RNA를 발현한다. 제2 선택 가능 마커와 관련하여 불안정한 표현형을 나타내는 형질전환체에 대한 이들 숙주 세포를 선별하면 DNA 작제물 삽입없이 변형된 관심 부위(예컨대, 공여체 DNA와의 상동성 재조합)를 갖는 숙주 세포를 수득할 수 있다.
다른 예로서, Cas 엔도뉴클레아제를 발현하는 숙주 세포를 유도하여 시험관내 합성된 가이드 RNA를 흡수하여 Cas 엔도뉴클레아제 활성을 가능하게 하고 게놈 내 정의된 부위를 표적화할 수 있다. 일부 경우에는, 가이드 RNA 및 선택 가능 마커 유전자를 지닌 분리된 DNA 작제물 모두의 흡수를 유도하여 DNA를 취했고 고주파수에서 가이드 RNA를 동시에 흡수할 것으로 예상되는 세포를 선택하게 하는 것이 바람직할 것이다. 상기와 같이, 벡터 DNA 삽입없이 관심 유전자 변형(예컨대, 공여체 DNA와의 상동성 재조합)에 대한 선택 가능 마커에 대하여 불안정한 표현형을 나타내는 형질전환체를 선별한다.
또 다른 예로서, Cas 엔도뉴클레아제를 발현하는 숙주 세포는 번역시 Cas 엔도뉴클레아제를 “표적 균주”에게 제공할 수 있는 “헬퍼 균주(helper strain)”를 생성하기 위해 사용될 수 있다. 간단히 말해서, 헤테로카리온은 헬퍼 균주 및 표적 균주 사이에서, 예를 들어 각 균주로부터의 원형질체의 융합 또는 사상균의 종에 따른 균사의 문합에 의해 생성될 수 있다. 헤테로카리온의 유지는, 부모 균주가 성장할 수 없지만 헤테로카리온이 보상으로 인해 성장할 수 있도록 각각의 부모 균주에서 적절한 영양의 또는 다른 마커 유전자 또는 돌연변이 및 적절한 선택 배지에서의 성장에 의존할 것이다. 헤테로카리온 형성시 또는 이후에, 가이드 RNA가 형질감염 (및 선택적으로 공여체 DNA)에 의해 도입된다. 가이드 RNA는 직접 도입되거나 Cas 엔도뉴클레아제 발현 카세트 및 선택 가능 마커 유전자를 갖는 DNA 작제물을 통해 도입될 수 있다. Cas 엔도뉴클레아제는 헬퍼 균주 핵 내의 유전자로부터 발현되고 헤테로카리온의 세포질에 존재한다. Cas 엔도뉴클레아제는 게놈에서 원하는 표적 부위(들)를 표적으로 하는 활성 복합체를 생성하기 위해 가이드 RNA와 결합한다. 이어서, 포자는 헤테로카리온으로부터 회수되고, 표적 부위에서 또는 표적 부위 근처에서 변형(예컨대, 게놈좌에서 공여체 DNA와의 상동성 재조합)으로 표적 균주를 회수하기 위해 선택 또는 선별된다. 발현 카세트가 가이드 RNA를 도입하는데 사용되는 경우, 가이드 RNA 발현 작제물이 안정적으로 유지되지 않는 헤테로카리온이 선택된다.
전술한 바와 같이, 본 개시의 방법은 Cas/가이드 RNA 복합체의 표적 부위의 각 측상의 염색체 DNA의 영역과 DNA 서열 상동성을 갖는 DNA 작제물을 세포(또는 공여체 DNA)에 도입하는 단계를 포함한다. 의도는 DNA 단편이 상동성 통합/재조합에 의해 통합되어, 표적 부위에서 DNA의 절단을 복구하고, 대부분의 경우 원하는 위치에서(예컨대, Cas/가이드 RNA 복합체의 표적 부위에서 또는 표적 부위 근처에서) 게놈에 변화를 도입하려는 것이다. 많은 생물체에서, 염색체 DNA의 이중 가닥 절단은 그 부위에서 선형 DNA 단편의 상동성 통합을 자극한다. 놀랍게도, 작용하는 NHEJ 경로를 갖는 사상 균류에서. 공여체 단편이 도입될 때조차, 이중 가닥 절단에서의 비상동성 삽입에 의한 DNA의 삽입이 선형 DNA 단편의 상동종 재조합보다 매우 선호된다는 것을 발견했다.
진균 세포에서의 DNA 복구에 대해서, 기능적 NHEJ 경로의 존재시, 오류가 발생하기 쉬운 복구가 이중 가닥 절단 부위에서 상동종 재조합보다 매우 선호된다는 것을 발견했다. 다시 말해서, 사상균 세포에서 이중 가닥 절단의 DNA 복구에 관해서, 기능적 NHEJ 경로의 존재 하에서, 절단시 DNA의 비상동성 삽입이 (1) DNA 삽입없이 비상동성 말단 결합 및 (2) 공여체 DNA로 이중 가닥 절단 부위에서 상동성 재조합보다 매우 선호된다는 것을 발견했다. 따라서, 본 발명의 특정 양태에서, 집단 내 진균 세포의 표적 부위에서 비상동성 말단 결합(NHEJ) 경로의 기능화는 억제되거나, 활성화되지 않거나, 비기능적이거나, 또는 감소된다. 이는 임의의 편리한 방식으로 달성될 수 있으며, 그 중 일부는 이하에 설명된다.
일부 구현예에서, 진균 세포의 표적 부위에서 비상동성 말단 결합(NHEJ) 경로의 기능화는, NHEJ 경로의 하나 이상의 성분이 비기능적이거나, 감소된 활성을 갖도록(예를 들어, 게놈으로부터 결실되거나 돌연변이되어 비기능적이거나 덜 활성인) 진균 숙주 세포를 변화시킴으로써 억제되거나, 활성화되지 않거나, 비기능적이거나, 또는 감소된다. 진균 세포의 이러한 변경은 유전자 결실, 유전자 돌연변이, 우성 간섭 재조합 단백질의 발현, 유전자 대체, 예컨대 안티센스 RNA/RNAi 방법론 등을 사용하는 유전자 발현 억제 등의 임의의 편리한 수단에 의해 달성될 수 있다. 특정 양태에서, NHEJ 경로의 하나 이상의 성분은 ku80, ku70, rad50, mre11, xrs2, lig4 및 xrs로 이루어진 군으로부터 선택된다. 하나의 예로서, 본 발명의 양태에서 사용되는 진균 세포는 ku80의 발현 및/또는 활성을 저해하는 유전자 변형을 포함한다.
추가의 구현예에서, 진균 세포의 표적 부위에서 비상동성 말단 결합(NHEJ) 경로의 기능화는 DNA 닉킹 활성을 갖는 Cas 엔도뉴클레아제를 사용함으로써 억제되거나, 활성화되지 않고, 비기능적이거나, 감소된다. 즉, 기능화는 표적 부위(Cas 닉카아제라고도 부름)에서 DNA의 한 가닥만을 절단한다. DNA의 이중 가닥 절단과는 달리, 닉은 NHEJ 경로를 활성화시키지는 않지만, 하나 이상의 상동성 영역을 갖는 공여체 DNA로 절단 부위에서 또는 절단 부위 근처에서 상동성 재조합을 촉진시키기에 충분하다. 야생형 Cas 엔도뉴클레아제의 재조합 변이체인 다수의 Cas 닉카아제가 당 기술에 기재되어 있으며(예를 들어 상기 정의 참조), 개시된 방법에서 사용될 수 있다.
일부 예에서, 공여체 DNA는 진균 세포의 게놈에서 상응하는 제1 및 제2 영역에 상동인 제1 영역 및 제2 영역을 포함하며, 상동성 영역은 일반적으로 게놈 DNA가 Cas 엔도뉴클레아제에 의해 절단되는 표적 부위를 포함하거나 둘러싼다. 상동성의 이들 영역은 상동성 재조합을 촉진하여 상동성의 상응하는 게놈 영역은 공여체 DNA와 게놈 간의 DNA 교환을 초래한다. 이와 같이, 제공된 방법은 진균 세포 게놈의 표적 부위의 절단 부위에서 또는 절단 부위 근처에서 공여체 DNA의 관심 폴리뉴클레오타이드의 통합을 초래하여, 원래의 표적 부위를 변경시킴으로써 변형된 게놈 표적 부위를 생성한다.
주어진 게놈 영역과 공여체 DNA에서 발견되는 해당 상동성 영역 사이의 구조적 유사성은 상동성 재조합이 일어날 수 있게 하는 임의의 서열 동일성의 정도가 될 수 있다. 예를 들어, 공여체 DNA의 "상동성 영역"과 진균 세포 게놈의 "게놈 영역"이 공유하는 상동성 또는 서열 동일성의 양은 적어도 50%, 55%, 60%, 65%, 70%, 75%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93% 95%, 96%, 97%, 98%, 99% 또는 심지어 100%의 서열 동일성을 가져서, 서열이 상동성 재조합을 겪는다.
공여체 DNA상의 상동성 영역은 표적 부위에 인접한 임의 서열과 상동성을 가질 수 있다. 일부 구현예에서 상동성의 영역들은 표적 부위의 바로 옆에 있는 게놈 서열에 대한 중요한 서열 상동성을 공유하지만, 상동성의 영역이 표적 부위에게 5’ 또는 3’ 더 길 수 있는 영역에 충분한 상동성을 갖도록 설계될 수 있다는 것이 인식된다. 또 다른 구현예에서, 상동성 영역은 하류 게놈 영역과 함께 표적 부위의 단편과 상동성을 가질 수 있다. 하나의 구현예에서, 상동성의 제1 영역은 표적 부위의 제1 단편을 추가로 포함하고, 상동성의 제2 영역은 표적 부위의 제2 단편을 포함하며, 제1 단편 및 제 2 단편은 상이하다.
Cas 엔도뉴클레아제 및 가이드 폴리뉴클레오티드 발현 작제물과 마찬가지로, 공여체 DNA는 임의의 편리한 수단(본원의 다른 곳에서 논의됨)에 의해 도입될 수 있다.
특정 구현예에서, Cas 엔도뉴클레아제는 Cas9 엔도뉴클레아제이다(예컨대, "RNA-directed DNA Cleavage by the Cas9-crRNA Complex"라는 제목의 WO 2013141680 참조). Cas9 엔도뉴클레아제의 예로는 스트렙토코쿠스 종(예컨대, S. 파이오제네스, S. 뮤탄스 및 S. 더모필루스), 캄피로박테르 종(Campylobacter sp.)(예컨대, C. 제주니), 나이세리아 종(Neisseria sp.)(예컨대, N. 메닝기티데스), 프란시셀라 종(Francisella sp.)(예컨대, F. 노비시다) 및 파스튜렐라 종(Pasteurella sp.)(예컨대, P. 물토시다)가 있다(본원에 참고로 통합된 Fonfara 등, Nucleic Acids Res., 2013, pages 1-14에 기재된 Cas9 엔도뉴클레아제를 참조). 일부 구현예에서, Cas 엔도뉴클레아제는 최적화된, 예컨대 진균 세포에서의 발현을 위해 최적화된 Cas9 엔도뉴클레아제 유전자(예를 들어, 하기 기재된 바와 같이 서열 번호 44, 예를 들어, 서열 번호 7을 포함하는 Cas9 인코딩 유전자)에 의해 인코딩된다.
특정 예에서, Cas 엔도뉴클레아제 유전자는 세포에서 발현되는 Cas 엔도뉴클레아제/가이드 폴리뉴클레오티드 복합체가 핵으로 효율적으로 운반되도록 핵 위치 신호를 인코딩하는 하나 이상의 폴리뉴클레오티드에 작동 가능하게 연결된다. 임의의 편리한 핵 위치 신호, 예를 들어, Cas 코돈 영역의 상류이자 Cas 코돈 영역과 동일 프레임에 존재하는 SV40 핵 위치 신호를 인코딩하는 폴리뉴클레오티드 및 Cas 코돈 영역의 하류이자 Cas 코돈 영역과 동일 프레임에 존재하는 T. 레에세이 blr2(청색광 조절인자 2)로부터 유도된 핵 위치 신호를 인코딩하는 폴리뉴클레오타이드가 사용될 수 있다. 다른 핵 위치 신호가 사용될 수 있다.
본 발명의 특정 구현예에서, 가이드 폴리뉴클레오티드는 유형 II Cas 엔도뉴클레아제와 복합체를 형성할 수 있는 유형 II CRISPR/Cas 시스템의 crRNA 영역(또는 crRNA 단편) 및 tracrRNA 영역(또는 tracrRNA 단편)을 포함하는 가이드 RNA이다. 상기한 바와 같이, 가이드 RNA/Cas 엔도뉴클레아제 복합체는 Cas 엔도뉴클레아제를 진균 세포 게놈 표적 부위로 향하게 할 수 있어, Cas 엔도뉴클레아제가 게놈 타겟 부위에 이중 가닥 절단을 도입할 수 있게 한다. 일부 경우에서, RNA/Cas9 엔도뉴클레아제 복합체를 안내하는 RNA는 crRNA와 별도의 tracrRNA를 포함하는 듀플렉스이다. 다른 예에서, 가이드 RNA는 (때때로 본원에서 융합된 가이드 RNA로 지칭되는) crRNA 영역 및 tracrRNA 영역 모두를 포함하는 단일 RNA 분자이다. 융합된 가이드 RNA 대 듀플렉스화된 crRNA-tracrRNA를 사용하는 한 가지 장점은 융합된 가이드 RNA를 발현하기 위해 하나의 발현 카세트만을 만들 필요가 있다는 것이다.
본원에 개시된 방법에 사용되는 숙주 세포는, 필라 아스코마이코타(phyla Ascomycota), 바시디오마이코타(Basidiomycota), 키트리디오마이코타Chytridiomycota), 및 자이고마이코타(Zygomycota)(Hawksworth 등, In, Ainsworth and Bisby 's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK에서 정의됨)뿐만 아니라 오오마이코타(Oomycota)(Hawksworth 등, supra에서 인용) 및 모든 불완전 균류(mitosporic fungi)(Hawksworth 등, supra)에서 유래된 임의의 진균 숙주 세포일 수 있다. 특정 구현예에서, 진균 숙주 세포는 효모 세포, 예컨대, 칸디다, 한세누라, 클루베로마이세스, 피키아, 사카로마이세스, 스키오사카로마이세스 또는 야로위아 세포이다. 효모 종은 다음을 포함하지만 이에 한정되지 않는다: 사카로마이세스 칼스베르겐시스(Saccharomyces carlsbergensis), 사카로마이세스 세레비시애(Saccharomyces cerevisiae), 사카로마이세스 디아스타티쿠스(Saccharomyces diastaticus), 사카로마이세스 도우글라시(Saccharomyces douglasii), 사카로마이세스 클루베리(Saccharomyces kluyveri), 사카로마이세스 노르벤시스(Saccharomyces norbensis), 사카로마이세스 오비포르미스(Saccharomyces oviformis), 클루베로마이세스 락티스(Kluyveromyces lactis), 및 야로위아 리폴리티카(Yarrowia lipolytica) 세포. 추가의 구현예에서, 진균 세포는 트리코데르마, 페니실리움, 아스퍼질러스, 휴미콜라, 크리소스포리움, 푸사리움, 뉴로스포라, 마이셀리오프토라, 하이포크레아 및 에메리셀라의 종을 포함하지만 이에 한정되지 않는 사상균 세포이다. 예를 들어, 사상 곰팡이 T. 레에세이 및 A. 나이거는 개시된 방법의 양태에서 사용된다.
표적 부위가 필요로 하는 프로토스페이서 인접 모티프(protospacer adjacent motiff(PAM))를 포함하는 한, 사실상 진균 세포 게놈의 임의의 부위는 개시된 방법을 사용하여 표적화될 수 있다. S. 파이로제네스 Cas9의 경우, PAM은 서열 NGG (5’에서 3'; N은 A, G, C 또는 T)를 가지며, 따라서 게놈의 표적 부위의 선택에 유의한 제한을 부과하지 않는다. 다른 공지된 Cas9 엔도뉴클레아제는 상이한 PAM 부위를 갖는다(예컨대, Fonfara 등, Nucleic Acids Res., 2013, pages 1-14에 기재된 Cas9 엔도뉴클레아제 PAM 부위 참조: 본원에 참고로 통합됨).
표적 부위의 길이는 변할 수 있으며, 예를 들어 적어도 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30 또는 그 이상의 뉴클레오티드 길이를 갖는 표적 부위를 포함한다. 표적 부위는 팔린드로믹일 수도 있다. 즉, 한 가닥 상에 있는 서열은 상보적 가닥에서 반대 방향으로 동일하게 판독된다. 절단 부위는 표적 서열 내에 존재할 수 있거나 절단 부위는 표적 서열 외부에 존재할 수 있다. 또 다른 변형에서, 절단은 블런트 엔드 컷을 생성하기 위해 서로 바로 마주 보는 뉴클레오티드 위치에서 발생할 수 있거나, 또는 다른 경우에는 "끈끈한 말단(sticky ends)"이라고도 부르는, 5’ 오버행 또는 3'오버행일 수 있는 단일 가닥 오버행을 생성하도록 절개가 틀어질 수 있다.
일부 경우에, 표적 부위가 (가이드 폴리뉴클레오티드의 crRNA 서열 내에서) 가이드 폴리뉴클레오티드의 관련 서열과 100% 동일하지 않다는 것을 의미하는, 진균 세포의 게놈에서의 활성 변이체 표적 서열도 사용될 수 있다. 이러한 활성 변이체는 주어진 표적 부위와 적어도 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 이상의 서열 동일성을 포함할 수 있고, 활성 변이체 표적 서열은 생물학적 활성을 보유하고, 따라서 Cas 엔도뉴클레아제에 의해 인식되고 절단 될 수 있다. 엔도뉴클레아제에 의한 표적 부위의 이중 가닥 절단을 측정하기 위한 분석은 당업계에 공지되어 있으며, 일반적으로 인식 부위를 포함하는 DNA 기질상의 작용제의 전체 활성 및 특이성을 측정한다.
관심 표적 부위는 관심 유전자의 영역 내에 위치하는 표적을 포함한다. 관심 유전자 내의 영역들의 비제한적 예로는 오픈 리딩 프레임, 프로모터, 전사 조절 요소, 번역 조절 요소, 전사 종결인자 서열, mRNA 스플라이스 부위, 단백질 코딩 서열, 인트론 부위, 및 인트론 증진 모티프가 있다.
특정 구현예에서, 진균 세포의 게놈의 변형은 검출될 수 있는 표현형 효과를 초래하고, 많은 예에서 사용자의 바람직한 결과이다. 비 제한적인 예로는 선택 가능한 세포 성장 표현형(예컨대, 항생제에 대한 저항성 또는 민감성, 영양 요구성 특성의 증가 또는 감소, 증가 또는 감소된 성장률 등), 검출 가능한 마커(예컨대, 형광 마커, 세포 표면 분자, 발색 효소 등), 그리고 배양 상등액에서 그 활성이 검출될 수 있는 효소의 분비가 있다.
진균 세포 게놈의 변형이 표현형 효과를 가져올 때, 표현형 마커인 관심 폴리뉴클레오티드를 포함하는 공여체 DNA가 종종 사용된다. 흔히 특정 배양 조건 하에서, 그것을 포함하는 진균 세포를 확인하거나 이를 혹은 이에 반하여 선택할 수 있게 하는 선택 가능하거나 선별 가능한 임의 마커를 포함하여 임의의 편리한 표현형 마커가 사용될 수 있다. 따라서, 본 발명의 일부 양태에서, 원하는 게놈 변형을 갖는 진균 세포의 확인은 표적 부위에서 변형을 갖는 세포를 선택하거나 선별하는 조건 하에서 Cas 엔도뉴클레아제 및 가이드 폴리뉴클레오티드(그리고 선택적으로 공여체 DNA)를 받은 진균 세포군을 배양하는 것을 포함한다. 진균 세포(선택 가능 마커라고도 지칭됨)에서의 효소 활성의 증가 또는 감소, 예를 들어 항생제 내성의 획득 또는 영양 요구성 마커의 증가/감소를 평가하는 것을 포함하여 임의 유형의 선택 시스템이 사용될 수 있다.
일부 예에서, 진균 세포에서의 게놈 변형은 시퀀싱, PCR, 서던 블롯, 제한 효소 분석 등을 포함하고, 이러한 방법들의 조합을 포함하는 임의의 편리한 방법을 사용하여 직접 검출된다.
일부 구현예에서, 효소, 예를 들어 아세틸 에스테라아제, 아미노펩티다아제, 아밀라아제, 아라비나아제, 아라비노푸라노시다아제, 카복시펩티다아제, 카탈라아제, 셀룰라아제, 키티나아제, 큐티나아제, 데옥시리보뉴클레아제, 에피메라아제, 에스테라아제, α- 갈락토시다아제, β- 갈락토시다아제, α- 글루카나아제, 글루카닐라아제, 엔도-β-글루카나아제, 글루코아밀라아제, 글루코스 옥시다아제, α-글루코시다아제, β-글루코시다아제, 글루쿠로니다아제, 헤미셀룰라아제, 헥소오스 옥시다아제, 하이드로라아제, 인베르타제, 이소메라제, 락카아제, 리파아제, 리아제, 만노시다제, 옥시다아제, 옥시도레덕타아제, 펙테이트 리아제, 펙틴 아세틸 에스테라아제, 펙틴 디폴리머라아제, 펙틴 메틸 에스테라아제, 펙틴 분해 효소, 페록시다아제, 페놀옥시다아제, 피타아제, 폴리갈락투로나아제, 프로테아제, 람노 - 갈락투로나아제, 리보뉴클레아제, 트랜스퍼라아제, 수송 단백질, 트랜스글루타미나아제, 크실라나아제, 헥소오스 옥시다아제 및 이들의 조합을 인코딩하는 유전자를 포함하여, 특정 유전자들이 개시된 방법을 사용하여 변형을 위해 표적화된다.
본원에 기재된 방법을 구현하기 위한 다양한 변형이 있다. 예를 들어, 진균 숙주 세포에 외인성 서열로서 존재하는 Cas 발현 카세트를 갖는 대신에, 이 카세트는 진균 숙주 세포의 게놈에 통합될 수 있다. 이러한 부모 세포주를 생성함으로써, 사용자는 본원의 다른 곳에서 상세히 설명된 바와 같이 관심 게놈 부위를 표적으로 하는 원하는 가이드 RNA(예컨대, 가이드 RNA 발현 벡터로서)를 간단히 도입 할 수있게 된다. 이들 구현예들의 일부에서, 통합된 Cas 유전자는 필요한 경우 게놈으로부터의 후속 루프-아웃/제거를 위해 거기에 인접한 폴리뉴클레오티드 리피트를 포함하도록 설계될 수 있다.
진균 세포의 조성물
본 발명의 양태는 변형된 게놈을 갖는 수득된 진균 세포뿐만 아니라 상기 기술된 방법을 수행하는데 사용되는 형질전환 진균 세포를 포함한다. 따라서, 본 발명의 구현예는 본원에 기재된 방법의 임의의 양태에 의해 생산된 재조합 진균 세포뿐만 아니라 이들을 생산하는데 사용되는 임의의 부모 진균 세포를 포함한다.
본 발명의 특정 구현예는 Cas 엔도뉴클레아제를 포함하는 재조합 진균 세포에 관한 것으로, 여기서 Cas 엔도뉴클레아제는 세포에서 재조합 DNA 작제물(제1 재조합 DNA 작제물)로부터 발현된다. 일부 구현예에서, 재조합 진균 세포는 비기능성 또는 활성이 감소된 NHEJ 경로를 갖는다. Cas 엔도뉴클레아제 및 위에서 상세히 기술된 Cas 엔도뉴클레아제를 인코딩하는 폴리뉴클레오티드의 구현예는 이 부분의 진균 세포 조성물에서 사용된다(그 중 일부는 하기에 기재됨). 이 진균 세포는 관심 대상인 원하는 게놈 변형을 갖는 진균 세포를 생성하기 위한 부모로서 사용되며, 게놈 변형을 생성하는 것은 세포 내로 가이드 폴리뉴클레오티드(예를 들어, 발현 카세트를 통해)를 도입하여 유전자 변형을 견인하는 Cas/가이드 폴리뉴클레오티드 복합체를 형성할 수 있다(위에서 기술 한 바와 같이). 특정 양태에서, NHEJ 경로의 하나 이상의 성분은 재조합 진균 세포에서 비기능적이거나 감소된 활성, 예컨대, ku80, ku70, rad50, mre11, xrs2, lig4, xrs 및 이들의 조합을 갖는다. 하나의 특별한 구현예에서, 재조합 진균 세포는 ku80의 발현 및/또는 활성을 저해하는 유전자 변형을 갖는다. 유전자 결실, 유전자 돌연변이, 우성 간섭 재조합 단백질의 발현, 유전자 대체, 예컨대 안티센스 RNA/RNAi 방법론 등을 사용하는 유전자 발현 억제를 포함하지만, 이들에 한정되지 않는 NHEJ 경로 성분(들)의 파괴를 이루기 위한 임의의 편리한 유전자 변형이 채용될 수 있다.
특정 양태에서, 재조합 진균 세포는 가이드 RNA를 발현할 수 있는 제2 재조합 DNA 작제물을 추가로 포함하며, 여기서 가이드 RNA 및 Cas 엔도뉴클레아제는 Cas 엔도뉴클레아제가 재조합 진균 세포의 게놈 내 표적 부위에서 이중 가닥 절단을 유도할 수 있게 하는 복합체를 형성할 수 있다. 가이드 폴리뉴클레오티드 및 위에서 상세히 기술된 가이드 폴리뉴클레오티드를 인코딩하는 폴리뉴클레오티드의 구현예는 이 부분의 진균 세포 조성물에서 사용된다(그 중 일부는 하기에 기재됨). 가이드 RNA의 발현은 진핵 생물 RNA 폴 III 프로모터에 의해 유도될 수 있으며, 특정 구현예에서 RNA 폴 III 프로모터는 사상균 세포 U6 유전자 프로모터(예컨대, 하기에서 더 상세히 기술된 바와 같은 서열 번호 40 또는 서열 번호 41) 및 이의 기능적 변이체이다. 이 복합체의 작용은 진균 세포의 표적 부위에서 게놈 DNA 서열을 변형시켜(위에서 기술된 바와 같이), 표적 부위에 (또는 근처에) 변형된 진균 세포를 생성할 수 있다. 변형은 하나 이상의 뉴클레오타이드의 결실, 하나 이상의 뉴클레오타이드의 삽입, 하나 이상의 뉴클레오타이드의 치환, 또는 이들의 임의의 조합을 포함할 수 있다. 일부 구현예에서, 진균 세포는 관심 폴리뉴클레오티드를 갖는 공여체 DNA를 추가로 포함한다. 공여체 DNA에서 관심 폴리뉴클레오티드는 표적 부위(예를 들어, 상동성 재조합에 의해 게놈에 삽입)의 게놈에 (또는 근처에) 존재할 수 있으며, 표적 부위에서 Cas/가이드 폴리뉴클레오티드 복합체의 작용에 의해 유도되는 과정.
특정 구현예에서, 재조합 폴리뉴클레오티드에 의해 인코딩된 Cas 엔도뉴클레아제는 Cas9 엔도뉴클레아제이다. Cas 엔도뉴클레아제 및 이의 기능적 단편을 포함하지만 이들에 한정되지 않는 임의의 Cas9 엔도뉴클레아제가 다음 박테리아 종으로부터 인코딩될 수 있다: 스트렙토코쿠스 종(예컨대, S. 파이오제네스, S. 뮤탄스 및 S. 더모필루스), 캄피로박테르 종(예컨대, C. 제주니), 나이세리아 종(예컨대, N. 메닝기티데스), 프란시셀라 종(예컨대, F. 노비시다) 및 파스튜렐라 종(예컨대, P. 물토시다)가 있다(본원에 참고로 통합된 Fonfara 등, Nucleic Acids Res., 2013, pages 1-14에 기재된 Cas9 엔도뉴클레아제를 참조). 일부 구현예에서, Cas 엔도뉴클레아제 유전자를 인코딩하는 폴리뉴클레오티드는 사상균 숙주 세포에서 발현을 위해 최적화된 폴리뉴클레오티드, 예를 들어 서열 번호 44(S. 파이오제네스 Cas9 엔도뉴클레아제의 사상균 세포 코돈 최적화 버전) 또는 서열 번호 7(서열 번호 44를 포함하고 또한 N- 및 C- 말단 NLS 서열을 포함)에 나타낸 폴리뉴클레오티드이다. 서열 번호 44 또는 서열 번호 7의 동의어 변이체를 포함하는 추가적인 코돈-최적화된 Ca9 유전자가 사용될 수 있다. 전술한 바와 같이, Cas 엔도뉴클레아제는 Cas 엔도뉴클레아제가 세포질에서 핵으로 그 작용 부위까지, 즉 세포의 핵 내로 이동하는 것을 향상시키는 기능을 하는 하나 이상의 핵 위치 신호에 작동 가능하게 연결될 수 있다. SV40 핵 위치 신호, T. 레에세이 blr2 (청색광 조절인자 2) 유전자로부터 유래된 핵 위치 신호, 또는 이 둘의 조합을 포함하는 임의의 편리한 핵 위치 신호가 사용될 수 있다.
필라 아스코마이코타(phyla Ascomycota), 바시디오마이코타(Basidiomycota), 키트리디오마이코타Chytridiomycota), 및 자이고마이코타(Zygomycota)(Hawksworth 등, In, Ainsworth and Bisby 's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK에서 정의됨)뿐만 아니라 오오마이코타(Oomycota)(Hawksworth 등, supra에서 인용) 및 모든 불완전 균류(mitosporic fungi)(Hawksworth 등, supra)로부터 유래된 진균 숙주 세포를 포함하는 매우 다양한 사상균 숙주 세포들 중 임의의 것이 본 발명에서 사용된다. 특정 구현예에서, 진균 숙주 세포는 효모 세포, 예컨대, 칸디다, 한세누라, 클루베로마이세스, 피키아, 사카로마이세스, 스키오사카로마이세스 또는 야로위아 세포이다. 효모 종은 다음을 포함하지만 이에 한정되지 않는다: 사카로마이세스 칼스베르겐시스(Saccharomyces carlsbergensis), 사카로마이세스 세레비시애(Saccharomyces cerevisiae), 사카로마이세스 디아스타티쿠스(Saccharomyces diastaticus), 사카로마이세스 도우글라시(Saccharomyces douglasii), 사카로마이세스 클루베리(Saccharomyces kluyveri), 사카로마이세스 노르벤시스(Saccharomyces norbensis), 사카로마이세스 오비포르미스(Saccharomyces oviformis), 클루베로마이세스 락티스(Kluyveromyces lactis), 및 야로위아 리폴리티카(Yarrowia lipolytica) 세포. 추가의 구현예에서, 진균 세포는 트리코데르마, 페니실리움, 아스퍼질러스, 휴미콜라, 크리소스포리움, 푸사리움, 뉴로스포라, 마이셀리오프토라, 더모마이세스, 하이포크레아 및 에메리셀라의 종을 포함하지만 이에 한정되지 않는 사상균 세포이다. 예를 들어, 사상균 트리코데르마 레에세이, P. 크리소제넘, M. 더모필라, 더모마이세스 라누지노수스, A. 오리재 및 A. 나이거가 본 개시의 양태에서 사용된다.
위에서 상세히 설명한 바와 같이, 본 발명은 일반적으로 사상균 세포의 게놈에서 관심 표적 부위를 변형시키는데 유용한 방법 및 조성물에 관한 것이다. 관심 대상인 특정 표적 부위는 이러한 방법 및 조성물의 사용자에 의해 결정되며, 프로모터, 조절 서열, 종결인자 서열, 조절 요소 서열, 스플라이스 부위, 코딩 서열, 폴리유비퀴티네이션 부위, 인트론 부위, 및 인트론 증진 모티프를 포함하는 관심 유전자의 영역 내에 위치한 부위를 포함한다. 아울러, 아세틸 에스테라아제, 아미노펩티다아제, 아밀라아제, 아라비나아제, 아라비노푸라노시다아제, 카복시펩티다아제, 카탈라아제, 셀룰라아제, 키티나아제, 큐티나아제, 데옥시리보뉴클레아제, 에피메라아제, 에스테라아제, α- 갈락토시다아제, β- 갈락토시다아제, α- 글루카나아제, 글루카닐라아제, 엔도-β-글루카나아제, 글루코아밀라아제, 글루코스 옥시다아제, α-글루코시다아제, β-글루코시다아제, 글루쿠로니다아제, 헤미셀룰라아제, 헥소오스 옥시다아제, 하이드로라아제, 인베르타제, 이소메라제, 락카아제, 리파아제, 리아제, 만노시다제, 옥시다아제, 옥시도레덕타아제, 펙테이트 리아제, 펙틴 아세틸 에스테라아제, 펙틴 디폴리머라아제, 펙틴 메틸 에스테라아제, 펙틴 분해 효소, 페록시다아제, 페놀옥시다아제, 피타아제, 폴리갈락투로나아제, 프로테아제, 람노 - 갈락투로나아제, 리보뉴클레아제, 트랜스퍼라아제, 수송 단백질, 트랜스글루타미나아제, 크실라나아제, 헥소오스 옥시다아제 및 이들의 조합을 포함하지만 이들에 한정되지 않는 관심 대상인 임의 유전자들이 사용자에 의해 선택될 수 있다.
재조합 폴리뉴클레오티드
본 발명의 양태는 본원에 기재된 방법 및 조성물에 사용되는 재조합 폴리뉴클레오티드에 관한 것이다.
본 발명의 구현예는 Cas 엔도뉴클레아제를 인코딩하는 진균 세포에 최적화된 뉴클레오티드 서열에 작동 가능하게 연결된 프로모터 서열을 갖는 재조합 폴리뉴클레오티드 DNA 작제물을 포함한다. 본 개시의 구현예는 Cas 엔도뉴클레아제를 인코딩하는 진균 세포에 최적화된 뉴클레오티드 서열 또는 Cas 엔도뉴클레아제를 인코딩하는 박테리아 세포에 최적화된 뉴클레오티드 서열에 작동 가능하게 연결된 프로모터 서열을 갖는 재조합 폴리뉴클레오티드 DNA 작제물을 포함한다. 전술한 바와 같이, 진균 세포에 최적화된 뉴클레오티드 서열 및 박테리아 세포에 최적화된 뉴클레오티드 서열에 인코딩된 Cas 엔도뉴클레아제는 가이드 RNA와 복합체 형성시에 표적 부위에서 작용할 수 있다. 임의의 Cas 엔도뉴클레아제는 Cas9 엔도뉴클레아제, 예를 들어 S. 피오게네스로부터의 Cas9를 포함하지만 이에 한정되지 않는, 최적화된 뉴클레오티드 서열에 의해 인코딩될 수 있다. 특정 구현예에서, 진균 세포에 최적화된 뉴클레오티드 서열은 사상균 세포에서의 발현에 위해 최적화된다. 예를 들어, 사상균 세포에 최적화된 서열은 Cas9 엔도뉴클레아제를 인코딩할 수 있고 서열 번호 44에 도시된 뉴클레오티드 서열(100% 동일성)을 포함하거나 Cas9 엔도 뉴클레아제를 인코딩할 수 있고, 서열 번호 44와 적어도 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 동일한 뉴클레오티드 서열을 포함한다. 특정 구현예에서, 박테리아 세포에 최적화된 뉴클레오티드 서열은 대장균 세포에서의 발현을 위해 최적화된다. 예를 들어, 대장균 세포에 최적화된 서열은 Cas9 엔도뉴클레아제를 인코딩할 수 있고 서열 번호 65에 도시된 뉴클레오티드 서열(100% 동일성)을 포함하거나 Cas9 엔도뉴클레아제를 인코딩할 수 있고, 서열 번호 65와 적어도 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 동일한 뉴클레오티드 서열을 포함한다.
본 개시의 구현예들은 추가로 트리코데르마 종 RNA 중합효소 III에 의해 구동된 프로모터에 관한 것이다. RNA 폴 III 유도 프로모터로부터 RNA 폴 III에 의한 유전자의 전사는 RNA 폴 II 의존 프로모터로부터의 RNA 중합효소 II에 의한 전사시에 발생하는 5 '캡 구조 또는 폴리아데닐화의 첨가로 이어지지 않는다. 아래의 실시예에서 기술된 바와 같이, 전사 종결인자 서열뿐만 아니라 U6 유전자와 관련된 T. 레에세이에서 RNA 폴 III에 의해 유도된 프로모터 서열을 확인하였다. 전체 프로모터 서열은 서열 번호 40에 기재되어 있고, 종결인자 서열은 서열 번호 43에 기재되어 있다. 또한, U6 유전자 RNA 폴 III 구동 프로모터의 보다 짧은 버전이 확인되었고 서열 번호 41에 제시되어 있다. 따라서, 본 발명의 양태는 RNA 폴 III 유도된 프로모터로서 기능하고 서열 버호 40 또는 서열 번호 41과 적어도 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% 또는 100%까지 동일한 뉴클레오티드 서열을 갖는 프로모터를 포함한다. 이 RNA 폴 III 유도 프로모터 서열은 임의의 이형 관심 서열을 발현하는데 사용된다. 따라서, 본 개시의 양태는 이종 관심 서열에 작동 가능하게 연결된 T. 레에세이 유래 RNA 폴 III 구동 프로모터 서열을 갖는 재조합 폴리뉴클레오티드 서열을 포함한다. 특정 구현예에서, 이종 서열은 가이드 폴리뉴클레오티드, 예를 들어 tracrRNA, crRNA 또는 단일 가이드 RNA를 인코딩하는 서열이다. 관심 대상인 특정 게놈 부위를 표적으로 삼은 가이드 RNA 인코딩 폴리뉴클레오티드는 실시예에 상세히 기술되어 있고 서열 번호 2 내지 6을 포함한다. 특정 구현예에서, 재조합 폴리뉴클레오티드는, 예를 들어 관심 이종 서열의 하류 부위(여기서, 하류 "는 당해 분야에서 통상적인 전사 방향을 지칭한다)에서 관심 이종 서열(RNA 폴 III 프로모터에 작동 가능하게 연결된)에 작동 가능하게 연결된 전사 종결인자 서열을 포함한다. 일부 구현예에서, 종결인자 서열은 서열 번호 43으로 도시된 폴리뉴클레오티드 서열 또는 이의 기능적 변이체를 포함한다. 따라서, 특정 구현예에서, 재조합 폴리뉴클레오티드는 종결인자에 작동 가능하게 연결된 관심 이종 서열(예를 들어, 가이드 RNA)에 작동 가능하게 연결된 RNA 폴 III 프로모터를 포함한다.
본원에 개시된 조성물 및 방법의 비제한적인 예 또는 구현예는 하기와 같다:
1. 사상균 세포의 게놈좌와 공여체 DNA를 상동성 재조합하기 위한 방법으로서: 상기 방법은:
a) 사상균 세포의 게놈좌에 대한 상동성을 가진 도메인을 포함하는 Cas 엔도뉴클레아제, 가이드 RNA, 및 공여체 DNA를 사상균 세포군에 도입하는 단계로서, Cas 엔도뉴클레아제 및 가이드 RNA는 Cas 엔도뉴클레아제가 사상균 세포의 게놈좌 내 또는 사상균 세포의 게놈좌 근처의 표적 부위에서 작용할 수 있게 하는 복합체를 형성할 수 있는 단계; 및
b) 게놈좌와 공여체 DNA의 상동성 재조합이 나타난 군으로부터 적어도 하나의 사상균 세포를 확인하는 단계를 포함하고,
Cas 엔도뉴클레아제, 가이드 RNA 또는 둘 모두는 사상균 세포군으로 일시적으로 도입되는, 방법.
2. 구현예 1에 있어서, 사상균 세포들 내 표적 부위에서 비상동성 말단 결합(NHEJ) 메커니즘은 활성화되지 않거나, 비기능적이거나 감소되는, 방법.
3. 구현예 2에 있어서, 사상균 세포들 내 비상동성 말단 결합(NHEJ) 경로는 하나 이상의 비기능적 또는 활성- 감소 성분들을 포함하는, 방법.
4. 구현예 3에 있어서, 하나 이상의 비기능적 또는 활성-감소 성분은 ku80, ku70, rad50, mre11, xrs2, lig4, xrs, 및 이의 조합들로 구성되는 군으로부터 선택되는, 방법.
5. 구현예 4에 있어서, 하나 이상의 비기능적 또는 활성-감소 성분은 ku80인, 방법.
6. 구현예 1 내지 5 중 어느 하나에 있어서, Cas 엔도뉴클레아제는 Cas 닉카아제인, 방법.
7. 구현예 1 내지 5 중 어느 하나에 있어서, Cas 엔도뉴클레아제는 Cas9 엔도뉴클레아제 또는 이의 변이체인, 방법.
8. 구현예 7에 있어서, Cas9 엔도뉴클레아제 또는 이의 변이체는 스트렙토코쿠스 종(Streptococcus sp.), S. 파이오제네스(pyogenes), S. 뮤탄스(mutans) 및 S. 더모필루스(thermophilus), 캄피로박테르 종(Campylobacter sp.), C. 제주니(jejuni), 나이세리아 종(Neisseria sp.), N. 메닝기티데스(meningitides), 프란시셀라 종(Francisella sp.), F. 노비시다(novicida), 및 파스튜렐라 종(Pasteurella sp.), P. 물토시다(multocida)로 구성되는 군으로부터 선택되는 종으로부터 전장 Cas9 또는 이의 기능적 단편을 포함하는, 방법.
9. 구현예 8에 있어서, Cas9 엔도뉴클레아제 또는 이의 변이체는 서열 번호 45 및 서열 번호 48 내지 53 중 어느 하나와 적어도 70% 동일성을 갖는 아미노산 서열을 포함하는, 방법.
10. 구현예 1 내지 9 중 어느 하나에 있어서, 공여체 DNA는 관심 폴리뉴클레오티드 서열을 포함하고 게놈좌에서 상동성 재조합은 게놈좌에서 관심 폴리뉴클레오티드 서열의 삽입을 초래하는, 방법.
11. 구현예 1 내지 10 중 어느 하나에 있어서, 상기 도입 단계는 Cas 엔도뉴클레아제용 발현 카세트를 포함하는 DNA 작제물을 사상균 세포들 내로 도입하는 단계를 포함하는, 방법.
12. 구현예 1 내지 11 중 어느 하나에 있어서, 상기 도입 단계는 가이드 RNA용 발현 카세트를 포함하는 DNA 작제물을 사상균 세포들 내로 도입하는 단계를 포함하는, 방법.
13. 구현예 1 내지 12 또는 구현예 48 내지 54 중 어느 하나에 있어서, 상기 도입 단계는 선택 가능 마커를 인코딩하는 서열을 포함하는 DNA 작제물을 사상균 세포들 내로 도입하는 단계를 포함하는 방법.
14. 구현예 13에 있어서, DNA 작제물은 선택 가능 마커를 인코딩하는 서열 및 공여체 DNA를 모두 포함하는, 방법.
15. 구현예 1 내지 14 중 어느 하나에 있어서, 상기 도입 단계는 Cas 엔도뉴클레아제를 인코딩하는 서열, 가이드 RNA를 인코딩하는 서열, 선택가능 마커를 인코딩하는 서열, 및 공여체 DNA를 포함하는 DNA 작제물을 사상균 세포들 내로 도입하는 단계를 포함하는, 방법.
16. 구현예 11 내지 15 중 어느 하나에 있어서, DNA 작제물은 선형 DNA 작제물인, 방법.
17. 구현예 11 내지 15 중 어느 하나에 있어서, DNA 작제물은 원형 DNA 작제물인, 방법.
18. 구현예 11 및 구현예 15 내지 17 중 어느 하나에 있어서, Cas 엔도뉴클레아제용 발현 카세트 또는 Cas 엔도뉴클레아제를 인코딩하는 서열은 사상균 세포에서 발현을 위해 최적화된 Cas 코딩 서열을 포함하는, 방법.
19. 구현예 18에 있어서, Cas 코딩 서열은 서열 번호 44와 적어도 70% 동일한 폴리뉴클레오티드 서열을 포함하는 Cas9 코딩 서열인, 방법.
20. 구현예 1 내지 10, 구현예 12 내지 14, 및 구현예 16 및 17 중 어느 하나에 있어서, 상기 도입 단계는 Cas 엔도뉴클레아제를 사상균 세포들 내로 직접 도입하는 단계를 포함하는, 방법.
21. 구현예 1 내지 11, 구현예 13, 14, 및 구현예 16 내지 20 중 어느 하나에 있어서, 상기 도입 단계는 가이드 RNA를 사상균 세포들 내로 직접 도입하는 단계를 포함하는, 방법.
22. 구현예 1 내지 21 중 어느 하나에 있어서, Cas 엔도뉴클레아제는 핵 위치 신호에 작동 가능하게 연결되는, 방법.
23. 구현예 1 내지 22 중 어느 하나에 있어서, 사상균 세포는 유미코티나 또는 페지조미코티나 사상균 세포인, 방법.
24. 구현예 1 내지 23 중 어느 하나에 있어서, 사상균 세포는 트리코데르마, 페니실리움, 아스퍼질러스, 휴미콜라, 크리소스포리움, 푸사리움, 마이셀리오프토라, 뉴로스포라, 하이포크레아, 및 에메리셀라로 구성되는 군으로부터 선택되는, 방법.
25. 구현예 1 내지 24 중 어느 하나에 있어서, 표적 부위는 오픈 리딩 프레임, 프로모터, 조절 서열, 종결인자 서열, 조절 요소 서열, 스플라이스 부위, 코딩 서열, 폴리유비퀴티네이션 부위, 인트론 부위, 및 인트론 증진 모티프로 구성되는 군으로부터 선택되는 관심 유전자의 영역 내에 위치되는, 방법.
26. 구현예 1 내지 25 중 어느 하나에 있어서, 상동성 재조합은 표적 부위에서 또는 표적 부위 근처에서 DNA 서열의 변형을 초래하고, 상기 변형은 하나 이상의 뉴클레오티드의 결실, 하나 이상의 뉴클레오티드의 삽입, 관심 단백질을 인코딩하는 발현 카세트의 삽입, 하나 이상의 뉴클레오티드의 치환, 및 이들의 임의 조합으로 구성되는 군으로부터 선택되는, 방법.
27. 구현예 1 내지 26 중 어느 하나에 있어서, 상기 확인 단계는 상동성 재조합 또는 변형을 선택하거나 선별하는 조건 하에서 단계 (a)로부터 세포군을 배양하는 단계를 포함하는, 방법.
28. 구현예 1 내지 27 중 어느 하나에 있어서, 상기 확인 단계는 불안정한 형질전환체를 선별하는 조건 하에서 단계 (a)로부터 세포군을 배양하는 단계를 포함하는, 방법.
29. 구현예 1 내지 28 중 어느 하나에 있어서, 상기 도입 단계는 선택가능 마커를 인코딩하는 서열 및 공여체 DNA를 포함하는 DNA 작제물을 사상균 세포들 내로 도입하는 단계를 포함하고, 상기 확인 단계는 선택가능 마커를 잃었지만 아직은 공여체 DNA를 보유하고 있는 불안정한 형질전환체를 선별하는 조건 하에서 단계 (a)로부터 세포군을 배양하는 단계를 포함하는, 방법.
30. 구현예 1 내지 29 중 어느 하나의 방법으로 생산된 재조합 사상균 세포.
31. Cas 엔도뉴클레아제용 발현 카세트를 포함하는 제1 재조합 DNA 작제물을 포함하는 재조합 사상균 세포.
32. 구현예 30 또는 31에 있어서, 재조합 사상균 세포는 NHEJ 경로에 하나 이상의 비기능성 성분 또는 활성-감소 성분을 포함하는, 재조합 사상균 세포.
33. 구현예 32에 있어서, NHEJ 경로의 하나 이상의 성분은 ku80, ku70, rad50, mre11, xrs2, lig4, 및 xrs로 구성되는 군으로부터 선택되는, 재조합 사상균 세포.
34. 구현예 33에 있어서, 사상균 세포는 ku80의 발현 및/또는 활성을 저해하는 유전자 변형을 포함하는, 재조합 사상균 세포.
35. 구현예 31에 있어서, Cas 엔도뉴클레아제는 Cas 닉카아제인, 재조합 사상균 세포.
36. 구현예 30 내지 35 중 어느 하나에 있어서, 가이드 RNA용 발현 카세트를 포함하는 제2 재조합 DNA 작제물을 추가로 포함하며, 여기서 가이드 RNA 및 Cas 엔도뉴클레아제는 Cas 엔도뉴클레아제가 재조합 사상균 세포의 게놈 내 표적 부위에서 작용할 수 있게 하는 복합체를 형성할 수 있는, 재조합 사상균 세포.
37. 구현예 30 내지 36 중 어느 하나에 있어서, 관심 폴리뉴클레오티드를 포함하는 공여체 DNA를 추가로 포함하는, 재조합 사상균 세포.
38. 구현예 30 내지 37 중 어느 하나에 있어서, Cas 엔도뉴클레아제는 Cas9 엔도뉴클레아제 또는 이의 변이체인, 재조합 사상균 세포.
39. 구현예 38에 있어서, Cas9 엔도뉴클레아제 또는 이의 변이체는 서열 번호 45 및 서열 번호 48 내지 53 중 어느 하나와 적어도 70% 동일성을 갖는 아미노산 서열을 포함하는, 재조합 사상균 세포.
40. 구현예 31 내지 39 중 어느 하나에 있어서, Cas 엔도뉴클레아제용 발현 카세트는 서열 번호 44와 적어도 70% 동일한 폴리뉴클레오티드 서열을 포함하는, 재조합 사상균 세포.
41. 구현예 31 내지 40 중 어느 하나에 있어서, Cas 엔도뉴클레아제용 발현 카세트는 사상균 세포에서의 발현을 위해 최적화된 Cas 엔도뉴클레아제 유전자를 포함하는, 재조합 사상균 세포.
42. 구현예 31 내지 41 중 어느 하나에 있어서, Cas 엔도뉴클레아제는 핵 위치 신호에 작동 가능하게 연결되는, 재조합 사상균 세포.
43. 구현예 42에 있어서, 핵 위치 신호는 SV40 핵 위치 신호(서열 번호 46), T. 레에세이 blr2(청색광 조절자 2) 유전자로부터 유래된 핵 표적화 신호(서열 번호 47), 및 이 둘의 조합으로 구성되는 군으로부터 선택되는, 재조합 사상균 세포.
44. 구현예 30 내지 43 중 어느 하나에 있어서, 사상균 세포는 트리코데르마, 페니실리움, 아스퍼질러스, 휴미콜라, 크리소스포리움, 푸사리움, 마이셀리오프토라, 뉴로스포라, 하이포크레아, 및 에메리셀라로 구성되는 군으로부터 선택되는 사상균 세포인, 재조합 사상균 세포.
45. Cas9 엔도뉴클레아제 또는 이의 변이체를 인코딩하는 사상균 세포에 최적화된 폴리뉴클레오티드 서열에 작동 가능하게 연결되는 프로모터를 포함하는, 재조합 DNA 작제물.
46. 구현예 45에 있어서, 사상균 세포에 최적화된 폴리뉴클레오티드 서열은 서열 번호 44와 적어도 70% 동일한, 재조합 DNA 작제물.
47. 가이드 RNA용 발현 카세트는 유아스코마이세테(Euascomycete) 또는 페지조마이세테(Pezizomycete)에서 기능적인 DNA 중합효소 III 의존성 프로모터를 포함하고, 여기서 상기 프로모터는 가이드 RNA를 인코딩하는 DNA에 작동 가능하게 연결되는, 구현예 12의 방법.
48. 구현예 47에 있어서, 프로모터는 트리코데르마 U6 snRNA 유전자로부터 유래되는, 방법.
49. 구현예 48에 있어서, 프로모터는 서열 번호 40 또는 41에 적어도 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 또는 99% 동일성을 갖는 뉴클레오티드 서열을 포함하는, 방법.
50. 구현예 49에 있어서, 프로모터는 서열 번호 40 또는 41을 포함하는, 방법.
51. 구현예 12 및 구현예 47 내지 50 중 어느 하나에 있어서, 가이드 RNA용 발현 카세트는 트리코데르마 U9 snRNA 유전자로부터의 인트론 서열을 갖는 가이드 RNA-인코딩 DNA를 포함하는, 방법.
52. 구현예 51에 있어서, 트리코데르마 U9 snRNA 유전자로부터 유래된 인트론 서열은 서열 번호 42와 적어도 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 또는 99% 동일성을 갖는 뉴클레오티드 서열을 포함하는, 방법.
53. 구현예 52에 있어서, 트리코데르마 U9 snRNA 유전자로부터 유래된 인트론 서열은 서열 번호 42의 서열을 포함하는, 방법.
실시예
다음 실시예들에서, 달리 언급되지 않으면, 부(parts) 및 백분율은 중량에 의하고, 도(degree)는 섭씨이다. 이 실시예들은 본 개시의 구현예들을 나타내면서 단지 예시로 제공되는 것임을 이해해야 한다. 상기 논의 및 이들 실시예로부터, 당업자는 다양한 용도 및 조건에 적응시키기 위해 본 개시의 다양한 변형 및 수정을 할 수 있다. 이러한 변형은 또한 첨부된 청구항의 범위 내에 속하는 것으로 의도된다.
섹션 A: 발현 벡터에 의한
Cas
/가이드 RNA의 도입
실시예
1: T.
레에세이
U6
snRNA
유전자의 확인
RNA 중합 효소 II에 의존적인 프로모터를 사용함으로써 초래될 5’ 캡 구조 또는 폴리아데닐화를 부가하지 않고서 T. 레에세이에서 가이드 RNA를 생산하기 위해서는 RNA 중합 효소 III 유도 프로모터가 바람직하다. 그러나, T. 레에세이에서 기능적인 RNA 중합 효소 III 의존성 프로모터는 기술되지 않았다. 사카로마이세스 세레비지애(Saccharomyces cerevisiae) snr52 유전자, 인간 U6 snRNA 유전자 또는 옥수수 U6 snRNA 유전자의 5 '상류 영역을 포함하여 다른 종으로부터 공지된 RNA 중합 효소 III 의존성 프로모터가 T. 레에세이에서 기능하는 능력을 시험받은 것으로 간주하였다.
보다 바람직한 것은 RNA 중합 효소 III 의존성 프로모터로서 작용할 천연 T. 레에세이 서열을 확인하는 것이었다. BLAST 알고리즘을 사용하여 T. 레에세이 v2 게놈 서열 (www.jgi.doe.gov)을 검색하기 위해 인간 U6 소형 핵 RNA(snRNA, 유전자은행 수탁번호 M14486)를 인코딩하는 DNA 서열을 사용하였다. T. 레에세이 DNA 서열의 짧은 영역이 인간 서열과 유사한 것으로 확인되었다. 주변 DNA 서열의 조사 및 효모, 특히 쉬조사카로마이세스 프롬베(Schizosaccharomyces pombe)의 U6 유전자와의 비교(Marck 등, 2006, Nucleic Acids Research 34 : 1816-1835)로 T. 레에세이 U6 유전자의 많은 특징을 추정하여 확인할 수 있었다(서열 번호 1, 하기에 나타냄). 전사된 서열의 시작과 종결인자는 상류 TATA 박스와 동일하게 확인되었다. 인트론은 전사된 영역을 분명히 방해하고, 가능한 A-박스 및 B-박스 프로모터 요소는 전사된 영역 내에서 인지될 수 있으며, 후자는 인트론 내에서 인식될 수 있다. (도 1 참조).
AAAAAACACTAGTAAGTACTTACTTATGTATTATTAACTACTTTAGCTAACTTCTGCAGTACTACCTAAGAGGCTAGGGGTAGTTTTATAGCAGACTTATAGCTATTATTTTTATTTAGTAAAGTGCTTTTAAAGTAAGGTCTTTTTTATAGCACTTTTTATTTATTATAATATATATTATATAATAATTTTAAGCCTGGAATAGTAAAGAGGCTTATATAATAATTTATAGTAATAAAAGCTTAGCAGCTGTAATATAATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATCGCCTTCGGGCATTTGGTCAATTTATAACGATACAGGTTCGTTTCGGCTTTTCCTCGGAACCCCCAGAGGTCATCAGTTCGAATCGCTAACAGGTCAACAGAGAAGATTAGCATGGCCCCTGCACTAAGGATGACACGCTCACTCAAAGAGAAGCTAAACATTTTTTTTCTCTTCCAAGTCGTGATGGTTATCTTTTTGCTTAGAGAATCTATTCTTGTGGACGATTAGTATTGGTAAATCCCTGCTGCACATTGCGGCGGATGGTCTCAACGGCATAATACCCCATTCGTGATGCAGCGGTGATCTTCAATATGTAGTGTAATACGTTGCATACACCACCAGGTTCGGTGCCTCCTGTATGTACAGTACTGTAGTTCGACTCCTCCGCGCAGGTGGAAACGATTCCCTAGTGGGCAGGTATTTTGGCGGGGTCAAGAA (서열 번호 1)
실시예
2: T.
레에세이
유전자를 표적으로 하기 위한
sgRNA
서열
단일 가이드 RNA(sgRNA) 분자가 스트렙토코쿠스 파이오제네스 Cas9 단백질과 상호 작용하여 이 엔도뉴클레아제를 생체 내에서 진핵 생물 게놈의 특정 유전자좌에 표적으로 할 수 있는 것으로 나타났다. sgRNA는 스트렙토코쿠스 파이오제네스 II 형 CRISPR-Cas 시스템의 구성 요소인 것으로 자연적으로 관찰되는 tracrRNA와 crRNA 사이의 융합체로 설계된 하이브리드 분자이다(Gasiunas 등 (2012) Proc. Natl. Acad. Sci.USA 109:E2579-86, Jinek 등 (2012) Science 337:816-21, Mali 등 (2013) Science 339:823-26, and Cong 등 (2013) Science 339:819-23). sgRNA의 처음 20개 뉴클레오티드는 게놈의 표적 부위에 상보적이다. 추가 서열인 포토스페이서 인접 모티프(PAM)도 sgRNA 상보 영역에 인접한 게놈의 표적 부위에 존재해야 한다. S. 파이오제네스 Cas9의 경우, PAM은 서열 NGG(N은 A, G, C 또는 T)를 갖는다.
이 실험에 사용된 sgRNA의 서열은 하기에 도시되고, 여기서 표적 부위에 상보적인 것으로 설계된 20개의 뉴클레오티드가 N 잔기(서열 번호 2)(N = A, G, C 또는 U)로 도시된다.
NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
sgRNA는 T. 레에세이 게놈 내 상이한 유전자좌를 표적으로 하도록 설계되었다. 표적 부위 1(TS 1)로 지정된 부위에서 T. 레에세이 ad3A 유전자(포스포리보실아미도이미다졸-숙시노카복사미드 신타아제)를 표적으로 하기 위한 (gAd3A TS1로 불리는) sgRNA의 서열이 아래에 도시된다(서열 번호 3). T. 레에세이 게놈 서열에 상보적인 20개 뉴클레오티드 영역이 소문자로 도시된다.
guccucgagcaaaaggugccGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
표적 부위 2(TS 2)로 지정된 부위에서 T. 레에세이 gla1(글루코아밀라아제) 유전자를 표적으로 하기 위한 (gTrGA TS2로 불리는) sgRNA의 서열이 아래에 도시된다(서열 번호 4). T. 레에세이 게놈 서열에 상보적인 20개 뉴클레오티드 영역이 소문자로 도시된다.
guucagugcaauaggcgucuGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
표적 부위 11(TS 11)로 지정된 부위에서 T. 레에세이 gla1(글루코아밀라아제) 유전자를 표적으로 하기 위한 (gTrGA TS11로 불리는) sgRNA의 서열이 아래에 도시된다(서열 번호 5). T. 레에세이 게놈 서열에 상보적인 20개 뉴클레오티드 영역이 소문자로 도시된다.
gccaauggcgacggcagcacGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
표적 부위 6(TS 6)으로 지정된 부위에서 T. 레에세이 pyr2(오로테이트 포스포리보실트란스페라아제(orotate phosphoribosyltransferase)) 유전자를 표적으로 하기 위한 (gPyr2 TS6으로 불리는) sgRNA의 서열이 아래에 도시된다(서열 번호 6). T. 레에세이 게놈 서열에 상보적인 20개 뉴클레오티드 영역이 소문자로 도시된다.
gcacagcgggaugcccuuguGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
실시예
3: T.
레에세이에서
발현을 위한
Cas9
DNA 및 단백질 서열
T. 레에세이에서의 발현을 위하여 NLS 서열을 포함하여 코돈이 최적화된 스트렙토코쿠스 파이오제네스 Cas9-인코딩 유전자를 설계하고, 합성하고 검사하였다(서열 번호 7). 인코딩된 단백질(서열 번호 8)은 N 말단 SV40 핵 위치 신호(NLS; 서열 번호 46) 및 T. 레에세이 blr2(청색 조절인자 2) 유전자로부터 유래된 C 말단 NLS(서열 번호 47; 둘 모두 아래의 서열 번호 8에 밑줄쳐져 있다)를 갖는다.
서열 번호 7:
acgcatcgacctgagccagctgggcggagacaagaagaagaagctcaagctctag
서열 번호 8:
실시예
4:
Cas9
발현 벡터의 구성
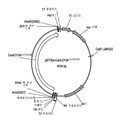
플랭킹 attL1 및 attL2 부위 사이에 있어 InvitrogenTM Gateway® 클로닝 기술(Thermo Fisher Scientific Inc., Grand Island, NY)에 의해 적합한 발현 벡터로의 전사가 가능하도록 상기에 도시된 Cas9를 인코딩하는 합성 DNA 서열을 pENTR/D-TOPO에 삽입하였다. 다음과 같은 특징을 포함하는 게이트웨이 호환 가능 발현 벡터, pTrex2gHyg를 사용할 수 있었다: 게이트웨이 클로닝 부위에 의해 분리된 T. 레에세이 pki1(피루베이트 키나아제) 유전자로부터의 프로모터 영역 및 T. 레에세이 cbh1(셀로비오하이드롤라아제 I) 유전자로부터의 종결인자 영역, 뉴로스포라 크라사 (Neurospora crassa ) cpc1(교차 경로 조절 1) 프로모터 영역 및 아스퍼질러스 니두란스 (Aspergillus nidulans ) trpC(글루타민 아미도 트랜스퍼라아제, 인돌글리세롤포스페이트 신타아제 및 포스포리보실란트라닐레이트 이소머라아제 활성을 갖는 3 기능성 단백질) 종결인자 영역에 기능적으로 연결된 박테리아 하이그로마이신 포스포트랜스퍼라아제 유전자, 및 대장균에서의 선택 및 유지를 위한 박테리아 벡터 서열. pTrex2gHygMoCas를 제공하기 위해 게이트웨이 클로닝 절차를 사용하여 cas9 유전자를 pTrex2gHyg에 복제하였다(도 2 참조).
실시예
5:
sgRNA
발현 벡터의 구성
상이한 추정 RNA 중합 효소 III 의존성 프로모터 및 종결인자에 의해 플랭킹된 gAd3A TS1 sgRNA를 인코딩하는 합성 DNA 서열을 수득하였다. 이들 합성 DNA 서열 각각은 또한 어느 한쪽 말단에 제한 효소 인식 부위(EcoRI 및 BamHI)를 갖는다.
다음 서열은 사카로마이세스 세레비지애 snr52 프로모터 및 S. 세레비지애 sup4 종결인자(gAd3A TS1-1로 표시, 서열 번호 9)로 gAd3A TS1 sgRNA(밑줄친)를 인코딩한다:
다음 서열은 T. 레에세이 U6 프로모터 및 종결인자(gAd3A TS1-2로 표시, 서열 번호 10)로 gAd3A TS1 sgRNA(밑줄친)를 인코딩한다:
다음 서열은 T. 레에세이 U6 프로모터, 종결인자 및 인트론(이탤릭체)(gAd3A TS1-3으로 표시, 서열 번호 11)로 gAd3A TS1 sgRNA(밑줄친)를 인코딩한다:
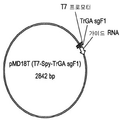
플라스미드 p219M(도 3)은 이의 천연 프로모터 및 종결인자를 포함하는 T. 레에세이 pyr4(오로티딘 모노포스페이트 디카복실라아제(orotidine monophosphate decarboxylase)) 유전자를 함유하는 대장균 벡터이다. 이 벡터를 EcoRI 및 BamHI로 절단하고, 말단을 탈인산화시켰다. 상기 합성 DNA 분자 각각을 EcoRI 및 BamHI로 절단하고 절단된 p219M과 연결하여 sgRNA 발현 카세트 및 pyr4 유전자를 포함하는 일련의 벡터를 생성하였다. 각 벡터는 암호화된 sgRNA의 이름으로 지정되었다(예컨대, p219M gAd3A TS1-1은 gAd3A 발현 카세트를 S. 세레비시애 snr52 프로모터 및 sup4 종결인자와 통합함).
보다 짧은 T. 레에세이 U6 프로모터 영역을 갖는 가이드 RNA 발현 카세트를 합성 DNA로서 수득 하였다. TS11에서의 T. 레에세이 gla1 유전자(서열 번호 12, 인트론 서열은 밑줄쳐진다)를 표적으로 하는 sgRNA에 대한 서열을 포함하는 예가 여기에 제공된다.
상기한 gRNA 발현 카세트를 프라이머 gRNA fwd aflII (5'-
; 서열 번호 13) 및 gRNA rev sfiI(5'-cgtcagggccacgtgggccAAGAGAAAAAAAAGCACCACCGACTCGG; 서열 번호 14)를 사용하여 PCR에 의해 증폭시켰다. 이들 프라이머는
aflII를 가이드 RNA 발현 카세트의 5’ 말단에,
sfiI 부위를 3’ 말단에 부가한다. PCR 생성물을 Qiagen PCR 정제 키트를 사용하여 제조사의 지침에 따라 정제하였다. 그런 다음, PCR 생성물을
SfiI 및
AflII로 절단하고 Qiagen PCR 정제 키트로 다시 세척하였다. 플라스미드 pTrex2g / HygMoCas를
SfiI 및
AflII로 절단하고 Roche Rapid 알칼라인 포스파타아제 키트(Roche Diagnostics Corp., IN)를 사용하여 탈인산화시켰다. 절단된 플라스미드 및 PCR 생성물을 최종적으로 Rapid DNA 리가아제 키트를 사용하여 결찰시켜 pTrex2g/Hyg MoCas gTrGA TS11B를 생성하였다. 다른 sgRNA 발현 카세트를 pTrex2g / Hyg MoCas에 유사한 방식으로 삽입하였다.
실시예
6: 트리코데르마
레에세이에서
Cas9
-
매개된
유전자 불활성화
트리코데르마 레에세이 균주가 두 개의 개별 발현 벡터, 즉 Cas9 생산용과 gRNA 생산용으로 동시에 형질전환되거나 Cas9 및 gRNA 둘 다 발현용 단일 벡터로 형질전환 된 일련의 실험이 하기에 기재되어 있다. 이 실험은 U6 인트론이 gRNA 전사 영역 내에도 존재할 때만 T. 레에세이 U6 유전자의 5’ 상류 영역이 gRNA 전사를 촉진한다는 것을 나타낸다. 실험은 또한 표적 유전자 불활성화가 T. 레에세이 형질전환체에서 높은 효율로 발생할 수 있음을 나타낸다.
ad3A
유전자의 불활성화
4 개의 주요 분비된 셀룰라아제를 인코딩하는 유전자(cbh1 , cbh2 , egl1 및 egl2)가 결실된 공개적으로 이용가능한 RL-P37 균주에서 유래된 트리코데르마 레에 세이 균주를 사용하였다. 이 균주에는 또한 기능적 pyr4 유전자가 결여되어 있다. (US20060003408A1에 기재된 바와 같은) 생물학적 형질전환을 사용하여 동일한 양의 pTrex2gHyg MoCas(도 2) 및 p219M gAd3A TS1-1, p219M gAd3A TS1-2 또는 p219M gAd3A TS1-3 중 어느 하나의 혼합물과 공변형시켰다. 2% 포도당, 100 mg/L 하이그로마이신 B 및 200 mg/L 아데닌을 함유한 보겔(Vogel)의 최소 배지를 갖는 한천 플레이트에서 형질전환체를 선택하였다. 제1 플레이트에서 선택한 후, 형질전환체 콜로니를 동일한 선택 배지의 신선한 플레이트에 집어 넣었다. 제 2 플레이트에서 성장하는 동안, 안정한 및 불안정한 하이그로마이신-내성 형질전환체를 구별하는 것이 가능하였다. 안정한 형질전환체가 더 빨리 자라났고, 콜로니는 윤곽이 부드러웠고 균사체는 더 조밀했다. 불안정한 형질전환체는 느리게 성장하고 밀도가 낮은 균사체가 있었으며 콜로니는 가장자리가 불규칙한 외형을 가졌다. 제2 플레이트에서 성장한 후 형질전환체를 포도당이 있고 하이그로마이신이 없는 보겔(Vogel) 배지에 옮기고 14 mg/L 아데닌을 사용하여 적색/갈색을 나타내는 것들을 선별 검사하여 이들이 아데닌 영양요구성 인자임을 나타냈다. p219M gAd3A TS1-1을 사용하여 5개의 안정한 형질전환체와 23개의 불안정한 형질전환체를 수득했는데, 모두 아데닌 (adenine) 원형질체였다. p219M gAd3A TS1-2를 사용하여 11개의 안정한 형질전환체와 38개의 불안정한 형질전환체를 수득했는데, 11개의 안정한 형질전환체 전부 및 불안정한 형질전환체 중 29개는 아데닌 원형질체였다. p219M gAd3A TS1-3으로 19개의 안정한 형질전환체와 2개의 불안정한 형질전환체를 수득했는데, 모두 아데닌 영양요구인자였다. 분명히, 아데닌 영양요구인자는 sgAd3A TS1의 전사를 조절하는 T. 레에세이 U6 프로모터, 인트론 및 종결인자를 이용하는 gAd3A TS1-3으로만 수득되었다. 아데닌 영양요법은 천연 T. 레에세이 ad3A 유전자좌에서 표적화된 Cas9 절단을 나타낸다. 시험된 gAd3A TS1-3을 갖는 모든 형질전환체는 아데닌 영양요구인자이므로 Cas9 매개 유전자 불활성화가 효율적이라는 결론을 내릴 수 있다.
pTrex2gHyg MoCas 및 p219M gAd3A TS1-3을 갖는 공동 형질전환체의 ad3A 유전자좌에서 돌연변이를 판단하기 위해, 10개의 안정한 아데닌 영양요구성 형질전환체로부터 게놈 DNA를 추출 하였다. 이 DNA를 Cas9 표적 부위에 걸쳐 있거나 표적 부위의 상류 또는 하류에 위치하는 생성물을 생성하도록 설계된 몇몇 상이한 프라이머 쌍을 사용하는 PCR을 위한 주형으로 사용하였다. 제조사의 지시에 따라 PfuUltra II Fusion HS DNA 중합 효소(Agilent Technologies, Santa Clara, CA)를 PCR에 사용하였다. 각각의 경우에 연장 시간은 아래에 설명된 것처럼 PCR 제품의 예상 크기에 대해 제조업체가 제안한 시간이었다. PCR 생성물의 크기는 아가로스 겔 전기 영동으로 평가하였다.
TS1 표적 부위의 5’ 측의 영역을 증폭시키는 Ad3 5' fwd + Ad3 5' rev 프라이머(각각 5'-tgaacacagccaccgacatcagc [서열 번호 15] 및 5'-gctggtgagggtttgtgctattg [서열 번호 16])를 사용하여 모든 형질전환체에서 예상 크기(872 bp)의 PCR 생성물을 수득하였다.
TS1 표적 부위의 5’ 측의 영역을 증폭시키는 Ad3 5' fwd + Ad3a 5005 rev 프라이머(각각 5'-tgaacacagccaccgacatcagc [서열 번호 15] 및 5'-gattgcttgggaggaggacat [서열 번호 17])를 사용하여 모든 형질전환체에서 예상 크기(1214 bp)의 PCR 생성물을 수득하였다.
TS1 표적 부위의 3’ 측의 영역을 증폭시키는 Ad3 3' fwd + Ad3 3' rev 프라이머(각각 5'-cgaggccactgatgaagttgttc [서열 번호 18] 및 5'-cagttttccaaggctgccaacgc [서열 번호 19])를 사용하여 모든 형질전환체에서 예상 크기(904 bp)의 PCR 생성물을 수득하였다.
TS1 표적 부위의 3’ 측의 영역을 증폭시키는 Ad3a 5003 fwd + Ad3mid rev 프라이머(각각 5'-ctgatcttgcaccctggaaatc [서열 번호 20] 및 5'-ctctctatcatttgccaccctcc [서열 번호 21])를 사용하여 모든 형질전환체에서 예상 크기(757 bp)의 PCR 생성물을 수득하였다.
상기 PCR 결과는 게놈 DNA 제제가 Cas9 표적 부위의 상류 또는 하류에서 PCR 생성물을 수득하기에 충분한 품질임을 나타내었다.
ad3A에서 TS1 표적 부위에 걸친 Adfrag fwd + Adfrag rev 프라이머(각각 5'-ctccattcaccctcaattctcc [서열 번호 22] 및 5'-gttcccttggcggtgcttggatc [서열 번호 23])를 사용하여 임의의 형질전환체에 대한 PCR 생성물을 수득할 수 없었다. Cas9 활성으로 인한 큰 크기 변화가 없다고 가정할 때 이 PCR 생성물의 예상 크기는 약 764 bp였다.
ad3A에서 TS1 표적 부위에 걸친 Adfrag fwd + Ad3 3 'rev 프라이머(각각 5'-ctccattcaccctcaattctcc [서열 번호 22] 및 5'-cagttttccaaggctgccaacgc [서열 번호 19])를 사용하여 임의의 형질전환체에 대해 PCR 생성물을 수득할 수 없었다. Cas9 활성으로 인한 큰 크기 변화가 없다고 가정할 때 이 PCR 생성물의 예상 크기는 약 2504 bp였다.
ad3A에서 TS1 표적 부위에 걸친 Ad3a 2k fwd + Ad3a 2k rev 프라이머(각각 5'-ctccattcaccctcaattctcc [서열 번호 24] 및 5'-cagttttccaaggctgccaacgc [서열 번호 25])를 사용하여 임의의 형질전환체에 대해 PCR 생성물을 수득할 수 없었다. Cas9 활성으로 인한 큰 크기 변화가 없다고 가정할 때 이 PCR 생성물의 예상 크기는 약 1813 bp였다.
형질전환체 중 5개는 또한 TS1 표적 부위에 걸친 Adfrag fwd + Ad3 mid rev 프라이머(각각 5'-ctccattcaccctcaattctcc [서열 번호 22] 및 5'-ctctctatcatttgccaccctcc [서열 번호 21])를 사용하여 PCR 생성물을 제공하지 못했다. Cas9 활성으로 인한 큰 크기 변화가 없다고 가정할 때 이 PCR 생성물의 예상 크기는 약 1438 bp였다.
공개된 데이터를 기초로, Cas9가 매개하는 유전자의 불활성화는 일반적으로 표적 부위의 DNA에서 이중 가닥 절단의 오류가 발생하기 쉬운 복구를 포함한다. 최종 결과는 표적 부위에서 작은 결실 또는 삽입(삽입 결실)이다. PCR 분석의 상기 결과는 표적 부위에 걸친 예상 크기의 PCR 생성물을 얻는 것이 불가능하다는 점에서 놀라웠으며, 이는 ad3A의 불활성화가 표적 부위에서 작은 삽입 또는 결실(삽입 결실)에 기인한 것이 아니라는 것을 시사한다. 대신, 이들 데이터는 ad3A의 불활성화가 표적 부위에서의 염색체 재배치 또는 큰 삽입에 의해 야기되었을 가능성과 일치한다.
글루코아밀라아제
(GA) 유전자의 불활성
4개의 주요 분비된 셀룰라아제를 인코딩하는 유전자(cbh1 , cbh2 , egl1 및 egl2)가 결실된 공개적으로 이용가능한 RL-P37 균주에서 유래된 트리코데르마 레에세이 균주를 사용하였다. 이 균주에는 기능적 pyr4 유전자가 결여되어 있다. 동량의 pTrex2gHyg MoCas 및 p219M gTrGATM TS2의 혼합물을 이용한 생물학적 방법을 사용하여 이 균주를 동시에 형질전환하였다. 1% 포도당, 100 ㎍/ml 하이그로마이신 B 및 2 mg/ml 우리딘을 함유하는 보겔 (Vogel)의 최소 배지를 사용하여 한천 플레이트 상에서 형질전환체를 선택하였다. 제1 플레이트에서 선택한 후, 형질전환체 콜로니를 동일한 선택 배지의 새로운 플레이트에 집어 넣었다. 제2 플레이트에서 성장하는 동안, 안정한 및 불안정한 하이그로마이신-내성 형질전환체를 구별하는 것이 가능하였다. 17개의 안정한 형질전환체 및 4개의 불안정한 형질전환체를 수득하였다. 분비된 글루코아밀라아제 존재 여부를 선별하기 위해, 이들 형질전환체를 글루코스가 없고 1% 불용성 전분을 함유한 보겔(Vogel) 한천 플레이트로 옮겼다. 글루코아밀라아제를 분비할 수 있는 콜로니는 잘 자라며 포자를 형성한다. 글루코아밀라아제를 분비할 수 없는 콜로니는 매우 드문 균사체로 자라며 명확하게 구별할 수 있다. 17개의 안정한 형질전환체 중 14개는 글루코아밀라아제를 분비할 수 없었고, 4개의 모든 불안정한 형질전환체는 글루코아밀라아제를 분비하지 않았다.
pTrex2gHyg MoCas 및 p219M gTrGA TS2와의 공동 형질전환체에서 gla1(글루코아밀라아제) 유전자좌에서 돌연변이를 판단하기 위해, 5개의 안정한 글루코아밀라제 비생산 형질전환체로부터 게놈 DNA를 추출하였다. 이 DNA는 Cas9 표적 부위에 걸쳐 있거나 표적 부위의 상류 또는 하류에 위치하는 생성물을 생성하도록 설계된 상이한 프라이머 쌍을 사용하여 PCR을 위한 주형으로 사용되었다. 제조사의 지시에 따라 PfuUltra II Fusion HS DNA 중합 효소(Agilent Technologies)를 PCR에 사용하였다. 각각의 경우에 연장 시간은 아래에 설명된 것처럼 PCR 제품의 예상 크기에 대해 제조업체가 제안한 시간이었다. PCR 생성물의 크기는 아가로스 겔 전기 영동으로 평가하였다.
gla 1에서 TS2 표적 부위에 걸친 glaA + glaB 프라이머(5'- ccgttagttgaagatccttgccg [서열 번호 26] 및 5'- gtcgaggatttgcttcatacctc [서열 번호 27])를 사용하여 임의의 형질전환체에 대한 PCR 생성물을 수득할 수 없었다. Cas9 활성으로 인한 큰 크기 변화가 없다고 가정할 때 이 PCR 생성물의 예상 크기는 약 1371 bp였다.
TS2 표적 부위의 5’ 측의 영역을 증폭시키는 glaA + glaJ 프라이머(각각 5'-ccgttagttgaagatccttgccg [서열 번호 26] 및 5'-tgccgactttgtccagtgattcg [서열 번호 30])를 사용하여 모든 형질전환체에서 예상 크기(364 bp)의 밴드를 수득하였다.
TS2 표적 부위의 3’ 측의 영역을 증폭시키는 glaK + glaB 프라이머(각각 5'-ttacatgtggacgcgagatagcg [서열 번호 31] 및 5'-gtcgaggatttgcttcatacctc [서열 번호 27])를 사용하여 형질전환체 중 4개에서 예상 크기(520 bp)의 밴드를 수득하였다. 형질전환체들 중 하나는 이 프라이머 쌍으로 PCR 생성물을 제공하지 못했다.
표적 Cas9 작용에 의해 gla1 유전자의 불활성화를 입증하고자 하는 별도의 실험을 RL-P37 유래되고 불활성화 pyr4 유전자를 갖는 T. 레에세이 균주를 사용하여 수행하였다. 이 균주의 원형질을 폴리에틸렌 글리콜 매개 절차를 사용하여 pTrex2gHyg MoCas gTrGA TS11로 형질전환시켰다(하기에 기술). 2% 포도당, 2 mg/ml 우리딘, 1.1M 소르비톨 및 100 ug/ml 하이그로마이신 B를 함유하는 Vogel 최소 배지의 한천 평판에서 형질전환체를 선택하였다. 제1 플레이트에서 선택한 후, 형질전환체 콜로니를 소르비톨이 없는 동일한 선택 배지의 새로운 플레이트에 집어 넣었다. 제 2 플레이트에서 성장하는 동안, 안정한 및 불안정한 하이그로마이신-내성 형질전환체를 구별하는 것이 가능하였다. 분비된 글루코아밀라아제 존재 여부를 선별하기 위해, 이들 형질전환체를 글루코스가 없고 1% 불용성 전분을 함유한 보겔(Vogel) 한천 플레이트로 옮겼다. 글루코아밀라아제를 분비하지 않는 B#1, B#2, B#4, B#5 및 B#6으로 명명된 5개의 안정한 형질전환체를 추가 분석을 위해 선택하였다. 이들 형질전환체 각각으로부터 게놈 DNA를 추출하였다.
게놈 DNA를 주형으로써 그리고 TS11 표적 부위에 걸쳐 야생형 gla 1 유전자좌로부터 983 bp의 생성물을 생성하는 gla1repF 및 gla1repR 프라이머(각각 5'- gtgtgtctaatgcctccaccac [서열 번호 32] 및 5'- gatcgtgctagcgctgctgttg [서열 번호 23])를 사용하여 PCR을 수행하였다. PCR 조건은 매 PCR 사이클마다 프라이머 어닐링 온도를 점차적으로 낮추고 표적 부위에 큰 삽입이 있었는지를 결정하기 위해 긴 연장 시간을 포함시켰다. 구체적인 PCR 조건은 다음과 같다.
단계 1: 94℃에서 1분
단계 2: 94℃에서 25초
단계 3: 63℃에서 30초(온도는 사이클마다 0.2℃씩 감소)
단계 4: 70℃에서 8분
단계 2~4를 24회 더 반복
단계 5: 4℃에서 유지
형질전환체 중 2개(B#1 및 B#6)로부터 12 kb를 초과하는 명확한 PCR 생성물을 수득하였는데, 이는 표적 부위에 걸친 DNA 영역에서 11 kb를 초과하는 증가를 나타낸다. 다른 3개의 형질전환체는 아가로스 겔 전기영동에 저 강도 밴드로 나타난 비특이적 PCR 생성물만을 제공했다. B#6로부터 12 kb 초과 PCR 생성물의 서열 분석은 플라스미드 pTrex2gHyg MoCas gTrGA TS11에서 유래된 DNA가 TS11 표적 부위에 삽입되었음을 입증했다.
게놈 DNA 샘플 B#2, B#4 및 B#5 및 프라이머 쌍 1553R 및 1555F(5'-CCGTGATGGAGCCCGTCTTCT [서열 번호 34] 및 5'-CGCGGTGAGTTCAGGCTTTTTC [서열 번호 35])를 각각 사용하여 PCR을 수행하였다. 프라이머 1553R은 표적 부위 11의 3’ 측의 gla1 유전자에 결합한다. 프라이머 1555F는 플라스미드 pTrex2gHyg MoCas gTrGA TS11 상의 하이그로마이신 포스포트랜스퍼라제(hygB) 유전자의 개시 코돈 근처에 결합한다. 위의 것과 동일한 PCR 조건을 사용하였다. 형질전환체 B#4 및 B#5 각각에 대해 4.5 kb 및 6.5의 PCR 생성물이 수득되었다. PCR 생성물은 hygB 유전자를 가진 플라스미드가 gla1 유전자에 삽입된 경우에만 수득되어야 한다. 추측컨대, 형질전환체 B#4 및 B#5 내 삽입된 플라스미드 DNA는 너무 커서 프라이머 gla1repF 및 gla1repR을 사용하여 PCR 생성물을 수득할 수 없었다.
종합하면, PCR 데이터는 글루코아밀라아제 불활성화를 갖는 안정한 하이그로마이신 내성 형질전환체가 gla1 유전자의 표적 부위에 Cas9 및 가이드 RNA 발현 벡터의 큰 세그먼트의 삽입을 통해 생성되었음을 입증하였다.
pyr2
유전자의 불활성화
T. 레에세이 pyr2 유전자 내의 상이한 위치를 표적으로 하는 가이드 RNA 발현 카세트를 포함하는 플라스미드 pTrex2gHyg MoCas의 유도체로 원형질체의 PEG-매개된 형질전환에 의해 T. 레에세이 균주의 형질전환체 QM6a 또는 RL-P37이 생성되었다. 이 유전자의 불활성화는 우리딘 영양요구성을 부여하고 5-플루오로오로트산(5-fluoroorotic acid(FOA))에 대한 내성을 부여한다. 형질전환체는 초기에 하이그로마이신 B를 함유하는 배지에서 선별되었다. 하이그로마이신 B를 함유하는 신선한 한천 플레이트로 옮길 때, 이들은 안정적이거나 불안정한 것으로 기록되었다. 이어서, 형질전환체를 2 mg/㎖의 우리딘 및 1.2 mg/㎖의 FOA를 갖는 보겔 최소 배지의 한천 플레이트로 옮겼다. FOA의 존재하에 성장할 수 있는 능력은 pyr2 유전자의 Cas9-매개 불활성화로 인한 우리딘 영양요구성의 지표이다.
PCR 분석을 위해 FOA 내성 하이그로마이신 안정 및 불안정 형질전환체의 일부로부터 게놈 DNA를 추출하였다. 이 분석을 위해 사용된 프라이머는 표적 부위에 걸친 pyr2 유전자좌의 영역을 증폭시키기 위해 설계된 pyr2F(5'-gtataagagcaggaggagggag [서열 번호 36]) 및 pyr2R(5'- gaacgcctcaatcagtcagtcg [서열 번호 37])였고 약 0.8 kb의 길이를 가졌다.
FOA 내성이 있는 것으로 나타난 QM6a 형질전환체 중, 안정한 18개 및 불안정한 5개의 하이그로마이신 내성 형질전환체를 야생형 균주와 유사한 크기로 추정하여 pyr2 유전자좌의 영역을 증폭시키기에 충분한 연장 시간을 갖는 PCR 프로토콜을 사용하여 시험하였다. 안정한 형질전환체 중 어느 것도 이 짧은 연장 시간을 갖는 PCR 생성물을 생성하지 않는 반면, 불안정한 형질전환체 중 2개는 PCR 생성물을 생성하였다. 이들 두 PCR 생성물에 대한 DNA 서열 분석 결과, 하나가 단일 뉴클레오타이드 결실을 가지며, 다른 하나는 예상된 표적 부위에서 111 nt 결실을 가졌다.
FOA 내성이 있는 것으로 나타난 RL-P37 형질전환체들 중, 안정한 4개 및 불안정한 4개의 하이그로마이신 내성 형질전환체를 짧은 연장 시간을 갖는 PCR 프로토콜을 사용하여 시험하였다. 안정한 형질전환체 중 어느 것도 이 짧은 연장 시간을 갖는 PCR 생성물을 생성하지 않는 반면, 불안정한 형질전환체 중 둘 모두는 PCR 생성물을 생성하였다. 이들 두 PCR 생성물에 대한 DNA 서열 분석 결과, 하나가 단일 뉴클레오타이드 결실을 가지며, 다른 하나는 예상된 표적 부위에서 134 nt의 삽입을 가졌다. 이 삽입은 pTrex2gHyg 벡터의 두 작은 단편으로 구성된다.
큰 DNA 단편이 pyr2 유전자좌의 표적 부위에 삽입된 것으로 추정하고서, pyr2 유전자좌 영역의 증폭을 가능하게 하도록 설계된 앞서 기술된 PCR 프로토콜을 사용하여 상이한 6개의 안정한 하이그로마이신 내성 RL-P37 형질전환체를 분석하였다. 6개의 모든 형질전환체는 이 긴 시간 연장 프로토콜로 큰 생성물 (형질전환체에 따라 약 5 kb와 12 kb 초과 사이)을 제공했다. 이들 PCR 생성물 중 5개에 대한 DNA 서열 분석 결과, pTrex2gHyg 벡터 DNA 또는 이의 단편이 모든 경우에 통합된 것으로 나타났다.
종합하면, 이들 데이터는 Cas9에 의해 야기되는 이중 가닥 절단의 수리가 안정한 형질전환체 내에서 큰 벡터 단편의 통합을 주로 포함한다는 것을 보여준다. 이는 유전자 불활성화의 매우 효율적인 방법이 될 수 있다. 이는 또한 기능적 유전자를 가지며 표적 부위와 서열 상동성이 없는 DNA 단편 또는 벡터가 Cas9 절단 및 이중 가닥 절단 형성 후 표적 부위에서 부위 특이적 방식으로 통합될 수 있음을 나타낸다. 대조적으로, 작은 결실 또는 삽입(삽입 결실)은 불안정한 형질전환체에서 Cas9에 의한 유전자의 불활성화와 연관되어있다. 이는 벡터 통합이 바람직하지 않은 경우 유전자 비활성화에 대한 선택가능한 방법이다.
실시예
7: 텔로미어를 갖는 발현 벡터를 이용한
cas9
및
sgRNA의
발현
Cas9 및 가이드 RNA 발현 벡터 pTrex2gHyg MoCAS gPyr2 TS6의 버전이 트리코데르마 레에세이 텔로미어 서열(도 6에 도시)을 포함하도록 구성되었다. 하기 DNA 서열(서열 번호 38)을 벡터에 삽입하였다. 밑줄 쳐진 영역은 반복된 텔로미어 서열을 포함하며, 각각은 이 단편의 중앙을 향하여 판독된다. 중앙 부분은 텔로미어 리피트의 유지를 보장하기 위해 대장균에서 선택을 가능하게 하는 프로모터 및 종결인자를 갖는 박테리아 카나마이신 내성 유전자이다. 트리코데르마에서, 텔로미어를 갖는 벡터는 각 말단에서 텔로미어 서열로 선형화 될 것으로 예상되며 염색체 DNA에 가끔씩 통합이 일어날 수도 있지만 낮은 카피 수에서 자율적으로 유지되어야 한다.
이 벡터는 원형질체의 PEG-매개 형질전환에 의해 T. 레에세이 균주 RL-P37에 삽입된다. 형질전환체는 하이그로마이신 저항성을 위해 선택되었고 하이그로마이신이 함유된 신선한 한천 플레이트로 옮겨졌다. 대다수의 형질전환체는 불안정한 하이그로마이신 내성 표현형을 보였다. 개별적으로 형질전환된 콜로니를 2 mg/ml 우리딘 및 1.2 mg/ml 5-플루오로오로트산이 함유된 최소 배지로 옮겨서 성장할 수 있는 개체를 선택하여 Pyr-마이너스 표현형을 얻었다. 142개의 불안정한 형질전환체 중 8개(6%)가 Pyr-마이너스였다. pyr2 유전자좌의 PCR 및 이들 형질전환체 3종의 서열 분석 결과, 2개는 표적 부위에서 작은 결실(각각 1 bp 및 27 bp)을 나타내었고, 하나는 pTrex2gHyg MoCAS gPyr2 TS6의 박테리아 벡터 부분에서 유래된 68 bp의 삽입과 결합된 1 bp 결실을 가졌다. 다른 5개의 형질전환체는 큰 DNA 단편을 증폭하기 위해 설계된 PCR 조건을 사용함에도 불구하고 PCR 생성물을 제공하지 않았다[PCR 조건: 단계 1: 94℃에서 1 분; 단계 2: 94℃에서 25 초; 단계 3: 63℃에서 30초(온도가 사이클마다 0.2℃씩 감소); 단계 4: 70℃에서 8 분; 단계 2~4는 24회 더 반복. 단계 5: 4℃에서 유지. 폴리머라아제: PfuUltra II Fusion HS DNA 중합 효소 (Agilent Technologies)].
이러한 결과는 자율적으로 복제하는 벡터로부터의 Cas9 및 가이드 RNA의 발현이 특정 유전자좌(이 경우 pyr2)에 대한 Cas9 표적화를 가능케 함을 나타낸다. 수득된 유전자 불활성화는 벡터 DNA를 표적 부위에 삽입하지 않고 일어날 수 있다.
실시예
8:
상동성
통합에 의한 유전자 편집
다음 실험을 위해 트리코데르마 레에세이 균주 T4(1)7을 사용했다. 이는 증가된 셀룰라아제 생산성을 선별함으로써 RL-P37로부터 유래된 균주로, pyr2 유전자를 불활성화하여 균주를 우리딘 영양요구인자로 만드는 단일 점 돌연변이를 갖는다.
하기 서열(서열 번호 39)을 갖는 Gla1rep로 불리는 합성 DNA 단편을 설계하고 수요자 맞춤형으로 만들었다.
Gla1rep 서열은 ORF 내 gla1 유전자좌의 982 bp이다. 이 서열은 TS11 표적 부위(밑줄)에 걸쳐 있다. Gla1 코딩 서열에서 프레임 쉬프트를 생성하고 TS11에 인접한 PAM을 파괴하기 위해, TS11 표적 부위의 바로 상류의 야생형 Gla1 유전자의 “CCG” PAM 서열 내 단일 "C” 뉴클레오티드를 결실시켜, Cas9에 의한 절단을 방지한다. PAM의 나머지 두 뉴클레오티드는 대문자 볼드체로 표시된다.
프라이머 gla1rep F 및 gla1rep R(각각 5'-gtgtgtctaatgcctccaccac [서열 번호 32] 및 5'-gatcgtgctagcgctgctgttg [서열 번호 33])을 사용하는 형질전환에 사용하기 위해 Gla1rep 단편을 PCR로 증폭시켰다.
T. 레에세이 균주 T4(1) 7의 원형질은 pTrex2gHyg MoCas GTrGA TS11B (2 ㎍) + Gla1rep (8 ㎍)로 PEG-매개 방법에 의해 공동 형질전환되었다. 50 ug/ml 하이그로마이신 B, 2 mg/ml 우리딘 및 1.1 M 소르비톨을 함유한 보겔의 최소 배지를 사용하여 한천 플레이트에서 형질전환체를 선택하였다. 플라스미드 pTrex2gHyg MoCas gTrGA TS11B는 TS11 가이드 RNA의 발현 카세트가 나머지 플라스미드에 대해 반대 방향으로 있다는 것을 제외하면 pTrex2gHyg MoCas gTrGA TS11과 동일하다.
형질전환체를 우리딘과 하이그로마이신을 갖는 보겔의 최소 배지의 새로운 한천 플레이트에 집어 넣었고, 안정한 및 불안정한 하이그로마이신 내성 표현형을 구별하는 것이 가능했다. 글루코아밀라아제 양성 또는 음성 표현형에 대해 점수를 매기기 위해 형질전환체를 유일한 탄소원으로서 우리딘 및 1% 불용성 전분을 갖는 보겔 최소 배지의 한천 플레이트로 옮겼다. 안정한 하이그로마이신 저항성 형질전환체의 약 83%는 글루코아밀라아제 생산에 대해 음성이었지만, 불안정한 하이그로마이신 내성 형질전환체의 15%는 글루코아밀라아제 생산에 대해 음성이었다. 글루코아밀라아제-마이너스 표현형을 갖는 7개의 불안정한 형질전환체를 비선택성 한천 배지(보겔의 배지 + 우리딘)로 옮기고 1주 동안 성장시켰다. 그 후 보겔의 배지 + 우리딘 + 하이그로마이신의 플레이트에 집어넣을 때 이들은 모두 하이그로마이신에 민감하여 pTrex2gHyg MoCas gTrGA TS11B와 연관된 하이그로마이신 내성 유전자의 손실을 나타냈다.
pTrex2gHyg MoCas GTrGA TS11B 및 Gla1rep(형질전환체 #31, 107, 114, 118 및 120)로 얻은 5개의 불안정한 하이그로마이신-민감성 및 글루코아밀라아제-음성 형질전환체로부터 게놈 DNA를 단리하여 약 3.2 kb에 걸친 TS11 또는 glaK[서열 번호 31](하기 참조) 및 glaH 5’-tgccgtgggtcattggcatattc[서열 번호 29]를 증폭하도록 설계된 프라이머 glaA 및 glaD(각각 5'-ccgttagttgaagatccttgccg [서열 번호 26] 및 5'-gagagacgcaggatgactcaaag [서열 번호 28])를 사용하는 PCR(위에서 기재된 프로그램)에서 주형으로 사용하였다. gla1rep F 및 gla1rep R(각각 5'-gtgtgtctaatgcctccaccac [서열 번호 32] 및 5'-gatcgtgctagcgctgctgttg [서열 번호 33])을 프라이머로 사용하여 PCR 생성물을 서열 분석하여 표적 부위 TS11에서의 변이를 결정하였다. 형질전환체 중 하나는 TS11과 관련된 PAM에서 단일 bp 결실을 도입하고 gla1 유전자를 불활성화시킨 gla1 유전자좌에서 Gla1rep의 상동종 재조합과 일치하는 PCR 및 서열 분석 결과를 나타내었다. 형질전환체 중 2개는 TS11 표적 부위에서 작은 삽입 결실을 가지지만 다른 2개는 이 부위를 통한 상동성 통합보다는 Gla1rep 단편의 Cas9 절단 부위 내로의 삽입을 보였다.
pTrex2gHyg MoCas GTrGA TS11A(가이드 RNA 발현 카세트가 벡터 내에서 반대 방향으로 존재하는 것을 제외하고 pTrex2gHyg MoCas GTrGA TS11B와 동일함) + gla 1 유전자좌에서 상동성 재조합에 의한 통합을 하도록 설계된 선형 DNA 단편을 사용하여 원형질체를 동시에 형질전환시킨 위의 실험을 반복하였다. 그러나, 글루코아밀라아제 유전자 내의 표적 부위 TS11에서 상동종 재조합을 위한 공여체로서 982 bp Gla1rep DNA 단편을 사용하는 대신, Gla1repL이라 불리는 더 길고 약 2 kb의 단편을 사용하였다. Gla1repL의 중앙 부분은 Gla1rep와 동일한 서열이었지만 단편의 5’ 및 3’ 말단은 gla1 유전자좌의 상류 및 하류 부분을 더 포함하도록 연장되었다. 상기에서 사용된 균주 T4(1)7 대신 트리코데르마 레에세이 균주 RL-P37을 이 실험에 사용하였다. 대조군으로 Gla1repL과 pTrex2gHyg MoCas로 원형질체를 공동 형질전환시켜 활성 Cas9가 없는 gla1 유전자좌에서 Gla1repL이 통합되는 빈도를 결정하였다. 형질전환 및 표현형 선별에 이어 형질전환체를 다음 범주에 할당할 수 있었다.
pTrex2gHyg MoCas gTrGA TS11A 및 Gla1repL(안정한 형질전환체 #51, 52, 60, 61 및 67, 불안정한 형질전환체 338, 41, 65, 66 및 67)로 얻은 5개의 안정한 및 5개의 불안정한 하이그로마이신-민감성 및 글루코아밀라아제-음성 형질전환체로부터 게놈 DNA를 단리하여 프라이머 glaA 및 glaD를 사용한 PCR에서 주형으로 사용하였다(상기 참조). gla1 유전자의 표적 부위 TS11에 삽입이 없거나 큰 결실이 발생하지 않으면 PCR 생성물은 3.2 kb가 될 것으로 예상된다.
5개의 안정한 형질전환체 중 3개(#52, 60 및 61)는 약 3.2 kb의 PCR 생성물을 제공하였고, 다른 2개는 TS11에서 DNA 삽입을 나타내는 더 큰 생성물을 제공하였다. glarepF를 프라이머로서 사용하여 약 3.2kb의 3개의 PCR 생성물을 서열분석하였다. 2개의 형질전환체(#52 및 #61)의 경우 서열분석 결과는 gla1 유전자좌에서 상동성 재조합에 의한 Gla1repL의 통합과 일치하고 다른 하나는 쉽게 해석 할 수 없는 혼합 신호를 가졌다.
5개의 불안정한 형질전환체 중 하나(#66)만이 약 3.2 kb의 PCR 생성물을 제공하였고, 나머지 4개는 TS11에서 DNA 삽입을 나타내는 더 큰 생성물을 제공하였다. glarepF를 프라이머로 사용하여 약 3.2kb의 하나의 PCR 생성물을 서열분석하였고 결과는 gla1 유전자좌에서 상동성 재조합에 의한 Gla1repL의 통합과 일치하였다.
종합하면, 이들 결과는 선형 DNA 단편의 상동성 통합이 표적 부위에서 Cas9 절단에 의해 자극될 수 있음을 보여준다. 그러나, 비상동성 말단 결합(NHEJ)에 의한 DNA의 작은 삽입 결실 또는 큰 삽입은 또한 일반적인 발생이다. 더 큰 상동성 선형 DNA 단편의 사용은 다른 경우에 비해 표적 부위의 상동성 통합의 빈도를 향상시키는 데 도움이 된다. 하이그로마이신이 없는 배지에서 성장을 허용함으로써 pTrex2g MoCas-기질 벡터가 후속적으로 제거될 수 있는 불안정한 하이그로마이신 내성 형질전환체 내 표적 부위에서 상동상 통합을 얻는 것이 가능하다.
실시예
9: T.
레에세이의
NHEJ
-결핍 균주에서 유전자 편집
트리코데르마 레에세이(RL-P37에서 유래되고 결실된 셀로바이오하이드로라아제 1, 셀로바이오하이드로라아제 2, 엔도글루카나아제 1, 및 엔도글루카나아제 2 유전자(Δcbh1, Δcbh2, Δegl1, 및 Δegl2 균주; WO 92/06184 및 WO 05/001036 참조))의 "사중-결실 균주” 로부터 유래되고 천연 엔도글루카나아제-3 및 베타글루카나아제-1 유전자에서 결실을 갖는 균주(MAD6)를 Cas9 표적 부위에서 비상동성 말단 결합(NHEJ) DNA 삽입의 역할을 판단하기 위해 설계된 실험을 위해 사용하였다. 또한 DNA 재조합을 위한 주요 NHEJ 경로에 필수적인 인간 ku80에 대해 병렬상동(orthologous)인 천연 유전자에 대해 MAD6 균주는 결실되었다 (MAD6 균주가 제조되는 방법의 설명을 위한 US20130149742 A1 "Filamentous fungal host strains and DNA constructs, and methods of use thereof" 참조). 상기 균주를 위에서 기술된 pTrex2gHyg MoCAS gTrGA TS11B + 공여체 Gla1rep 단편으로 상기 균주를 공동 형질전환시켰다. gla1 유전자좌에서 상동성 재조합에 의한 이 단편의 통합은 PAM 서열로부터 하나의 bp를 결실시킴으로써 gla1 유전자를 불활성화시키고 TS11 표적 부위를 제거할 것이다. 형질전환체는 PEG-매개 방법에 의해 원형질체로부터 얻어졌다. 1.1M 소르비톨 및 100 ug/mL 하이그로마이신 B를 함유하는 보겔의 최소 배지에서 형질전환체에 대한 선택을 수행하였다. 하이그로마이신만을 함유한 최소 배지의 새로운 한천 플레이트로 옮긴 91개의 형질전환체 중에서 4개는 안정한 하이그로마이신 내성 표현형을 가졌다(비선택적 조건 하에서의 성장 기간 후 하이그로마이신을 갖는 배지상에서 재도포시 성장하는 이들의 능력으로 확인됨). 유일한 탄소원으로서 1% 불용성 전분을 갖는 보겔l의 최소 배지로 모든 형질전환체를 옮기고, 4개의 안정한 형질전환체를 포함하여 17개(18%)가 글루코아밀라아제에 음성으로 나타났다. PCR 및 DNA 서열 분석 결과, 공여체 Gla1rep이 gla1 좌에서 상동성 재조합에 의해 통합된 경우, 13개의 불안정한 형질전환체 중 12개와 안정한 형질전환체 중 하나가 TS11 PAM에서 단일 bp 결실을 갖는 것으로 나타났다. 다른 불안정한 형질전환체는 글루코아밀라아제 음성 표현형을 가짐에도 불구하고 야생형 gla1 서열을 가졌다. 다른 3개의 안정한 형질전환체는 gla1 유전자좌에 대해 예상되는 크기의 명확한 PCR 생성물을 제공하지 않았고, 이것들에서 벡터 또는 공여체 Gla1rep 삽입이 일어날 수 있다. 모든 불안정한 글루코아밀라아제 음성 형질전환체를 하이그로마이신이 없는 배지에서 성장시켰고 하이그로마이신을 함유하는 배지로 다시 옮겼다. 어느 것도 성장할 수 없었는데, 이는 이들이 pTrex2gHyg MoCAS gTrGA TS11B 벡터를 잃었음을 나타낸다.
이러한 결과는 NHEJ가 결핍된 균주에서 Cas9 표적 부위에 벡터 또는 공여체 DNA 단편 삽입이 최소화되었음을 명확하게 보여준다. 결과적으로, Cas9 및 가이드 RNA의 일시적인 발현과 함께 불안정한 형질전환체에서 공여체 DNA 단편의 상동종 재조합을 통해 매우 특이적인 유전자 편집(단일 bp의 결실)의 높은 빈도가 가능하다.
섹션 B:
Cas
닉카아제
/가이드 RNA의 직접 도입
실시예
10: 대장균에서
CRISPR
SpyCas9
-
D10A
닉카아제의
이종 발현
Generay(중국, 상해)는 대장균 코돈에 최적화된 스트렙토코쿠스 파이오제네스 Cas9-D10A(SpyCas9-D10A) 닉카아제 유전자를 합성하여 NcoI 및 HindIII 부위에서 발현 벡터 pET30a에 삽입하여 플라스미드 pET30a-SpyCas9-D10A 닉카아제를 수득하였다(도 7). 도 7의 플라스미드 지도에 표시된 바와 같이, 발현 카세트의 전체 코딩 서열은 5 '에서 3'방향으로, 모든 작동가능한 연결에서 N- 말단 His6 태그/트롬빈/S ㆍ Tag ™/엔테로키나아제 영역을 인코딩하는 서열(서열 번호 68, 메티오틴을 위한 개시 코돈을 포함), SV40 핵 위치 신호를 인코딩하는 서열(서열 번호 69), SpyCas9-D10A 닉카아제를 인코딩하는 서열(서열 번호 65), 및 BLR2 핵 위치 신호를 인코딩하는 서열(서열 번호 70)을 포함한다. 이 전체 코딩 서열을 서열 번호 54에 나타난다. 서열 번호 68에 의해 인코딩되는 N-말단 His6 태그/트롬빈/S ㆍ 태그 ™/엔테로키나아제 영역의 아미노산 서열은 서열 번호 67(위치 1의 메티오닌을 포함함)에 나타나고, 서열 번호 69에 의해 인코딩되는 SV40 핵 위치 신호의 아미노산 서열은 서열 번호 46에 나타나고, 서열 번호 65에 의해 인코딩되는 SpyCas9-D10A 닉카아제의 아미노산 서열은 서열 번호 66에 나타나고, 서열 번호 70에 의해 인코딩되는 BLR2 핵 위치 신호의 아미노산 서열은 서열 번호 47에 나타난다. 서열 번호 54에 의해 인코딩되는 아미노산 서열은 서열 번호 55에 나타난다.
pET30a-SpyCas9-D10A 닉카아제 플라스미드를 Rosetta2 (De3)plysS 대장균 균주(독일, 다름슈타트의 Novagen®, EMD Biosciences, Inc., Merck KGaA)로 형질전환시키고, 형질전환 생성물을 34 ppm 클로로암페니콜(Chloramphenicol) 및 50 ppm 카나마이신(Kanamycin)으로 보충된 Luria 한천 플레이트 위에 펼쳤다. 콜로니를 집어 넣고 250 ml 쉐이크 플라스크에서 25 ml의 Invitrogen MagicMediaTM(Thermo Fisher Scientific Inc.)으로 24시간 동안 30℃에서 300 rpm으로 발효시켰다.
SpyCas9-D10A 서열이 유래된 스트렙토코쿠스 파이오제네스 유래의 야생형 Cas9 단백질의 아미노산 서열을 서열 번호 45로 나타낸다.
실시예
11:
SpyCas9
-
D10A의
정제
SpyCas9 (D10A)의 정제를 위해 친화성, 소수성 상호작용 및 크기 배제 크로마토그래피 단계들의 조합을 적용하였다. 2리터의 미정제 원액을 채취하여 원심분리 하였다. 세포를 펠릿화하고 400 ml 용해 완충액(20 mM HEPES, pH7.5, 500 mM NaCl, 0.1% Triton X-100, 1 mM DTT 및 1 mM TCEP, Roche에서 구입한 프로테아제 억제제 칵테일)에서 재현탁하였고, 초음파 분쇄기(35% 파워, 20분, 2초 온/3초 오프)(중국, 제징의 SCIENT2-II D, Ningbo Scientz Biotechnology Co., LTD.)로 세포를 용해시켰다. 용해물을 20,000 g에서 40분 동안 원심분리하여 제거하였다.
정화된 용해물을 롤링 인큐베이터(중국, 하이멘의 Kylin-Bell Lab Instruments Co., Ltd.)에서 4℃에서 30 rpm으로 Ni-NTA 수지(GE Healthcare)로 밤새 배양하였다. 원심 분리 후, 수지를 XK26/20 컬럼(GE Healthcare)에 옮기고 AKTA Explorer 시스템(GE Healthcare)에 연결하였다. 평형 완충액(20 mM HEPES, pH 7.5, 300 mM NaCl, 0.1 % Triton X-100) 및 세척 완충액(평형 완충액 내 25 mM 이미다졸)으로 광범위하게 세척한 후, 표적 단백질을 평형 완충액 내 50, 250 및 500 mM 이미다졸로 용출시켰다. 원하는 단백질은 50 및 250 mM 이미다졸 용출액에서 상대적으로 높은 순도로 발견되었고, 이들을 모으고 별도로 추가처리 하였다.
친화성 단계로부터 수집된 활성 분획에 0.6 M까지 황산암모늄을 첨가하고 20 ml 페닐-세파로스 HP 칼럼(GE Healthcare)에 로딩하였다. 칼럼을 HEPES 완충액(pH 7.5)에서 0.6 M 내지 0.0 M 황산암모늄의 구배로 용출시켰다. 각 분획의 순도는 SDS-PAGE 겔에 의해 평가되었으며, 관심 단백질이 주로 유류(flow-through) 분획에 존재함이 밝혀졌다.
최종적으로, 20 mM HEPES pH 7.5, 150 mM KCl 및 10% 글리세롤 중의 Superdex 200 16/60 칼럼(GE Healthcare) 상의 크기 배제 크로마토그래피로 단백질을 추가로 정제하였다. 단백질-함유 순수 분획들을 모으고 Amicon 30 KDa 멤브레인 필터(Millipore)를 사용하여 농축시켰다. 정제 단백질(각각 50 mM 이미다졸 및 250 mM 이미다졸 용출액)의 두 배치를 사용하기 전까지 150 mM KCl 및 40% 글리세롤을 함유한 20 mM HEPES 완충액에 pH7.5에서 -20℃에서 보관하였다.
실시예
12:
닉카아제의
시험관내
분석
시험관내
닉카아제
절단 분석을 위한 기질 DNA 단편의 제조
RL-P37에서 유래된 트리코데르마 레에세이 균주에서 추출되었으며 결실된 셀로바이오하이드로라아제 1, 셀로바이오하이드라아제 2, 엔도글루카나아제 1 및 엔도글루카나아제 2 유전자(Δcbh1, Δcbh2, Δegl1 및 Δegl2 균주; “사중-결실 균주”로도 불림; WO 92/06184 및 WO 05/001036 참조)를 갖는 게놈 DNA를 Zymo Research사(Irvine, CA)의 ZF Fungal/Bacterial DNA miniprep 키트를 사용하여 추출하였다. 1 ng의 추출 된 게놈 DNA를 주형으로 하여, 트리코데르마 레에세이 글루코아밀라아제(TrGA) 유전자(유전자 ID: 18483895) 및 이의 부분 5'-UTR (서열 번호 56)을 포함하는 DNA 단편을 KOD-Plus PCR 키트(일본, Toyobo사) 및 순방향 및 역방향 프라이머 각각 0.4 μM을 사용하여 PCR로 증폭시켰다: 5'-gactgtctccaccatgtaatttttc-3'(서열 번호 57) 및 5'-ggcagactacaagtctactagtactac-3’(서열 번호 58). PCR 생성물을 Zymo Research사의 DNA Clean & ConcentratorTM-5 키트를 사용하여 정제 및 농축하고 DNA 농도를 NanoDrop™ 분광광도계(Thermo Fisher Scientific Inc.)로 측정하였다.
서열 번호 56(하기)은 기질 DNA 단편의 뉴클레오티드 서열을 나타낸다. UTR 서열은 소문자로 나타나고 TrGA 유전자는 대문자로 나타난다. 두 개의 선택된 VT 도메인인 TrGA_sgF1과 TrGA_sgR1은 각각 굵은 글씨와 밑줄로 나타난다(이들 순서가 겹쳐있음에 주목).
시험관내
전사
TrGA 유전자의 두 가지 VT 도메인인 TrGA_sgF1과 TrGA_sgR1 및 이들 특유의 PAM을 하류 활성 및 형질전환 실험에 대해 확인하였다. TrGA_sgF1(서열 번호 59) 또는 TrGA_sgR1(서열 번호 60)을 갖는 T7 프로모터 및 단일 가이드 RNA 서열을 포함하는 올리고 뉴클레오타이드를 합성하였고 Generay에 의해 pMD18T 벡터에 삽입하여 pMD18T(T7-Spy- TrGA_sgF1)(도 8a) 또는 pMD18T(T7-Spy-TrGA_sgR1)(도 8b)를 각각 수득하였다. 시험관내 전사를 위한 DNA 단편을 순방향 및 역방향 프라이머 각각 0.4 μM로 PCR에 의해 pMD18T(T7-Spy-TrGA_sgF1) 또는 pMD18T(T7-Spy-TrGA_sgR1)로부터 증폭시켰다: 5'-tgatgacggtgaaaacctc-3'(서열 번호 71) 및 5'-aaaagcaccgactcgg-3'(서열 번호 72). PCR 생성물을 Zymo Research사의 DNA Clean & ConcentratorTM-5 키트를 사용하여 정제 및 농축하고 이의 DNA 농도를 NanoDrop™ 분광광도계로 측정하였다.
상기 특정 PCR 생성물을 주형으로 하여, VT 도메인인 TrGA_sgF1 또는 TrGA_sgR1에 대한 RNA를 Invitrogen, Thermo Fisher Scientific Inc.의 MEGAshortscriptTM T7 전사 키트를 사용하여 제조사의 지침에 따라 시험관내 전사에 의해 생성시켰다. 전사된 RNA를 Invitrogen, Thermo Fisher Scientific Inc.의 MEGAclear™ 전사 클린-업 키트를 사용하여 정제하였다. RNA 농도를 NanoDropTM에 의해 측정하였다.
합성된 단일 가이드 RNA의 기능을 확인하기 위해 SpyCas9 시험관내 분석을 수행했다. 분석을 시작하기 위해, 1 μg의 정제된 SpyCas9, 200 ng의 기질 DNA 단편 및 200 ng의 단일 가이드 RNA(또는 대조물로서의 물)를 50 mM HEPES pH 7.3, 150 mM KCl, 0.5 mM DTT 및 10 mM MgCl2를 함유하는 15 μl의 반응 완충액에서 함께 혼합하였다. 37℃에서 20 분간 분석을 수행한 후, 프로테이나제 K(Sigma, Cat No. P6556) 2 μg을 첨가하였다. 40℃에서 20분 동안 반응을 계속하였고 80℃에서 20분 동안 추가로 배양하여 반응을 종료하였다. 0.8% 아가로스 겔을 사용하여 140 볼트에서 30분 동안 반응 결과를 분석하였고, 분석한 결과를 도 9에 나타내었다. 도 9에서, 레인 1은 DNA 사다리(분자량은 좌측에 도시)이고, 레인 2는 대조 SpyCas9 반응(가이드 RNA 없음)이고, 레인 3 및 4는 TrGa_sgR1의 존재 하에 SpyCas9를 나타내고, 레인 5 및 6은 TrGA_sgF1의 존재 하에서 SpyCas9를 나타낸다. 손상되지 않은 DNA 기질(서열 번호 56)의 크기는 4.9 Kb(레인 2)인 반면, sgRNA 존재 하에서 절단된 생성물의 크기는 2.8 Kb 및 2.1 Kb이다. 도 9에 도시 된 바와 같이, 특정 단일 가이드 RNA의 존재 하에서, SpyCas9는 기질 DNA 단편을 원하는 크기로 성공적으로 절단하여 합성된 RNA의 올바른 기능을 확인할 수 있다.
서열 번호 59(하기)는 T7 프로모터, CER 도메인, 및 VT 도메인 TrGA_sgF1의 전사를 위한 올리고뉴클레오티드 서열을 나타낸다. VT 도메인은 대문자로 표시되어 있지만, T7 프로모터와 CER 도메인 영역은 각각 굵은 글씨와 소문자로 표시되어 있다.
서열 번호 60(하기)은 T7 프로모터, CER 도메인, 및 VT 도메인 TrGA_sgR1의 전사를 위한 올리고뉴클레오티드 서열을 나타낸다. VT 도메인은 대문자로 표시되어 있지만, T7 프로모터와 CER 도메인 영역은 각각 굵은 글씨와 소문자로 표시되어 있다.
정제된
SpyCas9(D10A)를
이용한
시험관내
닉카아제
절단 분석
시험관 내 닉카아제 절단 분석은 2단계 반응이다. 첫 번째 단계를 위해, 1 μg의 정제된 SpyCas9(D10A), 200 ng의 기질 DNA 단편 및 200 ng의 단일 가이드 RNA(또는 대조물로서의 물)를 50 mM HEPES pH 7.3, 150 mM KCl, 0.5 mM DTT 및 10 mM MgCl2를 함유하는 15 μl의 반응 완충액에서 함께 혼합하였다. 반응은 SpyCas9 핵산 분해 효소 분석법에 기재된 대로 수행하였다. 제1 단계 반응의 종료 후, 1 μg의 SpyCas9(D10A)와 200 ng의 특정 단일 가이드 RNA를 첨가하였다. 제1 단계 반응을 반복한 후, 그 후 반응 결과를 140 볼트에서 30분 동안 작동하는 0.8% 아가로스 겔을 사용하여 분석하였고 결과를 도 10에 나타내었다. 도 10에서, 레인 1은 DNA 사다리(분자량은 좌측에 도시)이고, 레인 2는 SpyCas9(D10A)와 기질 DNA 및 TrGA_sgF1만의 반응을 나타내고, 레인 3은 SpyCas9(D10A)와 기질 DNA 및 TrGA_sgR1만의 반응을 나타내고, 레인 4는 SpyCas9 (D10A)와 기질 DNA 및 두 sgRNA의 반응을 나타낸다. 손상되지 않은 DNA 기질(서열 번호 56)의 크기는 4.9 Kb인 반면, 두 sgRNA를 갖는 절단 생성물의 크기는 2.8 Kb 및 2.1 Kb가 될 것이다. 도 10에 도시된 바와 같이, 기질 DNA 단편은 두 RNA(레인 4)의 존재 하에서 절단만 되는데, 이는 SpyCas9(D10A)가 활성 닉카아제라는 것을 제시한다.
실시예
13:
생체내
닉카아제
흡수 실험
원형질체 준비
원형질체 준비를 위해, 엔도글루카나아제-3, 엔도글루카나아제-4, 엔도글루카나아제-5, 엔도글루카나아제-6, 만나아제-1, 및 알파-아밀라아제 유전자의 추가 결실을 갖지만, 정상 NHEJ 메카니즘(30℃에서 5일 동안 PDA 플레이트에서 성장한)을 이용하여, T. 레에세이(위에서 기재된)의 사중-결실 균주의 5×108개 포자를 4개의 배플을 갖는 250 ml 쉐이크 플라스크에서 50 ml 발아 배지(미국 특허 제8,679,815호에 기술된 방법)에 접종하였고 27℃에서 170 rpm으로 17시간 동안 배양하였다. 액체 부피를 50 ml의 원뿔형 튜브로 옮기고 3000 rpm으로 10분 동안 회전시킴으로써 균사체를 회수하였다. 상층액을 따라 내고, 균체 펠릿을 1.2 M MgSO4 -10 mM Na- 인산염 완충액을 사용하여 2회 세척하고, 15 ml 용해 효소 완충액(1.2 M MgSO4-10 mM Na-인산염 완충액(pH 5.8), 50 mg/ml을 사용하는 트리코데르마 하르지아눔(Sigma 카탈로그 #L1412)으로부터 용해 효소를 용해)에 재현탁시켰다. 세포 현탁액을 4개의 배플을 갖는 250 ml 쉐이크 플라스크로 옮기고 상온에서 200 rpm으로 적어도 2 시간 동안 흔들었다. 유리 깔때기에서 Greiner 튜브로 접힌 Miracloth(Calbiochem Art. No. 475855)를 통한 여과에 의해 원형질체를 수확하였다. 0.6 M 소르비톨-0.1 M Tris-HCl 완충액을 여과된 원형질체 위에 조심스럽게 첨가하였다. 원형질체를 4000 rpm에서 15분 동안 원심분리하여 수집하였다. 원형질체를 함유하는 중층을 새로운 튜브로 옮기고 적어도 같은 부피의 1.2 M 소르비톨-10 mM 트리스-염산 완충액을 첨가하였다. 원형질체를 4000 rpm에서 5분간 원심분리하여 수집하고, 1.2 M 소르비톨, 10 mM 트리스-염산 완충액을 사용하여 2회 세척하였다. 적어도 1 ml 1.2 M 소르비톨-10 mM Tris-HCl pH 7.5-10 mM CaCl2 완충액에 펠릿을 재현탁시키고, 원형질체의 수를 현미경으로 계수하였다. 1.2 M 소르비톨-10 mM Tris-HCl-10 mM CaCl2 4부 및 25% PEG6000-50 mM CaCl2-10 mM Tris-HCl 1부를 사용하여 원형질체 현탁액을 장래의 형질전환을 위해 ml당 5x108까지 희석시켰다.
결실 카세트의 준비
TrGA 결실 카세트를 구성하여 도 11에 개략적으로 도시하였다. 이는 pyr2 프로모터, pyr2 CDS 및 pyr2 종결인자를 포함하는 pyr2 발현 카세트를 포함하고, 이는 5’ 및 3’ TrGA-상동성 영역에 의해 플랭킹된 후속 루프 아웃을 위한 500 bp 반복 서열이다.
TrGA 녹아웃 형질전환체를 포도당이 없고 1% 불용성 전분(보겔의 전분 배지)이 있는 보겔 한천 접시에서 선별할 수 있다. 손상되지 않은 TrGA 유전자를 갖는 균주와 비교하여 TrGA 녹아웃 형질전환체는 이 배지에서 불충분하게 성장한다. TrGA 녹아웃 카세트의 뉴클레오티드 서열은 길이가 4248 염기 쌍이고: 1 내지 1000 염기는 TrGA 5’ 상동성 영역에 상응하고; 1001 내지 2730 염기는 pyr2 발현 카세트에 상응하고; 2739 내지 3248 염기는 500 bp 리피트에 상응하고; 3249 내지 4248 염기는 TrGA 3’ 상동성 영역에 상응한다. TrGA 녹아웃 카세트의 뉴클레오티드 서열은 서열 번호 61(아래에 표시)로 제공된다.
agatgccggccttgaagcagtaggtcgagaggctccacttcatggtgccgttgccgatcatggtgttgatgatgcggtcgtacgtctcgatggcgccgtagtagtcgccgtcgagggcggcgaggtcggcgtactgcgtccagagctt
형질전환
Spycas9 닉카아제 프리믹스와 결실 카세트를 사용하여 T. 레에세이에서 Trga 유전자를 녹아웃하기 위해, 저장 완충액(20 mM HEPES pH 7.5, 150 mM KCl, 40% 글리세롤) 내 Spycas9 닉카아제 단백질 25 μg을 3 μl NEB 완충액 3(뉴 잉글랜드 바이오랩스)의 뉴클레아제가 없는 물에 용해된 20 μg sgRNA(TrGA_sgR1)와 부드럽게 혼합하여 30 μl 프리믹스를 얻었고, 상온에서 30분간 배양했다. 20 μg(20 μl)의 결실 카세트와 함께 200 μl의 원형질체(1 × 108)에 30 μl의 프리믹스를 첨가하고 30분간 얼음 위에 유지시켰다. 얼음 위에서 30분 동안 배양한 후, 최소 보겔 한천 플레이트의 최상층으로 사용되도록, 냉각된 용융 소르비톨/보겔 한천(2% 글루코오스를 함유한 최소 보겔 한천에서 1.1 M 소르비톨)에 원형질체를 첨가했다(Davis 등 (1970) Methods in Enzymology 17A, pgs. 79-143 및 Davis, Rowland, NEUROSPORA, CONTRIBUTIONS OF A MODEL ORGANISM, Oxford University Press, (2000)). 플레이트를 30℃에서 1주간 배양하였다. 상세한 단계는 미국 특허 제8,679,815호(본원에 참고로 통합됨)에 기재되어 있다.
90개의 형질전환체를 선택하여 3개의 대조군과 함께 웰당 1 ug의 신선한 보겔 한천을 갖는 4개의 24- 웰 플레이트에 접종하였다: 엔도글루카나아제-3, 엔도글루카나아제-4, 엔도글루카나아제-5, 엔도글루카나아제-6, 만나아제-1의 추가 결실을 갖는 사중-결실 균주(위에 기재된 T. 레에세이 균주; 도 12의 "셀루(cellu)"), 알파-아밀라아제 유전자의 추가 결실(AA 결실, 도 12에서 ΔAA)을 갖는 "셀루"균주, 및 추가의 글루코아밀라아제 유전자 결실(GA 결실; 도 12에서 ΔGA)을 갖는 “셀루” 균주. 일주일 후, 형질전환체를 웰 당 1 ml의 신선한 보겔-전분 한천을 갖는 또 다른 4개의 24- 웰 플레이트로 옮겼다. 일주일 후, 이들 90개 균주의 형태를 관찰하였다. 47개의 형질전환체는 “셀루” 균주 및 AA 결실(ΔAA)을 갖는 것들과 비교하여 지연된 성장을 보였지만 보겔의 전분 플레이트에서 GA 결실(ΔGA)을 갖는 "셀루” 균주의 형질전환체와 유사하게 보였다(도 12). 보겔의 전분 한천에서, 탄소원으로 포도당을 함유한 정규 보겔 한천에 비해 유일한 탄소원은 전분이다. 글루코아밀라아제는 전분의 비환원 말단으로부터 a-1,4 글리코시드 결합을 연속적으로 가수분해하여 곰팡이 균주에 의해 이용될 수 있는 글루코오스를 생산한다. 글루코아밀라아제가 결실된 경우, 글루코오스의 이용가능성이 제한되어 지체된 성장이 관찰될 수 있다.
균주 검증
보겔의 전분 한천에서 90개의 균주 중 47개의 균주가 성장 지연을 나타내었다. 이들중 9개(도 12의 #1 내지 #9 참조)를 무작위로 선택하여 Fw1(5'-cactactacatccatacgcagcaaacatgg-3'(서열 번호 62)) 및 R3(5'-ggtcaagaagcacatgccagagttcg -3'(서열 번호 63))을 프라이머로 사용하는 PCR 선별을 수행하였다. 도 13a는 프라이머 어닐링 부위뿐만 아니라 야생형 및 TrGA 결실 균주들의 게놈 유전자좌를 나타낸다(PCR 반응에서 사용시 화살표는 중합의 방향을 나타낸다).
프라이머 쌍 FW1 및 R3을 갖는 PCR로 9개의 균주를 예비 선별하였다. PCR 생성물을 증폭하기 위한 PCR 조건은 다음과 같다: 단계 1: 95°C에서 5분. 단계 2: 95°C에서 30초. 단계 3: 60°C에서 30초. 단계 4: 68°C에서 3분. 단계 2, 3, 및 4를 추가 29 사이클 동안 반복. 단계 5: 68°C에서 10분. 이상적인 조건에서, 2개의 PCR 단편(1.9-kb 및 5.2-kb)이 PCR 예비 선별에서 예상될 것이다. 이 결과에서 5.2-kb 생성물을 명확하게 관찰할 수 없었다. 그러나 셀루라이터(C1) 포자의 5.1-kb 생성물 및 셀루라이터 + TrGA 결실 카세트(C2) 포자의 PCR 생성물과 비교시, 1.9-kb 단편은 TrGA 결손 카세트가 상동종 재조합에 의한 TrGA 유전자좌(도 13b, 상부 겔)에서 통합되었음을 확실히 확인할 수 있었다.
프라이머 쌍 FW1/R1, F4/R3 및 KOF1/KOR2를 사용하여 추가적인 PCR 확인을 각각 수행하기 위해 9개의 균주 중 2개(#1 및 #3)를 선택했다. C2(셀루라이터 포자 + TrGA 결실 카세트)의 결과와 비교하여 #1 및 #3의 포자로부터 2.0-kb 및 2.2-kb 단편이 수득되었을 때(도 13b, 좌측 하단 겔). 프라이머 쌍 FW1/R1 및 F4/R3을 사용한 PCR로 상동성 재조합에 의해 TrGA 유전자좌에 통합된 TrGA 결실 카세트를 추가로 확인하였다. 2.1 kb의 생성물이 셀루라이터(C1)로부터 증폭되는 동안 PCR 생성물이 #1 및 #3의 포자로부터 수득되지 않았을 때, 또 다른 한 쌍의 프라이머인 KOF1 및 KOR2를 사용한 PCR도 결과를 확인하였다(도 13b, 우측 하단 겔, 분자량 마커를 포함하여 처음 4 레인).
프라이머 TrGAF2(5'gactgtctccaccatgtaatttttc3’(서열 번호 64)) 및 R3(서열 번호 63)을 사용하여 TrGA 유전자좌의 전체 영역에 대한 #1 및 #3 포자로부터 PCR 생성물을 수득하였고, 이들의 DNA 서열이 결정되었다. 서열분석 결과는 TrGA 유전자가 pyr2 발현 카세트에 의해 대체되었음을 보여주었다(도 13b, 우측 하단 겔, 마지막 2레인의 PCR 결과 참조; 서열분석 결과에 대한 데이터는 나타내지 않음).
위의 결과는 SpyCas9 닉카아제와 sgRNA가 사상균류에서 상동종 재조합을 촉진하여 사상균류 T. 레에세이에 상동성 재조합에 기초한 유전자 결실을 허용할 수 있다는 것을 나타낸다. 구체적으로, 90개의 형질전환체 중 47개(약 52%)는 보겔의 전분 플레이트에서 성장 지연된 표현형을 보였는데, 이는 TrGA 유전자가 파괴되었음을 나타낸다. 의도한 상동성 재조합 사건은, 결손 카세트가 상동성 재조합에 의해 숙주 세포에서 TrGA 유전자좌로 성공적으로 혼입되고 결과적으로 TrGA 유전자가 pyr2 발현 카세트로 대체되었다는 것을 확인하는 PCR 생성물 서열분석에 의해 입증되었다. 우리의 데이터는 기능적 SpyCas9 닉카아제 복합체를 공여체 DNA (즉, 관심 게놈 유전자좌에서 상동성 재조합될 DNA) 이외에 표적 진핵 세포에 직접 도입할 때, 균류 내 상동성 재조합률이 현저히 증가될 수 있다는 것을 나타낸다.
섹션 C: 단일 발현 벡터 위에
Cas
/가이드 RNA 및 공여체 DNA의 도입
실시예
14: 유전자 편집을 위해 유도된
상동성
공여체 단편을 포함하는
cas9
발현 플라스미드 사용에 의한
gla1
유전자에서 단일 염기의 결실
이 실시예에서, 단일 염기 결실을 함유하는 상동성 gla1 공여체 단편을 플라스미드 pTrex2gHyg MoCas GTrGA TS11B(실시예 8에 기재)에 혼입시킴으로써 유전자 파괴를 유도하였다. 약 1 kb의 합성 DNA 단편 Gla1rep(서열 번호 39, 도 14에서 "Tr-gla 1 kb 상동성 단편"으로 표지됨)을 플라스미드 pTrex2gHygMoCasgTrGATS11B의 유일한 EcoRV 제한 부위에 삽입함으로써 플라스미드 pTrexMoCasGATS11-HDR(도 14)를 생성하였다. EcoRV 부위는 하이그로마이신 내성 마커 (hph)의 pSL1180 서열과 N. 크라사(crassa) cpc1 프로모터 사이의 폴리링커 서열에 존재한다. Gla1rep를 증폭시키고 말단 상동성 테일을 첨가하여 pTrex2gHygMoCasgTrGATS11B에서 EcoRV 부위의 양측을 중첩하여 EcoRV-절단 pTrex2gHygMoCasgTrGATS11B 플라스미드로 단편의 깁슨 어셈블리를 용이하게 하기 위해(매사추세츠, 베버리, 뉴잉글랜드 바이오랩, NEB Gibson Assembly Master Mix를 사용하여), 프라이머 1556F(5'-ATGCGCAAATTTAAAGCGCTGATgtgtgtctaatgcctccaccac[서열 번호 73]) 및 1557R(5'-ATATGGATCTGCGCGCGATCGATgatcgtgctagcgctgctgttg[서열 번호: 74])를 사용하였다.
pTrexMoCasGATS11-HDR 플라스미드를 사용하여 원형질체 및 PEG 매개된 DNA 흡수를 이용하여 정상적인 NHEJ 메카니즘을 갖는 균주인 T. 레에세이 균주 RL-P37을 형질전환시켰다. 이 균주는 이 실험을 위해 부수적이지만 우리딘(2 g/L)이 모든 성장 배지에 포함되는 것을 필요로 하는 pyr2에 대해 영양요구성이었다.
보겔의 최소 배지 + 1.2M 소르비톨 + 75 ppm 하이그로마이신의 선택성 배지 한천 플레이트에서 형질전환체를 초기에 성장시켰다. 성장 후, 안정형 및 불안정한 표현형 모두의 80 가지 형질전환체를 보겔의 전분 배지의 제2 한천 플레이트로 옮겼다. 제2 플레이트에서 성장 후, gla1 유전자가 파괴되었는지를 결정하기 위해 전분 제거 구역의 존재 또는 부재를 리포터로 사용하였다. 형질전환체를 제2 플레이트에서 보겔의 최소 배지의 비선택적 제3 플레이트로 옮겼다. 성장 며칠 후, 형질전환체를 보겔의 최소 배지의 제3 플레이트에서 보겔의 최소 배지 + 75 ppm의 하이그로마이신 B의 제4 플레이트로 옮겼다. 제2 및 제3 플레이트의 비선택성 배지상에서 성장한 후 형질전환체가 하이그로마이신-함유 배지 상에서 성장할 수 있는 능력을 유지하는지 여부를 결정하기 위해, 이러한 제4 플레이트 상에서 성장에 대해 형질전환체를 평가할 수 있었다.
보겔의 전분 배지의 제거 구역에서 생산된 80개의 형질전환체 중 단지 11개만이 gla1 파괴를 나타내며, 보겔의 최소 배지 + 75 ppm 하이그로마이신의 제4 하이그로마이신 선택성 플레이트에서는 성장하지 않는다는 것이 밝혀졌다. 이 형질전환체들은 아마도 gla1 유전자의 변이를 가지고 있었고 플라스미드에서 이들의 게놈으로 하이그로마이신 마커를 혼입하지 못했다.
제3 보겔의 최소 배지 플레이트의 균사체에서 이들 11개의 gla1- 형질전환체로부터 게놈 DNA를 단리하였다. 프라이머 glaK(서열 번호 31) 및 glaH(서열 번호 29)를 사용하여 게놈 DNA로부터 gla1 유전자를 PCR 증폭하여 TS11 유전자좌의 상태를 결정하였다. 프라이머 1538F 5'-CCACCACAGGAACCAAACC(서열 번호 75), 1539R 5'-CTGCGACGGAGGGAATGACG(서열 번호 76), 1540F 5'-GGGCAGGACTGGCAAGGATGT(서열 번호 77) 및 1541R 5’-GCCGTCACGCCAGGAACAAG(서열 번호 78)를 사용하여 PCR 생성물을 단리 및 서열분석하였다.
서열 분석 결과, 이들 11개의 gla1- 형질전환체 중 8개가 Gla1rep 서열의 단일 염기 결실을 포함하는 것으로 밝혀졌다. 2개의 gla1-형질전환체는 9 및 100 염기의 TS11 유전자좌에서 gla1 결실을 포함하였다. 1개의 gla1-형질전환체는 TS11에서 470 염기의 삽입을 포함하였다.
공여체 Gla1rep 단편을 플라스미드 pTrex2gHyg MoCas TrGA TS11에 혼입하여 유전자 편집을 유도한 단일 플라스미드를 생성하였다. 하이그로마이신-불안정 형질전환체의 높은 빈도는 공여체 Gla1rep 단편 내에서 조작 된 gla1 유전자에서 단일 염기 결실을 포함하였다. 불안정한 형질전환체가 제1 선택적 한천 배지인 보겔의 최소 배지 + 75 ppm 하이그로마이신 B에서 생성되었으며, 이후 변형된 cas9 플라스미드를 잃어 버렸다. 이러한 균주는 이들이 유전자 편집 플라스미드를 게놈에 NHEJ 혼입시키는 불편함을 갖고 있지 않기 때문에, 보다 유리하다.
데이터가 여기에 도시되지 않은 또 다른 실험에서, pTrex2gHyg MoCas gTrGA TS11B에 삽입된 결실 대신에 치환 뉴클레오티드를 포함하는 gla 1 상동성 DNA 단편을 사용하여, 본원에 기술된 것과 동일한 상동성 통합 방법을 사용하여 gla1의 치환 돌연변이를 T. 레에세이 균주 P37에 성공적으로 도입하였다.
이러한 결과는 cas9 및 표적 부위 가이드 RNA와 함께 상동성 단편을 혼입하는 단일 cas9 편집 플라스미드를 생성하는 유용성을 나타낸다. 표적 유전자좌에서 특정 cas9 표적 절단 및 유전자 편집 상동성 재조합 모두를 유도하는 하나의 플라스미드 벡터가 생성된다. 불안정한 하이그로마이신 저항성 형질전환체를 선별함으로써, 목표 유전자좌로 플라스미드 DNA의 NHEJ 삽입의 발생이 최소화된다. 또한, 본 실시예에서 불안정한 하이그로마이신 내성 형질전환체를 선별함으로써 상동성 재조합에 의해 상동성 공여체 단편을 표적 유전자좌로 혼입시킨 형질전환체를 발견할 수 있다. 그러나, 이러한 형질전환체는 표적화된 재조합을 유도한 플라스미드를 편리하게 잃어 버렸다.
전술한 조성물 및 방법이 이해의 명료성을 위해 예시 및 실시예로써 일부 상세히 기술되었지만, 본원의 교시에 비추어 첨부된 청구범위의 사상 또는 범위를 벗어나지 않으면서 특정 변경 및 변형이 이루어질 수 있다는 것이 당업자에게는 쉽게 명백하다.
따라서, 선행 기술은 단지 본 발명의 조성물 및 방법의 원리를 설명한다. 당업자는 본원에 명시적으로 기재되거나 도시되지는 않았지만 본 조성물 및 방법의 원리를 구현하고 이의 사상 및 범위 내에 포함되는 다양한 장치를 고안할 수 있음을 이해할 것이다. 또한, 본원에 인용된 모든 실시예 및 조건부 언어는 주로 본 발명의 조성물 및 방법의 원리 및 발명자가 당해 기술을 발전시키는 데 기여한 개념을 독자가 이해하도록 돕는 것을 목적으로 하며, 이렇게 구체적으로 열거된 실시예와 조건에 제한하지 않는 것으로 해석되어야 한다. 또한, 본 조성물 및 방법의 원리, 양태 및 실시예뿐만 아니라 이의 특정 실시예를 기재한 본원의 모든 기재 내용은 이의 구조적 및 기능적 등가물 모두를 포함하는 것으로 의도된다. 또한, 이러한 등가물은 현재 알려진 등가물 및 미래에 개발되는 등가물, 즉 구조와 상관없이 동일한 기능을 수행하는 임의의 개발된 요소를 모두 포함하는 것으로 의도된다. 따라서, 본 조성물 및 방법의 범위는 본원에 도시되고 기재된 예시적인 실시예에 한정되는 것으로 의도되지 않는다.
서열:
서열 번호 1:
추정 T. 레에세이 U6 유전자
AAAAAACACTAGTAAGTACTTACTTATGTATTATTAACTACTTTAGCTAACTTCTGCAGTACTACCTAAGAGGCTAGGGGTAGTTTTATAGCAGACTTATAGCTATTATTTTTATTTAGTAAAGTGCTTTTAAAGTAAGGTCTTTTTTATAGCACTTTTTATTTATTATAATATATATTATATAATAATTTTAAGCCTGGAATAGTAAAGAGGCTTATATAATAATTTATAGTAATAAAAGCTTAGCAGCTGTAATATAATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATCGCCTTCGGGCATTTGGTCAATTTATAACGATACAGGTTCGTTTCGGCTTTTCCTCGGAACCCCCAGAGGTCATCAGTTCGAATCGCTAACAGGTCAACAGAGAAGATTAGCATGGCCCCTGCACTAAGGATGACACGCTCACTCAAAGAGAAGCTAAACATTTTTTTTCTCTTCCAAGTCGTGATGGTTATCTTTTTGCTTAGAGAATCTATTCTTGTGGACGATTAGTATTGGTAAATCCCTGCTGCACATTGCGGCGGATGGTCTCAACGGCATAATACCCCATTCGTGATGCAGCGGTGATCTTCAATATGTAGTGTAATACGTTGCATACACCACCAGGTTCGGTGCCTCCTGTATGTACAGTACTGTAGTTCGACTCCTCCGCGCAGGTGGAAACGATTCCCTAGTGGGCAGGTATTTTGGCGGGGTCAAGAA
서열 번호 2:
sgRNA의 서열(N은 표적 부위에 상보적인 서열)
NNNNNNNNNNNNNNNNNNNNGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
서열 번호 3:
sgRNA: gAd3A TS1
guccucgagcaaaaggugccGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
서열 번호 4:
sgRNA: gTrGA TS2
guucagugcaauaggcgucuGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
서열 번호 5:
sgRNA: gTrGA TS11
gccaauggcgacggcagcacGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
서열 번호 6:
sgRNA: gPyr2 TS6
gcacagcgggaugcccuuguGUUUUAGAGCUAGAAAUAGCAAGUUAAAAUAAGGCUAGUCCGUUAUCAACUUGAAAAAGUGGCACCGAGUCGGUGGUGC
서열 번호 7:
코돈 최적화된 스트렙토코쿠스 파이오제네스 Cas9-인코딩 유전자; N- 및 C-말단 NLS 서열을 가짐
acgcatcgacctgagccagctgggcggagacaagaagaagaagctcaagctctag
서열 번호 8:
서열 번호 7에 의해 인코딩된 스트렙토코쿠스 파이오제네스 Cas9; N- 및 C-말단 NLS 서열을 가짐
DVRKMIAKSEQEIGKATAKYFFYSNIMNFFKTEITLANGEIRKRPLIETNGETGEIVWDKGRDFATVRKVLSMPQVNIVKKTEVQTGGFSKESILPKRNSDKLIARKKDWDPKKYGGFDSPTVAYSVLVVAKVEKGKSKKLKSVKELLGITIMERSSFEKNPIDFLEAKGYKEVKKDLIIKLPKYSLFELENGRKRMLASAGELQKGNELALPSKYVNFLYLASHYEKLKGSPEDNEQKQLFVEQHKHYLDEIIEQISEFSKRVILADANLDKVLSAYNKHRDKPIREQAENIIHLFTLTNLGAPAAFKYFDTTIDRKRYTSTKEVLDATLIHQSITGLYETRIDLSQLGGDKKKKLKL
서열 번호 9:
합성 DNA: gAd3A TS1-1(사카로마이세스 세레비지애 snr52 프로모터 및 S. 세레비지애 sup4 종결인자를 갖는 gAd3A TS1 sgRNA(서열 번호 3))
gaattcggatccTCTTTGAAAAGATAATGTATGATTATGCTTTCACTCATATTTATACAGAAACTTGATGTTTTCTTTCGAGTATATACAAGGTGATTACATGTACGTTTGAAGTACAACTCTAGATTTTGTAGTGCCCTCTTGGGCTAGCGGTAAAGGTGCGCATTTTTTCACACCCTACAATGTTCTGTTCAAAAGATTTTGGTCAAACGCTGTAGAAGTGAAAGTTGGTGCGCATGTTTCGGCGTTCGAAACTTCTCCGCAGTGAAAGATAAATGATCgtcctcgagcaaaaggtgccGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGGTGCTTTTTTTGTTTTTTATGTCTgaattcggatcc
서열 번호 10:
합성 DNA: gAd3A TS1-2(T. 레에세이 U6 프로모터 및 종결인자를 갖는 gAd3A TS1 sgRNA(서열 번호 3))
gaattcggatccAAAAAACACTAGTAAGTACTTACTTATGTATTATTAACTACTTTAGCTAACTTCTGCAGTACTACCTAAGAGGCTAGGGGTAGTTTTATAGCAGACTTATAGCTATTATTTTTATTTAGTAAAGTGCTTTTAAAGTAAGGTCTTTTTTATAGCACTTTTTATTTATTATAATATATATTATATAATAATTTTAAGCCTGGAATAGTAAAGAGGCTTATATAATAATTTATAGTAATAAAAGCTTAGCAGCTGTAATATAATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATCgtcctcgagcaaaaggtgccGTTTTAGAGCTAGAAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGGTGCTTTTTTTTCTCTTgaattcggatcc
서열 번호 11:
합성 DNA: gAd3A TS1-3(T. 레에세이 U6 프로모터, 종결인자 및 인트론을 갖는 gAd3A TS1 sgRNA(서열 번호 3))
gaattcggatccAAAAAACACTAGTAAGTACTTACTTATGTATTATTAACTACTTTAGCTAACTTCTGCAGTACTACCTAAGAGGCTAGGGGTAGTTTTATAGCAGACTTATAGCTATTATTTTTATTTAGTAAAGTGCTTTTAAAGTAAGGTCTTTTTTATAGCACTTTTTATTTATTATAATATATATTATATAATAATTTTAAGCCTGGAATAGTAAAGAGGCTTATATAATAATTTATAGTAATAAAAGCTTAGCAGCTGTAATATAATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATCgtcctcgagcaaaaggtgccGTTTTAGAGCTAGAGTTCGTTTCGGCTTTTCCTCGGAACCCCCAGAGGTCATCAGTTCGAATCGCTAACAGAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGGTGCTTTTTTTTCTCTTgaattcggatcc
서열 번호 12:
보다 짧은 T. 레에세이 U6 프로모터 영역을 갖는 가이드 RNA 발현 카세트를 합성 DNA로서 수득 하였다. TS11에서의 T. 레에세이 gla1 유전자를 표적으로 하는 sgRNA에 대한 서열을 포함하는 예가 여기에 제공된다.
AATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATCgccaatggcgacggcagcacGTTTTAGAGCTAGAGTTCGTTTCGGCTTTTCCTCGGAACCCCCAGAGGTCATCAGTTCGAATCGCTAACAGAATAGCAAGTTAAAATAAGGCTAGTCCGTTATCAACTTGAAAAAGTGGCACCGAGTCGGTGGTGCTTTTTTTTCTCTT
서열 번호 13:
프라이머: gRNA fwd aflII
cgtcagcttaagAATTCCTAAAGAAACAGCATGAAATGG
서열 번호 14:
프라이머: gRNA rev sfiI
cgtcagggccacgtgggccAAGAGAAAAAAAAGCACCACCGACTCGG
서열 번호 15:
프라이머: Ad3 5' fwd
tgaacacagccaccgacatcagc
서열 번호 16:
프라이머: Ad3 5' rev
gctggtgagggtttgtgctattg
서열 번호 17:
프라이머: Ad3a 5005 rev
gattgcttgggaggaggacat
서열 번호 18:
프라이머: Ad3 3' fwd
cgaggccactgatgaagttgttc
서열 번호 19:
프라이머: Ad3 3' rev
Cagttttccaaggctgccaacgc
서열 번호 20:
프라이머: Ad3a 5003 fwd
ctgatcttgcaccctggaaatc
서열 번호 21:
Ad3mid rev
ctctctatcatttgccaccctcc
서열 번호 22:
프라이머: Adfrag fwd
ctccattcaccctcaattctcc
서열 번호 23:
프라이머: Adfrag rev
gttcccttggcggtgcttggatc
서열 번호 24:
프라이머: Ad3a 2k fwd
caatagcacaaaccctcaccagc
서열 번호 25:
Ad3a 2k rev
gaacaacttcatcagtggcctcg
서열 번호 26:
프라이머: glaA
ccgttagttgaagatccttgccg
서열 번호 27:
프라이머: glaB
gtcgaggatttgcttcatacctc
서열 번호 28:
프라이머: glaD
gagagacgcaggatgactcaaag
서열 번호 29:
프라이머: glaH
tgccgtgggtcattggcatattc
서열 번호 30:
프라이머: glaJ
tgccgactttgtccagtgattcg
서열 번호 31:
프라이머: glaK
ttacatgtggacgcgagatagcg
서열 번호 32:
프라이머: gla1repF
gtgtgtctaatgcctccaccac
서열 번호 33:
프라이머: gla1repR
gatcgtgctagcgctgctgttg
서열 번호 34:
프라이머: 1553R
CCGTGATGGAGCCCGTCTTCT
서열 번호 35:
프라이머: 1555F
CGCGGTGAGTTCAGGCTTTTTC
서열 번호 36:
프라이머: pyr2F
gtataagagcaggaggagggag
서열 번호 37:
프라이머: pyr2R
gaacgcctcaatcagtcagtcg
서열 번호 38
트리코데르마 레에세이 텔로미어 서열간 박테리아 카나마이신 저항 유전자(프로모터 및 종결 인자를 가짐)
atagaccagctttacataataatcgcctgttgctactgactgatgaccttcttccctaaccagtttcctaattaccactgcagtgaggataaccctaactcgctctggggttattattatactgattagcaggtggcttatatagtgctgaagtactataagagtttctgcgggaggaggtggaaggactataaactggacacagttagggatagagtgatgacaagacctgaatgttatcctccggtgtggtatagcgaattggctgaccttgcagatggtaatggtttaggcagggtttttgcagagggggacgagaacgcgttctgcgatttaacggctgctgccgccaagctttacggttctctaatgggcggccgc
서열 번호 39:
합성 DNA: Gla1rep
gtgtgtctaatgcctccaccacaggaaccaaaccggctttgacctctgggaagaagtcaatgggagctcattctttactgttgccaaccagcaccgaggtatgaagcaaatcctcgacattcgctgctactgcacatgagcattgttactgaccagctctacagcacttgtcgagggcgccactcttgctgccactcttggccagtcgggaagcgcttattcatctgttgctccccaggttttgtgctttctccaacgattctgggtgtcgtctggtggatacgtcgactccaacagtatgtcttttcactgtttatatgagattggccaatactgatagctcgcctctagtcaacaccaacgagggcaggactggcaaggatgtcaactccgtcctgacttccatccacaccttcgatcccaaccttggctgtgacgcaggcaccttccagccatgcagtgacaaagcgctctccaacctcaaggttgttgtcgactccttccgctccatctacggcgtgaacaagggcattcctgCGgtgctgccgtcgccattggccggtatgcagaggatgtgtactacaacggcaacccttggtatcttgctacatttgctgctgccgagcagctgtacgatgccatctacgtctggaagaagacgggctccatcacggtgaccgccacctccctggccttcttccaggagcttgttcctggcgtgacggccgggacctactccagcagctcttcgacctttaccaacatcatcaacgccgtctcgacatacgccgatggcttcctcagcgaggctgccaagtacgtccccgccgacggttcgctggccgagcagtttgaccgcaacagcggcactccgctgtctgcgcttcacctgacgtggtcgtacgcctcgttcttgacagccacggcccgtcgggctggcatcgtgcccccctcgtgggccaacagcagcgctagcacgatc
서열 번호 40:
전체 U6 유전자 프로모터 서열(전사 개시 부위를 포함하지 않음)
AAAAAACACTAGTAAGTACTTACTTATGTATTATTAACTACTTTAGCTAACTTCTGCAGTACTACCTAAGAGGCTAGGGGTAGTTTTATAGCAGACTTATAGCTATTATTTTTATTTAGTAAAGTGCTTTTAAAGTAAGGTCTTTTTTATAGCACTTTTTATTTATTATAATATATATTATATAATAATTTTAAGCCTGGAATAGTAAAGAGGCTTATATAATAATTTATAGTAATAAAAGCTTAGCAGCTGTAATATAATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATC
서열 번호 41:
절단된/더 짧은 U6 프로모터 서열(전사 시작 부위를 포함하지 않음)
AATTCCTAAAGAAACAGCATGAAATGGTATTATGTAAGAGCTATAGTCTAAAGGCACTCTGCTGGATAAAAATAGTGGCTATAAGTCTGCTGCAAAACTACCCCCAACCTCGTAGGTATATAAGTACTGTTTGATGGTAGTCTATC
서열 번호 42:
U6 유전자 인트론
GTTCGTTTCGGCTTTTCCTCGGAACCCCCAGAGGTCATCAGTTCGAATCGCTAA
CAG
서열 번호 43:
U6 유전자 전사 종결인자 서열
TTTTTTTTCTCTT
서열 번호 44:
사상균 세포 코돈에 최적화된 스트렙토코쿠스 파이오제네스 Cas9-인코딩 유전자; NLS 없음
agac
서열 번호 45:
서열 번호 44에 의해 인코딩된 스트렙토코쿠스 파이오제네스 Cas9; NLS 없음
EQEIGKATAKYFFYSNIMNFFKTEITLANGEIRKRPLIETNGETGEIVWDKGRDFATVRKVLSMPQVNIVKKTEVQTGGFSKESILPKRNSDKLIARKKDWDPKKYGGFDSPTVAYSVLVVAKVEKGKSKKLKSVKELLGITIMERSSFEKNPIDFLEAKGYKEVKKDLIIKLPKYSLFELENGRKRMLASAGELQKGNELALPSKYVNFLYLASHYEKLKGSPEDNEQKQLFVEQHKHYLDEIIEQISEFSKRVILADANLDKVLSAYNKHRDKPIREQAENIIHLFTLTNLGAPAAFKYFDTTIDRKRYTSTKEVLDATLIHQSITGLYETRIDLSQLGGD
서열 번호 46:
SV40 NLS
PKKKRKV
서열 번호 47
T. 레에세이 blr2 (청색광 조절인자 2) 유전자 NLS
KKKKLKL
서열 번호 48:
스트렙토코쿠스 더모필루스 LMD-9 Cas9
FRERKSATEKVYFYSNIMNIFKKSISLADGRVIERPLIEVNEETGESVWNKESDLATVRRVLSYPQVNVVKKVEEQNHGLDRGKPKGLFNANLSSKPKPNSNENLVGAKEYLDPKKYGGYAGISNSFTVLVKGTIEKGAKKKITNVLEFQGISILDRINYRKDKLNFLLEKGYKDIELIIELPKYSLFELSDGSRRMLASILSTNNKRGEIHKGNQIFLSQKFVKLLYHAKRISNTINENHRKYVENHKKEFEELFYYILEFNENYVGAKKNGKLLNSAFQSWQNHSIDELCSSFIGPTGSERKGLFELTSRGSAADFEFLGVKIPRYRDYTPSSLLKDATLIHQSVTGLYETRIDLAKLGEG
서열 번호 49:
스트렙토코쿠스 뮤탄스 UA159 Cas9
AKKFFYSNIMNFFKKDDVRTDKNGEIIWKKDEHISNIKKVLSYPQVNIVKKVEEQTGGFSKESILPKGNSDKLIPRKTKKFYWDTKKYGGFDSPIVAYSILVIADIEKGKSKKLKTVKALVGVTIMEKMTFERDPVAFLERKGYRNVQEENIIKLPKYSLFKLENGRKRLLASARELQKGNEIVLPNHLGTLLYHAKNIHKVDEPKHLDYVDKHKDEFKELLDVVSNFSKKYTLAEGNLEKIKELYAQNNGEDLKELASSFINLLTFTAIGAPATFKFFDKNIDRKRYTSTTEILNATLIHQSITGLYETRIDLNKLGGD
서열 번호 50:
캄피로박테르 제주니 Cas9
MARILAFDIGISSIGWAFSENDELKDCGVRIFTKVENPKTGESLALPRRLARSARKRLARRKARLNHLKHLIANEFKLNYEDYQSFDESLAKAYKGSLISPYELRFRALNELLSKQDFARVILHIAKRRGYDDIKNSDDKEKGAILKAIKQNEEKLANYQSVGEYLYKEYFQKFKENSKEFTNVRNKKESYERCIAQSFLKDELKLIFKKQREFGFSFSKKFEEEVLSVAFYKRALKDFSHLVGNCSFFTDEKRAPKNSPLAFMFVALTRIINLLNNLKNTEGILYTKDDLNALLNEVLKNGTLTYKQTKKLLGLSDDYEFKGEKGTYFIEFKKYKEFIKALGEHNLSQDDLNEIAKDITLIKDEIKLKKALAKYDLNQNQIDSLSKLEFKDHLNISFKALKLVTPLMLEGKKYDEACNELNLKVAINEDKKDFLPAFNETYYKDEVTNPVVLRAIKEYRKVLNALLKKYGKVHKINIELAREVGKNHSQRAKIEKEQNENYKAKKDAELECEKLGLKINSKNILKLRLFKEQKEFCAYSGEKIKISDLQDEKMLEIDHIYPYSRSFDDSYMNKVLVFTKQNQEKLNQTPFEAFGNDSAKWQKIEVLAKNLPTKKQKRILDKNYKDKEQKNFKDRNLNDTRYIARLVLNYTKDYLDFLPLSDDENTKLNDTQKGSKVHVEAKSGMLTSALRHTWGFSAKDRNNHLHHAIDAVIIAYANNSIVKAFSDFKKEQESNSAELYAKKISELDYKNKRKFFEPFSGFRQKVLDKIDEIFVSKPERKKPSGALHEETFRKEEEFYQSYGGKEGVLKALELGKIRKVNGKIVKNGDMFRVDIFKHKKTNKFYAVPIYTMDFALKVLPNKAVARSKKGEIKDWILMDENYEFCFSLYKDSLILIQTKDMQEPEFVYYNAFTSSTVSLIVSKHDNKFETLSKNQKILFKNANEKEVIAKSIGIQNLKVFEKYIVSALGEVTKAEFRQREDFKK
서열 번호 51:
나이세리아 메닝기티데스 Cas9
GTGNINIRIHDLDHKIGKNGILEGIGVKTALSFQKYQIDELGKEIRPCRLKKRPPVR
서열 번호 52:
프란시셀라 투라렌시스 아종 노비시다 Cas9
EKKIADTIWDANKKDFKFGNYRSFINLTPQEQKAFRHALFLADENPIKQAVIRAINNRNRTFVNGTQRYFAEVLANNIYLRAKKENLNTDKISFDYFGIPTIGNGRGIAEIRQLYEKVDSDIQAYAKGDKPQASYSHLIDAMLAFCIAADEHRNDGSIGLEIDKNYSLYPLDKNTGEVFTKDIFSQIKITDNEFSDKKLVRKKAIEGFNTHRQMTRDGIYAENYLPILIHKELNEVRKGYTWKNSEEIKIFKGKKYDIQQLNNLVYCLKFVDKPISIDIQISTLEELRNILTTNNIAATAEYYYINLKTQKLHEYYIENYNTALGYKKYSKEMEFLRSLAYRSERVKIKSIDDVKQVLDKDSNFIIGKITLPFKKEWQRLYREWQNTTIKDDYEFLKSFFNVKSITKLHKKVRKDFSLPISTNEGKFLVKRKTWDNNFIYQILNDSDSRADGTKPFIPAFDISKNEIVEAIIDSFTSKNIFWLPKNIELQKVDNKNIFAIDTSKWFEVETPSDLRDIGIATIQYKIDNNSRPKVRVKLDYVIDDDSKINYFMNHSLLKSRYPDKVLEILKQSTIIEFESSGFNKTIKEMLGMKLAGIYNETSNN
서열 번호 53:
파스튜렐라 물토시다 Cas9
VKLALSFEKYQVDELGKNRQICRPQQRQ
PVR
서열 번호 54
플라스미드 pET30a-Cas9-D10A 닉카아제 내 합성된 Cas9-D10A 닉카아제 유전자의 뉴클레오티드 서열. N-말단 His6 태그를 인코딩하는 올리고뉴클레오티드는 밑줄이 쳐져 이탤릭체로 표기되고, SV40 핵 위치 신호를 인코딩하는 올리고뉴클레오티드는 이탤릭체 및 회색 하이라이트로 표시되고, BLR 핵 위치 신호를 인코딩하는 올리고뉴클레오티드는 밑줄이 쳐져 회색으로 강조 표시된다.
서열 번호 55:
플라스미드 pET30a-Cas9-D10A 닉카아제(서열 번호 54에 의해 인코딩됨)에서 발현된 Cas9-D10A 닉카아제 단백질의 아미노산 서열. N-말단 His6 태그는 밑줄과 이탤릭체로 표시되고, 돌연변이된 아미노산은 굵은 밑줄(D10A)로 표시되고, SV40 핵 위치 신호는 이탤릭 및 회색으로 표시되고, BLR 핵 위치 신호는 밑줄 쳐져 회색으로 표시된다.
서열 번호 56
기질 DNA 단편의 뉴클레오티드 서열. UTR 서열은 소문자로 나타나고 TrGA 유전자는 대문자로 나타난다. 두 개의 선택된 VT 도메인인 TrGA_sgF1과 TrGA_sgR1은 각각 굵은 글씨와 밑줄로 나타난다.
서열 번호 57:
서열 번호 56을 위한 정방향 프라이머:
5'- gactgtctccaccatgtaatttttc-3'
서열 번호 58:
TrGA 프라이머 KOR2 ; 서열 번호 56을 위한 역방향 프라이머:
5'-ggcagactacaagtctactagtactac-3'
서열 번호 59:
T7 프로모터, CER 도메인, 및 VT 도메인 TrGA_sgF1의 전사를 위한 올리고뉴클레오티드 서열. VT 도메인은 대문자로 표시되어 있지만, T7 프로모터와 CER 도메인 영역은 각각 굵은 글씨와 소문자로 표시되어 있다.
서열 번호 60:
T7 프로모터, CER 도메인, 및 VT 도메인 TrGA_sgR1의 전사를 위한 올리고뉴클레오티드 서열. VT 도메인은 대문자로 표시되어 있지만, T7 프로모터와 CER 도메인 영역은 각각 굵은 글씨와 소문자로 표시되어 있다.
서열 번호 61:
TrGA 녹아웃 카세트의 뉴클레오티드 서열:
agatgccggccttgaagcagtaggtcgagaggctccacttcatggtgccgttgccgatcatggtgttgatgatgcggtcgtacgtctcgatggcgccgtagtagtcgccgtcgagggcggcgaggtcggcgtactgcgtccagagctt
서열 번호 62:
Fw1 (5'-cactactacatccatacgcagcaaacatgg-3') 및
서열 번호 63:
R3 (5'-ggtcaagaagcacatgccagagttcg -3')
서열 번호 64:
TrGAF2 (5' gactgtctccaccatgtaatttttc 3')
서열 번호 65:
대장균 코돈에 최적화된 Cas9-D10A 닉카아제 유전자(중지 코돈 없음) agac
서열 번호 66:
스트렙토코쿠스 파이오제네스로부터의 Cas9-D10A 닉카아제의 아미노산 서열; 서열 번호 65에 의해 인코딩됨
eqeigkatakyffysnimnffkteitlangeirkrplietngetgeivwdkgrdfatvrkvlsmpqvnivkktevqtggfskesilpkrnsdkliarkkdwdpkkyggfdsptvaysvlvvakvekgkskklksvkellgitimerssfeknpidfleakgykevkkdliiklpkyslfelengrkrmlasagelqkgnelalpskyvnflylashyeklkgspedneqkqlfveqhkhyldeiieqisefskrviladanldkvlsaynkhrdkpireqaeniihlftltnlgapaafkyfdttidrkrytstkevldatlihqsitglyetridlsqlggd
서열 번호 67:
(시작 메티오닌을 갖는) N-말단 His6 tag/트롬빈/S·Tag™ /엔테로키나아제 영역 아미노산 서열
mhhhhhhssglvprgsgmketaaakferqhmdspdlgtddddkama
서열 번호 68:
(시작 코돈을 갖는) N-말단 His6 tag/트롬빈/S·Tag™ /엔테로키나아제 영역 폴리뉴클레오티드 서열; 서열 번호 67을 인코딩
atgcaccatcatcatcatcattcttctggtctggtgccacgcggttctggtatgaaagaaaccgctgctgctaaattcgaacgccagcacatggacagcccagatctgggtaccgacgacgacgacaaggccatggcc
서열 번호 69:
SV40 NLS 코딩 서열(서열 번호 46을 인코딩)
ccaaaaaagaaacgcaaggtt
서열 번호 70:
BLR 핵 위치 신호 코딩 서열(서열 번호 47을 인코딩)
Aagaagaaaaaactgaaactg
서열 번호 71:
tgatgacggtgaaaacctc
서열 번호 72:
aaaagcaccgactcgg
서열 번호 73:
Gla1rep 프라이머 1556F
ATGCGCAAATTTAAAGCGCTGATgtgtgtctaatgcctccaccac
서열 번호 74:
Gla1rep 프라이머 1557R
ATATGGATCTGCGCGCGATCGATgatcgtgctagcgctgctgttg
서열 번호 75:
서열분석 프라이머 1538F
CCACCACAGGAACCAAACC
서열 번호 76:
서열분석 프라이머 1539R
CTGCGACGGAGGGAATGACG
서열 번호 77:
서열분석 프라이머 1540F
GGGCAGGACTGGCAAGGATGT
서열 번호 78:
서열분석 프라이머 1541R
GCCGTCACGCCAGGAACAAG
서열 번호 79:
TrGA 프라이머 KOF1
gaacaatcttctttgcaatgttggtc
서열 번호 80:
Pyr2 프라이머 R1
gaggaagtcctgcttgtaggcaggc
서열 번호 81:
Pyr2 프라이머 F4
cgacagagcagtcatatggggatacg