FR2807764A1 - Mutants de bacteries lactiques surproducteurs d'exopolysaccharides - Google Patents
Mutants de bacteries lactiques surproducteurs d'exopolysaccharides Download PDFInfo
- Publication number
- FR2807764A1 FR2807764A1 FR0004971A FR0004971A FR2807764A1 FR 2807764 A1 FR2807764 A1 FR 2807764A1 FR 0004971 A FR0004971 A FR 0004971A FR 0004971 A FR0004971 A FR 0004971A FR 2807764 A1 FR2807764 A1 FR 2807764A1
- Authority
- FR
- France
- Prior art keywords
- sep
- gene
- lactic acid
- pgm
- eps
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Granted
Links
Classifications
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/10—Transferases (2.)
- C12N9/12—Transferases (2.) transferring phosphorus containing groups, e.g. kinases (2.7)
- C12N9/1241—Nucleotidyltransferases (2.7.7)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N1/00—Microorganisms; Compositions thereof; Processes of propagating, maintaining or preserving microorganisms or compositions thereof; Processes of preparing or isolating a composition containing a microorganism; Culture media therefor
- C12N1/20—Bacteria; Culture media therefor
- C12N1/205—Bacterial isolates
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/90—Isomerases (5.)
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12P—FERMENTATION OR ENZYME-USING PROCESSES TO SYNTHESISE A DESIRED CHEMICAL COMPOUND OR COMPOSITION OR TO SEPARATE OPTICAL ISOMERS FROM A RACEMIC MIXTURE
- C12P19/00—Preparation of compounds containing saccharide radicals
- C12P19/04—Polysaccharides, i.e. compounds containing more than five saccharide radicals attached to each other by glycosidic bonds
-
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12R—INDEXING SCHEME ASSOCIATED WITH SUBCLASSES C12C - C12Q, RELATING TO MICROORGANISMS
- C12R2001/00—Microorganisms ; Processes using microorganisms
- C12R2001/01—Bacteria or Actinomycetales ; using bacteria or Actinomycetales
- C12R2001/46—Streptococcus ; Enterococcus; Lactococcus
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Health & Medical Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Genetics & Genomics (AREA)
- Biotechnology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Medicinal Chemistry (AREA)
- Molecular Biology (AREA)
- Tropical Medicine & Parasitology (AREA)
- Chemical Kinetics & Catalysis (AREA)
- General Chemical & Material Sciences (AREA)
- Virology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
- Preparation Of Compounds By Using Micro-Organisms (AREA)
- Enzymes And Modification Thereof (AREA)
- Pharmaceuticals Containing Other Organic And Inorganic Compounds (AREA)
- Polysaccharides And Polysaccharide Derivatives (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
La présente invention a pour objet des mutants de bactéries lactiques surproducteurs d'exopolysaccharides à la suite d'une mutation dans le gène codant l'alpha-phosphoglucomutase.Ces mutants sont utilisables notamment pour la préparation de produits fermentés ou la production d'exopolysaccharides.
Description
<Desc/Clms Page number 1>
MUTANTS DE BACTERIES LACTIQUES SURPRODUCTEURS
D'EXOPOLYSACCHARIDES
L'invention est relative à la régulation de la production d'hétéropolysaccharides exocellulaires par des bactéries lactiques.
D'EXOPOLYSACCHARIDES
L'invention est relative à la régulation de la production d'hétéropolysaccharides exocellulaires par des bactéries lactiques.
De manière générale, les polysaccharides sont très utilisés comme additifs dans l'alimentation, mais aussi dans les cosmétiques et les produits pharmaceutiques, par exemple en tant qu'agents épaississants et/ou gélifiants, stabilisateurs de texture, succédanés de matière grasse, etc. Parmi les polysaccharides utilisés de la sorte, on citera notamment ceux produits par des microorganismes en particulier des bactéries, tels que des dextranes, les xanthanes, les gellanes, pullulanes etc.
De nombreuses espèces de bactéries lactiques, notamment des lactocoques tels que Lactococcus lactis, des Leuconostoc tels que Leuconostoc mesenteroide, des streptocoques tels que Streptococcus thermophilus, et des lactobacilles tels que Lactobacillus casei, Lactobacillus sake, Lactobacillus rhamnosus, Lactobacillus acidophilus, Lactobacillus delbrueckii subsp. bulgaricus, Lactobacillus helveticus, etc. produisent des polysaccharides.
Ceux-ci peuvent être regroupés en 2 catégories : les homopolysaccharides, tels que les dextranes, qui résultent de la polymérisation d'un seul sucre, et les hétéropolysaccharides, de structure complexe, associant des unités de base constituées de 2 ou plusieurs sucres différents (fréquemment le D-galactose, le D-glucose et le Lrhamnose).
Les hétéropolysaccharides des bactéries lactiques sont habituellement désignés sous le terme général d'EPS (pour exopolysaccharides), qui sera également utilisé ciaprès. Ils jouent un rôle majeur dans l'élaboration de la texture, de la perception en bouche et de la rhéologie des produits laitiers fermentés. En outre, il a été observé que certains d'entre eux possédaient des activités biologiques par lesquelles ils pourraient exercer divers effets bénéfiques sur la santé. [pour revue, cf. DE VUYST, et DEGEEST, FEMS Microbiology Reviews, 23,153-177, (1999)].
<Desc/Clms Page number 2>
Toutefois, la quantité d'hétéropolysaccharides produits par les bactéries lactiques est en général faible (de l'ordre de 10 à 200 mg par litre de produit fermenté).
Pour améliorer la texture, les fabricants de produits fermentés rajoutent d'autres agents texturants comme des stabilisateurs (amidons modifiés, carragheen, guar, pectine, gélatine...). Cependant, ces ajouts ne sont pas toujours autorisés (par exemple dans le yaourt nature), et affectent généralement le goût et l'arôme du produit. Une production optimisée d'hétéropolysaccharides exocellulaires (EPS) dans le produit est donc préférable.
Etant donné l'importance des EPS dans les industries agro-alimentaires, évoquée précédemment, de nombreux travaux ont porté sur des méthodes permettant d'augmenter leur production par optimisation des procédés biotechnologiques en agissant sur la température, le pH et la composition du milieu. Ces approches sont cependant parfois difficiles à appliquer dans le cadre de certains procédés agro-alimentaires comme la fabrication de produits laitiers fermentés pour lesquels le milieu et les conditions de fermentation sont propres à chaque type de produit. Ces contraintes de fabrication limitent donc l'utilisation des méthodes classiques d'optimisation des procédés pour améliorer la production des EPS. Une alternative à ces méthodes serait d'utiliser des souches adaptées à ces procédés, c'est-à-dire capables de produire des EPS en plus grande quantité, et/ou dont la production d'EPS puisse être contrôlée dans les conditions de production des produits fermentés.
L'une des limitations principales de la production d'EPS par les bactéries lactiques pourrait provenir d'une compétition entre la biosynthèse de ces EPS et d'autres voies métaboliques pour l'utilisation des sucres disponibles. En effet, alors que les homopolysaccharides sont principalement produits par des enzymes extracellulaires spécifiques à partir de substrats présents dans le milieu, la synthèse des hétéropolysaccharides s'effectue au moins en partie à l'intérieur de la cellule. En particulier la formation des précurseurs (sucres nucléotidiques) des
<Desc/Clms Page number 3>
hétéropolysaccharides, constitués par des sucres activés par réaction avec des nucléotides triphosphate pourrait faire intervenir des enzymes intracytoplasmiques qui participent également à d'autres voies métaboliques, et notamment à la glycolyse. Or, du fait du métabolisme fermentaire des bactéries lactiques, les réactions de la glycolyse sont plus actives que celles ayant trait à la synthèse des EPS.
Les Inventeurs se sont fixé pour but l'obtention de mutants de bactéries lactiques dont on puisse contrôler et notamment augmenter la capacité à utiliser des sucres disponibles dans le milieu, et en particulier le galactose, pour produire des EPS.
Dans ce but, ils ont recherché les gènes impliqués dans la synthèse sucres nucléotidiques des précurseurs des EPS, et/ou les gènes intervenant au carrefour des voies de la glycolyse et de la biosynthèse des EPS. Parmi ces derniers, ils se sont intéressés plus particulièrement au gène de l'alpha-phosphoglucomutase (a-PGM), qui intervient dans la transformation des dérivés métaboliques du galactose en intermédiaires de la glycolyse, ainsi qu'au gène galU qui code la Glucose-1-phosphate uridyl transférase, qui catalyse la formation des sucres nucléotidiques, précurseurs des EPS.
DE VUYST, et DEGEEST, (publication précitée) émettent l'hypothèse que la phosphoglucomutase pourrait jouer un rôle important de liaison entre la glycolyse et la biosynthèse des EPS, et que le détournement vers cette enzyme d'une partie du flux carboné pourrait permettre d'augmenter la production d'EPS. Toutefois, ils soulignent également qu'il reste à savoir si ceci est effectivement réalisable.
Il a été récemment rapporté [HARDY et al., J Bacteriol., 1854-1863, (2000)] que l'inactivation chez Streptococcus pneumoniae d'un gène [GENBANK AF165218] codant une phosphoglucomutase entraînait une diminution drastique de la production des EPS formant la capsule bactérienne, et était en outre fortement préjudiciable à la viabilité cellulaire, bien que cette bactérie possède un autre gène pgm .
Aucun gène susceptible de coder une a-PGM de bactérie lactique n'a été caractérisé génétiquement jusqu'à
<Desc/Clms Page number 4>
présent. L'existence d'une a-PGM et d'une P-PGM a été rapportée chez L. Lactis ; toutefois, seul le gène corrrespondant à la P-PGM a été isolé [QIAN et al., Microbiology, 143,855-865, (1997)].
Les Inventeurs sont maintenant parvenus à cloner et caractériser un gène pgm, codant une a-PGM de Streptococcus thermophilus.
Ce gène est représenté dans la liste de séquences en annexe sous le numéro SEQ ID NO: 1, et le polypeptide correspondant est représenté sous le numéro SEQ ID NO: 2.
La présente Invention a pour objet une séquence d'acide nucléique codant un polypeptide dont la séquence en acides aminés a au moins 70% d'identité ou au moins 85% de similarité, de préférence 80% d'identité ou au moins 90% de similarité, avantageusement au moins 90% d'identité ou au moins 95% de similarité, et de manière tout à fait préférée au moins 95% d'identité ou au moins 99% de similarité, avec le polypeptide SEQ ID NO: 2.
Le pourcentage d'identité d'une séquence avec une séquence de référence est défini ici comme le pourcentage de résidus de cette séquence qui sont identiques avec ceux de la séquence de référence lorsque les 2 séquences sont alignées pour une correspondance maximale entre les positions des résidus. Un polypeptide dont la séquence en acides aminés possède au moins X% d'identité avec une séquence de référence peut ainsi comprendre jusqu'à 100-X modifications pour 100 acides aminés de la séquence de référence. Ces modifications incluent la délétion, la substitution, ou l'insertion de résidus d'acides aminés, consécutifs ou non.
Le pourcentage de similarité d'une séquence avec une séquence de référence est défini ici comme le pourcentage de résidus de cette séquence qui sont identiques avec ceux de la séquence de référence ou qui n'en diffèrent que par une substitution conservative, lorsque les 2 séquences sont alignées pour une correspondance maximale entre les positions des résidus. On entend par "substitution conservative" la substitution d'un résidu d'acide aminé par un autre résidu possédant des caractéristiques physicochimiques (taille, charge, ou polarité) similaires, qui ne changent pas les
<Desc/Clms Page number 5>
propriétés fonctionnelles de la protéine. Un polypeptide dont la séquence en acides aminés possède au moins X% de similarité avec une séquence de référence peut ainsi comprendre jusqu'à 100-X modifications non-conservatives pour 100 acides aminés de la séquence de référence. Ces modifications incluent la délétion, la substitution nonconservative, ou l'insertion de résidus d'acides aminés, consécutifs ou non.
Les polypeptides présentant ainsi les pourcentages d'identité ou de similarité les plus importants avec la séquence SEQ ID NO: 2, identifiés par recherche sur la base de données "GENBANK nr" en utilisant le logiciel BLASTp [ALTSCHUL et al., Nucleic Acids Res., 25,3389-3402, (1997) ] sont les suivants : - la protéine ybbT de Bacillus subtilis . 57% d'identité, et 69% de similarité ; - la protéine femD de Staphylococcus aureus subtilis : 53% d'identité, et 69% de similarité ; la phosphoglucomutase hypothétique de Streptomyces coelicolor : 42% d'identité, et 55% de similarité ; - l'homologue de mrsA de Pseudomonas syr ingae : 41% d'identité, et 54% de similarité ; - l'homologue de mrsA de Mycobacterium leprae : 41% d'identité, et 54% de similarité ;
Aucune homologie significative avec le gène pgm de L. lactis décrit par QIAN et al. (publication précitée) n'a été observée. Le pourcentage d'identité avec le gène pgm de S. pneumoniae décrit par HARDY et al. (publication précitée) est inférieur à 31%.
Aucune homologie significative avec le gène pgm de L. lactis décrit par QIAN et al. (publication précitée) n'a été observée. Le pourcentage d'identité avec le gène pgm de S. pneumoniae décrit par HARDY et al. (publication précitée) est inférieur à 31%.
Les Inventeurs ont effectué la mutagenèse dirigée du gène pgm de S. thermophilus, et ont constaté, que, de manière surprenante, l'inactivation totale ou partielle de ce gène entraînait une augmentation de la production d'EPS.
La présente invention a également pour objet un mutant de bactérie lactique surproducteur d'EPS dans lequel le gène pgm de l'alpha-phosphoglucomutase est totalement ou partiellement inactivé.
<Desc/Clms Page number 6>
Ladite bactérie lactique sera de préférence une bactérie mésophile ou thermophile, choisie parmi les streptocoques et les lactobacilles. A titre d'exemple, il peut s'agir de Lactococcus lactis, Streptococcus thermophilus, Leuconostoc mesenteroide, Lactobacillus casei, Lactobacillus delbrueckii, Lactobacillus sake, etc.
L'inactivation du gène pgm peut être obtenue en effectuant une ou plusieurs mutations au niveau de la séquence codant pour l'a-PGM, et/ou au niveau de séquences contrôlant son expression.
On peut notamment utiliser des techniques de mutagenèse dirigée, connues en elles-mêmes de l'homme de l'art, et qui permettent d'introduire dans un gène une mutation définie, à l'emplacement souhaité.
On peut ainsi, par exemple, inactiver le gène pgm en introduisant à l'intérieur de la séquence codante, ou des séquences de régulation, une séquence exogène, par exemple un transposon.
Avantageusement, on peut également effectuer le remplacement, par recombinaison homologue, de la séquence sauvage du gène pgm par la séquence mutée. Dans ce cas on effectue à l'intérieur d'une séquence identique à celle de la région du gène que l'on souhaite muter, une ou plusieurs modification par insertion, délétion, ou substitution, d'un ou plusieurs nucléotides, consécutifs ou non.
Selon un mode de réalisation préféré de la présente invention, la séquence mutée est insérée dans un vecteur permettant l'intégration par recombinaison (simple crossing-over) entre le fragment d'ADN bactérien cloné dans le vecteur et la région homologue du génome bactérien.
L'excision des séquences du vecteur peut s'effectuer à la suite d'un second événement de recombinaison (double crossing-over), qui aboutit à la substitution de la forme sauvage chromosomique par la forme modifiée.
Des vecteurs permettant l'intégration d'une séquence exogène dans le chromosome d'une bactérie lactique sont connus en eux-mêmes, et disponibles pour la plupart des espèces de bactéries lactiques ; il peut par exemple s'agir de vecteurs non-réplicatifs, de vecteurs réplicatifs
<Desc/Clms Page number 7>
instables ou à réplication conditionnelle, de vecteurs portant des séquences d'insertion, etc..
On peut également, dans certains cas, effectuer la transformation directe de la bactérie avec l'ADN portant la séquence mutée que l'on souhaite insérer.
Des mutants du gène pgm conformes à l'invention peuvent aussi être obtenus par mutagenèse aléatoire (par exemple par mutagenèse chimique ou par radiations) ; peut aussi s'agir de mutants naturels sélectionnés à partir de cultures de bactéries lactiques par criblage sur la base de leurs propriétés phénotypiques.
En effet, les mutants dans lesquels l'a-PGM est partiellement ou totalement inactive ont une croissance normale sur lactose, et une croissance très ralentie sur glucose ou galactose seul. De tels mutants peuvent donc, qu'il s'agisse de mutants naturels, ou qu'ils soient issus d'une mutagenèse, être sélectionnes directement sur la base de cette propriété.
On peut aussi si on le souhaite, augmenter ou diminuer à volonté la production d'EPS en fonction, par exemple, des conditions de culture, en placant le gène pgm sous contrôle transcriptionnel d'un promoteur inductible.
Les inventeurs ont en outre cloné et caractérisé le gène galU de S. thermophilus. Ce gène est représenté dans la liste de séquences en annexes sous le numéro SEQ ID NO: 3.
La protéine GalU codée par ce gène est représentée sous le numéro SEQ ID NO: 4.
La présente Invention a également pour objet une séquence d'acide nucléique codant un polypeptide dont la séquence en acides aminés a au moins 85% d'identité ou au moins 90% de similarité, avantageusement au moins 90% d'identité ou au moins 95% de similarité, et de manière tout à fait préférée au moins 95% d'identité ou au moins 99% de similarité, avec le polypeptide SEQ ID NO: 4.
Les polypeptides présentant ainsi les pourcentages d'identité ou de similarité les plus importants avec la séquence SEQ ID NO: 4, identifiés par recherche sur la base de données "GENBANK nr" en utilisant le logiciel
<Desc/Clms Page number 8>
BLASTp [ALTSCHUL et al., Nucleic Acids Res., 25,3389-3402, (1997) ] sont les suivants : la glucose-1-phosphate uridyltransférase de Streptococcus mutans : 87% d'identité, et 93% de similarité ; - la protéine GalU de Streptococcus pneumoniae : 87% d'identité, et 93% de similarité ; l'UDP-glucose pyrophosphorylase de Streptococcus pyogenes . 84% d'identité, et 90% de similarité ; - la protéine Cap3C de Streptococcus pneumoniae : 76% d'identité, et 88% de similarité ; l'UDP-glucose pyrophosphorylase de Bacillus subtilis : 55% d'identité, et 74% de similarité.
La surexpression du gène galU permet d'accroître la synthèse des EPS, en augmentant la quantité de leurs précurseurs.
La présente invention a donc également pour objet un mutant de bactérie lactique surproducteur d'EPS dans lequel le gène galU est surexprimé.
Un tel mutant peut notamment être obtenu en introduisant dans une bactérie lactique une ou plusieurs copies de ce gène, et/ou en remplaçant son promoteur par un promoteur fort. De préférence, ledit mutant possède également une aPGM partiellement ou totalement inactive.
Des souches mutantes conformes à l'invention peuvent notamment être obtenues à partir de S. thermophilus.
Dans le cas de cette bactérie, un autre facteur limitant pour la synthèse d'EPS peut provenir du fait qu'un grand nombre de souches n'utilisent pas efficacement le galactose, que ce soit pour leur croissance ou la synthèse des EPS.
Les inventeurs ont constaté que des souches gal+ de S. thermophilus sélectionnées pour leur capacité de croissance sur galactose pouvaient synthétiser une quantité plus importante d'EPS. A partir de ces mutants, ou d'autres souches capables d'utiliser le galactose pour leur croissance, on peut effectuer une seconde sélection des souches produisant les colonies d'aspect plus volumineux, ce qui traduit une synthèse plus importante d'EPS.
<Desc/Clms Page number 9>
L'utilisation de souches gal+ de S. thermophilus, pour la production de mutants conformes à l'invention, améliore donc encore la capacité de ceux-ci à produire des EPS.
Des souches de bactéries lactiques conformes à l'invention peuvent avantageusement être utilisées pour la fabrication de produits fermentés, notamment de produits alimentaires, dont on souhaite contrôler, et en particulier augmenter, la teneur en EPS, ainsi que pour la production d'EPS.
La présente invention sera mieux comprise à l'aide du complément de description qui va suivre, qui se réfère à des exemples d'obtention et d'utilisation de souches de bactéries lactiques conformes à l'invention.
Exemple 1 : Sélection de souches de S. thermophilus utilisant le galactose plus efficacement pour la synthèse des EPS
A) Des souches capables d'utiliser efficacement le galactose ont été sélectionnées à partir de cultures de levains industriels de S. thermophilus des collections RHODIA et DANONE RD488 (JIM7446) (collection) et ext 1. 5 (JIM7459) (collection).
A) Des souches capables d'utiliser efficacement le galactose ont été sélectionnées à partir de cultures de levains industriels de S. thermophilus des collections RHODIA et DANONE RD488 (JIM7446) (collection) et ext 1. 5 (JIM7459) (collection).
Pour cela, une culture de 12 heures en milieu M17 [TERZAGHI et SANDINE, Appl. Microbiol., 29,807-813, (1975) ] contenant 1% de lactose, est déposée sur boîtes M17 agar contenant 1% de galactose et incubée à 42 C pendant 2 jours.
Les colonies ainsi obtenues sont striées sur boîte M17 agar contenant 1% de galactose et incubées à 42 C pendant la nuit.
Les colonies poussant ainsi sur M17 galactose sont récupérées et mises à pousser en M17 liquide contenant 1% galactose à 42 C pendant 6 heures avant d'être mises en collection.
D'autres milieux comme le milieu chimiquement défini décrit par [SISSLER et al., Proc. Natl. Acad. Sci. USA, 96, no. 16, 8985-8990, (1999)], où la source de sucre peut être contrôlée, peuvent être aussi utilisés pour cette sélection.
Pour vérifier que les souches obtenues sont bien capables de mieux utiliser le galactose, les croissances de ces souches et de leurs parents sont comparées.
Les croissances sont réalisées comme suit : des précultures de la nuit dans 10 ml de milieu Belliker
<Desc/Clms Page number 10>
(Laboratoires DIFCO) auquel est ajouté 10 g d'extrait de boeuf/1 (DIFCO), et contenant 1% de galactose sont diluées 100 fois et mises à pousser dans 2 ml de milieu M17 contenant 2% de sucre (saccharose, lactose, glucose ou galactose) pendant 4 heures à 42 C. Elles sont ensuite inoculées à 2% dans 200 l de milieu M17 contenant 1% de sucre. La croissance des souches est alors suivie au BIOSCREEN (LASBSYSTEMS).
La Figure 1 représente la croissance sur différents sucres, de la souche industrielle JIM7459 (lA) et d'un mutant gal+ sélectionné à partir de cette souche (1B).
B) On peut effectuer une sélection supplémentaire à partir de souches déjà capables d'utiliser le galactose pour leur croissance. Il est alors recherché des clones produisant des colonies d'aspect plus volumineux sur les boîtes. Ces souches utilisent le galactose encore plus efficacement.
La Figure 2 représente la croissance sur différents sucres, d'un mutant gal+ (2A) sélectionné comme décrit en A ci-dessus, à partir d'une culture d'une souche industrielle (JIM7446, collection DANONE), et d'un mutant gal++ (2B) sélectionné à partir d'une culture de ce mutant gal+, sur la base de la taille des colonies.
Après croissance de ces différentes souches en milieu chimiquement défini, ou en milieu M17, le culot bactérien des mutants gal+ est plus filamenteux que celui des souches sauvages, ce qui peut refléter une différence dans la production d'EPS entre les mutants obtenus et leur souche parentale. La production d'EPS de ces souches a été déterminée à partir de culture dans 80 ml de milieu chimiquement défini [SISSLER et al., Proc. Natl. Acad. Sci.
USA, 96, no. 16,8985-8990, (1999) ].
Après croissance, les cultures sont centrifugées à 16000 g pendant 10 min à 4 C. Le culot cellulaire est éliminé, et le surnageant est précipité (2 volumes d'éthanol 100% pour un volume de surnageant) pendant 24 heures à 4 C.
Après précipitation, une centrifugation est effectuée à 16000 g pendant 15 minutes à 4 C. Le surnageant est éliminé et le culot est resuspendu dans 60 ml d'eau. Une dialyse est
<Desc/Clms Page number 11>
effectuée contre l'eau pendant 4 jours en changeant l'eau 3 à 4 fois par jour. Les EPS sont dosés par la méthode au phénol/acide sulfurique [DUBOIS et al., Analytical Chemistry, 28,350-356, (1956) ].
Les résultats obtenus dans le cas de la souche 7446 Gal+, et d'un mutant de cette souche capable d'utiliser plus efficacement le galactose sont indiqués dans le Tableau I ci-après.

<tb>
<tb>
<tb>
Tableau <SEP> @
<tb> SOUCHES <SEP> EPS <SEP> (mg/l)
<tb> 7446 <SEP> Gal+ <SEP> 4
<tb> 7446 <SEP> Gal++ <SEP> 8
<tb>
<tb> SOUCHES <SEP> EPS <SEP> (mg/l)
<tb> 7446 <SEP> Gal+ <SEP> 4
<tb> 7446 <SEP> Gal++ <SEP> 8
<tb>
Après croissance en présence de galactose, les souches utilisant mieux le galactose produisent des quantités accrues d'EPS.
Exemple 2: Clonage du gène pgm de Streptococcus thermophilus et construction de mutants dans lesquels ce gène est inactivé.
Clonage du gène :
Le gène pgm de Streptococcus thermophilus peut êcre obtenu par PCR inverse [OCHMAN et al., Biorechnology (NY), 8, no. 8,759-760, (1990) ] à partir de l'ADN chromosomique de S. thermophilus. L'ADN chromosomique de S. thermophilus est digéré avec des enzymes de restriction (BamHI, EcoRI, HindIII, NcoI, PsI, XhI) puis les produits de coupure sont circularisés puis amplifiés par PCR en utilisant les amorces complémentaires au brin opposé OST15, OST16, OST23 à OST26 (Tableau II).
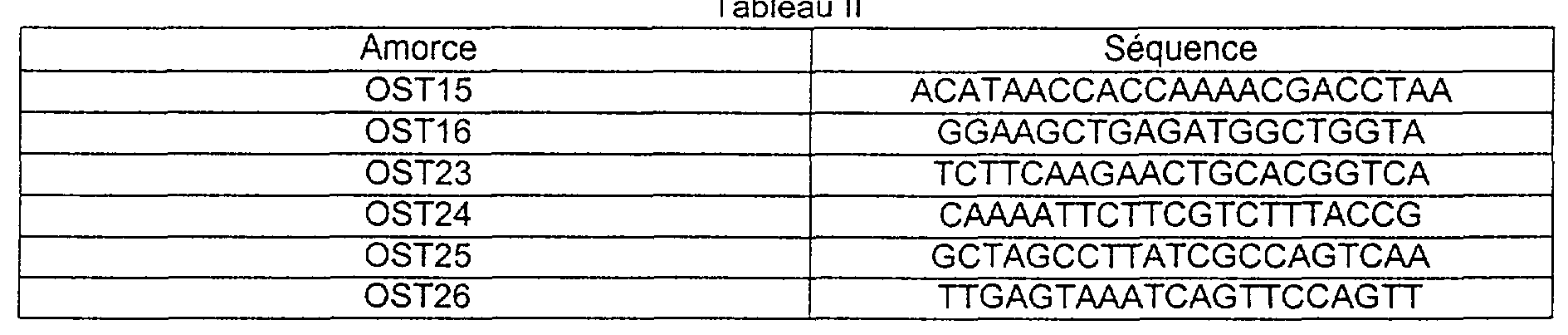
Le gène pgm de Streptococcus thermophilus peut êcre obtenu par PCR inverse [OCHMAN et al., Biorechnology (NY), 8, no. 8,759-760, (1990) ] à partir de l'ADN chromosomique de S. thermophilus. L'ADN chromosomique de S. thermophilus est digéré avec des enzymes de restriction (BamHI, EcoRI, HindIII, NcoI, PsI, XhI) puis les produits de coupure sont circularisés puis amplifiés par PCR en utilisant les amorces complémentaires au brin opposé OST15, OST16, OST23 à OST26 (Tableau II).
<tb>
<tb> i <SEP> aoieau <SEP> @@
<tb> Amorce <SEP> Séquence
<tb> OST15 <SEP> ACATAACCACCAAAACGACCTAA
<tb> OST16 <SEP> GGAAGCTGAGATGGCTGGTA
<tb> OST23 <SEP> TCTTCAAGAACTGCACGGTCA
<tb> OST24 <SEP> CAAAATTCTTCGTCTTTACCG
<tb> OST25 <SEP> GCTAGCCTTATCGCCAGTCAA
<tb> OST26 <SEP> TTGAGTAAATCAGTTCCAGTT
<tb>
<tb> i <SEP> aoieau <SEP> @@
<tb> Amorce <SEP> Séquence
<tb> OST15 <SEP> ACATAACCACCAAAACGACCTAA
<tb> OST16 <SEP> GGAAGCTGAGATGGCTGGTA
<tb> OST23 <SEP> TCTTCAAGAACTGCACGGTCA
<tb> OST24 <SEP> CAAAATTCTTCGTCTTTACCG
<tb> OST25 <SEP> GCTAGCCTTATCGCCAGTCAA
<tb> OST26 <SEP> TTGAGTAAATCAGTTCCAGTT
<tb>
Les bandes obtenues sont extraites du gel et séquencées. La séquence du fragment cloné, comprenant le gène pgm et ses régions flanquantes est représentée dans la liste de séquences en annexe sous le numéro SEQ ID NO: 1.
La taille du cadre ouvert de lecture du gène pgm de Streptococcus thermophilus est de 1350 pb.
<Desc/Clms Page number 12>
Construction de mutants du gène pgm
Pour diminuer l'activité du gène pgm, les Inventeurs ont adopté une stratégie d'inactivation par insertion d'un vecteur dans le gène par recombinaison homologue. La stratégie générale d'inactivation est représentée par la Figure 3. Dans un premier temps, des plasmides à réplication thermosensible, contenant des fragments internes au gène pgm ont été construits à partir du vecteur à réplication thermosensible pG+host [BISWAS et al., J. Bacteriol., 175,11, 3628-3635, (1993) ; Demande PCT WO/181164]. Deux fragments PCR respectivement de 650 à 835 bp dans le gène pgm, produits avec les oligonucléotides OST29+OST31 et OST28+OST31, ont été clonés dans pG+host générant les plasmides pST28 et pST29.

Pour diminuer l'activité du gène pgm, les Inventeurs ont adopté une stratégie d'inactivation par insertion d'un vecteur dans le gène par recombinaison homologue. La stratégie générale d'inactivation est représentée par la Figure 3. Dans un premier temps, des plasmides à réplication thermosensible, contenant des fragments internes au gène pgm ont été construits à partir du vecteur à réplication thermosensible pG+host [BISWAS et al., J. Bacteriol., 175,11, 3628-3635, (1993) ; Demande PCT WO/181164]. Deux fragments PCR respectivement de 650 à 835 bp dans le gène pgm, produits avec les oligonucléotides OST29+OST31 et OST28+OST31, ont été clonés dans pG+host générant les plasmides pST28 et pST29.
<tb>
<tb>
<tb>
Tableau <SEP> III
<tb> Amorce <SEP> Séquence
<tb> OST28 <SEP> TAAGGGCCCTAAATATTTTGGAACAGA
<tb> OST29 <SEP> CCGGGGCCCACTTCTCTCAGTAGGTAT
<tb> OST31 <SEP> AATATCGATATTTTCATCAACGGCAAT
<tb>
<tb> Amorce <SEP> Séquence
<tb> OST28 <SEP> TAAGGGCCCTAAATATTTTGGAACAGA
<tb> OST29 <SEP> CCGGGGCCCACTTCTCTCAGTAGGTAT
<tb> OST31 <SEP> AATATCGATATTTTCATCAACGGCAAT
<tb>
Le fragment de 650 bp porté par pST28 est central au gène, il est donc attendu que son insertion produise une inactivation totale du gène. Le fragment de 835 bp porté par pST29 contient la partie située en 5' du gène avec le codon de démarrage de traduction, mais en absence de son site de fixation au ribosome. Il est attendu que l'insertion de pST29 dans le chromosome provoque une diminution très importante de la traduction de pgm en ne permettant qu'une expression de base très faible.
Les plasmides pG+host, pST28 et pST29 ont été introduits par transformation dans les souches de S. thermophilus JIM7445 et JIM7459 à 30 C, puis intégrés par simple crossing-over comme schématisé sur la Figure 3.
L'intégration par simple crossing-over a été réalisée selon le protocole décrit par BISWAS et al. [publication précitée, (1993)] avec les modifications suivantes :
Les souches de Streptococcus thermophilus contenant le plasmide pG+host ou ses dérivés sont mises à pousser de nuit à 30 C en présence d'érythromycine, puis diluées 50 fois dans le même milieu. Elles sont ensuite transférées à 42 C pendant 6 heures. Les échantillons sont alors dilués et étalés d'une part à 42 C sur boîtes M17
Les souches de Streptococcus thermophilus contenant le plasmide pG+host ou ses dérivés sont mises à pousser de nuit à 30 C en présence d'érythromycine, puis diluées 50 fois dans le même milieu. Elles sont ensuite transférées à 42 C pendant 6 heures. Les échantillons sont alors dilués et étalés d'une part à 42 C sur boîtes M17
<Desc/Clms Page number 13>
contenant l'érythromycine pour détecter les événements d'intégration, et d'autre part à 30 C sur boîtes M17 sans antibiotique pour détecter le nombre total de cellules viables. La fréquence d'intégration par cellule est calculée en faisant le rapport de ces comptages. Les fréquences d'intégration de ces plasmides sont respectivement de 3.5 x 10-3, 3 x 10-3 et 10-2 pour pG+host, PST28 et pST29. Des clones issus de chacune de ces intégrations ont été isolés pour donner les souches STJ3, STJ1 et STJ2 respectivement.
Les souches STJ1 (pST28), STJ2 (pST29) et STJ3 (pG+host) ont été mises à pousser sur boîtes de milieu chimiquement défini contenant du glucose et du galactose ou sur boîtes des mêmes milieux contenant soit du glucose seul, soit du galactose seul. Les résultats obtenus sont indiqués dans le Tableau IV.

<tb>
<tb>
<tb>
Tableau <SEP> iv
<tb> Souches <SEP> plasmides <SEP> Croissance <SEP> sur <SEP> MCD <SEP> Croissance <SEP> sur <SEP> MCD <SEP> Croissance <SEP> sur <SEP> MCD
<tb> Glu <SEP> Gal <SEP> Ery <SEP> Glu <SEP> Ery <SEP> Gal <SEP> Ery
<tb> STJ1 <SEP> pST28 <SEP> Normale <SEP> Ne <SEP> pousse <SEP> pas <SEP> Ne <SEP> pousse <SEP> pas
<tb> STJ2 <SEP> pST29 <SEP> Normale <SEP> Ne <SEP> pousse <SEP> pas <SEP> Ne <SEP> pousse <SEP> pas
<tb> STJ3 <SEP> pG+host <SEP> Normale <SEP> Normale <SEP> Normale
<tb>
<tb> Souches <SEP> plasmides <SEP> Croissance <SEP> sur <SEP> MCD <SEP> Croissance <SEP> sur <SEP> MCD <SEP> Croissance <SEP> sur <SEP> MCD
<tb> Glu <SEP> Gal <SEP> Ery <SEP> Glu <SEP> Ery <SEP> Gal <SEP> Ery
<tb> STJ1 <SEP> pST28 <SEP> Normale <SEP> Ne <SEP> pousse <SEP> pas <SEP> Ne <SEP> pousse <SEP> pas
<tb> STJ2 <SEP> pST29 <SEP> Normale <SEP> Ne <SEP> pousse <SEP> pas <SEP> Ne <SEP> pousse <SEP> pas
<tb> STJ3 <SEP> pG+host <SEP> Normale <SEP> Normale <SEP> Normale
<tb>
Les clones STJ1 et STJ2 ne poussent donc pas sur glucose ou galactose seul, mais normalement en lactose ou sur un mélange glucose et galactose. Ceci montre que le métabolisme du glucose et du galactose a bien été découplé dans cette souche et que le gène dont l'activité a été affectée est bien pgm.
Ces résultats montrent que le gène inactivé code bien pour une enzyme connectant la voie des EPS et la glycolyse. Il code donc probablement pour l'a-PGM dont la séquence n'était pas encore caractérisée expérimentalement chez les bactéries lactiques.
Dosage des EPS produits par la souche de Streptococcus thermophilus portant une mutation dans le gène pgm
Le dosage des EPS est effectué comme décrit à l'exemple 1 ci-dessus.
Le dosage des EPS est effectué comme décrit à l'exemple 1 ci-dessus.
La production d'EPS de la souche STJ2 affectée dans l'expression du gène pgm est de 20 mg/1. Cette production, comparée aux 8 mg/1 obtenu pour la souche parentale 7446 Gal++ correspond donc à une augmentation de deux fois et demi.
<Desc/Clms Page number 14>
Exemple 2 : Construction de mutants surproduisant GalU Clonage du gène galU codant pour la Glucose-1-phosphate Uridyl Transférase
L' ensemble du gène galU peut être obtenu par PCR inverse [OCHMAN et al., (1990), publication précitée] à partir de l'ADN chromosomique de S. thermophilus, en utilisant des amorces complémentaires au brin opposé (OST13, OST14, OST21 et OST22) (Tableau V).

L' ensemble du gène galU peut être obtenu par PCR inverse [OCHMAN et al., (1990), publication précitée] à partir de l'ADN chromosomique de S. thermophilus, en utilisant des amorces complémentaires au brin opposé (OST13, OST14, OST21 et OST22) (Tableau V).
<tb>
<tb>
<tb>
Tableau <SEP> v
<tb> Amorce <SEP> Séquence
<tb> OST13 <SEP> CCAAGATATCTTCAATACCAGAC
<tb> OST14 <SEP> GAAAACACAACGCGTCTTTGC
<tb> OST21 <SEP> GCACCAGCACCGACTGCGATA
<tb> OST22ACTGGGTAGTTAAACGGTAAT
<tb>
<tb> Amorce <SEP> Séquence
<tb> OST13 <SEP> CCAAGATATCTTCAATACCAGAC
<tb> OST14 <SEP> GAAAACACAACGCGTCTTTGC
<tb> OST21 <SEP> GCACCAGCACCGACTGCGATA
<tb> OST22ACTGGGTAGTTAAACGGTAAT
<tb>
Les bandes obtenues sont extraites du gel et séquencées.
La séquence du fragment cloné, comprenant le gène galU est représentée dans la liste de séquences en annexe sous le numéro SEQ ID NO: 3.
La taille du gène galU est de 914 pb.
Surexpression de GalU
Le gène galU (avec son terminateur et son site de liaison des ribosomes) a été amplifié avec les oligonucléotides OST44 et OST45 (Tableau VI) et cloné en aval du promoteur p45 [SIBAKOV et al., Appl. Environ. Microbiol., 57,341-348, (1991) ] de L. lactis dans le site Apal.
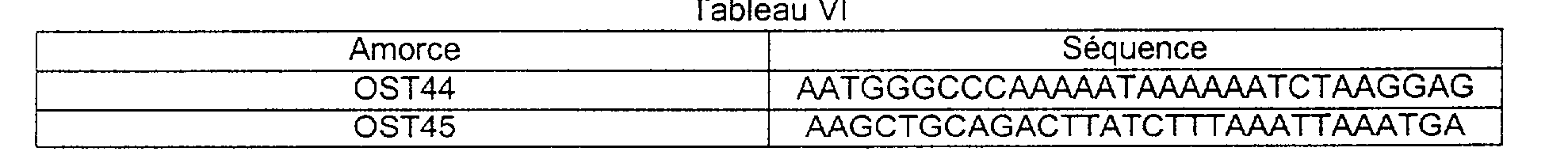
Le gène galU (avec son terminateur et son site de liaison des ribosomes) a été amplifié avec les oligonucléotides OST44 et OST45 (Tableau VI) et cloné en aval du promoteur p45 [SIBAKOV et al., Appl. Environ. Microbiol., 57,341-348, (1991) ] de L. lactis dans le site Apal.
<tb>
<tb> lableau <SEP> VI
<tb> Amorce <SEP> Séquence
<tb> OST44 <SEP> AATGGGCCCAAAAATAAAAAATCTAAGGAG
<tb> OST45 <SEP> AAGCTGCAGACTTATCTTTAAATTAAATGA
<tb>
<tb> lableau <SEP> VI
<tb> Amorce <SEP> Séquence
<tb> OST44 <SEP> AATGGGCCCAAAAATAAAAAATCTAAGGAG
<tb> OST45 <SEP> AAGCTGCAGACTTATCTTTAAATTAAATGA
<tb>
Un fragment Smal du plasmide résultant, portant P45 suivi de galU a été purifié et inséré dans le site SmaI du vecteur pGKV259 se répliquant chez S. thermophilus. Le plasmide pGKV259 [VAN DER VOSSEN et al., Appl. Environ. Microbiol., 53,2452-2457, (1987) ] exprimant le gène galU sous contrôle du promoteur p45 est appelé pSTJ4 (Figure 4).
Ce plasmide est introduit par transformation dans la souche S. thermophilus JIM7446, JIM7459 ou dans la souche contenant une mutation dans le gène pgm (STJ2).
Claims (5)
1) Mutant de bactérie lactique surproducteur d'exopolysaccharides dans lequel le gène pgm de l'alphaphosphoglucomutase est totalement ou partiellement inactivé, et/ou le gène galU est surexprimé.
2) Mutant selon la revendication 1, caractérisé en ce que ladite bactérie lactique est Streptococcus thermophilus.
3) Mutant selon la revendication 2, caractérisé en ce qu'il est obtenu à partir d'une souche de Streptococcus thermophilus capable d'utiliser le galactose.
4) Utilisation d'un mutant de bactérie lactique selon une quelconque des revendications 1 à 3 pour la fabrication d'un produit fermenté.
5) Utilisation d'un mutant de bactérie lactique selon une quelconque des revendications 1 à 3 pour la production d'un exopolysaccharide.
Priority Applications (12)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0004971A FR2807764B1 (fr) | 2000-04-18 | 2000-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
| HU0301029A HUP0301029A3 (en) | 2000-04-18 | 2001-04-18 | Lactic acid bacteria overproducing exopolysaccharides |
| PCT/FR2001/001198 WO2001079476A1 (fr) | 2000-04-18 | 2001-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
| AU2001254881A AU2001254881A1 (en) | 2000-04-18 | 2001-04-18 | Lactic acid bacteria overproducing exopolysaccharides |
| CA002406566A CA2406566A1 (fr) | 2000-04-18 | 2001-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
| CN01808204A CN1425063A (zh) | 2000-04-18 | 2001-04-18 | 过度产生胞外多糖的乳酸菌突变体 |
| BR0110128-5A BR0110128A (pt) | 2000-04-18 | 2001-04-18 | Mutante de bactéria láctica superprodutora de exopolissacarìdeos, utilização do mesmo e ácido nucleico codificando uma alfa-fosfoglucomutase |
| MXPA02010337A MXPA02010337A (es) | 2000-04-18 | 2001-04-18 | Mutantes de bacterias lacticas que sobreproducen exopolisacaridos. |
| EP01928002A EP1274836A1 (fr) | 2000-04-18 | 2001-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
| JP2001577459A JP2004500832A (ja) | 2000-04-18 | 2001-04-18 | 細胞外多糖類を過剰生産する乳酸菌の突然変異体 |
| US10/257,737 US7241610B2 (en) | 2000-04-18 | 2001-04-18 | Lactic acid bacteria overproducing exopolysaccharides |
| NO20025008A NO20025008L (no) | 2000-04-18 | 2002-10-17 | Melkesyrebakterier som overproduserer exopolysakkarider |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| FR0004971A FR2807764B1 (fr) | 2000-04-18 | 2000-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| FR2807764A1 true FR2807764A1 (fr) | 2001-10-19 |
| FR2807764B1 FR2807764B1 (fr) | 2004-09-10 |
Family
ID=8849365
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| FR0004971A Expired - Fee Related FR2807764B1 (fr) | 2000-04-18 | 2000-04-18 | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides |
Country Status (12)
| Country | Link |
|---|---|
| US (1) | US7241610B2 (fr) |
| EP (1) | EP1274836A1 (fr) |
| JP (1) | JP2004500832A (fr) |
| CN (1) | CN1425063A (fr) |
| AU (1) | AU2001254881A1 (fr) |
| BR (1) | BR0110128A (fr) |
| CA (1) | CA2406566A1 (fr) |
| FR (1) | FR2807764B1 (fr) |
| HU (1) | HUP0301029A3 (fr) |
| MX (1) | MXPA02010337A (fr) |
| NO (1) | NO20025008L (fr) |
| WO (1) | WO2001079476A1 (fr) |
Families Citing this family (7)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| RU2007129848A (ru) * | 2005-01-06 | 2009-02-20 | Кр. Хансен А/С (Dk) | Молочнокислые бактерии, устойчивые к бактериофагам |
| JP5944824B2 (ja) | 2009-09-01 | 2016-07-05 | セーホーエル.ハンセン アクティーゼルスカブ | エキソ多糖の過剰発現による、食品をテキスチャリングするための改変されたガラクトキナーゼ発現を伴う乳酸菌 |
| BR112012018882B1 (pt) | 2010-01-28 | 2020-12-01 | Chr. Hansen A/S | processo para fabricação de bactéria de ácido lático que gera tensão de cisalhamento e/ou rigidez de gel superior à cepa mãe quando as bactérias são usadas para fermentação de leite, cepa bacteriana, composição compreendendo a referida bactéria e processo para produção de produto de leite fermentado |
| MX346975B (es) | 2010-10-22 | 2017-04-07 | Chr Hansen As | Cepas texturizadoras de bacterias de acido lactico. |
| WO2017103051A1 (fr) * | 2015-12-18 | 2017-06-22 | Chr. Hansen A/S | Bactérie lactique pour préparer des produits alimentaires fermentés ayant un goût sucré naturel augmenté et une texture élevée |
| CN108251348A (zh) * | 2018-03-29 | 2018-07-06 | 上海理工大学 | 一种嗜热链球菌感受态细胞的制备及转化方法 |
| CN109182186B (zh) * | 2018-09-19 | 2022-02-01 | 内蒙古大学 | 一种乳酸菌胞外多糖及免疫佐剂 |
Family Cites Families (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| JP4469026B2 (ja) * | 1996-10-31 | 2010-05-26 | ヒューマン ジノーム サイエンシーズ,インコーポレイテッド | Streptococcus pneumoniaeの抗原およびワクチン |
| WO2001049721A2 (fr) | 1999-12-30 | 2001-07-12 | Bristol-Myers Squibb Company | Nouveaux genes bacteriens et proteines qui sont essentielles pour la viabilite cellulaire et leurs utilisations |
-
2000
- 2000-04-18 FR FR0004971A patent/FR2807764B1/fr not_active Expired - Fee Related
-
2001
- 2001-04-18 JP JP2001577459A patent/JP2004500832A/ja not_active Withdrawn
- 2001-04-18 CA CA002406566A patent/CA2406566A1/fr not_active Abandoned
- 2001-04-18 WO PCT/FR2001/001198 patent/WO2001079476A1/fr not_active Ceased
- 2001-04-18 MX MXPA02010337A patent/MXPA02010337A/es unknown
- 2001-04-18 EP EP01928002A patent/EP1274836A1/fr not_active Withdrawn
- 2001-04-18 US US10/257,737 patent/US7241610B2/en not_active Expired - Fee Related
- 2001-04-18 CN CN01808204A patent/CN1425063A/zh active Pending
- 2001-04-18 AU AU2001254881A patent/AU2001254881A1/en not_active Abandoned
- 2001-04-18 BR BR0110128-5A patent/BR0110128A/pt not_active Application Discontinuation
- 2001-04-18 HU HU0301029A patent/HUP0301029A3/hu unknown
-
2002
- 2002-10-17 NO NO20025008A patent/NO20025008L/no not_active Application Discontinuation
Non-Patent Citations (5)
| Title |
|---|
| DE VUYST LUC ET AL: "Heteropolysaccharides from lactic acid bacteria.", FEMS MICROBIOLOGY REVIEWS, vol. 23, no. 2, April 1999 (1999-04-01), pages 153 - 177, XP000971896, ISSN: 0168-6445 * |
| ESCALANTE A ET AL: "Enzymes involved in carbohydrate metabolism and their role on exopolysaccharide production in Streptococcus thermophilus.", JOURNAL OF APPLIED MICROBIOLOGY, vol. 84, no. 1, January 1998 (1998-01-01), pages 108 - 114, XP000978853, ISSN: 1364-5072 * |
| HARDY GAIL G ET AL: "Capsule biosynthesis and basic metabolism in Streptococcus pneumoniae are linked through the cellular phosphoglucomutase.", JOURNAL OF BACTERIOLOGY, vol. 182, no. 7, April 2000 (2000-04-01), pages 1854 - 1863, XP002157670, ISSN: 0021-9193 * |
| KLEEREBEZEM MICHIEL ET AL: "Exopolysaccharides produced by Lactococcus lactis: From genetic engineering to improved rheological properties?.", ANTONIE VAN LEEUWENHOEK, vol. 76, no. 1-4, July 1999 (1999-07-01), pages 357 - 365, XP000971897, ISSN: 0003-6072 * |
| MOLLERACH MARTA ET AL: "Characterization of the galU gene of Streptococcus pneumoniae encoding a uridine diphosphoglucose pyrophosphorylase: A gene essential for capsular polysaccharide biosynthesis.", JOURNAL OF EXPERIMENTAL MEDICINE, vol. 188, no. 11, 7 December 1998 (1998-12-07), pages 2047 - 2056, XP002157671, ISSN: 0022-1007 * |
Also Published As
| Publication number | Publication date |
|---|---|
| BR0110128A (pt) | 2003-02-11 |
| US20050164354A1 (en) | 2005-07-28 |
| CA2406566A1 (fr) | 2001-10-25 |
| AU2001254881A1 (en) | 2001-10-30 |
| WO2001079476A1 (fr) | 2001-10-25 |
| MXPA02010337A (es) | 2004-09-06 |
| JP2004500832A (ja) | 2004-01-15 |
| FR2807764B1 (fr) | 2004-09-10 |
| NO20025008L (no) | 2002-12-12 |
| US7241610B2 (en) | 2007-07-10 |
| EP1274836A1 (fr) | 2003-01-15 |
| HUP0301029A3 (en) | 2004-10-28 |
| CN1425063A (zh) | 2003-06-18 |
| NO20025008D0 (no) | 2002-10-17 |
| HUP0301029A2 (hu) | 2003-07-28 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| WO2019194062A1 (fr) | Enzyme dérivée de p aenibacillus pabuli et capable de produire un galacto-oligosaccharide, et procédé de production d'un galacto-oligosaccharide | |
| KR102624718B1 (ko) | 변이형 rpoc 암호화 서열을 포함하는 핵산 분자 | |
| FR2851575A1 (fr) | Nouveau gene de la lysine decarboxylase et procede de production de la l-lysine | |
| KR101091138B1 (ko) | 류코노스톡 락티스로부터 유래된 글루칸수크라제 및 그 제조방법 | |
| FR2822163A1 (fr) | Molecules d'acides nucleiques codant une dextrane-saccharase catalysant la synthese de dextrane portant des ramifications de type alpha-1,2 osidiques | |
| EP0750042A1 (fr) | Bactéries lactiques produisant des exopolysaccharides | |
| FR2807764A1 (fr) | Mutants de bacteries lactiques surproducteurs d'exopolysaccharides | |
| EP0750043B1 (fr) | Bactéries lactiques produisant des exopolysaccharides | |
| KR102149044B1 (ko) | 2-히드록시 감마 부티로락톤 또는 2,4-디히드록시-부티레이트 의 제조 방법 | |
| CN116670295A (zh) | 用于在抑制香草酸形成的情况下产生香草醛的拟无枝酸菌属菌株 | |
| WO1999054475A2 (fr) | Identification de genes de bacteries lactiques dans la biosynthese des exopolysaccharides | |
| CN115975965B (zh) | 乳酸脱氢酶及其编码基因、重组载体、重组菌体和发酵剂以及它们的应用 | |
| KR101153400B1 (ko) | 신규 리포산 합성효소와 리포산 단백질 리가제를 이용한 알파-리포산의 생산방법 | |
| JP2007189905A (ja) | 好アルカリ性サイクロデキストラン合成酵素遺伝子を含有するdna、組み換え体dna、および好アルカリ性サイクロデキストラン合成酵素の製造法 | |
| Bourel et al. | Métabolisme sucre-citrate chez Leuconostoc mesenteroides | |
| WO1999066052A1 (fr) | GENES CODANT POUR DES β-AGARASES ET LEUR UTILISATION POUR LA PRODUCTION D'ENZYMES DE BIODEGRADATION DES AGARS | |
| EP1137784A1 (fr) | Methode d'isolement et de selection de genes codant pour des enzymes, et milieu de culture approprie | |
| EP0957168A1 (fr) | Identification de gènes de Streptococcus thermophilus Sfi39 impliqués dans la biosynthèse d'exopolysaccharides | |
| FR2807766A1 (fr) | Operons de streptococcus thermophilus impliques dans la synthese des eps | |
| WO2026013147A1 (fr) | Région promotrice du gène régulateur gadr2 et son utilisation pour la surproduction d'acide gamma-aminobutyrique | |
| KR101124617B1 (ko) | 신규 리포산 합성효소와 리포산 단백질 리가제를 이용한 알파-리포산의 생산방법 | |
| EP0663955B1 (fr) | Acide nucleique codant pour une alpha-acetolactate decarboxylase et ses applications | |
| EP0957170A1 (fr) | Identification de gènes de Lactobacillus delbrueckii subsp bulgaricus Lfi5 impliqués dans la biosynthèse d'exopolysaccharides | |
| FR2822162A1 (fr) | Molecules d'acides nucleiques codant une dextrane-saccharase calatalysant la synthese de dextrane portant des ramifications de type alpha-1,2 osidiques | |
| EP1214419A2 (fr) | Bacteries lactiques transformees pour leur conferer un metabolisme respiratoire |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| CA | Change of address | ||
| ST | Notification of lapse |
Effective date: 20091231 |